
Abstract
Background
Metabolic dysregulation leading to diabetes is a major public health concern in India. While evidence has pointed to a role for genetic factors, there is still limited knowledge regarding the specific variants that play a part in this process. Recent studies have implicated Tumor protein, p53, a well-known tumor suppressor, in maintaining metabolic homeostasis in our body. Polymorphisms that can disrupt this function are thought to increase susceptibility to diabetic and prediabetic phenotypes like Metabolic syndrome (MetS). A common polymorphism at codon 72 (rs1042522) is associated with obesity and other metabolic disorders. However, its role may vary depending on the specific population and disease context. Our study aimed to evaluate whether the polymorphism at codon 72 of p53 (rs1042522) is associated with MetS and Diabetes, in a Central Indian population.
Results
A total of 66 individuals and 63 healthy controls, identified based on the National Cholesterol Education Program (NCEP)/Adult Treatment Panel-III (ATP-III) 2001 guidelines, were enrolled in the study. The carriers expressing mutant allele “G” for arginine at codon 72 had higher weight than those having wild-type allele “C” which codes for proline (p = 0.038). The majority of the subjects were heterozygous for p53 codon 72 polymorphism though the association was not statistically significant for either MetS or diabetes.
Conclusion
Our findings suggest that p53 codon 72 (rs1042522) varaints may trigger metabolic dysfunction by impacting weight. The polymorphism appears to confer a heterozygous advantage, as individuals with a heterozygous genotype exhibited the highest susceptibility to metabolic disease. Although further studies are required, our results for the first time indicate that the p53 codon 72 (rs1042522) polymorphism could be considered a genetic marker to predict the increased susceptibility to diabetic and prediabetic phenotypes among Central Indian population.
Similar content being viewed by others


Background
Diseases associated with dysregulation in metabolic homeostasis, namely diabetes and metabolic syndrome (MetS), have become a global epidemic, with India being particularly affected. Over the past three decades, the prevalence of diabetes in India has increased fivefold, and recent reports show prevalence rates of over 30% for MetS [1]. In most adults, the presentation of MetS precedes full-blown diabetes. The presence of MetS increases the risk for diabetes three to five times [2]. Multiple criteria for defining MetS have been proposed to exploit the utility of clubbing together various derangements in biochemical parameters seen in diabetes. This information is being used to understand its implications in diabetes development and management. The most recent is the Harmonized Definition, according to which MetS is considered present, when at least 3 of the five medical conditions coexist in an individual. The conditions are increased waist circumference, triglycerides, fasting glucose, or blood pressure or decreased HDL-cholesterol levels [3].
Studies have demonstrated a high prevalence of MetS and diabetes in both urban and rural Indian populations [4], emphasizing the involvement of a genetic component in MetS and diabetes. The distinct Indian phenotype marked by elevated insulin resistance and upper-body adiposity further highlight a genetic predisposition to MetS and diabetes [5, 6]. Recently, the product of p53 genes has been shown to play an essential role in maintaining metabolic homeostasis in our body [7]. p53 is a tumor suppressor gene that influences several biological activities that maintain genomic integrity, particularly in response to insults such as DNA damage, oncogene activation, and oxidative stress [8].
Studies have demonstrated that p53 is involved in the regulation of glucose homeostasis and hepatic glucose production [9]. Additionally, p53 has been found to have a direct role in adipocyte differentiation and function, which is important for metabolic health [10]. Furthermore, p53 is known to contribute to whole body energy homeostasis and is involved in multiple metabolic pathways, including glucose, lipid, and amino acid metabolism [11]. It has also been observed that p53 levels in adipose tissue can impact systemic energy homeostasis and insulin resistance [12]. Overall, these findings highlight the crucial role of p53 in maintaining metabolic homeostasis and its potential implications in metabolic diseases. Consequently, any polymorphism that disrupts its function in maintaining metabolic homeostasis may contribute to an increased susceptibility to MetS and diabetes. In vivo, humanized p53 knock-in mice analyses have shown that p53 codon 72 variants differentially regulate genes involved in cellular metabolism [13]. The variation at codon 72 (rs1042522) resulting in Arginine (Arg) instead of Proline (Pro), is widely studied due to its direct influence on obesity, body mass index (BMI), and higher risks for MetS and diabetes.
The p53 codon 72 variants have been found to differentially regulate genes involved in cellular metabolism [14,15,16]. In a study using isogenic cell lines, it was observed that the expression pattern of genes involved in metabolism was decreased in cells having the alleles for Arg compared to Pro containing cells [17]. Additionally, the Arg variant of mutant p53 was shown to enhance mitochondrial function and metastatic capability in tumor cells, indicating an impact on metabolism [18]. These findings suggest that the p53 codon 72 variants play a role in regulating genes involved in cellular metabolism, with the Arg- containing variant showing a distinct impact compared to the Pro-containing variant.
The p53 codon 72 polymorphisms have been studied in relation to metabolic syndrome and diabetes in various populations. The results from the available abstracts are mixed. One study in South Indian population found no significant association between p53 codon 72 polymorphism and diabetes [19]. However, another study in patients with type 1 diabetes found that the homozygous Pro/Pro genotype of TP53 codon 72 was associated with an increased risk of microvascular complications, dyslipidemia, and celiac disease, while the homozygous Arg/Arg variant was associated with a decreased risk of autoimmune thyroiditis and celiac disease [20]. Another study found that the homozygous Arg/Arg genotype of TP53 codon 72 might be a protective factor for diabetic complications in patients with type 2 diabetes in Chinese Han population [21]. Therefore, the association between p53 codon 72 polymorphism and metabolic diseases like MetS and diabetes may vary depending on the specific population and disease context. Given these facts, in the current study, we aimed to evaluate the prevalence of p53 polymorphism at codon 72 with MetS and diabetes and its various clinicopathological to understand its implications for MetS and diabetes in a Central Indian population.
Methods
In the present case–control study, p53 codon 72 polymorphism was determined by polymerase chain reaction (PCR)-restriction fragment length polymorphism (PCR–RFLP) in subjects diagnosed with or without metabolic syndrome. The subjects were selected among those attending the outpatient department of All India Institute of Medical Sciences-Raipur (AIIMS-Raipur), in Central India between October 2018 and April 2019. In total, one hundred twenty-nine participants were recruited, of which 66 subjects were diagnosed with MetS by the attending clinician, while 63 subjects were not diagnosed with metabolic syndrome. National Cholesterol Education Program (NCEP)/Adult Treatment Panel-III (ATP-III) (NCEP ATP III 2001) criteria were used to identify participants with MetS [22]. Accordingly, participants were categorized as having MetS if any three of the following five criteria were present in them: Waist circumference: > 40 inches in males or > 35 inches in females, Triglycerides: ≥ 150 mg/dL or on treatment, HDL: < 40 mg/dL in males or < 50 mg/dL (F) or on treatment, Fasting plasma glucose: ≥ 100 mg/dL or on treatment, Blood pressure: > 130 mmHg systolic or > 85 mmHg diastolic or on treatment. The controls, age, and gender-matched (with cases) individuals without metabolic syndrome were selected randomly among volunteers. In our study group, 47% of the subjects were prediabetic (fasting blood glucose between 101 and 125 mg/dL), and 47% were diabetic (fasting plasma glucose above 126 mg/dL). In 6% of the MetS subjects, the fasting plasma glucose was below 100 mg/dL. In contrast, in 95% of the controls, blood glucose levels were below 100 mg/dL, while prediabetic and diabetics were 2% and 3%, respectively (Additional file 1: Fig. 1). The study protocol was approved by the Institute Ethics Committee of AIIMS-Raipur and informed consent was obtained from all subjects.
Anthropometric and biochemical parameters
Measurements like height (cm) and weight (kg) were taken on subjects without shoes. Weight was measured using a digital weighing machine. For height measurement, subjects were asked to keep their feet flat on the floor and stand straight with their back flat against the wall, keeping the chin parallel to the floor. We placed a pencil flat against the top of the person's head and marked it on the wall. The distance was measured from the floor to the mark on the wall using a measuring tape. BMI was calculated using the formulae: weight in kilograms divided by the square of height in meters (kg/m2). We measured the waist circumference (inch) midway between the midaxillary line’s lowest rib and iliac crest. Fasting Plasma Glucose (FPG), Total Cholesterol, and Triglycerides were measured on Beckman CoulterAU680. HDL-cholesterol was measured after precipitation of apolipoprotein B containing lipoproteins, and LDL cholesterol level was calculated using the Fridewald formula.
DNA isolation and genotyping
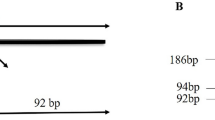
Whole blood was collected in an EDTA vacutainer and stored at − 20 °C until subsequent analysis. DNA was extracted from 200 μL of whole blood using a QIAamp® DNA Blood mini kit (Qiagen, CA, USA) and stored at − 80 °C. DNA yield and quality were assessed on NanoDrop spectrophotometer. Determination of p53 genotype for Arg/Pro polymorphism at codon72 was performed according to a protocol reported by Tandon et al. [23]. Primers, 5′-ATCTACAGTCCCCCTTGCCG-3′ and 5′-GCAACTGACCGTGCAAGTCA-3′, were used to amplify exon 4 region coding for 72nd amino acid of the p53 gene. PCR reaction was performed in 20 μL volume containing 50 ng of genomic DNA template, 12.5 pmol of each primer, 0.1 mM of each dNTP and 1 × PCR buffer. PCR conditions were as follows: an initial denaturation step at 94 °C for 4 min, 35 cycles of 94 °C for 40 s, 56 °C for 30 s, and 72 °C for 30 s and a final extension at 72 °C for 10 min. The PCR product (296 bp fragment) was digested with BstUI (Thermo Scientific™) for 16 h at 37 °C and analyzed on 2.5% agarose gel containing ethidium bromide. The p53 codon 72 polymorphism with Pro allele, which lacks the BstUI restriction site, shows only a single 296 bp band, whereas p53 codon 72 polymorphism with Arg allele, has the BstUI restriction site, and therefore produces bands of 169 and 127 bp. Accordingly, Pro/Pro subjects exhibit a single, 296 bp band, Arg/Arg subjects exhibit two bands of 169 and 127 bp, and heterozygote subjects (Pro/Arg) exhibit all three bands. The gel electrophoresis pattern is shown in Fig. 1.

PCR-restriction fragment length polymorphism analysis of p53 codon 72 polymorphism by 2% agarose gel electrophoresis. Shown are Lanes 1 and 9—50-bp DNA molecular weight marker; Lane 2—No template control; Lane 3, 5, 7–296 bp PCR product; Lanes 4, 6 and 8: PCR products following digestion with BstU1. Lane 4—Pro/Pro (C/C) having 296 bp band; Lane 6—Arg/Arg (G/G) having two bands of 169 and 127 bp, and Lane 8—Pro/Arg (C/G) with all three bands
Statistical analysis
Data was entered in MS Excel and statistical analysis was performed using SPSS 21.0 (SPSS Inc., Chicago, USA). Data are presented as mean ± standard deviation. The genetic model of inheritance was calculated as previously described [24]. Chi square test and t-test were performed for comparison of categorical and numerical data respectively. p-values of < 0.05 were considered significant.
Results
Clinical data and biochemical parameters
Because this was a pilot study, we calculated the sample size based on a previously published formula, n = Z2P(1 − P)/d2, where n is the sample size [25] Z, the standard normal variate was taken as 1.96 at 95% confidence interval (CI). P, the expected prevalence was taken as 21%. The p value used here represents the mean of diabetes prevalence in India, which is 11.6% [26], and MetS prevalence, set at 30%. The allowable error, d, was taken as 10%. Substituting the values in the equation, we got a sample size of 66 cases and 66 controls. Accordingly, we enrolled 132 randomly consenting individuals coming to the hospital for various medical care. However, three control samples did not amplify any genes. So, for calculation purpose our study group consists for 66 cases and 63 controls. The genotype distribution and allele frequency of the individual SNPs, p53 rs1042522 were consistent with Hardy Weinberg's law (Table 1). The MetS patients included 37 females and 29 males, whereas healthy controls included 29 females and 34 males. The samples were normally distributed. The observed genotype distributions agreed with those expected according to the Hardy–Weinberg equilibrium in this population (Table 1). The number of individuals carrying the heterozygous form of p53 rs1042522 was higher than those homozygous for either variants (Table 1). The average age of MetS patients and controls were 45.7 and 47.5, respectively (Fig. 2). There was a significant difference in anthropometric and biochemical measurements between the study groups (Additional file 1: Table 1). Weight, BMI, waist circumference, fasting plasma glucose, Triglyceride, and total cholesterol were significantly higher in the MetS group compared to the control. But HDL-Cholesterol was lower in the group with MetS compared to the healthy control group. Since selected MetS cases and controls were based on NCEP ATP III 2001 criteria, not all cases were diabetic.

Population distribution in cases and control
Relationship of p53 codon 72 (rs1042522) polymorphism on MetS
The genotypic and allele distribution of the studied subjects in MetS is shown in Tables 2 and 3 respectively. There was no significant impact of genotype or allele of p53 polymorphism at codon 72 (rs1042522) in cases or controls (p = 0.71 and 0.91 respectively). An odds ratio of 1 was observed between the cases (MetS) and controls which typically implies no association between the polymorphic allele and the outcome (metabolic syndrome). In order to assess the impact of allele variation, we calculated the odds for the disease (MetS) as previously described (24). The calculated value of OR2 was found to be equivalent to = 1/OR1 (Table 4) which implied that minor allele resulting in “arginine” at codon 72 had an over-dominant effect.
Next, we compared the effect of polymorphism on various clinical and biochemical parameters used to classify MetS (Table 5). The subgroups of MetS patients with different p53 codon 72 (rs1042522) genotypes showed a significant impact on weight (p = 0.03). To check in more detail, we did a Tukey multiple comparison of means at a 95% family-wise confidence level. We found a significant difference between carriers of the allele coding for “Pro” or “Arg”, which impacted the weight (p = 0.02). However, there was no significant difference between carriers of alleles coding for Arg + Pro and Arg + Arg (p = 0.4) or between Arg + Pro and Pro + Pro (p = 0.2), that impacted the weight (Table 6).
Relationship of p53 codon 72 (rs1042522) polymorphism on diabetes
Since selected MetS cases and controls were based on NCEP ATP III 2001 criteria, all the study participants could be further categorized as diabetic or prediabetic using the ADA criteria for diabetes diagnosis based on fasting blood glucose levels. Fasting is defined as no caloric intake for at least 8 h. The subjects were categorized as normal, prediabetic or diabetic if the fasting blood glucose levels were less than 100 mg/dL, 100 mg/dL to 125 mg/dL) and 126 mg/dL or higher respectively. The genotypic and allelic distribution of the p53 polymorphic forms among the three groups were analyzed. However, we did not see any significant impact of the polymorphism among the three groups (Tables 7 and 8). To evaluate the impact of individual alleles on diabetes, we clubbed the variables for prediabetes and diabetes and OR1 and OR2 was calculated as previously described (24). The calculated value to OR2 was 1, which implied that the presence of minor allele resulting in “Arginine” had a dominant role in the risk for diabetes (Table 7).
Discussion
The incidence of a cluster of deranged metabolic components termed MetS is a risk factor for diabetes and parallels the diabetic epidemic in India [1]. Recent studies have implicated the presence of a SNP in the tumor suppressor gene, p53 at codon 72 (rs1042522), resulting in arginine instead of proline in the development of metabolic diseases [14, 15]. However, the association between p53 codon 72 polymorphism and metabolic diseases like MetS and diabetes may vary depending on ethnic variations. Therefore, in the present study, we wanted to investigate the prevalence and impact of p53 codon 72 (rs1042522) polymorphism in the development of MetS and diabetes in a Central Indian population. We found a significant association between p53 codon 72 (rs1042522) polymorphism and weight. The presence of the Pro allele has a protective effect on weight. However, we did not find any significant association with other cardiometabolic components used to define MetS and diabetes, like fasting plasma glucose, total cholesterol, triglycerides, HDL-C, or LDL-C, even though their levels indicated a trend toward risk for type 2 diabetes and cardiovascular diseases.
Type 2 diabetes has become a significant health problem in India. Several factors are being analyzed to curb the prevalence of type 2 diabetes, but none have been very effective. Despite this, it is generally agreed that diabetes is a disease arising from a cluster of deranged metabolic pathways related to disturbances in energy metabolism and can be predicted by analyzing these biological parameters [27]. This cluster, called MetS indicates a predisposition to type 2 diabetes and cardiovascular diseases [28]. Coinciding with the increase in diabetic cases, there is also an increase in the incidence of MetS in India. So, understanding the etiology of MetS is equally crucial for managing and caring for diabetes. According to current estimates, nearly 30% of the population in India has MetS [1]. While the pathogenesis of the MetS and its components is complex and not fully elucidated, genetic profile, physical inactivity, aging, a proinflammatory state, and hormonal dysregulation have been implicated in its development [29]. Increasing evidence has linked the product of tumor suppressor gene, p53 and, in particular, SNP rs1042522 polymorphism having the minor allele “G” with the occurrence and development of metabolic diseases, including MetS and diabetes, in different ethnic populations [30,31,32].
MetS is the concurrence of obesity-associated cardiovascular risk factors. Though an association was found with weight in the studied population, we found no impact on other cardiovascular risk factors. Our observation agrees with the study by Lagares et al., who did not find any association between the p53 polymorphism at codon 72 (rs1042522) and risk factors of cardiovascular diseases or its risk factors in a Brazilian population [33]. Similarly, Alkhalaf et al. found similar risks of cardiovascular diseases and metabolic diseases in their study population from Kuwait, irrespective of p53 codon 72 (rs1042522) polymorphism status [34].
In the present study, we recruited 66 patients diagnosed with MetS and 63 controls. The sample size was consistent with the theoretical sample size calculated based on the dominant inheritance model of metabolic syndrome in an Indian population [1]. We found that the differential distribution of TP53 Pro72Arg genotypes (Pro/Pro, Pro/Arg, and Arg/Arg) was not statistically significant between the MetS patients and the controls (p = 0.71) (Table 2). Similarly, the allele frequency was statistically similar between the MetS patients and the controls (Table 3). However, the polymorphism seems to have a heterozygote advantage as the heterozygotes are more in the population than Arg/Arg or Pro/Pro homozygotes. This would mean that the mutant type p53 can antagonize wild-type p53 functions in a dominant negative manner. This inactivation could stem from the fact that the transcriptional activity of wild-type p53 relies on the formation of tetramers, whose DNA binding function may be interfered by mutant p53 [35]. Our observations that wild-type allele might have a protective effect on weight further confirms this hypothesis.
Since selected MetS cases and controls were based on NCEP ATP III 2001 criteria, we were able to classify our population into diabetic, prediabetic, and normal based on the levels of fasting blood glucose. We did not find any increased risk for diabetes due to polymorphisms in p53 at codon 72 (rs1042522). Our results concordance with the findings of Punja et al. in a population from South India. In their population, p53 codon72 polymorphism was not associated with an increased risk of type 2 diabetes [19]. However, this contrasts with the observation of Guo et al. in a Chinese population. The authors found a significant association of p53 codon 72 (rs1042522) polymorphism with susceptibility to diabetes. Further, they concluded that the homozygous Arg/Arg genotype might be a protective factor for diabetic complications [21]. Similarly, Gloria-Bottini et al. also found that carriers homozygous for “Arg/Arg” has an increased risk for diabetes in an Italian population [36].
Due to the complex, multifactorial disease that diabetes is, there should be considerable interaction between genetic and environmental factors, which may influence several intermediate traits like obesity and MetS. Through its influence on weight, our findings demonstrate the concept of an indirect effect of p53 codon72 as a genetic risk factor for MetS and diabetes. It is known that an increase in weight cause the release of proinflammatory cytokines, which may contribute to obesity-linked metabolic dysfunctions, leading to insulin resistance and type 2 diabetes mellitus [37, 38].
In this preliminary study, we found an indirect association between presence of alleles coding for “Arg” as 72nd amino acid of p53 and risk for MetS and diabetes. MetS and diabetes is a complex disease and their pathology involves interaction of multiple gene variants and environmental factors. In our investigation, we observed that minor allele coding for Arg had a dominant effect on the risk for diabetes. On the contrary, heterozygous alleles for p53 codon 72 polymorphism exhibited a pronounced over-dominant effect on MetS. An over-dominant model assumes the heterozygote has the strongest impact on the disease. MetS is considered as a precursor for diabetes and based on our findings, it can be inferred that the mere presence of a single copy of allele encoding for “Arg” may elevate the susceptibility to metabolic diseases. However, it is not clear if this effect was related to complexity of polygenic diseases like diabetes and MetS or due to limited sample size used in the study. Studies with larger groups as proposed should resolve these uncertainties.
Treating prediabetic conditions like obesity and MetS is more urgent than ever due to its drastic increase in recent years. In addition to contributing to chronic conditions like diabetes, and cardiovascular diseases, it can also impede the quality of life in other ways by limiting a person’s movement. Studies such as ours can be used to identify a subpopulation of people who are more prone to chronic metabolic conditions like MetS and diabetes. These subgroups can be subjected to rigorous lifestyle changes and/or combined with therapeutic interventions to control genetic factors influencing the risk of developing chronic metabolic diseases.
Availability of data and materials
All data generated or analyzed during this study are included in this published article.
Abbreviations
- MetS:
-
Metabolic syndrome
- Arg:
-
Arginine
- Pro:
-
Proline
References
Bhalwar R (2020) Metabolic syndrome: the Indian public health perspective. Med J Armed Forces India 76(1):8–16. https://doi.org/10.1016/j.mjafi.2019.12.001
Nsiah K, Shang VO, Boateng KA, Mensah FO (2015) Prevalence of metabolic syndrome in type 2 diabetes mellitus patients. Int J Appl Basic Med Res 5(2):133–138. https://doi.org/10.4103/2229-516X.157170
Lam DW, LeRoith D (2000) Metabolic syndrome. In: Feingold KR, Anawalt B, Boyce A et al (eds) Endotext. MDText.com, Inc., South Dartmouth
Mehta SR, Kashyap AS, Das S (2009) Diabetes mellitus in India: The modern scourge. Med J Armed Forces India 65(1):50–54. https://doi.org/10.1016/S0377-1237(09)80056-7
Ortega PEN, Meneses ME, Delgado-Enciso I et al (2021) Association of rs9939609-FTO with metabolic syndrome components among women from Mayan communities of Chiapas, Mexico. J Physiol Anthropol 40:11. https://doi.org/10.1186/s40101-021-00259-9
Khan S, Verma AK, Khan V, Bhatt D, Rafat S, Alsahli MA, Rahmani AH, Meena B, Naqvi IH, Dev K (2020) Role of FTO and MC4R polymorphisms in escalating obesity and their indirect association with risk of T2D in Indian population. Diabetes Ther 11(9):2145–2157. https://doi.org/10.1007/s13300-020-00896-w
Lahalle A, Lacroix M, De Blasio C, Cissé MY, Linares LK, Le Cam L (2021) The p53 pathway and metabolism: the tree that hides the forest. Cancers 13(1):133. https://doi.org/10.3390/cancers13010133
Gunaratna RT, Santos A, Luo L et al (2019) Dynamic role of the codon 72 p53 single-nucleotide polymorphism in mammary tumorigenesis in a humanized mouse model. Oncogene 38:3535–3550. https://doi.org/10.1038/s41388-018-0630-4
Gonzalez-Rellan MJ et al (2021) O-GlcNAcylated p53 in the liver modulates hepatic glucose production. Nat Commun 12(1):5068. https://doi.org/10.1038/s41467-021-25390-0
Lee YK, Chung Y, Lee JH, Chun JM, Park JH (2020) The intricate role of p53 in adipocyte differentiation and function. Cells 9(12):2621. https://doi.org/10.3390/cells9122621
Lacroix M, Riscal R, Arena G, Linares LK, Le Cam L (2020) Metabolic functions of the tumor suppressor p53: implications in normal physiology, metabolic disorders, and cancer. Mol Metab 33:2–22. https://doi.org/10.1016/j.molmet.2019.10.002
Krstic J, Reinisch I, Schupp M, Schulz TJ, Prokesch A (2018) p53 functions in adipose tissue metabolism and homeostasis. Int J Mol Sci 19(9):2622. https://doi.org/10.3390/ijms19092622
Kung CP, Leu JI, Basu S, Khaku S, Anokye-Danso F, Liu Q, George DL, Ahima RS, Murphy ME (2016) The P72R polymorphism of p53 predisposes to obesity and metabolic dysfunction. Cell Rep 14(10):2413–2425. https://doi.org/10.1016/j.celrep.2016.02.037
Kim BY, Lee SY, Chung SK (2021) Differential transcriptional regulation of polymorphic p53 codon 72 in metabolic pathways. Int J Mol Sci 22(19):10793. https://doi.org/10.3390/ijms221910793
Kung CP, Liu Q, Murphy ME (2017) The codon 72 polymorphism of p53 influences cell fate following nutrient deprivation. Cancer Biol Ther 18(7):484–491. https://doi.org/10.1080/15384047.2017.1323595
Leu JI, Murphy ME, George DL (2013) The p53 codon 72 polymorphism modifies the cellular response to inflammatory challenge in the liver. J Liver 2(1):117. https://doi.org/10.4172/2167-0889.1000117
Kung CP, Basu S, Murphy ME (2016) A link between TP53 polymorphisms and metabolism. Mol Cell Oncol 3(4):e1173769. https://doi.org/10.1080/23723556.2016.1173769
Basu S, Gnanapradeepan K, Barnoud T, Kung CP, Tavecchio M, Scott J, Watters A, Chen Q, Kossenkov AV, Murphy ME (2018) Mutant p53 controls tumor metabolism and metastasis by regulating PGC-1α. Genes Dev 32(3–4):230–243. https://doi.org/10.1101/gad.309062.117
Punja HK, Nanjappa DP, Babu N, Kalladka K, Shanti Priya Dias B, Chakraborty G, Rao SM, Chakraborty A (2021) TP53 codon 72 polymorphism and type 2 diabetes: a case-control study in South Indian population. Mol Biol Rep 48(6):5093–5097. https://doi.org/10.1007/s11033-021-06505-8
Słomiński B, Skrzypkowska M, Ryba-Stanisławowska M, Myśliwiec M, Trzonkowski P (2021) Associations of TP53 codon 72 polymorphism with complications and comorbidities in patients with type 1 diabetes. J Mol Med 99(5):675–683. https://doi.org/10.1007/s00109-020-02035-1
Guo D, Fang L, Yu X, Wang C, Wang Y, Guo W (2021) Different roles of TP53 codon 72 polymorphism in type 2 diabetes and its complications: evidence from a case-control study on a Chinese han population. Int J Gen Med 14:4259–4268. https://doi.org/10.2147/IJGM.S322840
Expert Panel on Detection, Evaluation, and Treatment of High Blood Cholesterol in Adults (2001) Executive summary of the third report of The National Cholesterol Education Program (NCEP) Expert panel on detection, evaluation, and treatment of high blood cholesterol in adults (Adult Treatment Panel III). JAMA 285(19):2486–2497. https://doi.org/10.1001/jama.285.19.2486
Tandon N, Srivastava AN, Fatima N, Raza ST, Kumar V (2017) p53 codon 72 gene polymorphism studies and p53 expression by immunohistochemistry in oral lesions as risk factor for malignancy. Int J Appl Basic Med Res 7(4):243–246. https://doi.org/10.4103/ijabmr.IJABMR_205_16
Horita N, Kaneko T (2015) Genetic model selection for a case-control study and a meta-analysis. Meta Gene 22(5):1–8. https://doi.org/10.1016/j.mgene.2015.04.003
Pourhoseingholi MA, Vahedi M, Rahimzadeh M (2013) Sample size calculation in medical studies. Gastroenterol Hepatol Bed Bench 6(1):14–17
Ramachandran A (2002) Epidemiology of type 2 diabetes in Indians. J Indian Med Assoc 100(7):425–427
Ashenhurst JR, Sazonova OV, Svrchek O, Detweiler S, Kita R, Babalola L, McIntyre M, Aslibekyan S, Fontanillas P, Shringarpure S; 23andMe Research Team; Pollard JD, Koelsch BL (2022) A polygenic score for type 2 diabetes improves risk stratification beyond current clinical screening factors in an ancestrally diverse sample. Front Genet 13:871260. https://doi.org/10.3389/fgene.2022.871260
Rochlani Y, Pothineni NV, Kovelamudi S, Mehta JL (2017) Metabolic syndrome: pathophysiology, management, and modulation by natural compounds. Ther Adv Cardiovasc Dis 11(8):215–225. https://doi.org/10.1177/1753944717711379
Alberti KG, Zimmet P, Shaw J (2006) Metabolic syndrome: a new world-wide definition—a consensus statement from the international diabetes federation. Diabet Med 23(5):469–480. https://doi.org/10.1111/j.1464-5491.2006.01858.x
Gaulton KJ, Willer CJ, Li Y, Scott LJ, Conneely KN, Jackson AU, Duren WL, Chines PS, Narisu N, Bonnycastle LL, Luo J, Tong M, Sprau AG, Pugh EW, Doheny KF, Valle TT, Abecasis GR, Tuomilehto J, Bergman RN, Collins FS, Boehnke M, Mohlke KL (2008) Comprehensive association study of type 2 diabetes and related quantitative traits with 222 candidate genes. Diabetes 57(11):3136–3144. https://doi.org/10.2337/db07-1731
Burgdorf KS, Grarup N, Justesen JM, Harder MN, Witte DR, Jørgensen T, Sandbæk A, Lauritzen T, Madsbad S, Hansen T; DIAGRAM Consortium; Pedersen O (2011) Studies of the association of Arg72Pro of tumor suppressor protein p53 with type 2 diabetes in a combined analysis of 55,521 Europeans. PLoS One 6(1):e15813. https://doi.org/10.1371/journal.pone.0015813
Sliwinska A, Kasznicki J, Kosmalski M, Mikolajczyk M, Rogalska A, Przybylowska K, Majsterek I, Drzewoski J (2017) Tumour protein 53 is linked with type 2 diabetes mellitus. Indian J Med Res 146(2):237–243. https://doi.org/10.4103/ijmr.IJMR_1401_15
Lagares MH, Silva KSF, Barbosa AM, Rodrigues DA, Costa IR, Martins JVM, Morais MP, Campedelli FL, Moura KKVO (2017) Analysis of p53 gene polymorphism (codon 72) in symptomatic patients with atherosclerosis. Genet Mol Res. https://doi.org/10.4238/gmr16039721
Alkhalaf M, Al-Bustan S, Hamoda H, Abdella N (2007) Polymorphism of p53 gene codon 72 in Kuwaiti with coronary artery disease and diabetes. Int J Cardiol 115(1):1–6. https://doi.org/10.1016/j.ijcard.2006.01.029
Rivlin N, Brosh R, Oren M, Rotter V (2011) Mutations in the p53 tumor suppressor gene: important milestones at the various steps of tumorigenesis. Genes Cancer 2(4):466–474. https://doi.org/10.1177/1947601911408889
Gloria-Bottini F, Banci M, Saccucci P, Magrini A, Bottini E (2011) Is there a role of p53 codon 72 polymorphism in the susceptibility to type 2 diabetes in overweight subjects? A study in patients with cardiovascular diseases. Diabetes Res Clin Pract 91(3):e64–e67. https://doi.org/10.1016/j.diabres.2010.11.031
Wu H, Ballantyne CM (2020) Metabolic inflammation and insulin resistance in obesity. Circ Res 126(11):1549–1564. https://doi.org/10.1161/CIRCRESAHA.119.315896
Wondmkun YT (2020) Obesity, insulin resistance, and type 2 diabetes: associations and therapeutic implications. Diabetes Metab Syndr Obes 9(13):3611–3616. https://doi.org/10.2147/DMSO.S275898
Acknowledgements
The authors would like to thank Director, AIIMS-Raipur and Head, Department of Biochemistry, AIIMS-Raipur for all the support.
Funding
STS (2020) ICMR funding to DM; Intramural funding from All India Institute of Medical Sciences, Raipur, India to PA.
Author information
Authors and Affiliations
Contributions
DM, PA: Sample collection, Investigation, MJJ: Statistical Analysis; JA: Conceptualization, Methodology, Writing.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The participants were recruited from the outpatient department of All India Institute of Medical Sciences, Raipur, India. The Institute Ethics Committee of All India Institute of Medical Sciences, Raipur, approved the study design. The study was conducted according to the criteria set by the Declaration of Helsinki. Each subject signed informed consent before participating in the study. The samples were assigned unique codes to deidentify and anonymize them.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Figure 1.
Distribution of Diabetes Mellitus cases in the studied population. Table 1. Arthrometric parameters in the subjects.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Abraham, J., Mahapatra, D., Agrawal, P. et al. Association of p53 codon 72 polymorphism with weight and metabolic diseases in a Central Indian population. Egypt J Med Hum Genet 25, 6 (2024). https://doi.org/10.1186/s43042-024-00472-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43042-024-00472-y