
Abstract
1H NMR spectroscopy was adopted to determine compositional changes (mainly sugars, organic acids and amino acids) involved in cold-stored immature soybean grains after exogenous spermine treatment. Significant changes of sugars, including sucrose, galactose, myo-inositol, glucose and fructose were detected in soybean after spermine treatment. As for the organic acids related to tricarboxylic acid cycle, the levels of malic and fumaric acids decreased but the level of citric acid increased. However, no significant changes were observed for amino acids in spermine-treated soybeans. By using metabolic profile analysis, a difference was observed between the aging of soybean grains as such and those treated with spermine. This study provides an insight into the accumulation of metabolites in postharvest immature soybeans after exogenous spermine-treatment.
Graphical abstract

Similar content being viewed by others
Introduction
Immature vegetable soybean (Glycine max L. Merr.) is a popular food in China, Japan and Korea, due to its favorable nutrition, flavor and taste (Young et al. 2000). However, immature vegetable soybean usually has a high respiratory rate and its quality declines rapidly after harvest. As compared to the intact soybean grains, soybean pod is more easily to perish when exposed to postharvest storage (Su et al. 2002). Even if soybean is stored in cold conditions (below 5 °C), they will suffer chilling injury (CI) (Aghdam and Bodbodak 2014). In recent years, a number of postharvest techniques, including polyamine, methyl jasmonate (MeJA) and 1-MCP treatment (Salvador et al. 2004; Cao et al. 2010; Zhang et al. 2009), and modified atmosphere packing (MAP) (Alasalvar et al. 2005) have been used to reduce the CI in order to prolong the storage period of soybean. Polyamines, such as spermine, spermidine and putrescine, etc., are low molecular-weight compounds with several biological activities, such as anti-stress, anti-aging and other effects. Polyamines have been used to improve the cold tolerance and extend the shelf life of mango, apricot and other fruits (Martínez-Romero et al. 2002; Malik and Singh 2005). In addition, spermine treatment could significantly inhibit the water loss, rust spots-forming and decay of common bean pod (Tian et al. 2013). Our previous study preliminarily revealed exogenous spermine treatment could reduce CI in vegetable soybeans. However, the impact of spermine treatment on the metabolic compounds (sugars, organic acids, amino acids, etc.) is still unclear (Song et al. 2018).
Low temperature can alter the balance between free radical production and free radical scavenging systems (Sala 1998). There are both advantages and disadvantages for low-temperature storage. Meanwhile, exogenous chemical treatment usually has synergetic effects with low-temperature storage, leading to the change and fluctuation of metabolites (Rudell et al. 2009; Lee et al. 2012). Nowadays, metabonomics have been widely used to analyze metabolic variations in organic plants. Lee et al. (2012) investigated the changes in metabolic profile of apple fruit during controlled atmosphere (CA) storage under low oxygen atmosphere, and found majority carbohydrates and organic acids were not significantly affected, however, the levels of amino acids and volatile metabolites were significantly affected by 1-MCP treatment. With the help of metabolomics methods, Zhu et al. (2015) found the stress-related metabolites, such as D-xylose, D-galactose, ornithine and γ-aminobutyric acid (GABA),increased in the 24-epibrassinolide-treated citrus fruit, which greatly accelerated the selection of new sustainable treatment.
Proton nuclear magnetic resonance (1H NMR) spectroscopy is one of the most effective techniques for the analysis and identification of plant samples (Simmler et al. 2014; Jiang et al. 2015). Sugimoto et al. (2010) investigated the temporal variation of edamame’s metabolic profiles and sensory characteristics during transportation and consumption. However, there is no available data about the effect of exogenous spermine on the properties of immature vegetable soybeans during postharvest storage. Therefore, the purpose of this study is to investigate the effect of exogenous spermine on the compositional changes (sugars, organic acids and amino acids) of cold-stored immature vegetable soybeans by using 1H NMR based metabolomics approach. The information obtained will reveal the metabolic characteristics of spermine-treated immature soybean.
Materials and methods
Plant materials
Vegetable soybean (Glycine max L. ‘Xindali No.1’) pods were harvested 45 days after flowering in October 2015 from Luhe Animal Science Base, Jiangsu Academy of Agricultural Sciences (Nanjing, China) situated at 32.08° N 118.40° E. Vegetable soybeans of similar size and maturity without physical injury or infection were selected. The fresh pods were quickly transported to the Institute of Agro-Products Processing in Jiangsu Academy of Agricultural Sciences and cooled down to 5 °C overnight.
Storage treatment
The homogeneous pods were soaked in appropriate amount of spermine (1.0 mmol L− 1) (Song et al. 2018) or distilled water (control) and stirred for 20 min. The ratio of pod to solution was about 1:2 (w/v). The treated pods were then transferred to the basket and drained at room temperature. The soybean pods were then stored in micro-perforated polyethylene bags at an ambient temperature of 1 ± 1 °C with a relative humidity of 85–95% in darkness. Each bag contained 500 g soybean pods, the bag had a thickness of 0.04 mm, the hole diameter was about 0.5 mm and the micro-perforations spacing of approximately 2 mm. The storage period was up to 60 days. After 0, 15, 30, 45 and 60 days (15-day intervals), hulled soybean samples were directly frozen in liquid nitrogen before being ground to a fine powder. The samples were then kept at − 80 °C until further determination of the main metabolites. Each treatment was replicated five times.
1H NMR analysis
The metabolites in stored immature vegetable soybeans were analyzed in accordance with the previous method (Song et al. 2013).
Sample preparation
The crushed material (250 mg) was transferred to a 2-ml centrifuge tube. 1 mL 50% (v/v) of methanol-water mixture was added to the test tube. The tube was vortexed for 30 s, and then the probe sonicator was used to sonicate it on ice three times, each time for 4 s. The material was then low-temperature centrifuged at 12, 200×g for 15 min. The aqueous phase was collected and blew with nitrogen near the surface for about 40 min. The mixture was evaporated in a vacuum centrifugal evaporator until dried. The dried sample was redissolved in 450 μL of 0.1 M phosphate buffer (pH 7.29) and 50 μL Anachro certified DSS (2, 2-dimethyl-2-silapentane-5-sulphonate) standard solution. The mixture was then vortexed for 10 s and centrifuged at 12, 200×g for 15 min, the supernatant was collected and used in 1H-NMR spectroscopy.
1H-NMR spectroscopy
All 1H NMR spectra were obtained at 298 K by a Bruker AV III-600 NMR spectrometer (Bruker BioSpin GmbH, Rheinstetten, Germany) equipped with an inverse cryoprobe operating at a proton NMR frequency of 600.13 MHz. Each sample was scanned 32 times by spectral width (SW) 12,019.23 Hz, pulse width (PW) 10 μs, and relaxation delay 1.0 s. The spectra referred to the peak height of DSS. To measure all the NMR signals, 5 mM DSS was used as an internal chemical shift standard.
Quantification of the metabolites
The quantification of metabolite concentrations was performed using the Chenomx NMR Suite v.7.5 module (Chenomx Inc., Edmonton, Canada). DSS was used as an internal standard for chemical shifts set to 0 ppm and as a quantitative reference signal. The quantification was carried out by comparing the integral of a given reference signal (DSS-d6) with a signal in a compound library containing chemical shift and peak multiplicity. The identifiable metabolites were quantified by matching them to the Chenomx 600 MHz Library.
Data analysis
Chemometrics was performed using unsupervised principal component analysis (PCA) in the R software environment. The differences of metabolite concentration were evaluated by Duncan’s multiple-range test with the SAS 9.1 software (SAS Institute, Cary, NC, USA). The difference was considered significant at P < 0.05.
Results and discussion
Identification of compounds by 1H-NMR
All the stored immature soybean grains were extracted with 50% (v/v) methanol-water mixture in order to collect hydrophilic constituents, such as sugars and amino acids. The chemical metabolites of all the samples were almost identical. The methanol aqeous extract of fresh immature soybean was characterized and shown in supplementary Fig. 1. The resonance of these metabolites was compared to the signals of Chenomx NMR Suite 7.5 database and internal databases measured under the same conditions. As a result 45 kinds of compounds including sugars, organic acids and amino acids were identified.
Classification of stored immature soybean samples
Immature soybeans were prone to decay in the initial storage stage regardless of spermine treatment. As reported, spermine could reduce the quality loss as prolonging storage time (Song et al. 2015). To compare the composition of metabolites between untreated and spermine-treated immature soybeans during postharvest storage, PCA was used to analyze the 1H NMR data set of the main extracts. Some groups were clearly discriminated in PC1 and PC2 which accounted for 98.7 and 0.6% of the total variance, respectively (Fig. 1). Groups SD15 and D15 formed a tight cluster, suggesting they had identical or closely related metabolite compositions in these stages. A similar pattern was also observed in groups SD30 and D30. However, a clear separation was observed between groups SD60/D60 and SD45/D45. The heat map of immature soybean samples with different storage periods exhibited three major clusters (Fig. 2).

Score and loading plots of effect of exogenous spermine on metabolite profiles of cold-stored immature vegetable soybeans

Heatmap of effect of exogenous spermine on metabolite profiles of cold-stored immature vegetable soybeans. Red colour indicates higher metabolite concentration than the median of the samples, blue indicates a lower concentration. The metabolites are listed on the right side of the heat map with metabolite clustering on the left. All samples are listed on the top of the map as well as the sample clustering tree
It is possible to find the metabolites that contributed most to the separation of distinct clusters using PCA approach. By observing the corresponding signals in loading scatter plots (Fig. 1), the compounds such as amino acids (glycine, serine, threonine, and β-alanine), organic acids (aspartic acid, succinic acid, and galactaric acid) and myo-inositol were all present at significantly higher levels (P < 0.05) in stages SD15 and D15 as compared to stages SD30 and D30. In contrast, stages SD60 and D60 showed significantly lower levels of sugars (P < 0.05) as compared to stages SD15 and D15. Spermine-treated groups contained considerably higher levels of sucrose, malic acid, aspartate and asparagine as compared to the non-treated groups during the same storage time, and these metabolites drastically decreased during postharvest when the grain was detached from the plant. Although these compounds appear to characterize the storage life of immature soybeans, this approach should be tested in other varieties, as they may be specific to the immature soybean varieties studied.
Effect of exogenous spermine on metabolite concentrations of immature soybeans
Sugars
The major sugars detected in immature soybeans during storage were sucrose, myo-inositol, fructose-6-phosphate and glucose, whereas fructose, galactaric acid and other sugars were only present in minor concentrations. In our previous study, postharvest sucrose metabolism induced by spermine was fully discussed (Song et al. 2015). Glucose and fructose in freshly harvested immature soybeans accumulated to higher levels as compared to those required for respiration. Both glucose and fructose occurred in equal amounts in immature soybeans during storage, which was caused by invertase activity that was responsible for decomposing sucrose (Oms-Oliu et al. 2011). During storage, the sucrose concentration of soybean was reduced by more than 90%. Sucrose concentration in spermine-treated immature soybeans was significant higher than the control group (P < 0.05), while glucose was not affected by spermine-treatment and gradually decreased when storage time prolonged. Meanwhile, fructose concentration in the spermine-treated immature soybeans was lower than that of the control group in the early storage stage, implying spermine inhibited sucrose exhaustion. Regardless of spermine treatment, as compared with fresh immature soybeans, the levels of sucrose, fructose and glucose all decreased, which was related with the inhibition of sucrose hydrolysis and the rapid utilization of fructose and glucose as carbon sources. Under spermine treatment, the hydrolysis of sucrose was inhibited, which was benefit to maintain the lipid bilayer structure. As a result, glucose acted as a direct substrate and played an important role in energy metabolism and oxygen free radical formation (Mbong et al. 2017).
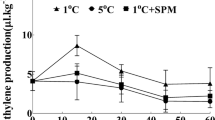
Sugar phosphates are vital intermediates of central metabolism, but its concentration is commonly constrained by the high turnover rate (Hasunuma et al. 2009). Figure 3 demonstrates the concentration of fructose 6-phosphate mostly in the spermine-treated immature soybeans decreased compared to the fresh soybeans. However, the catabolism of sugar and sugar phosphates through the glycolytic process and tricarboxylic acid cycle (TCA cycle) might increase with the change of respiration during postharvest storage, which may be the main cause of carbon flux in cereals (Egli 1999). In the early storage stage, the concentration of fructose 6-phosphate presented an increasing trend. Although spermine could inhibit the increase of fructose 6-phosphate, its inhibition ability was very limited especially at the late storage stage.

Effect of exogenous spermine on the concentration of major sugars in cold-stored immature vegetable soybeans
Galacturonic acid has been identified as the major cell wall component associated with ripening. The level of galacturonic acid increased with the softening of soybeans during maturation and ripening, indicating cell wall complexes destructed. Spermine-treated group had a significant lower galacturonic acid concentration than the control group (P < 0.05). Levels of myo-inositol were higher at the early storage stage than the late storage stage. Myo-Inositol is a vital precursor responsible for the biosynthesis of many cell wall polysaccharides. Spermine-treated group had a significant lower myo-inositol level than the control group after 45-day storage (P < 0.05).
Amino acids
Composition of amino acids is a key factor reflecting the flavor (umami taste) and nutritional quality of soybeans (Snowden et al. 2015). Effect of spermine-tretment on the composition of amino acids in immature soybeans was not obvious. Only some amino acids varied, which could be related with storage time (Fig. 4). Most of the amino acids in immature soybeans including GABA, arginine, asparagine, aspartic acid, glutamic acid, β-alanine, isoleucine, threonine, valine, leucine and serine dramatically increased at the early storage stage, and then decreased gradually to a certain extent at the late storage stage, implying that some protease activities increased at the initial storage stage. Similar results were obtained in broccoli and asparagus (Hurst and Clark 1993; Page et al. 2001). Meanwhile, phenylalanine increased in the whole storage period but spermine-treatment obviously inhibited the trend especially at the late storage stage. Spermine-treatment also inhibited the increase of arginine. Since the transamination of arginine was weak, the degradation of amino acids could easily form toxic ammonia (Hurst and Clark 1993; Winter et al. 2015). The amino acids (isoleucine, valine and leucine) with branched chains gradually increased at the early storage stage and then after 45-day storage. These amino acids were the main nitrogenous sources for cell membranes (Platell et al. 2000). Since membrane damage is the key signal of postharvest senescence, our result indicated that both low temperature and spermine-treatment could induce the resistance of self-aging.

Effect of exogenous spermine on the concentration of amino acids in cold-stored immature vegetable soybeans
There were no obvious differences in the levels of alanine, aspartic acid and glutamic acid between spermine-treatment and the control groups. All the above components first increased and then decreased, implying that they could act as active precursors of other macromolecules. Glutamic acid was involved in nitrogen metabolism and TCA circle, and aspartic acid and alanine were both involved in transamination. Glutamine is considered to be a vital compound of nitrogen transport in plants, thus this pattern was supposed to reflect the need growing with aging-related processes. On the other hand, glutamic acid is the most abundant free amino acid during storage. Its content in immature grains was very low but its levels progressively increased with grain aging.
Betaine, one of amino acid derivatives, usually acts as the defense product against adverse environmental conditions, is also an important non-toxitic osmotic adjustment in higher plant (Di Martino et al. 2003). In this study, both betaine and its synthetic substrate-choline accumulated as induced by spermine-treatment.
Organic acids
Organic acids are important components in immature soybean, which affect the taste and overall quality. Ten kinds of organic acids in immature soybeans were detected, and most of them are important TCA intermediates. Malic acid is the main organic acid, which was followed by citric acid, fumaric acid, succinic acid, pyruvic acid and tartaric acid (Fig. 5). The citric acid cycle involves the biosynthesis of fumaric acid, isocitrate, and succinic acid, many of which are precursors of amino acid biosynthesis (Carrari and Fernie 2006). Citric acid content increased and malic acid content decreased in cold-stored immature soybeans by spermine-treatment during the postharvest stage. The reason might be that the activity of malidase, which decarboxylated l-malic acid to pyruvate, was inhibited. The inhibited activities of malate dehydrogenase and citrate synthase led to a decline in malate and the prior accumulation of citrate. Fumaric acid, a precursor of malic acid in the citric acid cycle, decreased after harvest, while succinic acid increased during storage. This may indicate that these metabolites were up-regulated during aging. Although the general structure of plant TCA pathway is well known, little is known about its regulation (Fernie et al. 2004). The glycolysis and TCA cycle are the prevailing carbon fluxes during soybean aging.

Effect of exogenous spermine on the concentration of organic acids in cold-stored immature vegetable soybeans
Moreover, the concentrations of 2-hydroxyisobutyric acid, 2-oxoglutarate, citric acid, fumarate, succinate and acetic acid in spermine-treated group were higher than the control group. Nevertheless, no obvious differences in concentrations of tartaric acid, cis-aconitic acid and pyruvate were observed between spermine-treatment and the control groups. Organic acid changed with TCA due to spermine stress, and the metabolism of organic acid was affected. By adjusting the content of organic acids, the cellular homeostasis was restored. In vegetable soybean, malic acid was the highest among all the organic acids, malic acid is the intermediate of TCA and glyoxylate cycle. It could maintain cell osmotic pressure and charge-balance and was involved in the regulation of soybean stomatal opening, playing an important role in the stress adaptation of vegetable soybean. In this study, spermine-treatment promoted malic acid accumulation in vegetable soybean, which was similar to citric acid. Under stress conditions of spermine-treatment, the accumulations of citric acid and succinic acid in soybeans indicated TCA inhibited. Acetic acid, 2-oxoglutarate and succinic acid contents in vegetable soybeans were induced by spermine-treatment. Although lipids were not detected in the study, it was speculated that increased acid content may be related to lipid metabolism strengthened based on the literature (Thompson et al. 1998). Under stress conditions, the utilization of acetyl coenzyme A declined, which may be caused by acid accumulation. 2-oxoglutarate is the essential substrate in glutamic acid synthesis pathway, which is also an important organic acid in ammonia assimilation. Thus, 2-oxoglutarate accumulation had a positive effect on the synthesis of glutamate. It might be used as the substrate for the generation of ATP (Giovannoni 2004).
Conclusions
A metabolic profiling method was developed to describe the changes in the composition of immature vegetable soybeans during shelf life. Most compounds can be clustered together showing either increasing (e.g., sucrose, fructose, GABA, malate and fumarate) or decreasing (e.g., myo-inositol and galactaric acid) levels in sepermine-treated soybeans with some compounds being markedly characterized. Some of the observed changes may be related to phenomena known to be associated with grain aging. During cold storage, exogenous spermine effectively inhibited the conversion of sucrose to fructose and glucose and sugar consumption, in the meantime, reduced the contents of malic acid and citric acid, which indicated that spermine treatment might inhibit internal signal transduction, sugar metabolism and TCA rate, to delay the quality deterioration of immature vegetable soybeans. Exceptionally, no significant effects on major free amino acids were observed by spermine treatment. Metabonomics methods combined with appropriate multivariate data analysis may be a useful tool for investigating and assessing differences in edible legume metabolites under different storage conditions.
Additionally, we can speculate the mechanism of grain aging activates metabolic pathways which subsequently influence among others; these activities consume energy as well as carbon skeleton building block such as amino acids and organic acids. The overall results of the study may help in understanding the compositional changes and the mechanism underlying immature soybeans aging which later may be beneficial in developing effective ways to enhance shelf life and quality of grain especially during postharvest. Furthermore, as the nutritional quality of grain is closely related to the supply of metabolites such as sugars, organic acids, amino acids and secondary metabolites, a better understanding of the metabolite composition of the grain is required, it may help to improve the nutritional value of this important immature soybean for human consumption.
Availability of data and materials
Please contact the corresponding author for information.
Change history
31 July 2020
An amendment to this paper has been published and can be accessed via the original article.
Abbreviations
- TCA:
-
Tricarboxylic acid cycle
- NMR:
-
Nuclear magnetic resonance spectroscopy
- PCA:
-
Principal component analysis
- PC:
-
Principal component
- CI:
-
Chilling injury
- MeJA:
-
Methyl jasmonate
- 1-MCP:
-
1-methylcyclopropene
- MAP:
-
Modified atmosphere packing
- CA:
-
Controlled atmosphere
- DSS:
-
2, 2-dimethyl-2-silapentane-5-sulphonate
- GABA:
-
γ-aminobutyric acid
References
Aghdam, M. S., & Bodbodak, S. (2014). Postharvest heat treatment for mitigation of chilling injury in fruits and vegetables. Food and Bioprocess Technology, 7(1), 37–53.
Alasalvar, C., Al-Farsi, M., Quantick, P. C., Shahidi, F., & Wiktorowicz, R. (2005). Effect of chill storage and modified atmosphere packaging (MAP) on antioxidant activity, anthocyanins, carotenoids, phenolics and sensory quality of ready-to-eat shredded orange and purple carrots. Food Chemistry, 89(1), 69–76.
Cao, S., Zheng, Y., Wang, K., Rui, H., & Tang, S. (2010). Effect of methyl jasmonate on cell wall modification of loquat fruit in relation to chilling injury after harvest. Food Chemistry, 118(3), 641–647.
Carrari, F., & Fernie, A. R. (2006). Metabolic regulation underlying tomato fruit development. Journal of Experimental Botany, 57(9), 1883–1897.
Di Martino, C., Delfine, S., Pizzuto, R., Loreto, F., & Fuggi, A. (2003). Free amino acids and glycine betaine in leaf osmoregulation of spinach responding to increasing salt stress. The New Phytologist, 158(3), 455–463.
Egli, D. B. (1999). Variation in leaf starch and sink limitations during seed filling in soybean. Crop Science, 39(5), 1361–1368.
Fernie, A. R., Carrari, F., & Sweetlove, L. J. (2004). Respiratory metabolism: Glycolysis, the TCA cycle and mitochondrial electron transport. Current Opinion in Plant Biology, 7(3), 254–261.
Giovannoni, J. J. (2004). Genetic regulation of fruit development and ripening. Plant Cell, 16(suppl 1), S170–S180.
Hasunuma, T., Harada, K., Miyazawa, S. I., Kondo, A., Fukusaki, E., & Miyake, C. (2009). Metabolic turnover analysis by a combination of in vivo 13C-labelling from 13CO2 and metabolic profiling with CE-MS/MS reveals rate-limiting steps of the C3 photosynthetic pathway in Nicotiana tabacum leaves. Journal of Experimental Botany, 61(4), 1041–1051.
Hurst, P. L., & Clark, C. J. (1993). Postharvest changes in ammonium, amino acids and enzymes of amino acid metabolism in asparagus spear tips. Journal of the Science of Food and Agriculture, 63(4), 465–471.
Jiang, X., Kim, S. H., Oh, T. J., Huang, L. F., & Choi, H. K. (2015). Proton nuclear magnetic resonance spectrometry-based metabolic characterization of Panax Notoginseng roots. Analytical Letters, 48(8), 1341–1354.
Lee, J., Rudell, D. R., Davies, P. J., & Watkins, C. B. (2012). Metabolic changes in 1-methylcyclopropene (1-MCP)-treated ‘empire’ apple fruit during storage. Metabolomics, 8, 742–753.
Malik, A. U., & Singh, Z. (2005). Pre-storage application of polyamines improves shelf-life and fruit quality of mango. The Journal of Horticultural Science and Biotechnology, 80(3), 363–369.
Martínez-Romero, D., Serrano, M., Carbonell, L., Burgos, F., & Valero, D. (2002). Effects of postharvest putrescine treatment on extending shelf life and reducing mechanical damage in apricot. Journal of Food Science, 67(5), 1706–1711.
Mbong, V. B. M., Ampofo-Asiama, J., Hertog, M. L., Geeraerd, A. H., & Nicolai, B. M. (2017). The effect of temperature on the metabolic response of lamb’s lettuce (Valerianella locusta (L.) Laterr.) cells to sugar starvation. Postharvest Biology and Technology, 125, 1–12.
Oms-Oliu, G., Hertog, M. L. A. T. M., Van de Poel, B., Ampofo-Asiama, J., Geeraerd, A. H., & Nicolaï, B. M. (2011). Metabolic characterization of tomato fruit during preharvest development, ripening, and postharvest shelf-life. Postharvest Biology and Technology, 62, 7–16.
Page, T., Griffiths, G., & Buchanan-Wollaston, V. (2001). Molecular and biochemical characterization of postharvest senescence in broccoli. Plant Physiology, 125, 718–727.
Platell, C., Kong, S. E., McCauley, R., & Hall, J. C. (2000). Branched-chain amino acids. Journal of Gastroenterology and Hepatology, 15(7), 706–717.
Rudell, D. R., Mattheis, J. P., & Hertog, M. L. (2009). Metabolomic change precedes apple superficial scald symptoms. Journal of Agricultural and Food Chemistry, 57, 8459–8466.
Sala, J. M. (1998). Involvement of oxidative stress in chilling injury in cold-stored mandarin fruits. Postharvest Biology and Technology, 13, 255–261.
Salvador, A., Arnal, L., Monterde, A., & Cuquerella, J. (2004). Reduction of chilling injury symptoms in persimmon fruit cv. ‘Rojo Brillante’ by 1-MCP. Postharvest Biology and Technology, 33, 285–291.
Simmler, C., Napolitano, J. G., McAlpine, J. B., Chen, S. N., & Pauli, G. F. (2014). Universal quantitative NMR analysis of complex natural samples. Current Opinion in Biotechnology, 25, 51–59.
Snowden, C. J., Thomas, B., Baxter, C. J., Smith, J. A. C., & Sweetlove, L. J. (2015). A tonoplast Glu/asp/GABA exchanger that affects tomato fruit amino acid composition. The Plant Journal, 81(5), 651–660.
Song, J., Liu, C., Li, D., & Gu, Z. (2013). Evaluation of sugar, free amino acid, and organic acid compositions of different varieties of vegetable soybean (Glycine max [L.] Merr). Industrial Crops and Products, 50, 743–749.
Song, J., Wang, Y., Liu, C., & Li, D. (2015). Effect of exogenous spermine on quality and sucrose metabolism of vegetable soya bean (Glycine max L.) during cold storage. International Journal of Food Science & Technology, 50, 1697–1703.
Song, J., Wu, G., Liu, C., & Li, D. (2018). Effect of exogenous spermine on chilling injury and antioxidant defense system of immature vegetable soybean during cold storage. Journal of Food Science and Technology, 55(10), 4297–4303.
Su, X., Zheng, Y., Feng, L., & Zhang, L. (2002). Effects of exogenous me-JA on Postharvet senescence and decay of vegetable soybean pods. Journal of Plant Physiology and Molecular Biology, 29, 52–58 (in Chinese with English abstract).
Sugimoto, M., Goto, H., Otomo, K., Ito, M., Onuma, H., Suzuki, A., Sugawara, M., Abe, S., Tomita, M., & Soga, T. (2010). Metabolomic profiles and sensory attributes of edamame under various storage duration and temperature conditions. Journal of Agricultural and Food Chemistry, 58, 8418–8425.
Thompson, J. E., Froese, C. D., Madey, E., Smith, M. D., & Hong, Y. (1998). Lipid metabolism during plant senescence. Progress in Lipid Research, 37(2–3), 119–141.
Tian, W. N., Miao, Y., Cao, J. K., & Jiang, W. B. (2013). Effects of vacuum pulse infiltration pf exogenous spermine on quality of postharvest common bean. Transactions of the CSAM, 44(1), 131–136 (in Chinese with English abstract).
Winter, G., Todd, C. D., Trovato, M., Forlani, G., & Funck, D. (2015). Physiological implications of arginine metabolism in plants. Frontiers in Plant Science, 6, 534.
Young, G., Mebrahtu, T., & Johnson, J. (2000). Acceptability of green soybeans as a vegetable entity. Plant Food for Human Nutrition, 55(4), 323–333.
Zhang, W., Jiang, B., Li, W., Song, H., Yu, Y., & Chen, J. (2009). Polyamines enhance chilling tolerance of cucumber (Cucumis sativus L.) through modulating antioxidative system. Scientia Horticulturae-Amsterdam, 122(2), 200–208.
Zhu, F., Yun, Z., Ma, Q., Gong, Q., Zeng, Y., Xu, J., Cheng, Y., & Deng, X. (2015). Effects of exogenous 24-epibrassinolide treatment on postharvest quality and resistance of Satsuma mandarin (Citrus unshiu). Postharvest Biology and Technology, 100, 8–15.
Acknowledgements
Not applicable.
Funding
The work was financially supported by the North Jiangsu Science and Technology project (No. SZ-YC2018002) and the Tongzhou 510 talent project.
Author information
Authors and Affiliations
Contributions
All authors have made significant contributions to the analysis, writing, and improvement of the original manuscript, and have read and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors state that there is no competition for financial or non- financial benefits.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Song, J., Wu, G., Li, T. et al. Changes in the sugars, amino acids and organic acids of postharvest spermine-treated immature vegetable soybean (Glycine max L. Merr.) as determined by 1H NMR spectroscopy. Food Prod Process and Nutr 2, 7 (2020). https://doi.org/10.1186/s43014-020-00021-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s43014-020-00021-1