
Abstract
Background
Among the phytosanitary problems, affecting tomato crops in greenhouses heated by geothermal water in southern Tunisia, the tomato leaf miner, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) is gaining prominence from year to year.
Results
This study determined the susceptibility of eggs, second-instar larvae of T. absoluta to commercial formulations of the entomopathogenic fungi (EPF) Beauveria bassiana and Lecanicillium muscarium at different doses (1010, 109, 108, 107 and 106 spores/ml). Pathogenicity tests were carried out under controlled laboratory conditions at 27 ± 1 °C, and RH of 75 ± 5%, showed that the three commercial formulations of B. bassiana R444 (Bb-Protec®), B. bassiana ATCC 74,040 (Naturalis®) and L. muscarium Ve6 (Mycotal®) were effective against eggs and second instar larvae of T. absoluta. Mortality rates of eggs and larvae were high with increasing the concentrations of the fungi. B. bassiana ATCC 74,040 was the most effective one, with an egg mortality rate of (71.42%) and larval mortality was greater than (80%), at the highest concentration of 1010 spores/ml. The lethal concentration required to kill 50% of eggs was 1.56 × 108, 1.73 × 109 and 1.09 × 1010 conidia/ml for B. bassiana ATCC, B. bassiana R444, and L. muscarium, respectively. After 7 days, the median lethal concentration (LC50) values against larvae were the lowest for B. bassiana ATCC with 2.63 × 107 and 4.76 × 106 conidia/ml by way 1 and way 2 treatments, respectively. Thus, these three formulations affected the emergence of pest’s adults. The fungi significantly reduced pupation and adult emergence. Only 50.25, 60.15 and 69.16% of T. absoluta adults emerged in the treatments with B. bassiana ATCC, B. bassiana R444, and L. muscarium, respectively.
Conclusions
Biological performances of the EPF showed the role of a biological control agent that can be played against T. absoluta within the framework of an Integrated Pest Management program.
Similar content being viewed by others

Background
The tomato leaf miner, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) is an insect native to South America (Desneux et al. 2011). It is considered one of the most devastating pests of tomato crop (Solanum lycopersicum L., Solanaceae) (Cherif et al. 2019). T. absoluta has a high reproductive capacity. The duration of the development cycle depends essentially on the environmental conditions, in particular the temperature. (Tropea Garzia et al. 2012). It soon has spread to several countries outside its original region (Han et al. 2019). At the end of 2006, it was detected for the first time outside its native area of South America in the Mediterranean basin in the east of the Iberian Peninsula; southwestern Europe, including Spain and Portugal (Urbaneja et al. 2007). In the following years, this pest quickly reached destructive levels in the main tomato-growing Mediterranean coastal areas (Desneux et al. 2010). An ineffective early surveillance network, weak quarantine efforts, and increased trade are promoting its rapid spread in tomato growing areas (Han et al. 2019). Management of this leaf miner relies heavily on applications of chemical insecticides (Roditakis et al. 2013). But the failure of chemical control clearly shows the ability of this pest to develop resistance to several classes of insecticides (Roditakis et al. 2018). This genetic resistance of populations of T. absoluta is a common consequence of unbalanced use of insecticides worldwide (Guedes and Picanço 2012). Biological control represents one of the most promising options with entomopathogens (fungi, bacteria, nematodes or viruses) which can effectively control this pest (Urbaneja et al. 2012). Many studies have been conducted on EPF to control a wide range of pests which have proven to be very effective (Contreras et al. 2014). Specifically, Beauveria bassiana (Bals.) Vuill. (Ascomycota: Hypocreales) and Metarhizium anisopliae (Metsch.) Sorok (Ascomycota: Hypocreales) have been used against many insect pests for over 120 years (Contreras et al. 2014). The EPF, B. bassiana and M. anisopliae have been reported to be effective against tomato leaf miner (Tadele and Emana 2017).
Faced to the demands of the consumer, who has become more sensitive to pesticide residues, this study seemed to improve Integrated Pest Management (IPM) programs by developing biological means. This study aimed to evaluate the efficacy of EPF on eggs, second-instar larvae of T. absoluta to commercial formulations of B. bassiana and Lecanicillium muscarium fungi at different concentrations.
Methods
Insect rearing
Leaves infested by T. absoluta larvae were collected from a greenhouse culture located in the Technical Center for Protected and Geothermal Cultures in the experimental plot in Chenchou (Governorate of Gabes, ElHamma delegation in southeastern Tunisia). The insect population raised on tomato leaves was placed in plastic boxes (20*10*8) at a climatic chamber, at a temperature of 27 ± 1 °C, a RH of 75 ± 5% and a 16:8 h photoperiod. The nymphs were then transferred to other boxes containing fresh tomato leaves until adults’ emergence. Mating of T. absoluta adults was carried out on tomato plants in (9 cm diameter) pots containing peat. The pots were placed in wooden cages of dimensions (1 × 1 × 1.2 m), covered with insect-resistant fabric (20/10 threads/cm) in an experimental greenhouse. After oviposition, eggs and larvae of T. absoluta were used in bioassays.
Entomopathogenic fungi
Three commercial EPF (two isolates of B. bassiana and one of L. muscarium) were used in the study (Table 1). Viability of EPF conidia was confirmed on Potato Dextrose Agar (PDA) medium and incubated at 27 ± 1 °C for 7 days to confirm viability before use (Youssef 2015). Spore suspensions were obtained from Petri dishes containing EPF cultures. The concentrations of the suspensions were adjusted using the Malassez cell. Serial dilutions were carried out for each EPF product for the preparation of fungal suspensions and to reach the needed concentrations against T. absoluta: 1010, 109, 108,107et 106spores/ml, adding Tween 80 (0.01%) for each fungal suspension.
Pathogenicity test on Tuta absoluta eggs
On each tomato leaflet, 10 T. absoluta eggs, collected during rearing, were placed using a moistened brush. Five concentrations of each EPF were applied to T. absoluta eggs using a 0.5 l sprayer. As a control, eggs were treated with distilled water plus 80% Tween. Then, to avoid dehydration, each leaflet was placed face up on a disc of filter paper soaked in distilled water in a Petri dish 9 cm in diameter. Egg hatching and mycosis on eggs were assessed daily from the first day after treatment. Unhatched eggs were transferred to water-soaked filter papers in Petri dishes to observe the possible development of mycelium and conidia (Pires et al. 2010). The experimental protocol used was completely randomized, with five repetitions for each treatment; the experimental unit consisted of 10 eggs.
Pathogenicity test on Tuta absoluta larvae: the way 1
As in the previous experiment, five concentrations of each EPF were evaluated on second instar larvae of T. absoluta. Leaflets of tomatoes bearing active mines (stage L2) placed in 5*5 cm plastic dishes were sprayed with fungal suspensions for the two EPF (Ndereyimana et al. 2019). Control leaflets were sprayed only with distilled water and 0.01% Tween 80. In order to avoid rapid drying of the leaves, they were placed on filter paper soaked in distilled water. The experimental design was completely randomized, with five replicates, each consisting of leaflets containing 10 larvae.
Pathogenicity test on Tuta absoluta larvae: the way 2
Untreated T. absoluta larvae were brought by contact with tomato leaves treated with the same fungal suspensions and under the same conditions described above. Each batch of 10 larvae was transferred to dishes containing treated fresh tomato leaflets. Distilled water and 0.01% Tween 80 were used as a negative control. All experiments were repeated five times (Pires et al. 2010). Larval mortality was recorded daily for 7 days after inoculation. Dead larvae were incubated in Petri dishes on moistened filter papers to confirm fungal outgrowth on dead larvae.
Pathogenicity on pupation and adult emergence of Tuta absoluta
The reason for this bioassay was to see if these EPF formulations could have an effect on the fourth instar larvae of T. absoluta when they naturally drop onto silk threads to pupate in the soil (Desneux et al. 2010), therefore, whether there is an effect on adult emergence. Since these 4th instar larvae fall and pupate in the soil, sand was used to simulate natural field conditions in this trial. In this experiment, the sand was sterilized at 121 °C for 1 h in an oven and then cooled to room temperature for about 72 h before processing. Ten 4th instar larvae were exposed to 50 g of sterilized sand treated with 5 ml of fungal spore suspension at a concentration of 109/ml for each fungus with 0.01% Tween 80. Control treatments consisted of 50 g of sterilized sand treated with 5 ml of sterile distilled water containing 0.01% Tween 80. After 6 days of exposure, T. absoluta nymphs were transferred to tomato leaves placed in Petri dishes with droplets of honey solution to feed emerging adults. Emergence was observed and recorded up to the eleventh day (Akutse et al. 2020). The experiment was performed at 27 °C, 75 ± 5% RH at 16:8 h photoperiod and replicated five times.
Statistical analysis
Mortality data of T. absoluta were corrected using Abbott's formula (Abbott 1925) and checked for normality. These mortality rates were subjected to an analysis of variance and the means were compared according to Tukey's test (HSD) at a level of significance of 5%. Probit analysis (Finney 1971) was performed to estimate the median lethal concentrations (LC50) was calculated for the three EPF formulations. Values were considered significantly different if their 95% confidence limits did not overlap. All analyses were performed using SPSS 22 software.
where: A: Mean number of T. absoluta eggs/larvae untreated; B: Mean number of T. absoluta eggs/larvae treated.
Results
Pathogenicity test on Tuta absoluta eggs
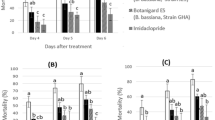
The corrected egg mortality of T. absoluta was significantly affected by the three commercial formulations based on B. bassiana ATCC, B. bassiana R444 and L. muscarium. Egg mortality increased significantly with concentrations of B. bassiana R444 (Bb-Protec®) (F = 3.957, P = 0.004), B. bassiana ATCC (Naturalis®) (F = 3.349, P = 0.011) and L. muscarium (Mycotal®) (F = 5.668, P = 0.0002). All the three EPF formulations revealed a direct relationship between concentration and mortality (Fig. 1). At the peak concentration of 1010 spores/ml, B. bassiana ATCC exhibited the highest ovicidal activity (71.42%), followed by B. bassiana R444 (55.10%) and L. muscarium (44.89%).

Corrected egg mortality of Tuta absoluta by Beauveria bassiana ATCC, B. bassiana R444 and Lecanicillium muscarium
Regression analysis of T. absoluta egg mortality caused by B. bassiana ATCC, B. bassiana R444, and L. muscarium is shown in Table 2. Indeed, the median lethal concentration required to kill 50% of the eggs was (1.56 × 108 conidia/ml) for B. bassiana ATCC. The LD90 of B. bassiana ATCC, B. bassiana R444 and L. muscarium against T. absoluta eggs was obtained by probit analysis as 6.4 × 1011, 3.65 × 1013 and 1.03 × 1014 conidia/ml (Table 2).
Pathogenicity test on Tuta absoluta larvae: the way 1
The evaluation of the corrected mortality rates recorded for the five concentration of B. bassiana ATCC, B. bassiana R444 and L. muscarium used by the first method against the second instar larvae of T. absoluta, followed the same pattern (Fig. 2). The concentration 1010 conidia/ml killed significantly more larvae than the concentration 109, 108,107 and 106 conidia/ml for the three fungi B. bassiana R444 (Bb-Protec®) (F = 5.394, P < 0.0001), B. bassiana ATCC (Naturalis®) (F = 5.830, P < 0.0001) et L. muscarium (Mycotal®) (F = 3.828, P = 0.005). The corrected mortality rates following the application of B. bassiana ATCC obtained after 7 days were the highest at 29.78, 42.55, 53.19, 61.70 and 80.85%, respectively for the concentration of 106, 107, 108, 109 and 1010conidia/ml.

Corrected mortality of second instar Tuta absoluta larvae by the first method with Beauveria bassiana ATCC, B. bassiana R444 and Lecanicillium muscarium
The median lethal concentration values (LC50) after 7 days of application were significantly different at the different fungi. The lowest LC50 value (2.63 × 107 conidia/ml) was observed in B. bassiana ATCC, while the highest LC50 value (3.33 × 109 conidia/ml) was observed in L. muscarium (Table 3). For LC90 estimated for L. muscarium, B. bassiana R444 and B. bassiana ATCC had concentrations of 2.52 × 1016, 2.64 × 1014 and 4.02 × 1011conidia/ml, respectively (Table 3).
Pathogenicity test on Tuta absoluta larvae: the way 2
Mortality rates of T. absoluta second-instar larvae by the second method, after treatment with EPF, differed significantly with increasing the concentration at the end of the experiment period; B. bassiana R444 (Bb-Protec®) (F = 3.602, P = 0.008), B. bassiana ATCC (Naturalis®) (F = 3.278, P = 0.012) et L. muscarium (Mycotal®) (F = 4.816, P = 0.001). The highest corrected mortality rate for T. absoluta (86.81%) was caused by B. bassiana ATCC. L. muscarium EPF was found to be the least virulent with a corrected mortality rate of 46.24% (Fig. 3).

Corrected mortality of 2nd instar Tuta absoluta larvae from by the second method with Beauveria bassiana ATCC, B. bassiana R444 and Lecanicillium muscarium
The values of the LC50 and LC90 of B. bassiana ATCC, B. bassiana R444 and L. muscarium were shown in (Table 4). The results showed that B. bassiana ATCC was the most virulent EPF on second instar of T. absoluta at the concentrations of 4.76 × 106 and 5.84 × 109 spores/ml to kill 50 and 90% of the population, respectively.
T. absoluta larvae infected with B. bassiana showed color changes to yellowish brown (Fig. 4A). Fungal hyphae began to grow from the edges of the larvae's body. The hyphae then covered most of the larvae's body (Fig. 4B).

Effect of Beauveria bassiana ATCC 74,040 against Tuta absoluta larvae. A: dead larva after 4 days of treatment; B: Development of mushroom mycelium on the body of the larva
Pathogenicity on pupation and adult emergence of Tuta absoluta
The three commercial EPF formulations affected pupation of T. absoluta compared to the untreated control, but non-significant differences for B. bassiana R444 and L. muscarium (F = 2.51, P = 0.095) were found. On the other hand, for B. bassiana ATCC, there was a significant difference than the control. In fact, only 82% of fourth-instar larvae exposed to soil treated with this fungus developed to the pupal stage (P = 0.021). Adult emergence was 69.16, 60.15 and 50.25% for fungal treatments with L. muscarium, B. bassiana R444 and B. bassiana ATCC, respectively (Fig. 5). These fungi significantly reduced the emergence of T. absoluta adults than the control (F = 2.51; P = 0.095).

Effects of commercial entomopathogenic fungi (EPF) preparations on pupation and adult emergence of Tuta absoluta. The means followed by the same letter are not significantly different by the Tukey test (HSD) (p < 0.05)
Discussion
The EPF used in this study showed pathogenicity to eggs and second instar larvae of T. absoluta in all bioassays performed. Corrected mortality rates increased by increasing concentration and over time. Various studies also showed the efficacy of EPFs against T. absoluta eggs. Rodríguez et al. (2006) showed that B. bassiana Qu-B912 and M. anisopliae Qu-M558 had pathogenicity towards T. absoluta eggs with mortality percentages of around 60 to 80%, respectively. Similarly, they showed that egg mortality rates were significantly high at the highest spore concentrations for M. anisopliae, B. bassiana (Abdel-Baky et al. 2021) and Verticillium lecanii (Abdel-Raheem et al. 2020). While M. anisopliae isolates tURPE-6 and URPE-19 were more pathogenic (Pires et al. 2010). On the other hand, other experiments indicated that B. bassiana showed superiority over M. anisopliae in its effect on T. absoluta eggs control under laboratory conditions (Abdel-Baky et al. 2021). Thus, one study suggested that B. bassiana was only effective at the egg stage (Inanli et al. 2012). Other results also indicated that B. bassiana was particularly effective against T. absoluta eggs, causing more than 79.7% mortality (Bayram 2019).
Several studies have been conducted on the pathogenicity of B. bassiana against T. absoluta larvae, and they have shown significant efficacy similar to this study. B. bassiana was virulent against 2nd or 3rd larval instars of T. absoluta by application of conidia to the leaf surface (Silva et al. 2020). This fungus exhibited epiphytic and endophytic activity against T. absoluta. Investigation of the efficacy of a commercial mycoinsecticides based on B. bassiana against all stages of T. absoluta showed that the corrected mortality reached 30–50% (Klieber and Reineke 2016). B. bassiana and M. anisopliae were virulent against T. absoluta larvae under laboratory and greenhouse conditions (Tadele and Emana 2017). Ndereyimana et al. (2019) reported that the two formulations of the commercial EPFs Metatech® WP (M. anisopliae Strain FCM Ar 23B3) and Beauvitech® WP (B. bassiana, Strain J25) had a high level of virulence against T. absoluta larvae under laboratory conditions. The larvicidal effect was also evaluated in the laboratory and in the greenhouse by spraying two types of endophytic fungi. Clonostachys spp. and B. bassiana showed remarkable efficacy, variable depending on the strain tested (Mahmoud et al. 2021). In addition, the application of a bioinsecticide based on M. anisopliae to the 2nd instar larvae of T. absoluta by spraying was effective in reducing the population of the pest by 87% mortality on the 7th day of application (Erol et al. 2021). Four virulent isolates of B. bassiana MN598666, MK046654, MK046652 and Purpureocillium lilacinum (thom) Samson (Ascomycota: Hypocreales) MK046655 indicated promising potentials in regulating T. absoluta (Hammad et al. 2021). Aynalem et al. (2021) showed the efficacy of 27 strains of Beauveria with a mortality rate of 65.7 to 95.7% and 68.3 to 95% against 2nd and 3rd larval instars of T. absoluta at the concentration of 1 × 107 spores/ml, 7 days after inoculation.
Other studies have investigated the importance of this fungus when combined with other products. The combination of Azadirachtin and B. bassiana resulted in reductions in T. absoluta infestation and fruit damage up to greenhouse tomato harvest (Jallow et al. 2019). As with eggs, the highest larval mortality was also at the highest concentration (Tadele and Emana 2017) and increased over time (Ndereyimana et al. 2019).
The biological test against 4th instar larvae placed on soil treated with these fungi showed significant efficacy against pupation and adult emergence. Thus, T. absoluta L4 larvae were exposed to EPF spores dipped in soil, and thus this fungus grew on the cuticle of the larvae. The development of spores in the body of the larvae prevented a small percentage of the larvae from pupating and prevented a large percentage of the pupae emerging to adults. Studies have shown that soil drench applications of B. bassiana and M. anisopliae effectively controlled T. absoluta nymphs and affected female fecundity (Erasmus et al. 2021). Pupation and emergence of adults were significantly reduced by different isolates of M. anisopliae (Akutse et al. 2020). In addition, a liquid formulation based on strains of the fungus M. anisopliae (MA-Prep) associated with irrigation could lead to high mortality of T. absoluta pupae (Contreras et al. 2014). In other study, Bali et al. (2022) indicated the efficiency of P. lilacinum against T. absoluta pupae.
Conclusion
The results showed that the EPF isolates B. bassiana and L. muscarium studied at different concentrations against T. absoluta under controlled conditions similar to that of geothermal greenhouses in southern Tunisia were effective, especially against the second-instar larvae of T. absoluta, eggs and the emergence of adults. Egg and larval mortality increased with the increase of concentration of fungal suspensions of the different fungi. The application of EPF during irrigation to the 4th instar larvae dropped into the soil to pupate can be an effective way to reduce the emergence of T. absoluta adults. Thus, EPF could play an important role, as a biological mean for controlling crop-associated T. absoluta in greenhouses as component in IPM programs.
Availability of data and materials
All data of the study have been presented in the manuscript, and high quality and grade materials were used in this study.
Abbreviations
- EPF:
-
Entomopathogenic fungi
- IPM:
-
Integrated pest management
- PDA:
-
Potato dextrose agar
- ANOVA:
-
Analysis of variance
- LC:
-
Lethal concentration
- CL:
-
Confidence limit
- P :
-
P-Value
- SE:
-
Standard error
- df:
-
Degrees of freedom
References
Abbott WS (1925) A method of computing the effectiveness of an insecticide. J Econ Entomol 18:265–267
Abdel-Baky NF, Alhewairini SS, Al-Azzazy MM, Qureshi MZ, Al-Deghairi MA, Hajjar J (2021) Efficacy of Metarhizium anisopliae and Beauveria bassiana against Tuta absoluta (Lepidoptera: Gelechiidae) eggs under laboratory conditions. Pak J Agri Sci 58:743–750. https://doi.org/10.21162/PAKJAS/21.52
Abdel-Raheem M, Youssif M, Helaly SM (2020) Use of Verticillium lecanii and Beauveria bassiana against Tomato leaf miner, Tuta absoluta (Meyrick) and Bemisia tabaci (Genn.) in Tomato Crop. Plant Arch 20:479–482
Akutse KS, Subramanian S, Khamis FM, Ekesi S, Mohamed SA (2020) Entomopathogenic fungus isolates for adult Tuta absoluta (Lepidoptera: Gelechiidae) management and their compatibility with Tuta pheromone. J Appl Entomol 144:777–787. https://doi.org/10.1111/jen.12812
Aynalem B, Muleta D, Venegas J, Assefa F (2021) Molecular phylogeny and pathogenicity of indigenous Beauveria bassiana against the tomato leafminer, Tuta absoluta Meyrick 1917 (Lepidoptera: Gelechiidae), in Ethiopia. J Gen Eng Biotechnol 19:1–15. https://doi.org/10.1186/s43141-021-00227-x
Bali GK, Singh SK, Maurya DK, Wani FJ, Pandit RS (2022) Morphological and molecular identification of the entomopathogenic fungus Purpureocillium lilacinum and its virulence against Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) larvae and pupae. Egypt J Biol Pest Control 32:1–10. https://doi.org/10.1186/s41938-022-00582-y
Bayram Y (2019) Efficacy of two entomopathogenic fungi and their combination with summer oil against tomato moth, Tuta absoluta. Fresenius Environ Bull 28:3435–3440
Cherif A, Attia-Barhoumi S, Mansour R, Zappalà L, Grissa-Lebdi K (2019) Elucidating key biological parameters of Tuta absoluta on different host plants and under various temperature and relative humidity regimes. Entomol General 39:1–7. https://doi.org/10.1127/entomologia/2019/0685
Contreras J, Mendoza J, Martínez-Aguirre M, García-Vidal L, Izquierdo J, Bielza P (2014) Efficacy of enthomopathogenic fungus Metarhizium anisopliae against Tuta absoluta (Lepidoptera: Gelechiidae). J Econ Entomol 107:121–124. https://doi.org/10.1603/EC13404
Desneux N, Wajnberg E, Wyckhuys KA, Burgio G, Arpaia S, Narváez-Vasquez CA, González-Cabrera J, Ruescas DC, Tabone E, Frandon J (2010) Biological invasion of European tomato crops by Tuta absoluta: ecology, geographic expansion and prospects for biological control. J Pest Sci 83:197–215. https://doi.org/10.1007/s10340-010-0321-6
Desneux N, Luna MG, Guillemaud T, Urbaneja A (2011) The invasive South American tomato pinworm, Tuta absoluta, continues to spread in Afro-Eurasia and beyond: the new threat to tomato world production. J Pest Sci 84:403–408. https://doi.org/10.1007/s10340-011-0398-6
Erasmus R, van den Berg J, du Plessis H (2021) Susceptibility of Tuta absoluta (Lepidoptera: Gelechiidae) pupae to soil applied entomopathogenic fungal biopesticides. InSects 12:515. https://doi.org/10.3390/insects12060515
Erol AB, Erdoğan O, Karaca İ (2021) Effects of some bioinsecticides on the tomato leaf miner, Tuta absoluta (Meyrick, 1917) (Lepidoptera: Gelechiidae). Egypt J Biol Pest Control 31:1–4. https://doi.org/10.1186/s41938-020-00346-6
Finney DJ (1971) Probit analysis. Cambridge University Press, Londres
Guedes R, Picanço M (2012) The tomato borer Tuta absoluta in South America: pest status, management and insecticide resistance. EPPO Bull 42:211–216. https://doi.org/10.1111/epp.2557
Hammad AMA, Bashir HAAA, Abdelbagi AO, Ishag AESA, Ali MMY, Bashir MO, Hur JH, Laing MD (2021) Efficacy of indigenous entomopathogenic fungi for the control of the tomato leafminer Tuta absoluta (Meyrick) in Sudan. Int J Trop Insect Sci. https://doi.org/10.1007/s42690-021-00663-9
Han P, Bayram Y, Shaltiel-Harpaz L, Sohrabi F, Saji A, Esenali UT, Jalilov A, Ali A, Shashank PR, Ismoilov K (2019) Tuta absoluta continues to disperse in Asia: damage, ongoing management and future challenges. J Pest Sci 92:1317–1327. https://doi.org/10.1007/s10340-018-1062-1
Inanli C, Yoldaș Z, Birgücü A (2012) Effects of entomopathogenic fungi, Beauveria bassiana (Bals.) and Metarhizium anisopliae (Metsch.) on larvae and egg stages of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Ege Üniv Ziraat Fakültesi Derg 49:239–242
Jallow MF, Dahab AA, Albaho MS, Devi VY (2019) Efficacy of some biorational insecticides against Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) under laboratory and greenhouse conditions in Kuwait. J Appl Entomol 143:187–195. https://doi.org/10.1111/jen.12588
Klieber J, Reineke A (2016) The entomopathogen Beauveria bassiana has epiphytic and endophytic activity against the tomato leaf miner Tuta absoluta. J Appl Entomol 140:580–589. https://doi.org/10.1111/jen.12287
Mahmoud FM, Bendebbah R, Benssaci B, Toudji F, Tafifet L, Krimi Z (2021) Entomopathogenic efficacy of the endophytic fungi: Clonostachys sp. and Beauveria bassiana on Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) larvae under laboratory and greenhouse conditions. Egypt J Biol Pest Control 31:1–6. https://doi.org/10.1186/s41938-021-00392-8
Ndereyimana A, Nyalala S, Murerwa P, Gaidashova S (2019) Pathogenicity of some commercial formulations of entomopathogenic fungi on the tomato leaf miner, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Egypt J Biol Pest Control 29:1–5. https://doi.org/10.1186/s41938-019-0184-y
Pires LM, Marques EJ, de Oliveira JV, Alves SB (2010) Selection of entomopathogenic fungi isolates for the control of Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) and their compatibility with some insecticides used in tomato crops. Neotrop Entomol 39:977–984. https://doi.org/10.1590/S1519-566X2010000600020
Roditakis E, Skarmoutsou C, Staurakaki M (2013) Toxicity of insecticides to populations of tomato borer Tuta absoluta (Meyrick) from Greece. Pest Manag Sci 69:834–840. https://doi.org/10.1002/ps.3442
Roditakis E, Vasakis E, Garcia-Vidal L, del Rosario M-A, Rison JL, Haxaire-Lutun MO, Nauen R, Tsagkarakou A, Bielza P (2018) A four-year survey on insecticide resistance and likelihood of chemical control failure for tomato leaf miner Tuta absoluta in the European/Asian region. J Pest Sci 91:421–435. https://doi.org/10.1007/s10340-017-0900-x
Rodríguez MS, Gerding MP, France AI (2006) Entomopathogenic fungi isolates selection for egg control of tomato moth Tuta absoluta Meyrick (Lepidoptera: Gelechiidae) eggs. Agric Téc 66:151–158
Silva ACL, Silva GA, Abib PHN, Carolino AT, Samuels RI (2020) Endophytic colonization of tomato plants by the entomopathogenic fungus Beauveria bassiana for controlling the South American tomato pinworm, Tuta absoluta. CABI Agric Biosci 1:1–9. https://doi.org/10.1186/s43170-020-00002-x
Tadele S, Emana G (2017) Entomopathogenic effect of Beauveria bassiana (Bals.) and Metarrhizium anisopliae (Metschn.) on Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) larvae under laboratory and glasshouse conditions in Ethiopia. J Plant Pathol Microbiol 8:2. https://doi.org/10.4172/2157-7471.1000411
Tropea Garzia G, Siscaro G, Biondi A, Zappalà L (2012) Tuta absoluta, a South American pest of tomato now in the EPPO region: biology, distribution and damage. EPPO Bull 42:205–210. https://doi.org/10.1111/epp.2556
Urbaneja A, Vercher R, Navarro-Llopis V, Porcuna Coto JL, García-Marí F (2007) The tomato leaf miner, Tuta absoluta. La revista profesional de sanidad vegetal, Phytoma España, pp 16–23
Urbaneja A, González-Cabrera J, Arno J, Gabarra R (2012) Prospects for the biological control of Tuta absoluta in tomatoes of the Mediterranean basin. Pest Manag Sci 68:1215–1222. https://doi.org/10.1002/ps.3344
Youssef NA (2015) Efficacy of the entomopathogenic nematodes and fungi for controlling the tomato leaf miner, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Arab Univ J Agric Sci 23:591–598
Acknowledgements
The authors are especially grateful to “Zina fresh” company for providing all the resources needed for this study.
Funding
This research was funded by “Zina fresh” company, the Technical Center for Protected and Geothermal Crops Gabes and by the Arid Land Institute Tunisia.
Author information
Authors and Affiliations
Contributions
Conceived and designed the experiments: SC and MSB. Performed the experiments: SC and BHA Analyzed the data: SC. Wrote the paper: SC, KLG, and MSB. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not Applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Chouikhi, S., Assadi, B.H., Lebdi, K. et al. Efficacy of the entomopathogenic fungi Beauveria bassiana and Lecanicillium muscarium in the control of the tomato leaf miner, Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Egypt J Biol Pest Control 32, 139 (2022). https://doi.org/10.1186/s41938-022-00640-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41938-022-00640-5