
Abstract
Background
Fluoride may induce oxidative stress and apoptosis. It may also lead to neurobehavioural defects including neuromuscular damage. The present study aimed to explore the effects of sub lethal concentrations of sodium fluoride (NaF) on the lifespan and climbing ability of Drosophila melanogaster. In total, 0.6 mg/L and 0.8 mg/L of NaF were selected as sublethal concentrations of NaF for the study. Lifespan was measured and climbing activity assay was performed.
Results
The study showed significant decrease in lifespan of flies treated with fluoride. With increasing age, significant reduction in climbing activity was observed in flies treated with sodium fluoride as compared to normal (control) flies. Flies treated with tulsi (Ocimum sanctum) and NaF showed increase in lifespan and climbing activity as compared to those treated with NaF only. Lipid peroxidation assay showed significant increase in malondialdehyde (MDA) values in the flies treated with NaF as compared to control. The MDA values decreased significantly in flies treated with tulsi mixed with NaF.
Conclusions
The results indicate that exposure to sub lethal concentration of NaF may cause oxidative stress and affect the lifespan and climbing activity of D. melanogaster. Tulsi extract may help in reducing the impact of oxidative stress and toxicity caused by NaF.
Similar content being viewed by others

Background
Fluoride may enter the human body through (i) drinking water, (ii) food and food products (e.g. contaminated with pesticides) and (iii) industrial emission of fluoride dust and fumes (Susheela, 2013). Excessive ingestion of fluoride results in development of dental, skeletal and non-skeletal fluorosis in humans. Acute pesticide poisoning during childhood may lead to neurobehavioural deficits (Berger, Friedman, Jaffar, Kofman, & Massarwa, 2006; Beseler, Bouchard, & London, 2012). Toxic effects of fluoride have been documented in other animals also. Chronic fluoride exposure can alter kidney structure, renal function and induce apoptosis in pigs (Zhan, Wang, Xu, & Li, 2006). It can cause severe health problems in rat, mice and fish due to oxidative stress, DNA damage and apoptosis (He & Chen, 2006; Mukhopadhyay & Chattopadhyay, 2014).
Fluoride is able to induce oxidative stress and apoptosis in both intrinsic and extrinsic pathways in the mammalian cells (Agalakova, Petrova, & Gusev, 2019; Lu et al., 2017; Ribeiro et al., 2017). NaF-induced oxidative damage inhibits enzymes involved in energy production, membrane bound ion transport and neurotransmission (Vani & Reddy, 2000). Pesticides containing fluoride have negative impacts on the biology of non-target organisms (Agalakova & Gusev, 2012). D. melanogaster (commonly called fruit fly) is not considered as a pest, as it grows and multiplies on rotten fruits. Instead, it is considered as a non-target organism and is used as a model for toxicity evaluation of chemicals (Rand, Montgomery, Prince, & Vorojeikina, 2014). Drosophila melanogaster is an acceptable model for understanding the human condition under stress of toxicants because of the abundance of highly conserved genes and pathways controlling development, stress response and xenobiotic metabolism across these two divergent species (Mackay & Anholt, 2006; Misra, Horner, Lam, & Thummel, 2011; Sykiotis & Bohmann, 2010). A number of studies have been conducted on Drosophila model using priority environmental contaminants and toxicants including mercury (Rand, Dao, & Clason, 2009), lead (Hirsch et al., 2003), arsenic (Ortiz, Opoka, Kane, & Cartwright, 2009), fluoride (Dutta, Rajak, Khatun, & Roy, 2017; Khatun, Mandi, Rajak, & Roy, 2018), manganese (Bonilla et al., 2012), ethanol (Guarnieri & Heberlein, 2003), nanoparticles (Posgai et al., 2011), pesticides (Gupta et al., 2007; Das, Podder, & Roy, 2010; Chmiel, Daisley, Burton, & Reid, 2019), nanopesticides (Demir, 2020) and solvents (Wasserkort & Koller, 1997).
NaF can cause genotoxic alterations and induce melanotic tumours in adult D. melanogaster (Herskowitz & Norton, 1963). Similarly, sublethal exposure of fluoride has been found to cause physiological and reproductive changes in D. melanogaster (Dutta et al., 2017; Singh, Chowdhary, Shah, & Yasmin, 2020).
The basic biological, physiological, and neurological properties are conserved between mammals and D. melanogaster, and nearly 75% of human disease-causing genes are believed to have a functional homologue in the fly (Lloyd & Taylor, 2010; Reiter, Potocki, Chien, Gribskov, & Bier, 2001). Therefore, D. melanogaster is used as a model system for investigating the roots of human diseases such as neurological diseases, including neuromuscular disease.
O. sanctum (Linn) commonly called ‘tulsi’ belongs to family Labiatae. Several medicinal properties have been attributed to the plant not only in Ayurveda and Siddha but also in Greek, Roman and Unani system of medicines. O. sanctum has been reported to possess antimicrobial, anti-stress, antidiabetic, hepatoprotective, anti-inflammatory, anti-carcinogenic, immunomodulatory, radioprotective, neuroprotective and cardioprotective properties. The leaves of O. sanctum contain alkaloids, flavonoids, glycosides, saponins, tannins, ascorbic acid and carotene (Mondal, Mirdha, & Mahapatra, 2009).
This paper reports about the effects of fluoride toxicity on the lifespan and climbing ability of D. melanogaster. Impaired climbing ability may be linked to premature ageing caused by fluoride due to oxidative stress. This is because fluoride inhibits bioenergetic reactions, in particular oxidative phosphorylation, reducing physical activity of muscles (Machoy-Mokrzyńska, 2004). An attempt was made to study the effect of O. sanctum on the lifespan and climbing activity of D. melanogaster exposed to NaF.
Methods
Native D. melanogaster was cultured in the laboratory at 25 °C in standard cornmeal medium. The standard cornmeal medium consisted of maize powder, sucrose, dextrose, yeast extract and agar. Single line culture (stock) of D. melanogaster was maintained to obtain flies of the same age and strain. For determining the sublethal concentration of NaF for D. melanogaster, 0.6 mg/L, 0.8 mg/L and 1 mg/L of NaF were tested following Singh et al. (2020) and Mishra, Kumari, Ranjan and Yasmin (2020). Four sets of cornmeal media were prepared. Each set consisted of three bottles. The media were changed after every 4 days.
The four sets of bottles were as follows:
-
1)
Control set—fruit flies were cultured in standard cornmeal media.
-
2)
NaF-treated sets (three sets)—three sets of standard cornmeal media were prepared and 1.0 mg/L, 0.8 mg/L and 0.6 mg/L of NaF were added respectively into these three sets.
Five flies from the stock were transferred into each set and monitored.
Preparation of O. sanctum extract
O. sanctum (tulsi) leaf extract was prepared following Mitra et al. (2014). The tulsi leaves were dried in a hot air oven and powdered using mortar and pestle. The dried O. sanctum leaf dusts were soaked overnight in distilled water (15 g leaf dust per 100 ml distilled water) and filtered through a fine muslin cloth. The filtrate was centrifuged at 5000 rpm for 10 min. The supernatant thus obtained, was filtered again using a fine muslin cloth and the filtrate was collected in sterile polypropylene tubes and frozen at 20 °C.
Published report on clinical trials conducted on humans till date suggests that tulsi is a safe herbal intervention. Tulsi dosage and frequency in such studies varied from 300 mg to 3000 mg given as 1–3 times per day as tulsi leaf aqueous extract to human subjects (Jamshidi & Cohen, 2017). In the present study, 10% v/v tulsi leaf extract (TLE) was taken which was roughly 50% of the above mentioned dosage. However, higher dose of TLE may also be tried.
For performing the experiments, five sets of cornmeal media were prepared. Each set had three bottles of cornmeal media.
-
1)
Control set with standard cornmeal media
-
2)
NaF (0.6 mg/L) treated cornmeal media
-
3)
NaF (0.8 mg/L) treated cornmeal media
-
4)
O. sanctum cornmeal media (with 10% v/v tulsi leaf extract in standard cornmeal media)
-
5)
O. sanctum + NaF (0.8 mg/L) containing media (with 10% v/v tulsi leaf extract and 0.8 mg/L of NaF in standard cornmeal media)
For studying the lifespan of native D. melanogaster in the different media mentioned above, newly ecloded flies were collected from the stock and raised in the respective media at 25 °C. Twenty flies were placed into each bottle and were transferred to bottles with fresh media after every 4 days. The numbers of dead flies were counted every day.
Climbing activity assay was performed following Manjila and Hasan (2018) in a glass cylinder of 50 ml. Batches of 20 flies from each experimental set were used to perform climbing activity assay. Timer of 10 s was set and the number of flies crossing the 50 ml border was counted. Each assay was repeated thrice and average climbing ability of each batch of flies was calculated. Climbing activity of each batch of flies was monitored at interval of 3 days.
Lipid peroxidation assay was performed following Ohkawa, Ohishi, and Yagi (1979) on third generation flies (flies exposed to NaF for three generations). In total, 0.3 g of the flies was taken and homogenised by adding 1 ml of 0.1% trichloroacetic acid (TCA) in a glass homogeniser. The homogenate was centrifuged at 5000 rpm for 15 min at room temperature. One millilitre of the supernatant was transferred into a clean and dry test tube and 2 ml of freshly prepared 0.5% thiobarbituric acid (TBA) in 20% TCA was added into it. This sample was incubated at 90 °C for 30 min and subsequently cooled at room temperature. Absorbance was measured by dual beam spectrophotometer at 532 and 600 nm. All the readings were taken in triplicates.
MDA level was calculated by following formula:
Where,
OD = optical density
TV = total volume of the sample
dw = dry weight of sample
Statistical analysis was performed using ANOVA and p<0.05 was considered as significant. One-way ANOVA was used to analyse climbing assay data and two-way ANOVA was used to analyse lifespan data.
Results
Flies cultured in media with 1.0 mg/L NaF survived for 1 week only. Some eggs but no larvae were found in these media after 1 week. Flies cultured in media with 0.6 mg/L and 0.8 mg/L NaF continued to survive and produce eggs and larvae. So, 0.6 mg/L and 0.8 mg/L of NaF were considered as sub lethal concentrations for the present study. Singh et al. (2020) and Mishra et al. (2020) also reported 0.8 mg/L as the sublethal concentration of NaF for D. melanogaster and Zaprionus indianus respectively.
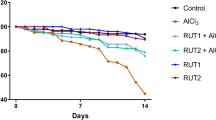
Survival of D. melanogaster was better in control media (Table 1) as compared to that in the media containing 0.6 mg/L NaF (Table 2), where only a single fly survived for 25 days. In the media containing 0.8 mg/L NaF (Table 3), a single fly survived for 22 days. Survival of flies in the media containing O. sanctum extract (Table 4) was similar to those in the control media whereas survival of flies in media containing NaF + O. sanctum extract was found to be better than those in media with NaF (Table 5). Fifty percent of the flies was dead by the end of 4th week in control media (Table 1), by the 2nd week in media with NaF (Tables 2 and 3), by the 5th week in media with tulsi extract (Table 4) and by the end of 3rd week in media with tulsi extract+0.8 mg/L NaF (Table 5). Lifespan of D. melanogaster was found to be ~75 days in control set, ~76 days in media with tulsi extract, ~40 days in media with 0.6 mg/L NaF, ~37 days in media with 0.8 mg/L NaF and ~57 days in media with tulsi extract+0.8 mg/L NaF. A comparative picture of survival of flies in different media is shown in Fig. 1a and b.

Mean life span of Drosophila melanogaster in different media
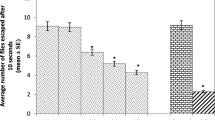
Climbing activity of D. melanogaster was maximum after 3 days of eclosion in control media (Fig. 2). On the other hand, the climbing activity of flies reared in media with 0.6 mg/L and 0.8 mg/L NaF was suppressed markedly after 3 days of eclosion. A progressive decline in climbing activity was seen in the flies exposed to NaF after 6, 9 and 12 days of eclosion (Figs. 3, 4 and 5). These flies could not show any climbing activity after 15 days of eclosion (Fig. 6). D. melanogaster cultured in media with tulsi extract showed improved climbing activity after 15 days of eclosion (Figs. 2, 3, 4, 5 and 6).

Climbing ability of Drosophila melanogaster after 03 days of eclosion. *, significant difference between control vs NaF; **, significant difference between tulsi vs NaF; ***, significant difference between tulsi + F in medium vs NaF

Climbing ability of Drosophila melanogaster after 06 days of eclosion. *, significant difference between control vs NaF; **, significant difference between tulsi vs NaF

Climbing ability of Drosophila melanogaster after 09 days of eclosion. *, significant difference between control vs NaF and tulsi; **, significant difference between tulsi vs NaF; ***, significant difference between tulsi+0.8 mg/L NaF vs NaF

Climbing ability of Drosophila melanogaster after 12 days of eclosion. *, significant difference between control vs NaF and tulsi; **, significant difference between tulsi vs NaF; ***, significant difference between tulsi+0.8 mg/L NaF vs NaF

Climbing ability of Drosophila melanogaster after 15 days of eclosion. *, significant difference between control vs tulsi
Lipid peroxidation (LPO) assay revealed altered malonyl dialdehyde (MDA) production in the tissues of flies after exposure to sub lethal concentration (0.8 mg/L) of NaF as compared to control and to those treated with tulsi extract+ 0.8 mg/L NaF (F(2,6) =11.95, P<0.05) (Fig. 7).

Lipid peroxidation (MDA) measured in D. melanogaster cultured in different media. *, significant difference between MDA values of flies cultured in Control vs media with NaF; **, significant difference between MDA values of flies cultured in media with NaF vs media with tulsi extract+0.8 mg/L NaF
Discussion
The significant reduction in lifespan of fluoride treated flies was similar to the findings of Khatun et al. (2018), where NaF exposure to D. melanogaster in the parental generation led to an increase in adult mortality. Significant reduction in the climbing activity was also seen in the flies treated with NaF. Similarly, Khatun et al. (2018) and Sarkar, Roy, and Roy (2018) also observed alteration in climbing behaviour in flies exposed to sub lethal concentrations of fluoride. Fluoride is thought to inhibit the activity of antioxidant enzymes, such as superoxide dismutase, glutathione peroxidase and catalase. Depletion of glutathione results in excessive production of reactive oxygen species at the mitochondrial level, leading to the damage of cellular components. Abolaji et al. (2019) treated D. melanogaster with NaF and found altered levels of oxidative stress markers (Glutathione-S-transferase (GST), catalase and acetylcholinesterase (AchE) activities, total thiol (T-SH), nitrites/nitrates and hydrogen peroxide (H2O2) levels). These parameters could be balanced by resveratrol (a natural polyphenol with antioxidant and anti-inflammatory properties).
In the present study, when tulsi extract was mixed in the media with NaF, the flies survived and maintained their climbing activity in comparison to the flies cultured in media with NaF only. Similar results were found by Siddique, Faisal, Naz, Jyoti, and Rahul (2014), where the flies exposed to various doses of O. sanctum extract showed a dose-dependent significant delay in the loss of climbing ability.
Higher MDA value in the flies treated with NaF indicated high oxidative stress which is similar to the results obtained by Patel and Chinoy (1998); Wang et al. (2004) and Dutta et al. (2017). Treatment with tulsi extract possibly reduced the oxidative stress. Siddique et al. (2014) also found that treatment with tulsi extract caused reduction in oxidative stress in the brain of Parkinson’s diseased model flies.
Oxidative stress plays a major role in ageing, and is associated with several neurodegenerative diseases. O. sanctum (tulsi) leaf extract possesses antioxidative properties (Mitra et al., 2014). O. sanctum leaves are rich in polyphenolic flavonoids which act as antioxidants (Hakkim, Gowri Shankar, & Girija, 2007) and are helpful in preventing lipid peroxidation (Geetha & Vasudevan, 2004). Aqueous extract of the O. sanctum leaves may function simply by quenching the free radicals generated during oxidative stress or may improve the antioxidant enzyme status of the tissue in the face of the oxidative stress (Mitra et al., 2014). Tulsi has been found to be a chief source of many biologically active compounds like ursolic acid, eugenol, rosmarinic acid, linalool, carvacrol and β caryophyllene and these compounds play a significant role in the treatment and prevention of many diseases (Almatroodi, Alsahli, Almatroudi, & Rahmani, 2020). The antioxidant nature of O. sanctum leaf extract might have reduced the oxidative stress caused by NaF in D. melanogaster used in this study. Shivananjappa and Joshi (2012) also found that aqueous extract of tulsi had putative potency to enhance the endogenous antioxidant defences in human hepatocyte cell line (HepG2) which can potentially effect faster dissipation of ROS. Free radical scavenging activity is a chief mechanism through which Ocimum sanctum products protect against cellular damage. Its role in free radicals scavenging property has confirmed its strong antioxidant activity and free radicals scavenging property (Ganasoundari et al., 1998; Keshari, Srivastava, Verma, & Srivastava, 2016).
In the present study, tulsi extract (alone) was not found to significantly increase the lifespan of flies as compared to control, but lifespan of flies treated with fluoride + tulsi significantly increased as compared to the flies treated with fluoride. The results of this study can be considered preliminary and further investigations are required to prove the worth of tulsi in increasing the lifespan. Research has shown that tulsi reduces stress, enhances stamina, relieves inflammation, lowers cholesterol, eliminates toxins, protects against radiation, prevents gastric ulcers, lowers fevers, improves digestion and provides a rich supply of antioxidants and other nutrients. The nutritional analysis of Ocimum sanctum has shown high level of ascorbic acid, N, P, K, total phenol, carbohydrates and proteins in their leaves, which may be very good for health. These properties may help to enhance the lifespan (Patel, 2020). Further, tulsi has been found to mediate a significant reduction in tumour cell size and an increase in lifespan of mice having sarcoma-180 solid tumours (Nakamura et al., 2004).
Fluoridated insecticides may be helpful in targeting the pests (Metcalf, 2015), but the toxic effects of fluoride on non-target animals should not be neglected (Dhar & Bhatnagar, 2009; Sauerheber, 2013). The results of the present study indicate that aqueous tulsi leaf extract acts as antioxidant by possibly scavenging the oxygen free radical and other reactive oxygen intermediates. Thus, O. sanctum has the potential to reduce fluoride toxicity in D. melanogaster. The study suggests that O. sanctum (tulsi) may be of future therapeutic relevance particularly in the area where humans are chronically exposed to fluoride either occupationally or through food chain.
Conclusion
The present study concluded that exposure of D. melanogaster to sub lethal concentrations of NaF caused oxidative stress induced damage in its body leading to reduced lifespan and climbing activity. It was also concluded that O. sanctum extract may reduce oxidative stress and fluoride toxicity. Therefore, O. sanctum can be of therapeutic relevance.
Availability of data and materials
The datasets used and analysed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- NaF:
-
Sodium fluoride
- TLE:
-
Tulsi leaf extract
- MDA:
-
Malondialdehyde
References
Abolaji, A. O., Ajala, V. O., Adigun, J. O., Adedara, I. A., Kinyi, H. W., & Farombi, E. O. (2019). Protective role of resveratrol, a natural polyphenol, in sodium fluoride-induced toxicity in Drosophila melanogaster. Experimental Biology and Medicine, 244(18), 1688–1694. https://doi.org/10.1177/1535370219890334.
Agalakova, N. A., Petrova, T. I., & Gusev, G. P. (2019). Activation of Fas receptors, caspase-8 and caspase-3 by fluoride ions in rat erythrocytes in vitro. Journal of Evolutionary Biochemistry and Physiology, 55, 97–103. https://doi.org/10.1134/S0022093019020029.
Agalakova, N. I., & Gusev, G. P. (2012). Molecular mechanism of cytotoxicity and apoptosis induced by inorganic fluoride. International Scholarly Research Network Cell Biology, 2012, 1–16. https://doi.org/10.5402/2012/403835 Article ID 403835.
Almatroodi, S. A., Alsahli, M. A., Almatroudi, A., & Rahmani, A. H. (2020). Ocimum sanctum: role in diseases management through modulating various biological activity. Pharmacognosy Journal, 12(5), 1198–1205.
Berger, A., Friedman, A., Jaffar, A. A., Kofman, O., & Massarwa, A. (2006). Motor inhibition and learning impairments in school-aged children following exposure to organophosphate pesticides in infancy. Journal of Pediatric Research, 60, 88–92.
Beseler, C., Bouchard, M. F., & London, L. (2012). Neurobehavioral and neurodevelopmental effects of pesticide exposures. Neurotoxicology, 33(4), 887–896. https://doi.org/10.1016/j.neuro.2012.01.004.
Bonilla, E., Contreras, R., Medina-Leendertz, S., Mora, M., Villalobos, V., & Bravo, Y. (2012). Minocycline increases the life span and motor activity and decreases lipid peroxidation in manganese treated Drosophila melanogaster. Toxicology, 294, 50–53.
Chmiel, J. A., Daisley, B. A., Burton, J. P., & Reid, G. (2019). Deleterious effects of neonicotinoid pesticides on Drosophila melanogaster immune pathways. mBio, 10, e01395–e01319.
Das, S. K., Podder, S., & Roy, S. (2010). Effects of fungicide, Thiovit Jet on several life history traits of Drosophila melanogaster (Diptera:Drosophilidae). Journal of Applied Biological Sciences, 4(3), 31–36.
Demir, E. (2020). Drosophila as a model for assessing nanopesticide toxicity. Nanotoxicology, 14(9), 1271–1279. https://doi.org/10.1080/17435390.2020.1815886.
Dhar, V. N., & Bhatnagar, M. (2009). Physiology and toxicity of fluoride. Indian Journal of Dental Research, 20(3), 350–355. https://doi.org/10.4103/0970-9290.57379.
Dutta, M., Rajak, P., Khatun, S., & Roy, S. (2017). Toxicity assessment of sodium fluoride in Drosophila melanogaster after chronic sub-lethal exposure. Chemosphere, 166, 255–266. https://doi.org/10.1016/j.chemosphere.2016.09.112.
Ganasoundari, A., Umadevi, P., & Rao, B. S. (1998). Enhancement of bone marrow radiation protection and reduction in WR-2721 toxicity by Ocimum sanctum. Mutation Research, 373, 271–275.
Geetha, R. K., & Vasudevan, D. M. (2004). Inhibition of lipid peroxidation by botanical extracts of Ocimum sanctum: In vivo and in vitro studies. Life Sciences, 76(1), 21–28. https://doi.org/10.1016/j.lfs.2004.05.036.
Guarnieri, D. J., & Heberlein, U. (2003). Drosophila melanogaster, a genetic model system for alcohol research. International Review of Neurobiology, 54, 199–228. https://doi.org/10.1016/S0074-7742(03)54006-5.
Gupta, S. C., Siddique, H. R., Mathur, N., Mishra, R. K., Mitra, K., Saxena, D. K., & Chowdhuri, D. K. (2007). Adverse effect of organophosphate compounds, dichlorvos and chlorpyrifos in the reproductive tissues of transgenic Drosophila melanogaster: 70kDa heat shock protein as a marker of cellular damage. Toxicology, 238(1), 1–14. https://doi.org/10.1016/j.tox.2007.05.017.
Hakkim, F. L., Gowri Shankar, C. G., & Girija, S. (2007). Chemical composition and antioxidant property of holy basil (Ocimum sanctum L.) leaves, stems, and inflorescence and their in vitro callus cultures. Journal of Agricultural and Food Chemistry, 55(22), 109–9117. https://doi.org/10.1021/jf071509h.
He, L. F., & Chen, J. G. (2006). DNA damage, apoptosis and cell cycle changes induced by fluoride in rat oral mucosal cells and hepatocytes. World Journal of Gastroenterology, 12, 1144–1148.
Herskowitz, I. H., & Norton, I. L. (1963). Increased incidence of melanotic tumors in two strains of Drosophila melanogaster following treatment with sodium fluoride. Genetics., 48(2), 307–310. https://doi.org/10.1093/genetics/48.2.307.
Hirsch, H. V., Mercer, J., Sambaziotis, H., Huber, M., Stark, D. T., Torno-Morley, T., … Ruden, D. M. (2003). Behavioral effects of chronic exposure to low levels of lead in Drosophila melanogaster. Neurotoxicology, 24(3), 435–442. https://doi.org/10.1016/S0161-813X(03)00021-4.
Jamshidi, N., & Cohen, M. M. (2017). The clinical efficacy and safety of tulsi in humans: a systematic review of the literature. Evidence-Based Complementary and Alternative Medicine : eCAM, 2017, 9217567–9217513. https://doi.org/10.1155/2017/9217567.
Keshari, A. K., Srivastava, A., Verma, A. K., & Srivastava, R. (2016). Free radicals scavenging and protein protective property of Ocimum sanctum (L). British Journal of Pharmaceutical Research, 14(4), 1–10. https://doi.org/10.9734/BJPR/2016/31445.
Khatun, S., Mandi, M., Rajak, P., & Roy, S. (2018). Interplay of RDS and behavioural pattern in fluoride exposed Drosophila melanogaster. Chemosphere, 209, 220–231.
Lloyd, T. E., & Taylor, J. P. (2010). Flightless flies: Drosophila models of neuromuscular disease. Annals of the New York Academy of Sciences, 1184(1), e1–e20. https://doi.org/10.1111/j.1749-6632.2010.05432.x.
Lu, Y., Luo, Q., Cui, H., Deng, H., Kuang, P., Liu, H., … Zhao, L. (2017). Sodium fluoride causes oxidative stress and apoptosis in the mouse liver. Aging, 9(6), 1623–1639. https://doi.org/10.18632/aging.101257.
Machoy-Mokrzyńska, A. (2004). Fluorine as a factor in premature aging. Annales Acadamiae Medicae Stetinensis, 50(1), 9–13.
Mackay, T. F., & Anholt, R. R. (2006). Of flies and man: Drosophila as a model for human complex traits. Annual Review of Genomics and Human Genetics, 7(1), 339–367. https://doi.org/10.1146/annurev.genom.7.080505.115758.
Manjila, S. B., & Hasan, G. (2018). Flight and Climbing assay for assessing motor functions in Drosophila. Bio-protocol, 8(5), e2742.
Metcalf, R. L. (2015). Fluorine-containing insecticides. Florida Entomologist, 98(3), 987–988.
Mishra, D., Kumari, R., Ranjan, S., Yasmin, S. (2020). Effect of fluoride on the learning and memory ability of larvae of Zaprionus indianus. Journal of Basic and Applied Zoology, 81(1), 27. https://doi.org/10.1186/s41936-020-00166-y.
Misra, J. R., Horner, M. A., Lam, G., & Thummel, C. S. (2011). Transcriptional regulation of xenobiotic detoxification in Drosophila. Genes and Development, 25(17), 1796–1806. https://doi.org/10.1101/gad.17280911.
Mitra, E., Ghosh, D., Ghosh, A. K., Basu, A., Chattopadhayay, A., Pattari, S. K., … Bandyopadhyay, D. (2014). Aqueous tulsi leaf extract possess antioxidant properties and protects against cadmium induced oxidative stress in rat heart. International Journal of Pharmacy and Pharmaceutical Sciences, 6(1), 500–513.
Mondal, S., Mirdha, B. R., & Mahapatra, S. C. (2009). The science behind sacredness of tulsi (Ocimum sanctum Linn.) [J]. Indian Journal of Physiology and Pharmacology, 53(4), 291–306.
Mukhopadhyay, D., & Chattopadhyay, A. (2014). Induction of oxidative stress and related transcriptional effects of sodium fluoride in female zebrafish liver. Bulletin of Environmental Contamination and Toxicology, 93(1), 64–70. https://doi.org/10.1007/s00128-014-1271-0.
Nakamura, C. V., Ishida, K., Faccin, L. C., Filho, B. P. D., Cortez, D. A. G., Rozental, S., … Ueda-Nakamura, T. (2004). In vitro activity of essential oil from Ocimum gratissimum L. against four Candida species. Research in Microbiology, 155(7), 579–586. https://doi.org/10.1016/j.resmic.2004.04.004.
Ohkawa, H., Ohishi, N., & Yagi, K. (1979). Assay for lipid peroxidation in animal tissues by thiobarbituric acid reaction. Analytical Biochemistry, 95(2), 351–358. https://doi.org/10.1016/0003-2697(79)90738-3.
Ortiz, J. G., Opoka, R., Kane, D., & Cartwright, I. L. (2009). Investigating arsenic susceptibility from a genetic perspective in Drosophila reveals a key role for glutathione synthetase. Toxicological Sciences, 107(2), 416–426. https://doi.org/10.1093/toxsci/kfn192.
Patel, D., & Chinoy, N. J. (1998). Influence of fluoride on biological free radical reactions in ovary of mice and its reversal. Fluoride, 31, 27–32.
Patel, R. R. (2020). Tulsi: The queen of medicinal herbs. Journal of Bioequivalence & Bioavailability, 12, 407.
Posgai, R., Cipolla-McCulloch, C. B., Murphy, K. R., Hussain, S. M., Rowe, J. J., & Nielsen, M. G. (2011). Differential toxicity of silver and titanium dioxide nanoparticles on Drosophila melanogaster development, reproductive effort, and viability: size, coatings and antioxidants matter. Chemosphere, 85(1), 34–42. https://doi.org/10.1016/j.chemosphere.2011.06.040.
Rand, M. D., Dao, J. C., & Clason, T. A. (2009). Methylmercury disruption of embryonic neural development in Drosophila. Neurotoxicology, 30(5), 794–802. https://doi.org/10.1016/j.neuro.2009.04.006.
Rand, M. D., Montgomery, S. L., Prince, L., & Vorojeikina, D. (2014). Developmental toxicity assays using the Drosophila model. Current Protocols in Toxicology, 59(1), 1.12.1–1.12.20. https://doi.org/10.1002/0471140856.tx0112s59.
Reiter, L. T., Potocki, L., Chien, S., Gribskov, M., & Bier, E. (2001). A systematic analysis of human disease-associated gene sequences in Drosophila melanogaster. Genome Research, 11(6), 1114–1125. https://doi.org/10.1101/gr.169101.
Ribeiro, D. A., Cardoso, C. M., Yujra, V. Q., Viana, M. D. B., Aguiar, O., Pisani, L. P., & Oshima, C. T. F. (2017). Fluoride induces apoptosis in mammalian cells: in vitro and in vivo studies. Anticancer Research, 37, 4767–4777. https://doi.org/10.21873/anticanres.11883.
Sarkar, S., Roy, R., & Roy, S. (2018). Flubendiamine affects visual and locomotory activities of D. melanogaster for three successive generations (P, F1 and F2). Invertebrate Neuroscience, 18, 6.
Sauerheber, R. (2013). Physiological conditions affect toxicity of ingested industrial fluoride. Journal of Environmental and Public Health, 2013, 1–13. https://doi.org/10.1155/2013/439490 Article ID 439490.
Shivananjappa, M., & Joshi, M. (2012). Aqueous extract of tulsi (Ocimum sanctum ) enhances endogenous antioxidant defenses of human hepatoma cell line (HepG2). Journal of Herbs, Spices & Medicinal Plants, 18(4), 331–348. https://doi.org/10.1080/10496475.2012.712939.
Siddique, Y. H., Faisal, M., Naz, F., Jyoti, S., & Rahul (2014). Role of Ocimum sanctum leaf extract on dietary supplementation in transgenic Drosophila model of Parkinson’s disease. Chinese Journal of Natural Medicines, 12(10), 777–781. https://doi.org/10.1016/S1875-5364(14)60118-7.
Singh, V., Chowdhary, R., Shah, S., & Yasmin, S. (2020). Effect of fluoride on the reproductive output of Drosophila melanogaster. Global Journal of Science Frontier Research. C, Biological Science, 20(1), 17–20.
Susheela, A. K. (2013). Dental fluorosis and its extended effects. The Indian Journal of Pediatrics, 80, 715–717.
Sykiotis, G. P., & Bohmann, D. (2010). Stress-activated cap’n’collar transcription factors in aging and human disease. Science Signalling, 3, re3.
Vani, M. L., & Reddy, K. P. (2000). Effects of fluoride accumulation on some enzymes of brain and gastrocnemius muscle of mice. Fluoride, 33, 17–26.
Wang, A. G., Xia, T., Chu, Q. L., Zhang, M., Liu, F., Chen, X. M., & Yang, K. D. (2004). Effects of fluoride on lipid peroxidation, DNA damage and apoptosis in human embryo heaptocytes. Biomedical and Environmental Sciences, 17(2), 217–222.
Wasserkort, R., & Koller, T. (1997). Screening toxic effects of volatile organic compounds using Drosophila melanogaster. Journal of Applied Toxicology, 17(2), 119–125. https://doi.org/10.1002/(SICI)1099-1263(199703)17:2<119::AID-JAT415>3.0.CO;2-9.
Zhan, X. A., Wang, M., Xu, Z. R., & Li, J. X. (2006). Toxic effects of fluoride on kidney function and histological structure in young pigs. Fluoride, 39, 22–26.
Acknowledgements
We are grateful to the Principal, Patna Women’s College, for providing us all the facilities for carrying out the study. We are also grateful to Mr. Sumeet Ranjan for the helpful support and to Dr. M C Arunan of HBCSE, Mumbai, and CUBE mailing group for their valuable suggestions.
Funding
No funding was availed for this study.
Author information
Authors and Affiliations
Contributions
The manuscript was drafted by SP; life tables and graphs were prepared by SK and data were compiled and analysed partly by HR. SY designed the experiment, performed final data analyses and revision of manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Perveen, S., Kumari, S., Raj, H. et al. Effects of sodium fluoride and Ocimum sanctum extract on the lifespan and climbing ability of Drosophila melanogaster. JoBAZ 82, 32 (2021). https://doi.org/10.1186/s41936-021-00229-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41936-021-00229-8