
Abstract
Background
The wide unregulated use of malathion has produced severe health hazards. N-acetylcysteine (NAC) is a known glutathione precursor, and there is a growing attention concerning its beneficial effects against pesticide-induced toxicity. The present study was designed to investigate the therapeutic effects of NAC against malathion-induced hepatotoxicity, oxidative stress, genotoxicity, immunotoxicity, inflammation, and acetylcholinestrase alteration in rats.
Methods
Four groups comprised of 25 male rats each. Group 1 received distilled water, group 2 received NAC 150 mg/kg/day, group 3 received malathion 50 mg/kg/day, and group 4 received malathion 50 mg/kg/day followed by NAC 150 mg/kg/day for 90 consecutive days. Aspartate transaminase; alanine transaminase; alkaline phosphatase and lactate dehydrogenase; lipid peroxidation; reduced glutathione and total antioxidant capacity; DNA fragmentation; apoptosis and antiapoptosis-related gene expression; leukocyte counts; myeloperoxidase and immunophenotyping of CD4+ and CD8+; interleukin-1β, interleukin-6, and interferon-γ expression; and acetylcholinestrase were assessed.
Results
Malathion administration resulted in significant hepatic injury, immunotoxicity, genotoxicity, oxidative stress injury, inflammation, and significant reduction in acetylcholinestrase activity. Furthermore, malathion showed damaging histopathological effects on liver tissue. NAC treatment significantly attenuated all the previously mentioned biochemical, molecular, and histopathological alterations induced by malathion.
Conclusion
NAC had therapeutic effects against the detrimental hazards of malathion. Administering NAC to vulnerable risk groups is recommended.
Similar content being viewed by others
Background
Pesticides are considered to be absolutely necessary for the production of an adequate food supply for an increasing world population. The remaining residues of pesticides in air, water, and harvested crops could have a deleterious effect on human health (Prodhan et al. 2009). There is a lack in official reports of pesticide poisoning in Egypt, but suffering from chronic toxicity was reported in more than 60% of workers involved in pesticide applications (Mansour 2008).
Malathion is an organophosphate insecticide that has been extensively used throughout the world for some time as a dichlorodiphenyltrichloroethane (DDT) substitute. In this concern, contamination of the environment with insecticides, including malathion, is considered deleterious because of mutagenic and carcinogenic effects, and numerous other toxic effects on the brain, lung, mucous membrane, skin, immune system, kidney, liver, and blood, through different proposed mechanisms (Mohamed et al. 2010).
Organophosphorus compounds, including malathion, are well known for their mechanism of toxicity in living organisms through inhibition of acetylcholinesterase, with subsequent accumulation of the neurotransmitter acetylcholine and activation of both cholinergic muscarinic and cholinergic nicotinic receptors. Added to this previously mentioned mechanism, induction of oxidative stress, reduction of antioxidant enzyme activity, and triggering inflammatory response have been noted as co-lethal mechanisms of toxicity (Govindarajan et al. 2019).
Lately, there has been a considerable attention to find protective compounds with a considerable role in protecting living organisms from the toxic consequences of pesticides (Nurulain et al. 2015).
N-Acetylcysteine (NAC) is a thiol-containing compound, known for its powerful antioxidant and anti-inflammatory actions. Its antioxidant action originates mainly from its ability to stimulate reduced glutathione (GSH) synthesis, preserving intracellular GSH levels and scavenging reactive oxygen species (ROS). Recent researches reported that NAC may have beneficial roles against organophosphorus toxicity through acting by different mechanisms (Yurumez et al. 2007; Dhouib et al. 2016).
Methodology
Animals
One hundred (100) adult male albino Wistar rats, 10 weeks old, were obtained from Animal House of Research Institute of Ophthalmology, weighting 150–200 g. Rats were housed one per clean plastic cage. Animals were acclimatized to standard room temperature and to 12-h light/dark cycles. Rats were supplied with balanced food and water. The experiment was approved by the Ethical Committee of Faculty of medicine, Cairo University, and by the Institutional Animal Care and Use Committee (IACUC)—Cairo University with approval number CU III F 50 17.
Chemicals
Malathion (dimethoxythiophosphorylthiosuccinate) 57% was purchased from Directorate of Agriculture, Ministry of Agriculture, Giza, Egypt, and was manufactured by Cheminova Agro A/S Company, Denmark. N-Acetylcysteine was purchased from the pharmacy of National Center of Clinical and Environmental Toxicology, Faculty of Medicine, Cairo University, and was produced by South Egypt Drug Industries Company, Egypt.
Study design
Rats were randomly divided into four groups, with 25 rats in each group, and were treated daily by oral gavage, around 9 a.m., as follows for 90 successive days:
Group 1: Control group received distilled water daily.
Group 2: Received N-acetylcysteine “150 mg/kg/day” dissolved in distilled water with a 2 ml dose volume. (Yurumez et al. 2007).
Group 3: received malathion “50 mg/kg/day” dissolved in distilled water with a 0.2 ml dose volume. This dose is equivalent to occupational exposure level (El-Gharieb et al. 2010) and also corresponds to 1/40 LD50 (since LD50 = 2100 mg/kg of body weight for rats) (Lasram et al. 2014).
Group 4: received malathion “50 mg/kg/day” dissolved in distilled water followed by N-acetylcysteine “150 mg/kg/day” dissolved in distilled water, 2 h after malathion ingestion.
On the 90th day and 2 h after the last administration, blood samples were collected from retro-orbital venous plexus for biochemical analysis, then all rats were anesthetized by high-dose anesthetic agent [ketamine hydrochloride (100 mg/kg, i.p.)] then liver and spleen tissues were excised and washed by phosphate buffered saline, and frozen at − 80°C.
Biochemical and molecular parameters
-
1.
Liver function enzymes: Serum aspartate transaminase (AST), alanine transaminase (ALT), alkaline phosphatase (ALP), and lactate dehydrogenase (LDH) were assessed by routine laboratory investigations using kit supplied by BioMed, Egypt.
-
2.
Lipid peroxidation (LPO): The extent of LPO in liver homogenate was estimated as the concentrations of malondialdehyde (MDA) which is the end product of LPO according to the method of Ohkawa et al. (1979).
-
3.
Reduced glutathione (GSH): GSH level in hepatic liver tissue was determined according to the method of Griffith (1980)
-
4.
Total antioxidant capacity (TAC): It was measured by ferric reducing ability of plasma (FRAP) method (Benzie and Strain 1996).
-
5.
Quantitative real-time PCR for analysis of Bax, Bcl-2, and inflammatory markers; interleukin 1β (IL-1β), interleukin 6 (IL-6), interferon gamma (IFN-γ) genes’ expressions.
Total RNA extraction
According to the manufacturer’s instruction of SV Total RNA Isolation System (Promega, Madison, WI, USA), total RNA was extracted from tissue homogenate. RNA concentrations and purity were measured using an ultraviolet spectrophotometer.
Complementary DNA (cDNA) synthesis
According to the manufacturer’s protocol of SuperScript III First-Strand Synthesis System (#K1621, Fermentas, Waltham, MA, USA), the cDNA was synthesized from 1 μg RNA.
Real-time quantitative PCR (RT-PCR)
An Applied Biosystem with software version 3.1 (StepOne™, USA) was used to perform real-time PCR amplification and analysis. The reaction included SYBR Green Master Mix (Applied Biosystems) gene-specific primer pairs (Table 1) and was designed with Gene Runner Software (Hasting Software, Inc., Hasting, NY) from RNA sequences from the gene bank. All primer sets had a calculated annealing temperature of 60 °C. Quantitative RT-PCR was performed in a 25-μl reaction volume consisting of 2X SYBR Green PCR Master Mix (Applied Biosystems), 900 nm of each primer, and 2 μl of cDNA. Relative expression of studied gene mRNA was calculated using the comparative Ct method. All values were normalized to beta actin which was used as the control housekeeping gene.
-
6.
DNA damage by DNA fragmentation:
DNA was extracted from tissue lysate using the kit supplied by Qiagen following the recommended steps, then DNA fragmentation was detected in the extracted DNA through gel electrophoresis and was visualized and photographed under UV light.
-
7.
Leukocytes count: It was detected by homocytometer (Improved Neubauer, China).
-
8.
Myeloperoxidase activity (MPO): Liver MPO was determined using a spectrophotometry according to the method of Mullane et al. (1985).
-
9.
Immunophenotyping of CD4+ and CD8+: It was estimated by flow cytometry according to Novelli et al. (2000).
-
10.
Acetylcholinestrase (AChE): Activity of AChE in liver was estimated using spectrophotometry according to the method of Ellman et al. (1961).
Histopathological examination
Small pieces from the right lobes of the livers of the rats in the different studied groups were dissected and fixed for 24 h with 10% neutral formalin solution. They were processed in a sequence of ethanol and solutions and finally embedded in paraffin wax blocks. Tissues blocks were sectioned at 4 μm thickness, followed by deparaffinization by xylene and staining with hematoxylin and eosin (H&E) to detect liver injury as well as Masson trichrome stain to highlight fibrosis. The sections were viewed and photographed using Olympus light microscope (Olympus CX41) with mounted photographic camera (Olympus SC100).
Statistical analysis
Data was coded and entered using the statistical package SPSS version 25. Data was summarized using mean and standard deviation. Comparison between groups was done using one-way analysis of variance (ANOVA) test and post hoc pairwise comparison. Probability (P) values equal or less than 0.05 were considered as statistically significant.
Results
Effect of malathion and malathion plus NAC treatment on liver function enzymes
There was significant (P < 0.001) increase in mean values of serum levels of ALT, AST, ALP, and LDH in group 3 compared to controls, also there was significant (P < 0.001) decrease in their mean values in group 3 (Table 2).
Effect of malathion and malathion plus NAC treatment on oxidant/antioxidant status markers
As shown in Table 2, significant (P < 0.001) reduction in mean values of both GSH and TAC levels was observed in group 3; in contrast, mean value of MDA level showed significant (P < 0.001) increase in group 3 compared to control groups. Group 4 showed significant (P < 0.001) increase in mean values of both GSH and TAC levels and showed significant (P < 0.001) decrease in mean values of MDA level compared to group 3. Furthermore, there was no significant difference in TAC between group 4 and control group 1.
Effect of malathion and malathion plus NAC treatment on genotoxicity markers
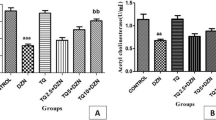
Apoptosis gene “Bax” expression and DNA fragmentation (Fig. 1) showed the highest mean values in group 3 while antiapoptosis gene “Bcl-2” expression showed the lowest mean value in group 3. While group 4 showed a significant (P < 0.001) decrease in Apoptosis gene “Bax” expression and DNA fragmentation and significant (P < 0.001) increase in antiapoptosis gene “Bcl-2.” Moreover, there was no significant difference in DNA fragmentation between group 4 and control groups 1 and 2 (Table 2).

An agarose gel electrophoresis show DNA fragmentation, where M is DNA marker with 100 bp, lanes 1, 2, and 4 representing groups 1, 2, and 4, respectively, show DNA without streaks or laddering while lane 3 representing group 3 shows DNA with marked streaks and laddering (fragmented)
Effect of malathion and malathion plus NAC treatment on immune-modulatory, inflammatory markers and AChE
Mean values of immune-modulatory marker (leukocytes, MPO, CD4%, and CD8%) levels and expression of inflammatory markers (IL-1β, IL-6, and IFN-γ) were found to be increased significantly (P < 0.001) in group 3 compared to control groups, whereas group 4 showed significant (P < 0.001) reduction in inflammatory markers expression compared to group 3 (Table 3).
Mean value of AChE activity showed a significant (P < 0.001) decrease in group 3 compared to control groups, while group 4 showed a significant (P < 0.001) increase in AChE activity compared to group 3. In addition, there was no significant difference in AChE activity between group 4 and control group 1 (Table 3).
Histopathological findings
Histopathological examination of the livers showed that the livers of the control and NAC-treated rats exhibited unremarkable pathological changes, preserved hepatic lobular architecture, arrangement of the hepatocytes in thin double cell thick plates, patent sinusoids, and regular hepatocytes cytomorphology in the form of central vesicular nuclei and abundant eosinophilic cytoplasm and revealed no evident portal or bridging fibrosis through Masson trichrome staining (Fig. 2). In contrast, the livers of the malathion-treated animals showed severe hepatic lesion displayed as remarkable cell loss, focal necrosis (severe inflammatory reaction), cytoplasmic clearing, focal nucleomegaly with hyperchromasia, focal nuclear pyknosis, and congested portal veins with areas of interstitial hemorrhage as well as focal obliterated sinuses and revealed marked expansion of portal areas by fibrous tissue evidenced by presence of collagen, yet there was no evident bridging fibrosis through Masson trichrome staining (Fig. 3). While the livers of the malathion plus NAC-treated animals exhibited less hepatic injury, unremarkable cell loss (minimal inflammatory reaction), and normal cytoplasmic staining with minimal clearing, the nuclei were relatively hyperchromatic, yet with regular nuclear-cytoplasmic ratio, the sinusoids were patent with no evident interstitial hemorrhage; however, the portal veins showed mild congestion and revealed mild expansion of portal areas by fibrous tissue evidenced by the presence of minimal collagen deposition through Masson trichrome staining (Fig. 4). Table 4 shows grading and summarization of these histopathological findings.

Photomicrography of sections in rat’s liver of control group (a) and NAC administered group (b) showed normal hepatocytes with vesicular nuclei and abundant eosinophilic cytoplasm, arranged in thin cords with preserved lobular architecture, normal portal tracts, central vein, and patent sinusoids. No remarkable fibrosis in sections examined by Masson trichrome for both control groups

Photomicrography of sections in rat’s liver administered malathion showing markedly congested portal vein (triangle), focal necrosis (short arrow), focal obliterated sinusoids (arrow head), and interstitial hemorrhage (star) as well as pyknosis (long arrow), vacuolation (square head), nuclear enlargement, and hyperchromasia (circle head) of hepatocytes. Marked portal fibrous tissue deposition (X), evidenced by Masson trichrome stain

Photomicrography of sections in rat’s liver administered malathion and treated by NAC showed mildly congested portal vein (triangle), unremarkable focal necrosis (arrow), minimal cellular degeneration (arrow head), more or less patent sinusoids (square head), and hyperchromatic regular nuclei (circle head). Minimal portal fibrous tissue deposition (X) was traced by Masson trichrome stain
Discussion
The widely spread use of organophosphate pesticides by public health, and industrial and agricultural programs has resulted in significant potential health hazards for all living organisms, including human beings (Akbel et al. 2018).
In the present study, despite that malathion was given at 1/40 of the oral LD50, numerous biochemical and pathological changes were observed. According to the current study, oral exposure to 50 mg/kg malathion in rats for 90 days has led to significant increase in ALT, AST, ALP, and LDH serum concentrations. This could be attributed to the liver injury evidenced by disturbed biochemical parameters and also confirmed by histopathological findings including hemorrhage, congestion, and collagen deposition as will be discussed below. Studies conducted by Akbel et al. (2018) and Lasram et al. (2014) also reported liver injury after malathion administration. After NAC treatment, there was significant decrease in ALT, AST, ALP, and LDH serum concentrations; this goes in agreement with Lasram et al. (2014) after NAC supplementation, and this may be attributed to the hepatoprotective effects of NAC and resulting attenuation of liver damage as reported by biochemical parameters analysis and by histopathological findings in the present study.
The oxidative stress created by malathion biotransformation into malaoxon and by malathion detoxification through conjugation with glutathione which leads to generation of ROS and depletion of antioxidant markers was evidenced in the current study by increased LPO, as evidenced by the formation of MDA, and depressed antioxidant status, as evidenced by depletion of GSH and TAC, in the liver of rats treated by malathion. Attachment of resulting free radicals to hepatocyte membranes leads to LPO which damages these membranes and causes necrosis, thereby causing structural breakdowns in hepatocytes membranes, leading to their damage and release of intracellular cytosol (AST, ALT, ALP, and LDH) into the blood (Celik and Suzek, 2008). Analysis of GSH and MDA by Akbel et al. (2018), Lasram et al. (2014), and Bhatti et al. (2013) showed consistent results with those of the current study.
As NAC is known for its antioxidant activity, so that it has the ability to detoxify or remove malathion via a GSH-dependent pathway. The demonstrated hepatoprotective effect might be attributed to the homeostasis in the oxidant/antioxidant status supplied by NAC as proved by the current study as NAC restored the altered oxidative stress markers. This offered protection by NAC against oxidative stress injury keep the structural integrity of hepatic cells preventing release of intracellular enzymes into the blood (Izadia et al. 2011). Results of the current study regarding MDA and GSH are in accordance with Lasram et al. (2014) and Yurumez et al. (2007) after using NAC against oxidative injury induced by malathion and fenthion, respectively.
Molecular mechanisms of genotoxicity of malathion include induction of oxidative stress, alkylation, and immunotoxicity (R´ EUS et al. (2008). DNA is one of the main cellular targets of ROS in addition to cellular lipids and proteins. High levels of DNA damage due to released free radicals may exceed the capacity of cellular repair, thus triggering mutations and apoptosis through causing persistent activation of apoptosis-related genes and inhibition of antiapoptosis-related genes which leads to induction of apoptosis and eventually cell death (Deavall et al. 2012; Arulselvan et al. 2016). This was proved by the current study as malathion-treated rats showed significantly increased expression of apoptosis gene “Bax” and DNA fragmentation and significantly decreased expression of antipoptosis gene “|Bcl-2” in rat livers. On the other hand, the antioxidant activity of NAC prevents DNA damage by ROS leading to inhibition of apoptosis-related pathway and stimulation of antiapoptosis-related pathway, and to the best of our knowledge, this is the first study investigating the therapeutic effect of NAC against malathion-induced genotoxicity in rats.
Initial injury from malathion toxic metabolites and overproduction of ROS could directly disturb the hepatocyte membranes as a result of LPO; this is followed by activation of the Kupffer cells in the liver, triggering a cascading series of cellular processes in the form of massive release of inflammatory cytokines such as IL-1β, IL-6, and IFN-γ. So, the total peripheral circulating leucocyte count increases and neutrophils are attracted by these cytokines to the site of injury, as evidenced by increasing activity of MPO. Also, cellular immunity is stimulated, as evidenced by the increasing CD4% and CD8%, subpopulations of T lymphocytes, as noted in the current study and study conducted by Lasram et al. (2014).
Protection against liver immunotoxicity and inflammation offered by NAC are attributed to its anti-inflammatory and immunomodulatory effects. The anti-inflammatory effect of NAC is accompanied by production of specific proteins which inhibit IKKβ/NF-κB axis so NAC can adjust the expression and the activity of these transcription factors which are involved in triggering the inflammatory cascading series (Pajonk et al. 2002; Samuni et al. 2013). It was also reported that NAC has attenuated cytokine release during the earlier phase of immune proliferation training (Omara et al., 1997). Moreover, NAC has reduced TNFα, IL-6, and IL-1β in patients subjected to hemodialysis or septic shock. Also, the antioxidant effect of NAC protects against cell injury, thus limits cytokine release and immune stimulation (Emet et al. 2004; Nascimento et al. 2010). This was proved in the current study as the malathion plus NAC-treated group showed significant decrease in immunotoxicity and inflammatory markers.
The reduction in AChE activity in malathion-treated rats is due to the fact that malathion acts essentially through irreversible inhibition of AChE at cholinergic junctions of the nervous system (Galloway and Handy 2003). This result was also approved by previous studies as Ouardi et al. (2019) who analyzed AChE in mice brain tissue after low-dose malathion administration. Improvement in AChE activity in malathion plus NAC-treated group may be attributed to the ability of NAC to facilitate rapid elimination of toxic malathion metabolites from the body thus diminishing its action (Lasram et al. 2014).
Regarding the histopathological findings in the present study, severe damaging lesions were noted in the livers of the malathion-treated rats. These changes are consistent with the changes in various biochemical parameters that were also observed in the present study. Analogical histopathological changes in the livers of the malathion-treated rats were reported by Kalender et al. (2010) and El-Gharieb et al. (2010). Histopathological changes showed a great improvement of the present study in malathion plus NAC-treated rats. Again, this is the first study investigating the effect of NAC against malathion-induced histopathological changes in liver sections of rats to the best of our knowledge.
Conclusion
The present study concluded that NAC treatment had attenuated all the biochemical, molecular, and histopathological alterations induced by malathion especially DNA fragmentation, total antioxidant capacity (TAC), and AChE levels as they were restored to their normal levels. Administering NAC to vulnerable risk groups is highly recommended.
Availability of data and materials
Data will not be shared with public access. Please contact author for data requests.
Abbreviations
- AChE:
-
Acetylcholinesterase
- ALP:
-
Alkaline phosphatase
- ALT:
-
Alanine transaminase
- ANOVA:
-
Analysis of variance
- AST:
-
Aspartate transaminase
- Bax:
-
Bacl2-associated X
- Bcl-2:
-
B cell lymphoma-2
- CD4:
-
Cluster of differentiation 4
- CD8:
-
Cluster of differentiation 8
- cDNA:
-
Complementary deoxyribonucleic acid
- Ct:
-
Cycle threshold
- DDT:
-
Dichlorodiphenyltrichloroethane
- DNA:
-
Deoxyribonucleic acid
- FRAP:
-
Ferric reducing ability of plasma
- GSH:
-
Reduced glutathione
- H&E:
-
Hematoxylin and eosin
- i.p.:
-
Intraperitoneal
- IACUC:
-
Institutional Animal Care and Use Committee
- IFN-γ:
-
Interferon gamma
- IKKβ/NF-κB:
-
IκB kinase/IκB kinase beta
- IL-1β:
-
Interleukin 1β
- IL-6:
-
Interleukin 6
- LD50:
-
Median lethal dose
- LDH:
-
Lactate dehydrogenase
- LPO:
-
Lipid peroxidation
- MDA:
-
Malondialdehyde
- MPO:
-
Myeloperoxidase
- NAC:
-
N-Acetylcysteine
- PCR:
-
Polymerase chain reaction
- RNA:
-
Ribonucleic acid
- ROS:
-
Reactive oxygen species
- RT-PCR:
-
Reverse transcription polymerase chain reaction
- SPSS:
-
Statistical Package for the Social Sciences
- TAC:
-
Total antioxidant capacity
- TNFα:
-
Tumor necrosis factor alpha
- UV:
-
Ultraviolet
References
Akbel E, Arslan-Acaroz D, Demirel HH, Kucukkurt I, Ince S (2018) The subchronic exposure to malathion, an organophosphate pesticide, causes lipid peroxidation, oxidative stress, and tissue damage in rats: the protective role of resveratrol. Toxicol Res 7(3):503–512
Arulselvan P, Fard MT, Tan WS, Gothai S, Fakurazi S, Norhaizan ME, Kumar SS (2016) Role of antioxidants and natural products in inflammation. Oxidative Med Cell Longev. https://doi.org/10.1155/2016/5276130
Benzie IF, Strain J (1996) The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: the FRAP assay. Anal Biochem 239(1):70–76
Bhatti GK, Sidhu IPS, Bhatti JS (2013) Protective effect of melatonin against malathion-induced alterations in antioxidant defense system and morphology of erythrocytes in wistar rats. J Basic Appl Sci 9:438–446
Celik I, Suzek H (2008) The hematological effects of methyl parathion in rats. J Hazard Mater 153(3):1117–1121
Deavall DG, Martin EA, Horner JM, Roberts R (2012) Drug-induced oxidative stress and toxicity. J Toxicol. https://doi.org/10.1155/2012/645460
Dhouib IE, Annabi A, Jallouli M, Elfazaa S, Lasram MM (2016) A minireview on N-acetylcysteine: an old drug with new approaches. Life Sci 151:359–363
El-Gharieb MA, El-Masry TA, Emara AM, Hashem MA (2010) Potential hepatoprotective effects of vitamin E and Nigella sativa oil on hepatotoxicity induced by chronic exposure to malathion in human and male albino rats. Toxicol Environ Chem 92(2):391–407
Ellman GL, Countney DK, Andres V Jr, Featheretone RH (1961) A new and colorimetric determination of acetylcholineesterase activity. Biochem Pharmacol 7:88–95
Emet S, Memis D, Pamukçu Z (2004) The influence of N-acetyl-l-cystein infusion on cytokine levels and gastric intramucosal pH during severe sepsis. Crit Care 8:172–179
Galloway T, Handy R (2003) Immunotoxicity of organophosphorous pesticides. Ecotoxicology 12(1-4):345–363
Govindarajan D, Chatterjee C, Shakambari G, Varalakshmi P, Jayakumar K, Balasubramaniem A (2019) Oxidative stress response, epigenetic and behavioral alterations in Caenorhabditis elegans exposed to organophosphorus pesticide quinalphos. Biocatal Agric Biotechnol 17:702–709
Griffith OW (1980) Determination of glutathione and glutathionedisulfide using glutathione reductase and 2-vinylpyridine. Anal Biochem 106:207–212
Izadia F, Jafarib M, Asgarib A, Salehib M (2011) Protective role of N-acetyl-cysteine on diazinon induced oxidative stress in rat kidney. Clin Biochem 44(13):S44
Kalender S, Uzun FG, Durak D, Demir F, Kalender Y (2010) Malathion-induced hepatotoxicity in rats: the effects of vitamins C and E. Food Chem Toxicol 48(2):633–638
Lasram MM, Lamine AJ, Douib IB, Bouzidb K, Annabi A, Belhadjhmida N, Ahmed M, El Fazaa S, Abdelmoula J, Gharbi N (2014) Antioxidant and anti-inflammatory effects of N-acetylcysteine against malathion-induced liver damage and immunotoxicity in rats. Life Sci 107(1-2):50–58
Mansour S (2008) Environmental impact of pesticides in Egypt. In: Reviews of Environmental Contamination and Toxicology, vol 196. Springer, New York, pp 1–51
Mohamed Z, Ahmed M, Fetyan N, Elnagdy S (2010) Isolation and molecular characterisation of malathion-degrading bacterial strains from waste water in Egypt. J Adv Res 1(2):145–149
Mullane KM, Kraemer R, Smith B (1985) Myeloperoxidaseactivity as a quantitative assessment of neutrophil infiltrationinto ischemic myocardium. J Pharmacol Methods 14(3):157–167
Nascimento MM, Suliman ME, Silva M, Chinaglia T, Marchioro J, Hayashi SY, Riella MC, Lindholm B, Anderstam B (2010) Effect of oral N-acetylcysteine treatment on plasma inflammatory and oxidative stress markers in peritoneal dialysis patients: a placebocontrolled study. Perit Dial Int 30:336–342
Novelli M, Savoia P, Cambieri I, Ponti R, Comessatti A, Lisa F, Bernengo MG (2000) Collagenase digestion a mechanical disaggregation as a method to extract and immunophenotype tumour lymphocytes in cutaneous T cell lymphomas. Clin Exp Dermatol 25:423–431
Nurulain SM, Ojha S, Tekes K, Shafiullah M, Kalasz H, Adem A (2015) Efficacy of N-acetylcysteine, glutathione, and ascorbic acid in acute toxicity of paraoxon to Wistar rats: survival study. Oxid Med Cell Longev. https://doi.org/10.1155/2015/329306
Ohkawa H, Ohishi N, Yagi K (1979) Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal Biochem 95(2):351–358
Omara FO, Blakley BR, Bermier J, Fournier M (1997) Immunomodulatory and protective effects of N-acetylcysteine in mitogen-activated murine splenocytes in vitro. Toxicology 116:219–226
Ouardi FZ, Anarghou H, Malqui H, Ouasmi N, Chigr M, Najimi M, Chigr F (2019) Gestational and lactational exposure to malathion affects antioxidant status and neurobehavior in mice pups and offspring. J Mol Neurosci 12:1–11
Pajonk F, Riess K, Sommer A, McBride WH (2002) N-acetyl-l-cysteine inhibits 26S proteasome function: implications for effects on NF-kappaB activation. Free Radic Biol Med 32(6):536–543
Prodhan MDH, Rahman MA, Akon MW, Ahmed MS, Kabir KH (2009) Determination of pre harvest interval for quinalphos, malathion, cypermethrin and diazinon in major vegatables. Annu Rep 10:146–158
R´ EGZ, Valvassori SS, Nuernberg H, Comim CM, Buss R, Teodoro P, Leffa D, Tavares P, Dagostim G, Paula MM, Andrade VM, Quevedo J (2008) DNA damage after acute and chronic treatment with malathion in rats. J Agric Food Chem 56(16):7560–7565
Samuni Y, Goldstein S, Dean OM, Berk M (2013) The chemistry and biological activities of nacetylcysteine. Biochim Biophys Acta 1830:4117–4129
Yurumez Y, Cemek M, Yavuz Y, Birdane YO, Buyukokuroglu ME (2007) Beneficial effect of N-acetylcysteine against organophosphate toxicity in mice. Biol Pharm Bull 30(3):490–494
Acknowledgements
Not applicable.
Funding
None.
Author information
Authors and Affiliations
Contributions
HMA did the experimental work and writing the paper. EAE is responsible for establishing the experimental design of the research, starting from the idea, interpretation of the results, to the critical revision of the paper. AAA provided assistance in writing, establishing the experimental design of the research, and the final revision. NKE did the final revision of the paper. LAR is responsible for the biochemical and molecular work up of the research. ASA is responsible for the histopathological part of the work. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval
The study work was conducted after the approval of Ethical Committee, Faculty of medicine, Cairo University and by the Institutional Animal Care and Use Committee (IACUC) – Cairo University with approval number CU III F 50 17.
Consent for publication
Not applicable.
Competing interests
The authors declared that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Aboubakr, H.M., Elzohairy, E.A., Ali, A.A. et al. Therapeutic effects of N-acetylcysteine against malathion-induced hepatotoxicity. Egypt J Forensic Sci 9, 34 (2019). https://doi.org/10.1186/s41935-019-0142-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41935-019-0142-6