
Abstract
Background
Behcet’s disease (BD) is a multisystem and multifactorial autoimmune disease characterized by relapsing episodes of oral aphthae, genital ulcers, and ocular and skin lesions. Toll-like receptor 9 (TLR9) has pro-inflammatory roles and its genetic variants might be involved in the pathogenesis of inflammatory diseases.
Methods
Two hundred five BD patients and 207 age and sex-matched healthy controls were evaluated for TLR9 single nucleotide polymorphisms − 1486 T/C (rs187084) and + 2848:G/A (rs352140) using polymerase chain reaction-restriction fragment length polymorphism (RFLP-PCR).
Results
Healthy individuals had a significantly higher frequency of rs187084 AG and AG + GG genotypes than BD patients (p = 0.02 and p = 0.018; respectively). Of interest, healthy males had a significantly higher frequency of rs187084 AG + GG genotype and G allele than male BD patients (p = 0.035 and p = 0.045; respectively). However, rs187084 AG genotype and G allele frequencies were significantly higher in male patients with genital aphthous (p = 0.01 and p = 0.046; respectively). Furthermore, a significantly higher frequency of rs352140 CT and TT + CT genotypes was detected in healthy individuals than in BD patients (p = 0.01, and p = 0.032; respectively). Such results were also seen in healthy females than female patients (p = 0.001, and p = 0.004; respectively). Haplotype analysis revealed a significantly higher frequency of A-C and G-C haplotypes among patients and healthy subjects, respectively (p = 0.002 and p = 0.000; respectively).
Conclusion
Our data suggested that rs187084 AG and AG + GG genotypes and rs352140 CT and TT + CT genotypes protect Iranian individuals from BD but rs187084 AG genotype and G allele predispose male BD individuals to genital aphthous. However, additional studies are required to verify these results.
Similar content being viewed by others

Introduction
Behcet’s disease (BD) is a complex, multisystem inflammatory condition, mainly characterized by mucocutaneous ulcers, and ocular and vascular involvement. Gastrointestinal and central nervous system (CNS) are involved in most severe cases [1]. The prevalence of BD depends on geographical region. Particularly, the outbreak of BD in the ancient Silk Road, which contains Italy, Turkey, Israel, Saudi Arabia, Iran, China, Korea, and Japan, is higher than in other countries [2, 3]. The disease is distributed equally between men and women but globally, males are more frequently affected than women and experience more severe forms of disease [4, 5]. The exact pathogenic reason for BD is not exactly known but some evidence pointed to crucial roles of immunological abnormalities and environmental factors [6, 7]. With recent progress in genetic research, many genes such as human leukocyte antigen (HLA), especially HLA-B51, have been introduced as candidates that increase the susceptibility to BD [8]. Toll-like receptors (TLRs) which mediate innate immune responses are suspected to play vital roles in autoimmune diseases [9]. Increasing evidence has revealed possible associations between gene expression, single nucleotide polymorphisms (SNP), and copy number variation of TLRs and molecules involved in their signaling and the development of BD [8, 10, 11]. TLR9 gene which is located on chromosome 3p21.3 is one of the susceptible regions associated with BD [12]. TLR9 which recognizes cytosine-phosphate-guanosine (CPG) motifs is expressed by B cells and acts as a sensor for bacterial infection to activate the innate immune cells [13, 14]. Besides, autoantibody production and induction of inflammatory cells are modulated through the TLR9 signaling pathway [14,15,16]. Previous studies have revealed the importance of TLR9 gene variants in the pathogenesis of autoimmune diseases such as Systemic lupus erythematosus (SLE) [17,18,19], rheumatoid arthritis (RA) [20], ankylosing spondylitis (AS) [21], acute myeloid leukemia (AML) [22], Hashimoto’s autoimmune thyroiditis [23], and Graves’ disease (GD) [24]. However, their possible association with BD has provided controversial results [24]. TLR9 rs187084 and rs352140 are located at the promoter region and second exon, respectively, where they regulate TLR9 expression [17, 25]. These SNPs are the most functional and their special genotypes might lead to changes in TLR9 gene expression. Therefore, in this study, the possible association between these TLR9 genetic variants and the pathogenesis of BD was analyzed among Iranian individuals.
Materials and methods
Participants
This study included 205 BD patients, including 89 males and 116 females with a mean age of 36 ± 0.72 years, who were diagnosed based on the International Criteria for Behcet Disease in Hafez Rheumatology Outpatient Clinic, Shiraz, Iran [26]. Pregnant patients and those with malignancy, cancer, and other autoimmune diseases were excluded from the study. Two hundred seven age- and sex-matched healthy blood donors (100 males and 107 females with a mean age of 37 ± 0.4) who had no history of autoimmune or inflammatory disease participated as the healthy controls. Our study was approved by the Ethics Committee of the Shiraz University of Medical Science (Code: IR.SUMS.REC.1400.695) and was conducted following the ethical principles outlined in the Declaration of Helsinki (https://www.wma.net/policies-post/wma-declaration-of-helsinki-ethical-principles-for-medical-research-involving-human-subjects/). Before participation, all the individuals provided written informed consent. The clinical, laboratory, and demographic parameters of participants are presented in Table 1.
DNA extraction and SNPs genotyping
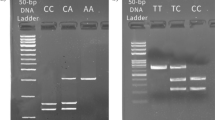
A total of 5 mL of whole blood samples in EDTA was collected. Genomic DNA was extracted using the salting out method and was stored at -20˚C until analysis [27]. A polymerase chain reaction followed by restriction fragment length polymorphism (PCR-RFLP) was used for genotyping TLR9 SNPs (rs187084 and rs352140) [28]. The PCR was done in 10 µl reaction volume containing 2 µl of DNA, 0.7 µM of each primer (Pioneer, South-Korea, Table 2), 0.1 µl Taq DNA polymerase (CinnaGen, Iran), 0.3 µl dNTPs (CinnaGen, Iran), 1 µl of 10X buffer, 0.7 µl MgCl2 and 4.5 µl distilled water. For the detection of DNA contamination, no template control (NTC) was included. The amplification conditions were initial denaturation at 95˚C for 5 min, then 30 cycles of denaturation at 95 °C for 30 s, annealing at 62 °C for 30 s, extension at 72 °C for 30 s, followed by a final extension at 72 °C for 5 min. Subsequently, PCR products were digested and incubated at 37˚C for 24 h. After that, fragments were assessed using 3% agarose gel with DNA safe stain (CinnaGen, Iran) and visualized in a UV transilluminator (Upland, CA) (Supplementary Fig. 1). The sequences of forward and reverse primers for each SNP, required restriction enzyme are shown in Table 2.
Statistical analysis
All statistical analyses were performed using Statistical Package for Social Sciences (SPSS) and Epi-info 7.2.2.6 software packages. Hardy–Weinberg equilibrium was calculated by Arlequin (version 3.1) software to assess the consistency of the genotype distribution. Comparison of the genotype and allele frequencies of TLR9 polymorphisms between studied groups was done by a two-tailed person’s Chi-square (χ²) test. Statistical significance in all tests was set at p values less than 0.05.
Results
Demographic and clinical history of BD patients
Clinical manifestation and demographic data of BD patients and controls are shown in Table 1. These data show no significant differences in age and sex between patients and controls (p = 0.511 and p = 0.214; respectively).
Genotype and allele frequencies of TLR9 gene polymorphisms
Both rs187084 and rs352140 followed the Hardy–Weinberg equilibrium (p = 0.06 and p = 0.78; respectively). A significantly higher frequency of rs352140 CT and TT + CT genotypes was observed in the healthy group compared to the BD patients (p = 0.01 and p = 0.032; respectively; Table 3). Further analysis revealed such results in healthy females compared with the female patients (p = 0.001 and p = 0.004; respectively). In the case of rs187084, significantly higher frequencies of AG and AG + GG genotypes were indicated in healthy individuals compared with the patients (p = 0.02 and p = 0.018; respectively). rs187084 GG + AG genotype combination and G allele frequencies were also significantly higher among healthy males compared with the male BD patients (p = 0.035 and p = 0.045; respectively). As shown in Table 4, analysis of patients according to their disease manifestation revealed only a significant association between rs187084 AG genotype and G allele and genital aphthous in male BD patients (p = 0.01 and p = 0.046; respectively).
Haplotype analysis
Table 5 indicates that A-C haplotype frequency is significantly higher in BD patients (p = 0.002) while G-C haplotype is more frequent in the control group (p = 0.000) (Table 5).
Discussion
Although the etiopathogenesis of BD is not completely understood, genetic predisposition, environmental factors, microbial agent triggers, endothelial cell dysfunction, neutrophil hyperfunction, and autoimmune mechanisms have been supposed [28, 29]. It is confirmed that the interaction between genetic and environmental factors plays a vital role in the pathogenesis of BD [30]. Genome-wide association studies (GWAS) support that single nucleotide gene polymorphisms might predispose individuals to such autoimmune diseases [31]. Recently some studies were published to point out the possible role of TLR9 genetic variants rs352140 and rs187084, in the pathogenesis of some autoimmune diseases such as SLE, RA, AS, AML, and GD [17]- [24]. Here, in line with these studies, an association between rs352140 and rs187084 polymorphisms and BD was found among the Iranian population. To our knowledge, this is the first study to evaluate the possible role of TLR9 in the pathogenesis of BD and its manifestations among Iranians. Our results revealed a significantly higher frequency of rs187084 AG and AG + GG genotypes in healthy subjects, as well as AG + GG genotype and G allele in healthy males. These findings indicate a protective role for these genotypes and alleles among Iranian individuals against BD. Nevertheless, a positive association between rs187087 AG genotype and G Allele and genital aphthous was found in male BD patients. Surprisingly, these results indicate that despite the protective role of the AG genotype against BD, it might increase the risk of genital aphthous in male BD patients. Besides, a significantly higher frequency of rs352140 CT and CT + TT genotypes was shown in healthy individuals, especially healthy females than in BD patients. Again, it is concluded that these genotypes and alleles protect Iranian individuals from BD. There have been limited investigations into the association between TLR9 genotypes and the risk of BD. In a study by Ito et al., no significant association between rs187084 and rs352140 and susceptibility to BD has been found among Japanese individuals [12]. Fang et al. also evaluated these SNPs among Chinese BD patients and reached no significant association [32]. However, a study by Sakamoto et al. revealed a significantly higher frequency of rs352140 CC genotype among Japanese BD patients compared to the healthy control [33]. Furthermore, Dhifallah et al. showed an approximate to significant level difference for rs187084 TT genotype and T allele (p = 0.07 and p = 0.08, respectively) among Tunisian BD patients compared with the healthy subjects [34]. The differences in ethnicity, environmental factors that may impact gene expression, clinical heterogeneity, and limited sample size could explain the discrepancies between our findings and those of other studies. One limitation of the study is the absence of serum levels of TLR-9 that could improve the quality of the study.
Conclusion
In summary, our study indicated that rs187084 AG and AG + GG genotypes and rs352140 CT and TT + CT genotypes protect Iranian individuals from BD but rs187084 AG genotype and G allele predispose male BD individuals to genital aphthous. However, additional studies are required to verify these results. Besides, further functional and linkage analyses are necessary to elucidate the precise role of rs187084 and rs352140 in the pathogenesis of BD.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author upon reasonable request.
References
Rodríguez-Carrio J, Nucera V, Masala IF, Atzeni F. Behçet disease: from pathogenesis to novel therapeutic options. Pharmacol Res. 2021;167:105593.
Ortiz-Fernández L, Sawalha AH. Genetics of Behcet’s disease: functional genetic analysis and estimating disease heritability. Front Med. 2021;8:625710.
Kilian NC, Sawalha AH. Behçet’s disease in the United States: a single center descriptive and comparative study. Eur J Rheumatol. 2017;4(4):239.
Leccese P, Alpsoy E. Behçet’s disease: an overview of etiopathogenesis. Front Immunol. 2019;10:1067.
Escudier M, Bagan J, Scully C. Number VII Behcet’s disease (Adamantiades syndrome). Oral Dis. 2006;12(2):78–84.
Mohammad-Ebrahim H, Kamali-Sarvestani E, Mahmoudi M, Beigy M, Karami J, Ahmadzadeh N, et al. Association of killer cell immunoglobulin-like receptor (KIR) genes and their HLA ligands with susceptibility to Behçet’s disease. Scand J Rheumatol. 2018;47(2):155–63.
Tong B, Liu X, Xiao J, Su G. Immunopathogenesis of Behcet’s disease. Front Immunol. 2019;10:665.
Van der Houwen T, van Hagen P, van Laar J, editors. Immunopathogenesis of Behçet’s disease and treatment modalities. Semin Arthritis Rheum. 2022:52:e151956.
Cavalcante P, Barzago C, Baggi F, Antozzi C, Maggi L, Mantegazza R, et al. Toll-like receptors 7 and 9 in myasthenia gravis thymus: amplifiers of autoimmunity? Ann N Y Acad Sci. 2018;1413(1):11–24.
Bulur I, Onder M. Behçet disease: new aspects. Clin Dermatol. 2017;35(5):421–34.
Puccetti A, Fiore PF, Pelosi A, Tinazzi E, Patuzzo G, Argentino G, et al. Gene expression profiling in Behcet’s disease indicates an autoimmune component in the pathogenesis of the disease and opens new avenues for targeted therapy. J Immunol Res. 2018;2018:e4246965.
Song GG, Choi SJ, Ji JD, Lee YH. Toll-like receptor polymorphisms and vasculitis susceptibility: meta-analysis and systematic review. Mol Biol Rep. 2013;40:1315–23.
Wang D, Zhang C, Zhou Z, Pei F. TLR9 polymorphisms and systemic lupus erythematosus risk: an update meta-analysis study. Rheumatol Int. 2016;36:585–95.
Akira S, Takeda K. Toll-like receptor signalling. Nat Rev Immunol. 2004;4(7):499–511.
Christensen SR, Kashgarian M, Alexopoulou L, Flavell RA, Akira S, Shlomchik MJ. Toll-like receptor 9 controls anti-DNA autoantibody production in murine lupus. J Exp Med. 2005;202(2):321–31.
Leadbetter EA, Rifkin IR, Hohlbaum AM, Beaudette BC, Shlomchik MJ, Marshak-Rothstein A. Chromatin–IgG complexes activate B cells by dual engagement of IgM and toll-like receptors. Nature. 2002;416(6881):603–7.
Gharbia OM, Bassiouni SAR, Zaki MES, El-Beah SM, El-Desoky MM, Elmansoury EA, et al. Toll-like receptor 5 and toll-like receptor 9 single nucleotide polymorphisms and risk of systemic lupus erythematosus and nephritis in Egyptian patients. Egypt Rheumatol Rehabilitation. 2021;48(1):1–10.
Zhang J, Zhu Q, Meng F, Lei H, Zhao Y. Association study of TLR-9 polymorphisms and systemic lupus erythematosus in northern Chinese Han population. Gene. 2014;533(1):385–8.
Bashir MA, Afzal N, Hamid H, Kashif M, Niaz A, Jahan S. Analysis of single nucleotide polymorphisms encompassing toll like receptor (TLR)-7 (rs179008) and (TLR)-9 (rs352140) in systemic lupus erythematosus patients. Advancements Life Sci. 2021;8(2):103–7.
Kim W, Kim TH, Oh SJ, Kim HJ, Kim JH, Kim H-A, et al. Association of TLR 9 gene polymorphisms with remission in patients with rheumatoid arthritis receiving TNF-α inhibitors and development of machine learning models. Sci Rep. 2021;11(1):e20169.
Oliveira-Toré CF, Moraes AG, Martinez GF, Neves JS, Macedo LC, Rocha-Loures MA, et al. Genetic polymorphisms of and as susceptibility factors for the development of Ankylosing spondylitis and Psoriatic Arthritis. J Immunol Res. 2019;2019:e1492092.
Banescu C, Tripon F, Bojan AS, Trifa AP, Muntean C, Crauciuc GA, et al. Association of TLR4 Rs4986791 polymorphism and TLR9 haplotypes with acute myeloid leukemia susceptibility: a case-control study of adult patients. J Personalized Med. 2022;12(3):e409.
Dvornikova KA, Bystrova EY, Platonova ON, Churilov LP. Polymorphism of toll-like receptor genes and autoimmune endocrine diseases. Autoimmun rev. 2020;19(4):e102496.
Zhang Y, Liu J, Wang C, Liu J, Lu W. Toll-like receptors gene polymorphisms in autoimmune disease. Front Immunol. 2021;12:e672346.
Hassan AE, Shoeib S, Abdelmohsen E, Nazir A, Dawood A, Gamal H, et al. Toll-like receptor 9 (TLR9) gene C/T (rs352140) polymorphisms in adult primary immune thrombocytopenia. Clin Appl Thromb Hemost. 2020;26:e1076029620940050.
O’NEILL T, Rigby A, Silman A, Barnes C. Validation of the International Study Group criteria for Behçet’s disease. Rheumatology. 1994;33(2):115–7.
John S, Weitzner G, Rozen R, Scriver C. A rapid procedure for extracting genomic DNA from leukocytes. Nucleic Acids Res. 1991;19(2):e408.
Bouali E, Kaabachi W, Hamzaoui A, Hamzaoui K. Interleukin-37 expression is decreased in Behçet’s disease and is associated with inflammation. Immunol Lett. 2015;167(2):87–94.
Kacem O, Kaabachi W, Dhifallah IB, Hamzaoui A, Hamzaoui K. Elevated expression of TSLP and IL-33 in Behçet’s disease skin lesions: IL-37 alleviate inflammatory effect of TSLP. Clin Immunol. 2018;192:14–9.
Inal EE, Rüstemoğlu A, Inanır A, Ekinci D, Gül Ü, Yiğit S, et al. Associations of rs4810485 and rs1883832 polymorphisms of CD40 gene with susceptibility and clinical findings of Behçet’s disease. Rheumatol Int. 2015;35:837–43.
Kahmini FR, Gholijani N, Amirghofran Z, Daryabor G. Single nucleotide polymorphisms rs7799039 and rs2167270 in leptin gene and elevated serum levels of adiponectin predispose iranians to Behçet’s disease. Cytokine. 2023;162:e156100.
Fang J, Hu R, Hou S, Ye Z, Xiang Q, Qi J, et al. Association of TLR2 gene polymorphisms with ocular Behcet’s disease in a Chinese Han population. Investig Ophthalmol Vis Sci. 2013;54(13):8384–92.
Sakamoto N, Sekine H, Kobayashi H, Sato Y, Ohira H. Association of the toll-like receptor 9 gene polymorphisms with behcet’s disease in a Japanese population. Fukushima J Med Sci. 2012;58(2):127–35.
Dhifallah IB, Lachheb J, Houman H, Hamzaoui K. Toll-like-receptor gene polymorphisms in a Tunisian population with Behcet’s disease. Clin Experimental Rheumatol. 2009;27(2):58–62.
Acknowledgements
The authors express their gratitude to Mr. Mohammad-Reza Malekmakan and Mr. Mohammad-Reza Yazdani for their assistance in conducting the experiments.
Funding
Shiraz University of Medical Sciences, Iran provided the financial support for this study (Grant Numbers: 23811 and 23967).
Author information
Authors and Affiliations
Contributions
Z. Tadayon. performed the analysis and prepared a preliminary manuscript draft. A. Shahzadeh Fazeli helped in designing the study. G. Daryabor. conceived and designed the study, wrote the paper, and was involved in critically revising the manuscript. N. Gholijani. participated in drafting the article.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Our study was approved by the Ethics Committee of the Shiraz University of Medical Science (SUMS). It was conducted following the ethical principles outlined in the Declaration of Helsinki (https://www.wma.net/policies-post/wma-declaration-of-helsinki-ethical-principles-for-medical-research-involving-human-subjects/). Before participation, all the individuals provided written informed consent.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Tadayon, Z., Shahzadeh Fazeli, S.A., Gholijani, N. et al. Toll-like receptor 9 (TLR9) genetic variants rs187084 and rs352140 confer protection from Behcet’s disease among Iranians. BMC Rheumatol 8, 13 (2024). https://doi.org/10.1186/s41927-024-00382-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41927-024-00382-x