
Abstract
In this study, we isolated and characterized the acid-soluble skin collagen of Pacific bluefin tuna (PBT, Thunnus orientalis). The PBT skin collagen was composed of two α chains (α1 and α2) and one β chain. The denaturation temperature of PBT collagen was low although it was rich in proline and hydroxyproline. The primary structure of PBT skin collagen was almost identical to that of calf and salmon skin collagen; however, it differed with respect to the epitope recognition of the antibody against salmon type I collagen. These results suggest that the primary structure of skin collagen was highly conserved among animal species, although partial sequences that included the epitope structure differed among collagens.
Similar content being viewed by others

Background
Collagen is a major structural protein that is widely distributed in animal connective tissues. The primary structure of collagen is unique as it contains a glycine-rich repeat sequence (Gly-X-Y), in which prolyl and hydroxyprolyl residues at the X and Y positions determine the triple helical secondary structure (Gordon and Hahn, 2010; Ramshaw et al., 1998). Collagen is widely used in the food, cosmetic, biomedical, and pharmaceutical industries. Commercial sources of collagen are mainly derived from mammals such as cows and pigs. Marine collagen is advantageous over mammalian collagen because (i) marine animals are not affected by infectious diseases such as avian influenza, bovine spongiform encephalopathy (BSE), transmissible spongiform encephalopathy (TSE), and foot and mouth disease (FMD) observed in pigs and cattle, (ii) the consumption of marine collagen is acceptable to people with religious restrictions, and (iii) it has a lower thermal denaturation temperature than collagen of terrestrial animals (Nagai et al., 1999; Nagai et al., 2010; Senaratne et al., 2006), which is conducive for assimilation by the human digestive system.
In 2002, the first full-cycle aquaculture of Pacific bluefin tuna (PBT) was successfully performed at Kindai University, Japan (Sawada et al., 2005). Currently, more than 40,000 cultured juveniles are available from the bioventure company, A-Marine Kindai (Wakayama, Japan). This increased supply of bluefin tuna has triggered research into the effective use of the unused parts of the tuna, such as its skin and organs, to avoid environmental pollution and to promote economic sufficiency. Therefore, we have focused on PBT skin as a collagen-rich underused resource for functional food. Previously, we reported that dietary PBT skin protein and collagen hydrolysis exerts hepato-protective action in CCl4-intoxicated mice (Tanaka et al., 2012). In addition, the collagen from PBT, but not from salmon, mackerel, and carp, also reduced HepG2 and HeLa cell growth in a dose-dependent manner, suggesting the existence of a PBT skin collagen-specific primary structure and/or higher-order structural conformation (Han et al., 2011). However, little is known about characteristic feature and structural information of isolated PBT skin collagen.
Studies on the early life history of PBT, which addressed the morphological changes (Kaji et al. 1996; Miyashita et al. 2001), chemical content, enzyme activities (Takii et al. 1997), and the development of the digestive system during PBT embryogenesis (Miyashita et al. 1998), have provided information required for the development of mass seeding techniques. However, laboratory-reared PBT suffer high mortality during the rapid somatic growth stage in their early life (Sawada et al. 2005; Tanaka et al. 2007). For example, PBT possesses very sensitive skin, which renders its handling difficult during rearing of this species. Over 40% PBT juveniles die of skin injuries that are incurred during transportation with hand nets in the first week of transfer of these land-based farmed juveniles to open net cages (Ishibashi et al., 2009). Therefore, it is important to understand the property of type I collagen, which is a major component of PBT skin.
In this study, we isolated skin collagen from PBT and characterized certain properties.
Methods
Materials
Calf and salmon skin type I collagens were purchased from Wako Pure Chemicals (Osaka, Japan). All chemicals used in this study were of the highest purity available.
Isolation of PBT skin collagen
PBT (24–32 days after hatching) was obtained in an unfrozen state at 4 °C within 24 h after catching the tuna from culture fields of Aquaculture Research Institute, Uragami Station, Kindai University, Japan. The skin was dissected from the body and stored at − 20 °C. Bluefin tuna skin collagen was isolated using a previously reported procedure (Han et al., 2011) with slight modifications. All steps of the extraction was performed at 4 °C. The skin of PBT without the muscles and scales was cut into small pieces. The pieces were soaked in 0.1 M NaOH for 24 h with stirring. The NaOH solution was changed every 8 h to remove non-collagenous proteins and pigments. The pieces were washed with distilled water until neutral pH was obtained. The pieces were then defatted with methanol/chloroform (2:3) and washed with methanol and distilled water. For extracting collagen, the defatted pieces were stirred in 10 volumes (w/v) of 0.5 M acetic acid for 24 h. Pepsin (3130 U/mg solid; Nacalai Tesque Inc. Kyoto, Japan) was then added to the supernatant (7 μg/L), and the mixture was gently stirred for 48 h. Collagen was precipitated by salting out with 25% (w/v) NaCl and centrifuged at 5000×g for 30 min. The precipitate was dissolved in 0.5 M acetic acid and centrifuged (15,000×g, 60 min). The supernatant was dialyzed with stirring for 24 h against five changes of distilled water and lyophilized. The collagen sample was stored at − 20 °C until further analysis.
Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and peptide mapping
SDS-PAGE was performed using the Tris-HCl/glycine buffer system and 7.5% polyacrylamide gel described by Laemmli (1970) using the Tris-HCl/glycine buffer system with a 7.5% resolving gel and 4% stacking gel. The collagen sample was dissolved in sample buffer (0.5 M Tris-HCl, pH 6.8, containing 8% SDS, 30% glycerol, 0.2% bromophenolblue) containing 5% β-mercaptoethanol and then boiled for 5 min. Collagen samples (50 μg/well) were applied to sample wells and electrophoresed. The separated proteins were stained with Coomassie Brilliant Blue R-250. Peptide mapping was performed as described by Yata et al. (2001). The isolated collagens were digested with lysyl endopeptidase (Wako Pure Chemicals, Japan) at an enzyme/substrate ratio of 1:100 (w/w). Peptides generated by the protease digestion were separated by SDS-PAGE using 7.5% gel. The separated proteins and peptide were stained with Coomassie Brilliant Blue R-250.
Amino acid composition
The collagen sample was hydrolyzed in 6 N HC1 at 110 °C for 24 h. The hydrolysates were analyzed using an L-8800 automated amino acid analyzer (Hitachi High-Technologies, Tokyo, Japan).
Denaturation temperature
As previously reported by Nomura et al. (1996), the denaturation temperature of PBT skin collagen in 0.5 M acetic acid was measured using an Autopol III automatic polarimeter (Rudolph Research Co. Flanders, N J) at 589 nm.
UV-Vis spectra
The ultraviolet absorption spectra of collagen were recorded using a spectrophotometer (U-0080D, HITACHI, Japan) from 190 to 400 nm. The isolated collagen was dissolved in 0.5 M acetic acid to obtain a concentration of 0.05% (m/v).
Fourier transform infrared spectroscopy (FTIR)
Attenuated total reflection (ATR)-FTIR spectra of collagen was obtained using a Nicolet 6700 FTIR Spectrometer (Thermo Fisher Scientific, USA) equipped with ATR accessory. Spectra were recorded from 4000 to 500 cm− 1 at a data acquisition rate of 0.5 cm− 1 per point.
Cross-reactivity of PBT type I skin collagen with salmon collagen antibody
The cross-reactivity of PBT type I skin collagen with IgG-purified guinea pig antibody against salmon type I collagen was examined using three enzyme-linked immunosorbent assays (ELISA).
Direct ELISA
ELISA plates were coated with serial dilutions of collagen in phosphate-buffered saline (PBS) to determine the linear range of the sigmoid curve. Next, the plates were blocked with blocking buffer (200 μL) (Blocking One-P, Nacalai Tesque Inc. Kyoto, Japan) for 1 h at room temperature. Horseradish peroxidase (HRP)-labeled antibody against salmon collagen (100 μL) was added at 1/1000 dilution in PBS and incubated for 1 h at room temperature. The specificity for binding with immunoglobulins of salmon collagen antibodies was previously tested using western blot (data not shown). After incubation with 3, 3′, 5, 5′-tetramethylbenzidine (TMB) substrate buffer, absorbance was measured using a plate spectrophotometer at 405 nm.
Sandwich ELISA
ELISA plates were coated with 100 μL of 10 μg/mL salmon collagen antibody for 13 h at 4 °C. After blocking, the plates were incubated with serial dilutions of collagen (100 μL) in PBS for 1 h at room temperature. Next, HRP-labeled salmon collagen antibody (100 μL) was added at 1/1000 dilution in PBS and incubated for 1 h at room temperature. The colorimetric method was performed as mentioned above using the TMB substrate.
Inhibition ELISA
Serial dilutions of collagen were coated on ELISA plates. After coating, the plate was incubated for 1 h at room temperature (200 μL). Pre-incubated (1 h at room temperature) dilutions of a salmon collagen and HRP-labeled salmon collagen antibody were added and incubated for 1 h at room temperature. The colorimetric method was performed as mentioned above using the TMB substrate.
Results
Isolation of PBT skin collagen
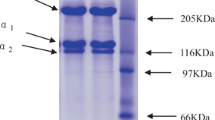
In this study, the acid-soluble skin collagen of PBT was isolated. The final protein recovery rate of the PBT skin collagen was 2.1 g/100 g and the dry yield was 5.4%. The isolated PBT skin collagen was analyzed using SDS-PAGE. The separation pattern shows that PBT skin collagen was composed of two α chains (α1 and α2) and one β chain similar to calf and salmon collagen (Fig. 1). The estimated molecular weights for the α1 and α2 chains were approximately 120 and 112 kDa, respectively, which is similar to previous observations (Nalinanon et al., 2007).

SDS polyacrylamide gel (7.5%) electrophoretic pattern for acid-soluble collagen. M.W. Marker: molecular weight marker
Amino acid composition of PBT skin collagen
Table 1 shows the amino acid composition of the PBT skin collagen. Glycine was the most abundant amino acid in the PBT skin collagen with a content of 27.58%. This is similar to the glycine content of calf (Giraud-Guille et al., 2000) and salmon skin gelatin (Arnesen and Gildberg, 2007). In addition, PBT skin collagen had high content of proline, alanine, and arginine; however, cysteine and phenylalanine were not detected.
UV-Vis spectra of PBT skin collagen
Figure 2 shows the UV-Vis spectra of PBT skin collagens scanned at 190–590 nm. The major peak was observed at 238 nm. There was also a swell distribution between 250 and 280 nm.

Ultraviolet spectra of PBT skin collagen
Denaturation temperature of PBT skin collagen
As shown in Fig. 3, the change in optical rotation of PBT skin collagen in solution started at 20 °C and finished at 29 °C. Thus the denaturation temperature (mid-point, Tm) of the PBT skin collagen was estimated as 24.5 °C.

Denaturation curve of PBT skin collagen estimated from specific rotation
Peptide mapping
Peptide mapping was performed to compare the primary structure of PBT skin collagen with calf and salmon skin collagen. The electrophoretic patterns of lysyl endopeptidase-digested PBT, calf, and salmon skin collagen were observed on a 7.5% denaturing polyacrylamide gel. As shown in Fig. 4, the electrophoretic pattern of PBT skin collagen was similar to those of calf and salmon skin collagen, indicating that the cleavage site of PBT skin collagen by lysyl endopeptidase was almost identical to those of calf and salmon skin collagen.

Peptide maps of lysyl endopeptidase digests of PBT, calf, and salmon skin collagens. M.W. Marker: molecular weight marker
Cross-reactivity of PBT type I skin collagen with salmon collagen antibody
To compare the partial sequences and higher-order structure of PBT skin collagen with calf, and salmon skin collagen, direct, sandwich, and inhibition ELISA were established. We examined cross-reactivity between the isolated PBT skin collagen and IgG-purified guinea pig antibody against salmon type I collagen. As shown in Fig. 4, the calibration ranges established using direct, sandwich, and inhibition ELISA were 10–1000, 10–10,000, and 10–100,000 ng/mL, respectively. In the direct ELISA, the PBT skin collagen reacted with the antibody against salmon type I collagen, but the reactivity was almost similar to that of calf collagen (Fig. 5a). In addition, the PBT skin collagen was not detected in sandwich ELISA, suggesting that its epitope structure differed from that of salmon collagen (Fig. 5b). The PBT collagen recognized the antibody in inhibition ELISA, although the reactivity to the antibody was appreciably weaker compared to that of salmon collagen (Fig. 5c). The difference in cross-reactivity of PBT and salmon collagen might reflect the variations in epitope recognition of the antibody.

Determination of the antibody reactivities of PBT, calf, and salmon collagen. a Direct ELISA, b sandwich ELISA, and c inhibition ELISA. An IgG-purified guinea pig antibody against salmon type I collagen was used. All ELISA were performed in triplicate and the data were expressed as the mean value
FTIR spectra of PBT skin collagen
Figure 6 shows the FTIR spectra of PBT and calf skin collagen. The spectra of PBT skin collagen were roughly similar to those of calf collagen. The spectra of PBT dispersions demonstrated a characteristic pattern reflecting the amide I band at 1657 cm− 1, the amide II band at 1553 cm− 1, and amide III band at 1241 cm− 1, derived from C=O stretching, N–H bending vibrations, and C–H stretching (Payne and Veis, 1988), respectively. The amide I band, which is associated with the secondary structure of the protein, and the amide III band demonstrated the existence of a helical structure (Muyonga et al., 2004, 2004). These results suggest the existence of helical arrangements in the extracted PBT collagen.

FTIR spectra of PBT skin collagen
Discussion
In this study, we isolated and characterized certain properties of PBT skin collagen. The PBT skin collagen was composed of two α chains (α1 and α2) and one β chain. This result is similar to previous reports on collagen characteristics of other fish species (Tan and Chang 2018; Muyonga et al., 2004; Yata et al., 2001). UV-vis and FTIR spectra of PBT skin collagen resembled that of type I collagen reported previously. All these data suggest that the isolated collagen is a typical type I collagen. In the present study, we did not perform proximate analysis of fish skin during the isolation process. The possible differences in the yield obtained during the isolation process between these species are a limitation of this study.
The denaturation temperature of the PBT skin collagen was lower than that of other fish collagen. The thermal denaturation temperature of collagen is related to the proline and hydroxyproline content (Wong, 1989). The Pro and Hyp content in PBT skin type I collagen were 10.5 and 6.4%, respectively; the ratio of Pro to Hyp in PBT is higher than that in salmon (Arnesen and Gildberg, 2007), big eye snapper (Kittiphattanabawon et al., 2005), and skate (Hwang et al., 2007). However, the thermal denaturation temperature of PBT skin collagen was lower than that of salmon (28.7 °C), torafugu, and skate (28.8 °C).
Previous studies have revealed the primary structure of type I and II procollagen α1 chain in some fishes (Saito et al., 2001; Hwang et al., 2006; Zhang et al., 2016). We cloned the cDNA for PBT procollagen α1 (I) (Tanaka et al., 2014) and predicted that the PBT procollagen α1 (I) might contain high numbers of Gly-Gly sequences (Gly-Gly and Gly-Gly-Gly) in the triple-helical region. The number of Gly-Gly sequences in PBT procollagen α1 (I) was 14, whereas the number in zebrafish, rainbow trout, and torafugu were 4, 22, and 11, respectively. Since Gly is the smallest amino acid, the Gly-Gly sequence likely contributes to the partial skew in the triple helix structure and the decrease in thermal stability. While the PBT procollagen α1 (I) contains a high number of Gly-Gly sequence, it is not the highest among fish procollagen α1 (I) reported previously. Thus, further rationalization for the low thermal stability of PBT skin collagen is required. In addition, two Ser residues (1253 and 1270) that play a crucial role in the interactions of the procollagen α chains (Dion and Myers, 1987) were not found in the C-terminal region of the PBT procollagen α1 (I) chain. This indicated that PBT collagen might easily accrue distortion in its protein structure, which might contribute to its low denaturation temperature. PBT possesses delicate skin, which renders handling difficult during rearing this species. The primary structure of the PBT skin collagen could possibly explain the sensitive nature of its skin.
Tryptophan and phenylalanine are not present in the PBT collagen and the tyrosine content was 0.35%. Because, it is generally considered that most proteins that absorb at 280 nm of the UV-Vis spectra contain tyrosine, tryptophan, and phenylalanine, the absorption peak at 280 nm was weak. The major peak at 238 nm was slightly different from the skin collagen of largefin longbarbel (Zhang et al., 2009) at 232 nm and collagen of abalone gastropod muscle (Dong et al., 2012) at 233 nm. These differences might be due to differences in amino acid content between PBT collagen and other collagen.
The electrophoretic patterns of lysyl endopeptidase-digested PBT was similar to those of calf and salmon skin collagen as well as the electrophoretic pattern for acid-soluble collagen. Therefore, the primary structure of PBT skin collagen, including the cleavage site by lysyl endopeptidase, was almost identical to that of calf and salmon skin collagen. However, the cross-reactivity of PBT type I skin collagen with salmon collagen antibody was weak. The difference in cross-reactivity of PBT and salmon collagen might reflect the variations in epitope recognition of the antibody. These results suggest that although the primary structure of collagen type I is highly conserved in animal species, the partial sequences that include the epitope structure differ significantly. An antibody against PBT collagen is required for more accurate characterization of tuna collagen.
Further studies
Most fish collagens are composed of two α1 and one α2 chains (Gómez-Guillén et al., 2002; Muyonga et al., 2004). Piez (1965) reported that cod skin collagen has three variants of α chains (α1, α2, and α3) that differ in amino acid composition. Subsequently, the α3 chain was identified in collagen of other fish skin. Although the PBT skin collagen may contain the α3 chain, its presence was not determined using ion exchange chromatography in this study. Therefore, further studies are required to elucidate this point.
In the present study, we did not calculate the extraction efficiency of skin collagen halfway during the extraction process. However, this efficiency will be calculated by determining the hydroxyproline content in the sample in our next study. In addition, the proximate analysis of fish skin and the yield during the isolation process was not performed. The differences in the yield obtained during the isolation process between these animal species are a limitation of this study.
In addition, type I collagen has been identified as cross-reactive allergen for fish allergies (Hamada et al., 2001). Although the difference in cross-reactivity of PBT and salmon collagen was showed in this study, Kobayashi et al. (2016) clarified that pooled serum obtained from patients with fish collagen-specific allergies exhibited IgE reactivity to extracts from Atlantic salmon (Salmo salar) and yellowfin tuna (Thunnus albacares) by direct and inhibition ELISA. The cross-reactivity of bluefin tuna collagen with salmon collagen antibody provided information relevant for structural studies. Therefore, epitope recognition by anti-collagen antibody might differ among tuna species. However, further studies are required to understand its structural integrity.
Conclusion
In summary, the PBT skin collagen is composed of two α chains (α1 and α2) and one β chain. The PBT collagen has low denaturation temperature, although it is rich in proline and hydroxyproline. The primary structure of PBT skin collagen was approximately identical to that of calf and salmon skin collagen; however, it varied from the others with respect to epitope recognition of the antibody against salmon type I collagen. Further studies are required for understanding the specific primary or higher-order structure of PBT collagen.
Abbreviations
- BSE:
-
Bovine spongiform encephalopathy
- ELISA:
-
Enzyme-linked immunosorbent assays
- FMD:
-
Foot and mouth disease
- PBS:
-
Phosphate-buffered saline
- PBT:
-
Pacific bluefin tuna
- SDS-PAGE:
-
Sulfate-polyacrylamide gel electrophoresis
- TSE:
-
Transmissible spongiform encephalopathy
References
Arnesen JA, Gildberg A. Extraction and characterisation of gelatine from Atlantic salmon (Salmo salar) skin. Bioresour Technol. 2007;98:53–7.
Dion AS, Myers JC. COOH-terminal propeptides of the major human procollagens: structural, functional and genetic comparisons. J Mol Biol. 1987;193:127–43.
Dong X, Yuan Q, Qi H, Yang J, Zhu B, Zhou D, Murata Y, Ye W. Isolation and characterization of pepsin-soluble collagen from abalone (Haliotis discus hannai) gastropod muscle part II. Food Sci Technol Res. 2012;18:271–8.
Giraud-Guille MM, Besseau L, Chopin C, Durand P, Herbage D. Structural aspects of fish skin collagen which forms ordered arrays via liquid crystalline states. Biomaterials. 2000;21:899–906.
Gómez-Guillén MC, Turnay J, Fernández-Dıaz MD, Ulmo N, Lizarbe MA, Montero P. Structural and physical properties of gelatin extracted from different marine species: a comparative study. Food Hydrocoll. 2002;16:25–34.
Gordon MK, Hahn RA. Collagens. Cell Tissue Res. 2010;339:247–57.
Hamada Y, Nagashima Y, Shiomi K. Identification of collagen as a new fish allergen. Biosci Biotechnol Biochem. 2001;65:285–91.
Han SH, Uzawa Y, Moriyama T, Kawamura Y. Effect of collagen and collagen peptides from bluefin tuna abdominal skin on cancer cells. Health. 2011;3:129–34.
Hwang JH, Mizuta S, Yokoyama Y, Yoshinaka R. Purification and characterization of molecular species of collagen in the skin of skate (Raja kenojei). Food Chem. 2007;100:921–5.
Hwang JH, Yokoyama Y, Mizuta S, Yoshinaka R. cDNA cloning and characterization of type I procollagen α1 chain in the skate Raja Kenojei. Comp Biochem Physiol B: Biochem Mol Biol. 2006;144:1–10.
Ishibashi Y, Honryo T, Saida K, Hagiwara A, Miyashita S, Sawada Y, Okada T, Kurata M. Artificial lighting prevents high night-time mortality of juvenile Pacific bluefin tuna, Thunnus orientalis, caused by poor scotopic vision. Aquaculture. 2009;293:157–63.
Kaji T, Tanaka M, Takahashi Y, Oka M, Ishibashi N. Preliminary observations on development of Pacific bluefin tuna Thunnus thynnus (Scombridae) larvae reared in the laboratory, with special reference to the digestive system. Mar Freshw Res. 1996;47:261–9.
Kittiphattanabawon P, Benjakul S, Visessanguan W, Nagai T, Tanaka M. Characterisation of acid-soluble collagen from skin and bone of bigeye snapper (Priacanthus tayenus). Food Chem. 2005;89:363–72.
Kobayashi Y, Huge J, Imamura S, Hamada-Sato N. Study of the cross-reactivity of fish allergens based on a questionnaire and blood testing. Allergol Int. 2016;65:272–9.
Laemmli UK. Cleavage of structural proteins during assembly of head bacterriophage T4. Nature. 1970;277:680–5.
Miyashita S, Kato K, Sawada Y, Murata O, Ishitani Y, Shimizu K, Yamamoto S, Kumai H. Development of digestive system and digestive enzyme activity of larval and juvenile bluefin tuna, Thunnus thynnus, reared in the laboratory. Aquac Sci. 1998;46:111–20.
Miyashita S, Sawada Y, Okada T, Murata O, Kumai H. Morphological development and growth of laboratory-reared larval and juvenile Thunnus thynnus (Pisces:Scombridae). Fish Bull. 2001;99:601–16.
Muyonga JH, Cole CGB, Doudu KG. Charaterisation of acid soluble collagen from skins of young and adult nile perch (Lates niloticus). Food Chem. 2004;85:81–9.
Muyonga JH, Cole CGB, Duodu KG. Fourier transform infrared (FTIR) spectroscopic study of acid soluble collagen and gelatin from skins and bones of young and adult Nile perch (Lates niloticus). Food Chem. 2004;86:325–32.
Nagai T, Ogawa T, Nakamura T, Ito T, Nakagawa H, Fujiki K, Nakao M, Yano T. Collagen of edible jellyfish exumbrella. J Sci Food Agric. 1999;79:855–8.
Nagai T, Suzuki N, Tanoue Y, Kai N, Nagashima T. Characterization of acid-soluble collagen from skins of surf smelt (Hypomesus pretiosus japonicus Brevoort). Food Nutr Sci. 2010;1:59–66.
Nalinanon S, Benjakul S, Visessanguan W, Kishimura H. Use of pepsine for collagen extraction from the skin of bigeye smapper (Priacanthus tayenus). Food Chem. 2007;104:591–601.
Nomura Y, Sakai H, Ishii Y, Shirai K. Preparation and some properties of type I collagen from fish scales. Biosci Biotechnol Biochem. 1996;60:2092–4.
Payne KJ, Veis A. Fourier transform IR spectroscopy of collagen and gelatin solutions: deconvolution of the amide I band for conformational studies. Biopolymers. 1988;27:1749–60.
Piez KA. Characterization of a collagen from codfish skin containing three chromatographically different α chains. Biochemistry. 1965;4:2590–6.
Ramshaw JA, Shah NK, Brodsky B. Gly-X-Y tripeptide frequencies in collagen: a context for host-guest triple-helical peptides. J Struct Biol. 1998;122:86–91.
Saito M, Takenouchi Y, Kunisaki N, Kimura S. Complete primary structure of rainbow trout type I collagen consisting of α1 (I) α2 (I) α3 (I) heterotrimers. FEBS J. 2001;268:2817–27.
Sawada Y, Okada T, Miyashita S, Murata O, Kumai H. Completion of the Pacific bluefin tuna Thunnus orientalis (Temminck et Schlegel) life cycle. Aquac Res. 2005;36:413–21.
Senaratne LS, Park PJ, Kim SK. Isolation and characterization of collagen from brown backed toadfish (Lagocephalus gloveri) skin. Bioresour Technol. 2006;97:191–7.
Takii K, Miyashita S, Seoka M, Tanaka Y, Kubo Y, Kumai H. Changes in chemical contents and enzyme activities during embryonic development of bluefin tuna. Fish Sci. 1997;63:1014–8.
Tan Y, Chang SK. Isolation and characterization of collagen extracted from channel catfish (Ictalurus punctatus) skin. Food Chem. 2018;242:147–55.
Tanaka T, Takahashi K, Adachi K, Ohta H, Yoshimura Y, Agawa Y, Zaima N, Moriyama T, Kawamura Y. Molecular cloning and expression profiling of procollagen α1 (I) of cultured Pacific bluefin tuna. Fish Sci. 2014;80:603–12.
Tanaka T, Takahashi K, Iwamoto N, Agawa Y, Sawada Y, Yoshimura Y, Zaima N, Moriyama T, Kawamura Y. Hepatoprotective action of dietary bluefin tuna skin proteins on CCl4-intoxicated mice. Fish Sci. 2012;78:911–21.
Tanaka Y, Gwak WS, Tanaka M, Sawada Y, Okada T, Miyashita S, Kumai H. Ontogenetic changes in RNA, DNA and protein contents of laboratory-reared Pacific bluefin tuna Thunnus orientalis. Fish Sci. 2007;73:378–84.
Wong DWS. Mechanism and theory in food chemistry. New York: Van Nostrand Reinhold; 1989.
Yata M, Yoshida C, Fujisawa S, Mizuta S, Yoshinaka R. Identification and characterization of molecular species of collagen in fish skin. J Food Sci. 2001;66:247–51.
Zhang M, Liu WT, Li GY. Isolation and characterization of collagens from the skin of largefin longbarbel catfish (Mystus Macropterus). Food Chem. 2009;115:826–31.
Zhang X, Azuma N, Hagihara S, Adachi S, Ura K, Takagi Y. Characterization of type I and II procollagen α1chain in Amur sturgeon (Acipenser schrenckii) and comparison of their gene expression. Gene. 2016;579:8–16.
Acknowledgements
The authors would like to thank the hatchery staff of the Fish Nursery Centers, Uragami Station, Kindai University for their assistance in this study.
Funding
This work was supported in part by a Grant-in-Aid for the Global COE Program from the Ministry of Education, Culture, Sports, Science, and Technology of Japan and the Sapporo Bioscience Foundation.
Availability of data and materials
All datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
Author information
Authors and Affiliations
Contributions
TT designed the study, collected the data, analyzed the data, and drafted the manuscript. KT, KT, MH, KY, and AB collected the data. TM designed the study and drafted the manuscript. YK designed the study and completed the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval
All experimental animals used in this study were maintained under a protocol approved by the Kindai University Animal Care and Use Committee.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Tanaka, T., Takahashi, K., Tsubaki, K. et al. Isolation and characterization of acid-soluble bluefin tuna (Thunnus orientalis) skin collagen. Fish Aquatic Sci 21, 7 (2018). https://doi.org/10.1186/s41240-018-0084-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41240-018-0084-1