
Abstract
N6-methyladenosine (m6A) is the most prevalent and well-characterized internal chemical modification in eukaryotic RNA, influencing gene expression and phenotypic changes by controlling RNA fate. Insulin-like growth factor-2 mRNA-binding proteins (IGF2BPs) preferentially function as m6A effector proteins, promoting stability and translation of m6A-modified RNAs. IGF2BPs, particularly IGF2BP1 and IGF2BP3, are widely recognized as oncofetal proteins predominantly expressed in cancer rather than normal tissues, playing a critical role in tumor initiation and progression. Consequently, IGF2BPs hold potential for clinical applications and serve as a good choice for targeted treatment strategies. In this review, we discuss the functions and mechanisms of IGF2BPs as m6A readers and explore the therapeutic potential of targeting IGF2BPs in human cancer.
Similar content being viewed by others
Background
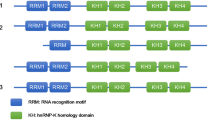
Insulin-like growth factor-2 mRNA-binding proteins (IGF2BPs, also known as IMPs or VICKZs), including IGF2BP1, IGF2BP2, and IGF2BP3, are a highly conserved family of RNA-binding proteins (RBPs) identified in humans since the late 20th century [1, 2]. They are named IGF2BPs due to their initial target, insulin-like growth factor II leader 3 mRNA [1]. In mammals, the three paralogues exhibit significant similarity in mass and structure, with molecular weight ranging from 58 to 66 kDa and amino acid sequence identities exceeding 56% [2]. This sequence conservation is most prominent in their domains, contributing to functional similarities (Fig. 1). IGF2BP proteins consist of two RNA recognition motif (RRM) domains (RRM-1 and RRM-2) and four heterogeneous nuclear ribonucleoprotein (hnRNP) K homology (KH) domains (KH-1 to KH-4) [1]. The C-terminal KH domains are responsible for recognizing and binding RNAs, while the N-terminal RRM domains likely contribute to the stability of RNA-protein complexes and interactions with other RBPs [3, 4]. With these domains, the three IGF2BP paralogues together target thousands of RNAs, of which approximately 92% are coding RNAs, with 55–70% co-targets. They play an essential role in mRNA processes such as transport, localization, stability, and translation [2, 5, 6]. Mechanically, IGF2BPs recruit their target transcripts to cytoplasmic messenger ribonucleoprotein particles (mRNPs) which subsequently condense into non-membrane-enclosed RNA granules, including processing bodies (PBs) and stress granules (SGs), thereby preventing mRNA decay and regulating translation-related events (Fig. 2A) [2, 7].

Features of human full-length IGF2BP protein family. Full-length IGF2BPs have a high identity in their domains, with two RRM (RNA recognition motif) and four KH (heterogeneous nuclear ribonucleoprotein K homology) domains. The RRM domain may mediate interactions with other RBPs (RNA-binding proteins) and the stability of RNA-protein complexes, and the KH domain is responsible for recognizing and binding RNA. The amino acid sequences of each domain in IGF2BPs are marked above them. Data from UniProt [130].

The post-transcriptional regulatory mechanisms of IGF2BPs. A IGF2BPs with other RBPs (RNA-binding proteins) regulate mRNA stability, transport, and translation by recruiting cytoplasmic transcripts into protective mRNPs (messenger ribonucleoprotein particles) including PBs (processing bodies) and SGs (stress granules). B IGF2BPs function as m6A readers to promote their stabilization and translation by selectively recognizing m6A-modified mRNAs under stress conditions in SGs or under normal conditions in PBs.
Additionally, IGF2BPs participate in various physiological and pathological processes throughout human life, from embryonic development to death, by controlling the expression of multitudinous genes. During embryogenesis, IGF2BPs are highly expressed and closely associated with cell differentiation and organ development [8]. Physiologically, all IGF2BPs, especially IGF2BP2, play a vital role in nervous system development and activity, as well as cell energy metabolism, including lipid metabolism, glycolysis, and aerobic respiration [8]. In contrast, IGF2BP1 and IGF2BP3 expression levels are negligible in adult tissues except for certain organs, while IGF2BP2 expression remains widespread [9]. However, gene expression analyses from Gene Expression Profiling Interactive Analysis (GEPIA) 2 (http://gepia2.cancer-pku.cn) [10] reveal significant re-expression of IGF2BP genes in most human cancer types. Moreover, mounting evidence suggests that aberrant expression of IGF2BPs contributes to various human tumors and participates in numerous malignant behaviors, including cell proliferation, invasion, stemness, apoptosis resistance, chemosensitivity, and immune escape. These effects are achieved by mediating several oncogene transcripts, such as KRAS, MYC, PTEN, and MDR1 [2, 8]. Consequently, IGF2BPs possess significant cancer-promoting properties, with IGF2BP1 and IGF2BP3 abnormally re-expressed in malignancies considered bona fide oncofetal proteins. IGF2BPs have regained widespread attention since their identification as a family of reader proteins that control RNA fate in an m6A-dependent manner [5]. Here, we mainly focus on recent advances in IGF2BPs’ roles as reader proteins and review the therapeutic potential of targeting IGF2BPs in cancers.
RNA m6A modification and IGF2BPs
Epigenetics is the study of heritable changes in gene expression or cellular phenotype resulting from factors such as lifestyle and environment, rather than alterations of DNA sequence [11]. This field includes DNA methylation, RNA modification, histone modification, histone variants, non-coding RNA (ncRNA), and more [12]. Epitranscriptomics refers to epigenetics in RNA, encompassing internal chemical modifications such as 7-methylguanosine (m7G), m1G, m6A, m1A, 5-methylcytosine (m5C), m3C, and pseudouridine (Ψ), which plays a critical role in post-transcriptional control [13]. Nevertheless, m6A, the methylation at the nitrogen-6 position of adenosine, is the best-studied RNA modification since the 1970s. It has been identified as the most widespread and dynamic modification in both messenger RNA (mRNA) and ncRNA in eukaryotes, including mammals [14, 15]. Although m6A modifications are ubiquitous in mRNAs, transcriptome-wide m6A mapping has uncovered that m6A sites predominantly cluster near stop codons and within 3’ untranslated regions (3’UTRs), situated in a specific consensus motif RR (m6A) CH ( R = G/A; H = U/A/C) [16, 17].
As an epigenetic regulatory mechanism, m6A participates in diverse biological processes in most organisms through controlling gene expression and cell fate at the post-transcriptional level. These processes include spermatogenesis, cell self-renewal and differentiation, embryonic development, immunoregulation, and stress responses [18, 19]. The whole-transcriptome m6A profile of human fetal tissue shows a positive correlation between m6A methylation and gene expression homeostasis [20]. Disruption of m6A homeostasis can lead to various human diseases, such as psychiatric disorders, osteoporosis, obesity, type 2 diabetes, viral infection, and cancer [21].
This homeostasis is maintained by its primary regulators, including methyltransferases, demethylases, and effector proteins, which are also metaphorically referred to as writers, erasers, and readers, respectively [22, 23]. Methyltransferases are multi-subunit complexes comprising methyltransferase-like 3/14 (METTL3/14), Wilms tumor 1-associated protein (WTAP), RNA binding motif protein 15/15B (RBM15/15B), zinc finger CCCH-type containing 13 (ZC3H13), Vir like m6A methyltransferase associated (VIRMA), and Cbl proto-oncogene like 1 (CBLL1). These complexes recognize RNA targets and catalyze m6A methylation. However, only METTL3 functions as the catalytic subunit, with METTL3/14 considered core subunits [24, 25]. Notably, a recent study reported another m6A methyltransferase, METTL16, which can attach m6A to the 3’UTR of MAT2A to regulate its RNA splicing efficiency, further adjusting the homeostasis of S-adenosylmethionine (SAM), an influential methyl donor for m6A [26]. In contrast, N6-methylated adenosine in RNA can be reversed by demethylases, including AlkB homolog 5 (ALKBH5) and fat mass and obesity-associated protein (FTO), with the former playing a primary role. Writers and erasers are localized in the nucleus, where m6A is installed or removed in a co-transcriptional manner [18, 22]. Once the m6A modification in RNA is complete, effector proteins play an essential role in m6A-mediated RNA metabolism, including RNA splicing, translocation, stability, decay, and translation, through the selective recognition and binding of m6A sites (Table 1) [18, 23, 24]. In general, writers and erasers jointly maintain m6A homeostasis and confer reversible and dynamic properties to m6A methylation, while readers play a key role in determining the fate of m6A-modified RNAs.
IGF2BPs, as a family of readers, preferentially recognize and selectively bind m6A-modified mRNAs to promote their stabilization and translation in an m6A-dependent manner under stress conditions in SGs or under normal conditions in PBs (Fig. 2B) [5]. The effect of IGF2BPs on target stability is likely exerted through recruiting co-factors such as ELAV-like RNA-binding protein 1 (ELAVL1; also known as HuR), matrin 3 (MATR3), and polyadenylate-binding protein 1 (PABPC1) [5]. Furthermore, numerous studies have shown that IGF2BPs play a carcinogenic role by stabilizing m6A-modified ncRNAs. The KH domains in IGF2BPs, particularly the KH3–4 di-domain, are confirmed to be responsible for m6A recognition and binding [5], which is consistent with the aforementioned KH functionality and has been corroborated in many subsequent studies. Remarkably, more than 80% of the thousands of IGF2BP targets contain at least one m6A-enriched region, and these regions spatially align with global m6A distribution [5], suggesting the significant role of IGF2BPs as reader proteins in the post-transcriptional regulation of m6A-methylated RNAs. However, the exact molecular mechanisms and functions of IGF2BPs in human cancers remain poorly understood.
IGF2BPs function as m6A readers in human cancer
Since 2018, increasing evidence has demonstrated that dysregulated IGF2BPs play vital roles as m6A effector proteins in various malignancies, as summarized in Table 2.
IGF2BPs in cancer stemness, survival and metastasis
Fundamental characteristics of cancers include immortalization, resistance to cell death, invasion, and metastasis.
IGF2BP1. Müller S et al. have reported that IGF2BP1 enhances cancer cell growth and invasiveness by increasing the expression of transcription factor SRF via m6A modification and obstructing microRNA-dependent decay of SRF mRNA [41]. In endometrial cancer, IGF2BP1 regulates cell cycle and promotes cell proliferation through employing the cofactor PABPC1 and stabilizing m6A-modified PEG10 mRNA [42].
IGF2BP2. IGF2BP2 maintains stemness and proliferation of pancreatic cancer cells by stabilizing long noncoding RNA (lncRNA) DANCR in an m6A-dependent manner [43]. LncRNA DIAPH1-AS1 is stabilized by IGF2BP2 via m6A modification that is mediated by WTAP, accelerating the growth and metastasis of nasopharyngeal carcinoma [44]. Increased MTA1 regulating colorectal cancer metastasis relies on FTO/IGF2BP2 regulated m6A methylation [45]. The formation of LINC00460/IGF2BP2/DHX9 complex reinforces HMGA1 mRNA stability in an m6A-dependent manner, promoting epithelial-mesenchymal transition (EMT), tumor growth, and metastasis in colorectal cancer [46]. METTL3 and IGF2BP2 control m6A of lncRNA PCAT6 and keep its stability. PCAT6 in turn intensifies the stability effects of IGF2BP2 on IGF1R mRNA, further promoting prostate cancer growth and bone metastasis [47]. ZNF677, which plays a tumor suppressor role in renal cell carcinoma, can be stabilized by METLL3/IGF2BP2-regulated m6A modification at the post-transcription level [48]. IGF2BP2 stabilizes MSX1 and JARID2 transcripts in an m6A-dependent manner, regulating cell migration and survival in colorectal cancer [49]. METTL14/IGF2BP2 prevents m6A-modified lncRNA ARHGAP5-AS1 degradation, playing an important part in hepatocellular carcinoma growth and metastasis [50]. IGF2BP2 functions as the m6A effector to stabilize E2F6 and E2F3, activating the Wnt/β-catenin pathway and facilitating liver tumor initiating cells (TICs) self-renewal and metastasis [51]. METTL3/IGF2BP2 regulates m6A modification of PLK1 and increases its stability and expression, leading to radio-resistance in pancreatic adenocarcinoma [52]. IGF2BP2 improves PRMT6 expression for leukemia stem cell (LSC) maintenance and acute myeloid leukemia development via stabilizing its transcript in an m6A-dependent form [53]. NFE2L2, upregulated by ALKBH5/IGF2BP2-mediated m6A methylation after transcription, inhibits ferroptosis of hypopharyngeal squamous cell carcinoma [54]. SHMT2 improves the m6A abundance of PPAT mRNA by offering methyl donor SAM, allowing IGF2BP2 to enhance the stability and expression of PPAT, regulating cell cycle to promote proliferation in renal cell carcinoma [55].
IGF2BP3. In colon cancer, IGF2BP3 boosts DNA replication and cell proliferation by binding to m6A-modified sites in CCND1 mRNA and preventing its degradation [56]. ACIN1 is stabilized through METTL3/IGF2BP3, boosting proliferation and migration of cervical cancer cells [57]. In hepatocellular carcinoma, lnc-CTHCC, stabilized by METTL3/IGF2BP1/IGF2BP3 via m6A modification, promotes tumor initiation and development by binding hnRNPK and activating YAP1 transcription [58]. LncRNA KCNMB2-AS1 that improves IGF2BP3 expression via sponging miR-130b-5p and miR-4294 is stabilized by IGF2BP3 in an m6A-mediated manner, synergistically accelerating cervical cancer cell growth [59]. TMBIM6, a downstream target of RBM15 and IGF2BP3, is involved in proliferation, invasion, migration, and apoptosis of laryngeal squamous cell carcinoma cells [60]. In clear cell renal cell carcinoma, the complex of IGF2BP3/lncRNA DMDRMR selectively interacts with their m6A-modified co-targets (including CDK4, COL6A1, LAMA5, and FN1) to promote cell proliferation [61]. In prostate cancer, hsa_circ_0003258 directly binds IGF2BP3 and enhances its function of stabilizing m6A-methylated HDAC4 transcript, contributing to EMT and metastasis by stimulating the MAPK signaling pathway [62]. In gastric cancer, IGF2BP3 physically binds and stabilizes PKMYT1 mRNA in an m6A-dependent manner, facilitating cell invasion and migration [63]. IGF2BP3 recognizes and reads m6A sites in KPNA2 mRNA to promote its stability and expression, activating tumor growth and metastasis in nasopharyngeal carcinoma [64]. In acute myeloid leukemia, IGF2BP3 is indispensable for leukemia cell survival by enhancing RCC2 expression through the interaction between IGF2BP3 and m6A sites in RCC2 mRNA [65].
IGF2BPs. In hepatocellular carcinoma cells and HeLa cells, knockdown of each IGF2BP can hinder MYC expression and tumor proliferation, colony formation, migration, and invasion by suppressing MYC mRNA stability and translation in an m6A-dependent manner [5]. In HEK293T and HeLa cells, reactive oxygen species (ROS) signaling improves m6A abundance by weakening the demethylase activity of ALKBH5, which withstands ROX-induced DNA damage and apoptosis through stabilization of m6A–modified FAAP20 and ATRX mRNA mediated by IGF2BP proteins [66]. IGF2BPs facilitate cell proliferation and migration in renal cell cancer by stabilizing S1PR3 mRNA in an m6A-dependent manner and activating the PI3K/AKT pathway [67].
IGF2BPs in drug resistance
IGF2BP1. In seminoma cells, cisplatin induces a significant elevation of METTL3 and m6A levels, and IGF2BP1 stabilizes TFAP2C gene in an m6A-dependent manner in response to the chemotherapy damage [68].
IGF2BP2. In colorectal carcinoma, m6A modification of SOX2 is installed by METTL3 and recognized by IGF2BP2, resulting in upregulated SOX2 that strengthens cell stemness, invasion property, and resistance to oxaliplatin [69]. In T-cell acute lymphoblastic leukemia, IGF2BP2 impels cell chemoresistance and survival via directly binding and stabilizing m6A-modified NOTCH1 mRNA [70]. In addition, METTL14-mediated MN1 methylation raises the stability and translation of MN1 transcript through the IGF2BP2-dependent pathway, prompting all-trans-retinoic acid resistance and progression in osteosarcoma [71].
IGF2BP3. In cervical and liver cancers, m6A in PDK4 transcript regulated by IGF2BP3 and METTL3 plays an important role in tumor growth and chemoresistance [72]. Overexpression of ABCB1 inducing multidrug resistance (MDR) desensitizes colon cancer cells to chemotherapy drugs with ABCB1 substrate specificity, resulting from enhanced stability of ABCB1 transcript via IGF2BP3 reading its m6A region [73]. In clear cell renal cell carcinoma, IGF2BP3 suppresses cancer sensitivity to CDK4/6 inhibitor palbociclib by stabilizing CDK4 mRNA in an m6A- dependent manner [61].
IGF2BPs in reprogramming of metabolism
The Warburg effect, a hallmark of cancer cells, describes their preference for obtaining energy through aerobic glycolysis instead of oxidative phosphorylation [74, 75]. Glutamine presents metabolic dysregulation in cancer and serves as a significant source of nitrogen and carbon for cancer development [76]. These processes support cancer progression directly or through enhancing crosstalks with the surrounding tumor microenvironment (TME), which includes immune cells, fibroblasts, endotheliocytes, and extracellular matrix [77]. Besides, autophagy, responsible for the degradation and recycling of dysfunctional organelles and proteins, plays dual roles in both promoting and suppressing cancer through adjusting cellular metabolism [78].
IGF2BP2. HK2 and GLUT1, the key factors of glucose metabolism, undergo m6A modification by METTL3 and are individually stabilized by IGF2BP2 and IGF2BP2/3 at the post-transcription level, activating the Warburg effect to promote colorectal cancer progression[79]. In colorectal cancer, IGF2BP2 stabilizes lncRNA ZFAS1 in an m6A-dependent manner, which activates OLA1 to reinforce adenosine triphosphate (ATP) hydrolysis and aerobic glycolysis [80]. IGF2BP2 also can enhance colorectal cancer cell glycolysis through the downstream MYC gene [81]. In papillary thyroid cancer, low-level FTO results in m6A modification in APOE mRNA, which is then read and stabilized by IGF2BP2, activating glycolysis through IL-6/JAK2/STAT3 signaling [82]. In acute myeloid leukemia, IGF2BP2 enhances the mRNA stability and translation initiation of m6A-containing targets GPT2, SLC1A5, and MYC by employing PABPC1 and eukaryotic translation initiation factor (eIF) complexes eIF4A, respectively, facilitating glutamine uptake and metabolism for tumor cell stemness and development [83]. In clear cell renal cell carcinoma, SIK2 downregulation promotes tumor progression by inhibiting autophagy, depending on FTO/IGF2BP2-m6A axis-mediated SIK2 mRNA stabilization [84].
IGF2BP3. IGF2BP3 can increase the expression of PDK4 by binding to the m6A site and enhancing its mRNA stability, thereby promoting glycolysis and ATP production in cervical and liver cancer cells [72]. In addition, IGF2BP3 and METTL3 improve the expression of transcription factor HDGF by directly recognizing and binding the m6A-modified HDGF, which transcriptionally accelerates the expression of GLUT4 and ENO2 to facilitate glycolysis in gastric cancer. This ultimately results in pro-tumorigenic activities, including cancer growth and liver metastasis [85].
IGF2BPs in tumor microenvironment
The TME comprises various cell types (including fibroblasts, endothelial cells, and immune cells) and noncellular components (including extracellular matrix, cytokines, and metabolites). The TME can be considered the “soil” in which cancer cells, the “seeds”, grow. The crosstalk between the two creates a complex ecosystem involved in multiple aspects of malignant development, including tumor growth, metastasis, drug resistance, angiogenesis, and immune escape.
IGF2BP2/3. In clear cell renal cell carcinoma, IGF2BP3 promotes extracellular matrix (ECM) deposition, including COL6A1, LAMA5, and FN1, by stabilizing their m6A-methylated transcripts [61]. As previously discussed, HDGF expression is fostered via the METTL3/IGF2BP3-HDGF mRNA axis in an m6A-dependent manner in gastric cancer. Intriguingly, while nuclear HDGF involves in glycolysis, secretory HDGF in the TME promotes tumor angiogenesis [85]. Another different mechanism of angiogenesis in gastric cancer has been revealed: the disruption of IGF2BP3 weakens the stability of m6A-enriched HIF1A, consequently inhibiting hypoxia-induced cell migration and angiogenesis [86]. In colon cancer, IGF2BP3 expression promotes angiogenesis by targeting m6A-methylated VEGF mRNA [56]. Beyond traditional tumor angiogenesis, cancer cells can form channels resembling blood vessels, which are responsible for supplying blood, nutrients, and oxygen. This process is termed vasculogenic mimicry [87]. In colorectal cancer, IGF2BP2 and IGF2BP3 can stimulate vasculogenic mimicry formation by separately stabilizing the transcripts of EphA2 and VEGF via interaction with their m6A sites. This subsequently activates downstream PI3K/AKT/mTOR and MAPK/ERK1/2 signaling pathways, promoting cell proliferation, migration, and invasion [88]. Additionally, the METTL3/IGF2BP3 axis reinforces the immune evasion capacity of breast cancer cells in the TME through increasing the stability of downstream target PD-L1 mRNA and the expression of its transmembrane protein [89].
To sum up, IGF2BPs function as oncogenes promoting various aspects of tumor properties, including tumorigenesis, survival, metastasis, drug resistance, metabolic reprogramming, matrix deposition, angiogenesis, and immune evasion, by binding to m6A-containing RNAs and increasing their stability and/or translation. In addition to their role as RBPs, the identification of m6A readers means a deeper exploration of IGF2BPs. Further systematic studies are needed to fully elucidate the essence of IGF2BPs, including their effects on hallmarks across cancer types, alternative mechanisms, target selectivity, the roles of each domain, and more.
The value of IGF2BPs in clinical prediction
As discussed above, IGF2BPs typically paly a pro-carcinogenic role in human malignancies and are involved in numerous aspects of tumor characteristics. Evidence has shown that IGF2BPs are often aberrantly overexpressed and could serve as biomarkers for diagnosis and prognosis across various cancer types.
Tumor-associated antigens (TAAs), such as carcinoma embryonic antigen (CEA), alpha-fetoprotein (AFP), and carbohydrate antigen 125 (CA125), are widely used in clinical tumor diagnosis. IGF2BPs, as embryonic proteins, have been found to be valuable for early screening and diagnosis of tumors. In colorectal cancer, IGF2BP1 and IGF2BP3 might act as potential biomarkers for screening high-risk groups and cancer patients [90]. In renal cell carcinoma, IGF2BP3 serves as an indicator of high risk for metastasis and informs systemic treatment decisions [91]. Yang et al. suggested that IGF2BP3 and IGF2BP2 could be considered specific genes of triple-negative breast cancer, the molecular subtype with the poorest prognosis, which would aid in more precise diagnosis and treatment for breast cancer patients [92]. Gong Y et al. reviewed IGF2BP3 as a competent molecular marker for diagnosis in a majority of malignancies [93].
While numerous studies have highlighted the potential of histological detection of IGF2BP proteins for tumor diagnosis, body fluid tests offer a more convenient and minimally invasive approach for patients. The detection of serum autoantibodies to IGF2BP1 in esophageal squamous cell carcinoma [94] and IGF2BP1/3 in colon cancer [95] contributes to identifying patients with cancer, and when combined with other TAAs, achieves greater sensitivity and specificity for diagnosis. Moreover, autoantibodies to IGF2BP2 were detected in the serum of some hepatocellular carcinoma patients but not in precancerous lesions like chronic hepatitis or liver cirrhosis hepatitis [96]. Zhang J et al. expanded autoantibody testing of IGF2BP2/3 to more cancers and believed that they could become valuable biomarkers for pan-cancer clinical applications [97]. Given the broad expression and oncogenic roles of IGF2BPs in cancers, their detection could be beneficial for the diagnosis of other tumors not mentioned above.
In addition, IGF2BPs have been suggested for tumor prognosis assessment. IGF2BP3 might be a pan-cancer oncogene, as its overexpression is associated with poor patient survival in various cancers, including kidney renal clear cell carcinoma, kidney renal papillary cell carcinoma, brain lower-grade glioma, and more [98]. Moreover, IGF2BP3 is upregulated and its high expression conveys tumor progression and worse prognosis in patients with lung adenocarcinoma [99], esophageal cancer [100], gastric cancer [63], pancreatic adenocarcinoma [101], papillary renal cell carcinoma [102], colon cancer [56], clear cell renal cell carcinoma [61], bladder cancer [103], prostate cancer [104], nasopharyngeal carcinoma [64], laryngeal squamous cell cancer [60], melanoma [105], glioma [106], acute myelocytic leukemia [65], among others. IGF2BP1 has been identified as an unfavorable prognostic indicator in tumors such as lung adenocarcinoma [107], endometrial cancer [42], gastric cancer [108], hepatocellular carcinoma, and serous ovarian cancer [41]. Upregulated IGF2BP2 is significantly correlated with weak prognosis, providing a hopeful predictor for pancreatic cancer [43], colorectal cancer [69, 80, 81], osteosarcoma [71], and lung adenocarcinoma [99].
The immune microenvironment and immunotherapy have become research hotspots, especially since the discovery of immune checkpoint therapy earned a Nobel Prize. The mechanism by which IGF2BP3 promotes PD-L1 expression has been stated above. Both proteins are highly expressed and positively correlated in breast cancer, especially in HER2-enriched and triple-negative breast cancer, the more aggressive subtypes [89]. IGF2BP3 also exerts a significant impact on PD-L1 expression in bladder cancer [109]. Besides, IGF2BPs have been found to be associated with immunomodulators and immune infiltration levels in diverse cancers [99, 101, 105, 107, 110,111,112,113,114]. Consequently, IGF2BPs may provide guidance for screening patients responsive to immunotherapy and could become potential targets for boosting immunotherapy response.
In conclusion, IGF2BPs show promise as broad-spectrum tumor markers for the diagnosis and prediction of tumors, and their performance improves when combined with other TAAs. However, IGF2BPs only provide diagnostic clues and lack tissue and organ specificity. Additionally, IGF2BP2 is expressed in multiple normal organs, which reduces tumor specificity. As a result, a baseline value must be established to evaluate normal or abnormal expression. Further research is needed to determine whether the expression of IGF2BPs is related to disease severity and tumor stage, and whether they can monitor treatment efficacy, recurrence, and metastasis.
The therapeutic potential of targeting IGF2BPs
The therapeutic potential of targeting IGF2BPs is gaining attention due to advances in understanding their molecular structures and carcinogenic mechanisms. We summarized all the current agents and strategies for targeting IGF2BPs (Fig. 3).

Potential therapeutic strategies of targeting IGF2BPs in cancer. A The structure of small molecule inhibitors for IGF2BP1/2. B Oligonucleotide sequences designed according to IGF2BP1-RNA binding sites. C Natural plant extracts that effectively inhibit IGF2BP3 expression. d-ICD: derivative of isocorydine. D Torin1 (mTORC2 kinase inhibitor) and AZD0530 (Src kinase inhibitor) impede IGF2BP1-RNA interaction by hindering IGF2BP1 phosphorylation.
Small molecule inhibitors
Small molecule inhibitors have emerged as a promising strategy for cancer treatment, with many small molecule drugs already used in clinical and showing encouraging results. Recently, a specific small molecule inhibitor, JX5 (Kd value = 93.2 ± 3.9 µM), has been identified for IGF2BP2 through high-throughput virtual screening based on the three-dimensional structure of IGF2BP2 [70]. It inhibits IGF2BP2 activity by physically docking with its KH3-4 di-domains and has shown effectiveness in hampering T-cell acute lymphoblastic leukemia cell amplification and marrow/spleen infiltration with minimal toxicities [70]. Furthermore, IGF2BP2 inhibitors, CWI1-1 and CWI1-2, have also been identified, with CWI1-2 (IC50 = 203.1-781.6 nM in leukemia cell lines) demonstrating stronger anti-tumor effects on acute myelocytic leukemia in vitro and in vivo [83]. Importantly, CWI1-2 has shown synergistic effects when combined with conventional acute myelocytic leukemia chemotherapy [83]. Several compounds, belonging to either benzamidobenzoic acid or ureidothiophene, have been found to selectively restrain the interaction of IGF2BP2 with its target RNAs [115]. Among them, compound 4 (IC50 = 18.2–35.5 µM), compound 6 (IC50 = 42.9–52.6 µM), and compound 9 (IC50 = 24.9–39.8 µM) demonstrate higher activity and effectively kill IGF2BP2-containing colorectal cancer and hepatocellular carcinoma cells in vitro and in vivo [115]. Small molecule allosteric inhibitor BTYNB (IC50 = 2.3 µM in ES-2, = 3.6 µM in IGROV-1, = 4.5 µM in SK-MEL2) has been identified to hinder the proliferation of IGF2BP1-overexpressing ovarian cancer and melanoma cells by altering the functional site of IGF2BP1 and decreasing the expression of downstream oncogenes, including MYC, BTRC, and eEF2 [116]. Intriguingly, BTYNB has been found to reduce both IGF2BP1 and IGF2BP2 protein levels [83]. The compound 2-[(8-bromo-5-methyl-5 H-[1,2,4]triazino[5,6-b]indol-3-yl)thio]-N-(1-phenylethyl)acetamide (C20H18BrN5OS, ChemBridge ID: 6896009) has been regarded as a lead IGF2BP1 inhibitor to obstruct the binding of IGF2BP1 to MYC mRNA and can obviously restrain IGF2BP1-containing ovarian cancer cell proliferation [117]. In non-small-cell lung carcinoma and ovarian clear cell carcinoma, another IGF2BP1 small molecule inhibitor compound 7773 competitively binds to a hydrophobic surface around KH3-4 domains and blocks their binding activity to target transcripts, which inhibits cell migration and growth without any toxicity [118].
Unfortunately, no specific small molecule inhibitors have been discovered for IGF2BP3 yet. However, given the high similarity among IGF2BP paralogues, especially in their structural domains, it is possible that some small molecule inhibitors targeting one IGF2BP could also target the other two proteins, potentially having therapeutic effects on the tumors they drive. For example, BTYNB has already been verified to target both IGF2BP1 and IGF2BP2, inhibiting leukemia initiation and development [115]. Further studies are warranted to explore this possibility and to develop more specific inhibitors targeting IGF2BPs for cancer therapy.
Oligonucleotides
Oligonucleotides, including antisense oligonucleotides (ASOs), small interfering RNAs (siRNAs), microRNAs (miRNAs), and aptamers, hold promise as cancer therapeutics by regulating gene expression and product function [119].
IGF2BPs interact with specific RNA sites to promote gene expression, and ASOs can be designed to target these specific sequences and disrupt these interactions. For example, ASOs CRD-ODN4 [120] and DD4/DD7 [121] have been designed to target the specific sequences recognized by IGF2BP1 on MYC and CD44 mRNA, respectively, and have been shown to efficiently reduce intracellular MYC and CD44 levels.
In addition to inhibiting gene expression through complementary base pairing, oligonucleotides can also interact with and block proteins through their three-dimensional secondary structures. The structured oligonucleotide S1, containing two distinct stem loops, has been shown to target IGF2BP1 domains and interferes with IGF2BP1-GLI1 mRNA interaction [122]. Moreover, 2’-O-methyl derivatives of these oligonucleotides can lower mRNA levels of corresponding oncogenes in diverse cancer cells [120,121,122].
siRNAs are principal components of the RNA-induced silencing complex (RISC), which mediates gene silencing of complementary target transcripts. siRNA drugs have been approved by the Food and Drug Administration (FDA) for the treatment of clinical diseases [119]. There is substantial evidence that siRNA can be used to specifically knock down IGF2BPs in experiments and effectively suppress IGF2BP-mediated malignant phenotypes in vitro and in vivo.
However, there are significant challenges associated with oligonucleotide therapeutics, including the development of efficient delivery systems, the potential for off-target effects, and unknown toxicities [119]. Continued research is needed to address these challenges and optimize oligonucleotide-based therapies targeting IGF2BPs for cancer treatment.
Plant extracts
Plant extracts and their derivatives have been widely applied to clinical antitumor strategies, with well-known examples including vinca alkaloids, camptothecins, taxanes, anthracyclines, and podophyllotoxin [123]. Some plant extracts, such as isoliquiritigenin [124], berberine [125], and an isocorydine derivative (d-ICD) [126] have been shown to decrease IGF2BP3 expression, subsequently inhibiting the malignant behavior of cancers.
Targeting regulators of IGF2BPs
Targeting the dysregulated upstream regulators of IGF2BPs, including their transcription factors, epigenetic ncRNAs, post-translational modification, and E3 ligase-mediated ubiquitination, is another potential treatment strategy for IGF2BP-driven cancer. For example, IGF2BP1 phosphorylation at Ser181 and Tyr396, catalyzed by mTORC2 and Src kinase, respectively, is involved in the interaction and post-transcriptional regulation of target transcripts such as MYC [127,128,129]. And mTORC2 kinase inhibitor Torin1 and Src kinase inhibitor AZD0530 have been identified to synergistically inhibit the growth of IGF2BP1-expressing cancer cells in vitro and in vivo by disrupting IGF2BP1 phosphorylation [129].
Given the significant tumor-promoting abilities of IGF2BPs in various malignancies, targeting dysregulated IGF2BPs is an appealing approach for cancer treatment. However, the clinical application of agents targeting IGF2BPs is still in its early stages. Several challenges need to be addressed, including improving the understanding of IGF2BPs’ structures and carcinogenic mechanisms, selecting suitable drug candidates, conducting credible preclinical studies, and performing rigorous clinical trials.
Conclusion
As the most widespread and energetic RNA internal modification, m6A is involved in almost every aspect of RNA metabolism to control gene expression and cell phenotype. The imbalance of m6A modification caused by maladjusted m6A regulators, including IGF2BPs, has become a driver of tumor initiation and progression. Since the identification of IGF2BPs as a family of reader proteins, substantial studies that implicate the role of IGF2BPs in diverse malignancies have emerged, providing novel insights into the carcinogenic mechanisms of IGF2BPs. IGF2BPs play a crucial role in m6A-mediated post-transcriptional regulation. They preferentially recognize and bind m6A-modified target RNAs to promote their stabilization and translation in an m6A-dependent manner. IGF2BPs also function in RNA processes independently of m6A modification. However, the mechanism by which IGF2BPs affect RNA metabolism remains unclear, representing further research direction. Consequently, dysregulated IGF2BPs accumulate oncogenic products to promote various malignant phenotypes, including proliferation, resistance to cell death, metastasis, drug resistance, metabolism reprogramming, angiogenesis, and immune escape.
IGF2BPs, particularly IGF2BP1 and IGF2BP3, are oncofetal proteins and exhibit significantly up-regulated expression and prognostic correlation in various human cancers. Furthermore, IGF2BPs could serve as broad-spectrum tumor markers for early screening and prognosis evaluation. Combining these markers with others in specific cancers may enhance the accuracy of diagnosis, outcome prediction, and treatment guidance.
Understanding the underlying mechanisms of IGF2BPs in tumorigenic processes and their molecular structures is essential for developing therapeutic strategies. Small molecule inhibitors targeting IGF2BP1 and IGF2BP2 have been developed based on their KH3-4 structural domain, the core region that recognizes and binds RNA. However, further in vivo efficacy and toxicity studies are highly warranted. In addition, targeting IGF2BP1 and IGF2BP3 could be more specific for tumors and safer for patients, given their absence in normal tissues and high levels in many cancers.
In summary, IGF2BPs play a wide range of roles in cancer biology through post-transcriptional regulation of gene expression in both m6A-dependent and m6A-independent forms. They have shown potential as new biomarkers for tumor early screening, diagnosis, and prognosis. Current cancer therapies targeting IGF2BPs are still in their infancy. Further research into the molecular structure of IGF2BPs and their regulatory mechanisms in cancer is necessary to develop novel effective cancer therapies.
Data Availability
Not applicable.
Change history
21 July 2023
A Correction to this paper has been published: https://doi.org/10.1186/s40364-023-00512-6
Abbreviations
- m6A:
-
N6-methyladenosine.
- IGF2BPs:
-
Insulin-like growth factor-2 mRNA-binding proteins
- RBPs:
-
RNA-binding proteins
- RRM:
-
RNA recognition motif
- hnRNP:
-
Heterogeneous nuclear ribonucleoprotein
- KH:
-
Heterogeneous nuclear ribonucleoprotein K homology domain
- mRNPs:
-
Messenger ribonucleoprotein particles
- PBs:
-
Processing bodies
- SGs:
-
Stress granules
- ncRNA:
-
Non-coding RNA
- m7G:
-
7-methylguanosine
- m5C:
-
5-methylcytosine
- Ψ:
-
Pseudouridine
- mRNA:
-
Messenger RNA
- UTRs:
-
Untranslated regions
- METTL3/14:
-
Methyltransferase-like 3/14
- WTAP:
-
Wilms tumor 1 associated protein
- RBM15/15B:
-
RNA binding motif protein 15/15B
- ZC3H13:
-
Zinc finger CCCH-type containing 13
- VIRMA:
-
Vir like m6A methyltransferase associated
- CBLL1:
-
Cbl proto-oncogene like 1
- SAM:
-
S-adenosylmethionine
- ALKBH5:
-
AlkB homolog 5
- FTO:
-
Fat mass and obesity-associated protein
- YTHDC1/2:
-
YTH domain-containing protein 1/2
- YTHDF1/2/3:
-
YTH domain-containing family protein 1/2/3
- eIF3:
-
Eukaryotic translation initiation factor 3
- FMR1:
-
Fragile X messenger ribonucleoprotein 1
- HNRNPC/G/A2B1:
-
Heterogeneous nuclear ribonucleoprotein C/G/A2B1
- ELAVL1:
-
ELAV-like RNA-binding protein 1
- MATR3:
-
Matrin 3
- PABPC1:
-
Polyadenylate-binding protein 1
- lncRNA:
-
Long noncoding RNA
- ROS:
-
Reactive oxygen species
- TME:
-
Tumor microenvironment
- ATP:
-
Adenosine triphosphate
- ECM:
-
Extracellular matrix
- TAA:
-
Tumor-associated antigen
- ASO:
-
Antisense oligonucleotide
- siRNA:
-
Small interfering RNA
- CDS:
-
Coding sequence region
References
Nielsen J, Christiansen J, Lykke-Andersen J, Johnsen AH, Wewer UM, Nielsen FC. A family of insulin-like growth factor II mRNA-binding proteins represses translation in late development. Mol Cell Biol. 1999;19(2):1262–70.
Bell JL, Wächter K, Mühleck B, Pazaitis N, Köhn M, Lederer M, et al. Insulin-like growth factor 2 mRNA-binding proteins (IGF2BPs): post-transcriptional drivers of cancer progression? Cell Mol Life Sci. 2013;70(15):2657–75. https://doi.org/10.1007/s00018-012-1186-z.
Wächter K, Köhn M, Stöhr N, Hüttelmaier S. Subcellular localization and RNP formation of IGF2BPs (IGF2 mRNA-binding proteins) is modulated by distinct RNA-binding domains. Biol Chem. 2013;394(8):1077–90. https://doi.org/10.1515/hsz-2013-0111.
Nielsen J, Kristensen MA, Willemoës M, Nielsen FC, Christiansen J. Sequential dimerization of human zipcode-binding protein IMP1 on RNA: a cooperative mechanism providing RNP stability. Nucleic Acids Res. 2004;32(14):4368–76.
Huang H, Weng H, Sun W, Qin X, Shi H, Wu H, et al. Recognition of RNA N(6)-methyladenosine by IGF2BP proteins enhances mRNA stability and translation. Nat Cell Biol. 2018;20(3):285–95. https://doi.org/10.1038/s41556-018-0045-z.
Lederer M, Bley N, Schleifer C, Hüttelmaier S. The role of the oncofetal IGF2 mRNA-binding protein 3 (IGF2BP3) in cancer. Semin Cancer Biol. 2014;29:3–12. https://doi.org/10.1016/j.semcancer.2014.07.006.
Ivanov P, Kedersha N, Anderson P. Stress granules and Processing Bodies in Translational Control. Cold Spring Harb Perspect Biol. 2019;11(5):a032813. https://doi.org/10.1101/cshperspect.a032813.
Du Q-Y, Zhu Z-M, Pei D-S. The biological function of IGF2BPs and their role in tumorigenesis. Invest New Drugs. 2021;39(6):1682–93. https://doi.org/10.1007/s10637-021-01148-9.
Hammer NA, Hansen TvO, Byskov AG, Rajpert-De Meyts E, Grøndahl ML, Bredkjaer HE, et al. Expression of IGF-II mRNA-binding proteins (IMPs) in gonads and testicular cancer. Reproduction. 2005;130(2):203–12.
Li C, Tang Z, Zhang W, Ye Z, Liu F. GEPIA2021: integrating multiple deconvolution-based analysis into GEPIA. Nucleic Acids Res. 2021;49(W1):W242–W6. https://doi.org/10.1093/nar/gkab418.
Goldberg AD, Allis CD, Bernstein E. Epigenetics: a landscape takes shape. Cell. 2007;128(4):635–8.
Peixoto P, Cartron P-F, Serandour AA, Hervouet E. From 1957 to nowadays: a brief history of Epigenetics. Int J Mol Sci. 2020;21(20):7571. https://doi.org/10.3390/ijms21207571.
Barbieri I, Kouzarides T. Role of RNA modifications in cancer. Nat Rev Cancer. 2020;20(6):303–22. https://doi.org/10.1038/s41568-020-0253-2.
Lan Q, Liu PY, Haase J, Bell JL, Hüttelmaier S, Liu T. The critical role of RNA m6A methylation in Cancer. Cancer Res. 2019;79(7):1285–92. https://doi.org/10.1158/0008-5472.CAN-18-2965.
Perry RP, Kelley DE. Existence of methylated messenger RNA in mouse L cells. Cell. 1974;1(1):37–42. https://doi.org/10.1016/0092-8674(74)90153-6.
Meyer KD, Saletore Y, Zumbo P, Elemento O, Mason CE, Jaffrey SR. Comprehensive analysis of mRNA methylation reveals enrichment in 3’ UTRs and near stop codons. Cell. 2012;149(7):1635–46. https://doi.org/10.1016/j.cell.2012.05.003.
Dominissini D, Moshitch-Moshkovitz S, Schwartz S, Salmon-Divon M, Ungar L, Osenberg S, et al. Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq. Nature. 2012;485(7397):201–6. https://doi.org/10.1038/nature11112.
Yang Y, Hsu PJ, Chen Y-S, Yang Y-G. Dynamic transcriptomic m6A decoration: writers, erasers, readers and functions in RNA metabolism. Cell Res. 2018;28(6):616–24. https://doi.org/10.1038/s41422-018-0040-8.
Yue Y, Liu J, He C. RNA N6-methyladenosine methylation in post-transcriptional gene expression regulation. Genes Dev. 2015;29(13):1343–55. https://doi.org/10.1101/gad.262766.115.
Xiao S, Cao S, Huang Q, Xia L, Deng M, Yang M, et al. The RNA N6-methyladenosine modification landscape of human fetal tissues. Nat Cell Biol. 2019;21(5):651–61. https://doi.org/10.1038/s41556-019-0315-4.
Yang C, Hu Y, Zhou B, Bao Y, Li Z, Gong C, et al. The role of m6A modification in physiology and disease. Cell Death Dis. 2020;11(11):960. https://doi.org/10.1038/s41419-020-03143-z.
Zaccara S, Ries RJ, Jaffrey SR. Reading, writing and erasing mRNA methylation. Nat Rev Mol Cell Biol. 2019;20(10):608–24. https://doi.org/10.1038/s41580-019-0168-5.
Meyer KD, Jaffrey SR. Rethinking m6A readers, Writers, and Erasers. Annu Rev Cell Dev Biol. 2017;33:319–42. https://doi.org/10.1146/annurev-cellbio-100616-060758.
Jiang X, Liu B, Nie Z, Duan L, Xiong Q, Jin Z, et al. The role of m6A modification in the biological functions and diseases. Signal Transduct Target Ther. 2021;6(1):74. https://doi.org/10.1038/s41392-020-00450-x.
Liu J, Yue Y, Han D, Wang X, Fu Y, Zhang L, et al. A METTL3-METTL14 complex mediates mammalian nuclear RNA N6-adenosine methylation. Nat Chem Biol. 2014;10(2):93–5. https://doi.org/10.1038/nchembio.1432.
Pendleton KE, Chen B, Liu K, Hunter OV, Xie Y, Tu BP, et al. The U6 snRNA m6A methyltransferase METTL16 regulates SAM synthetase Intron Retention. Cell. 2017;169(5):824–35e14. https://doi.org/10.1016/j.cell.2017.05.003.
Xiao W, Adhikari S, Dahal U, Chen Y-S, Hao Y-J, Sun B-F, et al. Nuclear m(6)a reader YTHDC1 regulates mRNA splicing. Mol Cell. 2016;61(4):507–19. https://doi.org/10.1016/j.molcel.2016.01.012.
Roundtree IA, Luo G-Z, Zhang Z, Wang X, Zhou T, Cui Y, et al. YTHDC1 mediates nuclear export of N6-methyladenosine methylated mRNAs. Elife. 2017;6:e31311. https://doi.org/10.7554/eLife.31311.
Hsu PJ, Zhu Y, Ma H, Guo Y, Shi X, Liu Y, et al. Ythdc2 is an N6-methyladenosine binding protein that regulates mammalian spermatogenesis. Cell Res. 2017;27(9):1115–27. https://doi.org/10.1038/cr.2017.99.
Wang X, Zhao BS, Roundtree IA, Lu Z, Han D, Ma H, et al. N(6)-methyladenosine modulates Messenger RNA translation efficiency. Cell. 2015;161(6):1388–99. https://doi.org/10.1016/j.cell.2015.05.014.
Wang X, Lu Z, Gomez A, Hon GC, Yue Y, Han D, et al. N6-methyladenosine-dependent regulation of messenger RNA stability. Nature. 2014;505(7481):117–20. https://doi.org/10.1038/nature12730.
Shi H, Wang X, Lu Z, Zhao BS, Ma H, Hsu PJ, et al. YTHDF3 facilitates translation and decay of N6-methyladenosine-modified RNA. Cell Res. 2017;27(3):315–28. https://doi.org/10.1038/cr.2017.15.
Meyer KD, Patil DP, Zhou J, Zinoviev A, Skabkin MA, Elemento O, et al. 5’ UTR m(6)a promotes Cap-Independent translation. Cell. 2015;163(4):999–1010. https://doi.org/10.1016/j.cell.2015.10.012.
Lin S, Choe J, Du P, Triboulet R, Gregory RI. The m(6)a methyltransferase METTL3 promotes translation in Human Cancer cells. Mol Cell. 2016;62(3):335–45. https://doi.org/10.1016/j.molcel.2016.03.021.
Hsu PJ, Shi H, Zhu AC, Lu Z, Miller N, Edens BM, et al. The RNA-binding protein FMRP facilitates the nuclear export of N6-methyladenosine-containing mRNAs. J Biol Chem. 2019;294(52):19889–95. https://doi.org/10.1074/jbc.AC119.010078.
Zhang F, Kang Y, Wang M, Li Y, Xu T, Yang W, et al. Fragile X mental retardation protein modulates the stability of its m6A-marked messenger RNA targets. Hum Mol Genet. 2018;27(22):3936–50. https://doi.org/10.1093/hmg/ddy292.
Edupuganti RR, Geiger S, Lindeboom RGH, Shi H, Hsu PJ, Lu Z, et al. N6-methyladenosine (m6A) recruits and repels proteins to regulate mRNA homeostasis. Nat Struct Mol Biol. 2017;24(10):870–8. https://doi.org/10.1038/nsmb.3462.
Liu N, Dai Q, Zheng G, He C, Parisien M, Pan T. N(6)-methyladenosine-dependent RNA structural switches regulate RNA-protein interactions. Nature. 2015;518(7540):560–4. https://doi.org/10.1038/nature14234.
Liu N, Zhou KI, Parisien M, Dai Q, Diatchenko L, Pan T. N6-methyladenosine alters RNA structure to regulate binding of a low-complexity protein. Nucleic Acids Res. 2017;45(10):6051–63. https://doi.org/10.1093/nar/gkx141.
Alarcón CR, Goodarzi H, Lee H, Liu X, Tavazoie S, Tavazoie SF. HNRNPA2B1 is a mediator of m(6)A-Dependent Nuclear RNA Processing events. Cell. 2015;162(6):1299–308. https://doi.org/10.1016/j.cell.2015.08.011.
Müller S, Glaß M, Singh AK, Haase J, Bley N, Fuchs T, et al. IGF2BP1 promotes SRF-dependent transcription in cancer in a m6A- and miRNA-dependent manner. Nucleic Acids Res. 2019;47(1):375–90. https://doi.org/10.1093/nar/gky1012.
Zhang L, Wan Y, Zhang Z, Jiang Y, Gu Z, Ma X, et al. IGF2BP1 overexpression stabilizes PEG10 mRNA in an m6A-dependent manner and promotes endometrial cancer progression. Theranostics. 2021;11(3):1100–14. https://doi.org/10.7150/thno.49345.
Hu X, Peng W-X, Zhou H, Jiang J, Zhou X, Huang D, et al. IGF2BP2 regulates DANCR by serving as an N6-methyladenosine reader. Cell Death Differ. 2020;27(6):1782–94. https://doi.org/10.1038/s41418-019-0461-z.
Li Z-X, Zheng Z-Q, Yang P-Y, Lin L, Zhou G-Q, Lv J-W, et al. WTAP-mediated m6A modification of lncRNA DIAPH1-AS1 enhances its stability to facilitate nasopharyngeal carcinoma growth and metastasis. Cell Death Differ. 2022;29(6):1137–51. https://doi.org/10.1038/s41418-021-00905-w.
Ruan D-Y, Li T, Wang Y-N, Meng Q, Li Y, Yu K, et al. FTO downregulation mediated by hypoxia facilitates colorectal cancer metastasis. Oncogene. 2021;40(33):5168–81. https://doi.org/10.1038/s41388-021-01916-0.
Hou P, Meng S, Li M, Lin T, Chu S, Li Z, et al. LINC00460/DHX9/IGF2BP2 complex promotes colorectal cancer proliferation and metastasis by mediating HMGA1 mRNA stability depending on m6A modification. J Experimental Clin Cancer Research: CR. 2021;40(1):52. https://doi.org/10.1186/s13046-021-01857-2.
Lang C, Yin C, Lin K, Li Y, Yang Q, Wu Z, et al. m6A modification of lncRNA PCAT6 promotes bone metastasis in prostate cancer through IGF2BP2-mediated IGF1R mRNA stabilization. Clin Transl Med. 2021;11(6):e426. https://doi.org/10.1002/ctm2.426.
Li A, Cao C, Gan Y, Wang X, Wu T, Zhang Q, et al. ZNF677 suppresses renal cell carcinoma progression through N6-methyladenosine and transcriptional repression of CDKN3. Clin Transl Med. 2022;12(6):e906. https://doi.org/10.1002/ctm2.906.
Wu H, Ding X, Hu X, Zhao Q, Chen Q, Sun T, et al. LINC01021 maintains tumorigenicity by enhancing N6-methyladenosine reader IMP2 dependent stabilization of MSX1 and JARID2: implication in colorectal cancer. Oncogene. 2022;41(13):1959–73. https://doi.org/10.1038/s41388-022-02189-x.
Liu J, Zhang N, Zeng J, Wang T, Shen Y, Ma C, et al. N6-methyladenosine-modified lncRNA ARHGAP5-AS1 stabilises CSDE1 and coordinates oncogenic RNA regulons in hepatocellular carcinoma. Clin Transl Med. 2022;12(11):e1107. https://doi.org/10.1002/ctm2.1107.
Chen Z, Huang L, Wang K, Zhang L, Zhong X, Yan Z, et al. rtcisE2F promotes the self-renewal and metastasis of liver tumor-initiating cells via N6-methyladenosine-dependent E2F3/E2F6 mRNA stability. Sci China Life Sci. 2022;65(9):1840–54. https://doi.org/10.1007/s11427-021-2038-5.
Tatekawa S, Tamari K, Chijimatsu R, Konno M, Motooka D, Mitsufuji S, et al. N(6)-methyladenosine methylation-regulated polo-like kinase 1 cell cycle homeostasis as a potential target of radiotherapy in pancreatic adenocarcinoma. Sci Rep. 2022;12(1):11074. https://doi.org/10.1038/s41598-022-15196-5.
Cheng Y, Gao Z, Zhang T, Wang Y, Xie X, Han G, et al. Decoding m6A RNA methylome identifies PRMT6-regulated lipid transport promoting AML stem cell maintenance. Cell Stem Cell. 2023;30(1):69–85e7. https://doi.org/10.1016/j.stem.2022.12.003.
Ye J, Chen X, Jiang X, Dong Z, Hu S, Xiao M. RNA demethylase ALKBH5 regulates hypopharyngeal squamous cell carcinoma ferroptosis by posttranscriptionally activating NFE2L2/NRF2 in an m6A-IGF2BP2-dependent manner. J Clin Lab Anal. 2022;36(7):e24514. https://doi.org/10.1002/jcla.24514.
Huo F-C, Xie M, Zhu Z-M, Zheng J-N, Pei D-S. SHMT2 promotes the tumorigenesis of renal cell carcinoma by regulating the m6A modification of PPAT. Genomics. 2022;114(4):110424. https://doi.org/10.1016/j.ygeno.2022.110424.
Yang Z, Wang T, Wu D, Min Z, Tan J, Yu B. RNA N6-methyladenosine reader IGF2BP3 regulates cell cycle and angiogenesis in colon cancer. J Exp Clin Cancer Res. 2020;39(1):203. https://doi.org/10.1186/s13046-020-01714-8.
Su C, Zhang Y, Chen P, Yang W, Du J, Zhang D. Methyltransferase-like 3 induces the development of cervical cancer by enhancing insulin-like growth factor 2 mRNA-binding proteins 3-mediated apoptotic chromatin condensation inducer 1 mRNA stability. Bioengineered. 2022;13(3):7034–48. https://doi.org/10.1080/21655979.2022.2044261.
Xia A, Yuan W, Wang Q, Xu J, Gu Y, Zhang L, et al. The cancer-testis lncRNA lnc-CTHCC promotes hepatocellular carcinogenesis by binding hnRNP K and activating YAP1 transcription. Nat Cancer. 2022;3(2):203–18. https://doi.org/10.1038/s43018-021-00315-4.
Zhang Y, Wang D, Wu D, Zhang D, Sun M, Long Noncoding RNA. KCNMB2-AS1 stabilized by N6-Methyladenosine modification promotes cervical Cancer Growth through acting as a competing endogenous RNA. Cell Transpl. 2020;29:963689720964382. https://doi.org/10.1177/0963689720964382.
Wang X, Tian L, Li Y, Wang J, Yan B, Yang L, et al. RBM15 facilitates laryngeal squamous cell carcinoma progression by regulating TMBIM6 stability through IGF2BP3 dependent. J Exp Clin Cancer Res. 2021;40(1):80. https://doi.org/10.1186/s13046-021-01871-4.
Gu Y, Niu S, Wang Y, Duan L, Pan Y, Tong Z, et al. DMDRMR-Mediated regulation of m(6)A-Modified CDK4 by m(6)a reader IGF2BP3 drives ccRCC progression. Cancer Res. 2021;81(4):923–34. https://doi.org/10.1158/0008-5472.CAN-20-1619.
Yu YZ, Lv DJ, Wang C, Song XL, Xie T, Wang T, et al. Hsa_circ_0003258 promotes prostate cancer metastasis by complexing with IGF2BP3 and sponging miR-653-5p. Mol Cancer. 2022;21(1):12. https://doi.org/10.1186/s12943-021-01480-x.
Hu Y, Gong C, Li Z, Liu J, Chen Y, Huang Y, et al. Demethylase ALKBH5 suppresses invasion of gastric cancer via PKMYT1 m6A modification. Mol Cancer. 2022;21(1):34. https://doi.org/10.1186/s12943-022-01522-y.
Du M, Peng Y, Li Y, Sun W, Zhu H, Wu J, et al. MYC-activated RNA N6-methyladenosine reader IGF2BP3 promotes cell proliferation and metastasis in nasopharyngeal carcinoma. Cell Death Discov. 2022;8(1):53. https://doi.org/10.1038/s41420-022-00844-6.
Zhang N, Shen Y, Li H, Chen Y, Zhang P, Lou S, et al. The m6A reader IGF2BP3 promotes acute myeloid leukemia progression by enhancing RCC2 stability. Exp Mol Med. 2022;54(2):194–205. https://doi.org/10.1038/s12276-022-00735-x.
Yu F, Wei J, Cui X, Yu C, Ni W, Bungert J, et al. Post-translational modification of RNA m6A demethylase ALKBH5 regulates ROS-induced DNA damage response. Nucleic Acids Res. 2021;49(10):5779–97. https://doi.org/10.1093/nar/gkab415.
Ying Y, Ma X, Fang J, Chen S, Wang W, Li J, et al. EGR2-mediated regulation of m6A reader IGF2BP proteins drive RCC tumorigenesis and metastasis via enhancing S1PR3 mRNA stabilization. Cell Death Dis. 2021;12(8):750. https://doi.org/10.1038/s41419-021-04038-3.
Wei J, Yin Y, Zhou J, Chen H, Peng J, Yang J, et al. METTL3 potentiates resistance to cisplatin through m6A modification of TFAP2C in seminoma. J Cell Mol Med. 2020;24(19):11366–80. https://doi.org/10.1111/jcmm.15738.
Li T, Hu P-S, Zuo Z, Lin J-F, Li X, Wu Q-N, et al. METTL3 facilitates tumor progression via an m6A-IGF2BP2-dependent mechanism in colorectal carcinoma. Mol Cancer. 2019;18(1):112. https://doi.org/10.1186/s12943-019-1038-7.
Feng P, Chen D, Wang X, Li Y, Li Z, Li B, et al. Inhibition of the m6A reader IGF2BP2 as a strategy against T-cell acute lymphoblastic leukemia. Leukemia. 2022;36(9):2180–8. https://doi.org/10.1038/s41375-022-01651-9.
Li H-B, Huang G, Tu J, Lv D-M, Jin Q-L, Chen J-K, et al. METTL14-mediated epitranscriptome modification of MN1 mRNA promote tumorigenicity and all-trans-retinoic acid resistance in osteosarcoma. EBioMedicine. 2022;82:104142. https://doi.org/10.1016/j.ebiom.2022.104142.
Li Z, Peng Y, Li J, Chen Z, Chen F, Tu J, et al. N(6)-methyladenosine regulates glycolysis of cancer cells through PDK4. Nat Commun. 2020;11(1):2578. https://doi.org/10.1038/s41467-020-16306-5.
Yang Z, Zhao F, Gu X, Feng L, Xu M, Li T, et al. Binding of RNA m6A by IGF2BP3 triggers chemoresistance of HCT8 cells via upregulation of ABCB1. Am J Cancer Res. 2021;11(4):1428–45.
Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144(5):646–74. https://doi.org/10.1016/j.cell.2011.02.013.
Warburg O. On the origin of cancer cells. Science. 1956;123(3191):309–14.
Zhang J, Pavlova NN, Thompson CB. Cancer cell metabolism: the essential role of the nonessential amino acid, glutamine. EMBO J. 2017;36(10):1302–15. https://doi.org/10.15252/embj.201696151.
Pavlova NN, Thompson CB. The emerging Hallmarks of Cancer Metabolism. Cell Metab. 2016;23(1):27–47. https://doi.org/10.1016/j.cmet.2015.12.006.
Levy JMM, Towers CG, Thorburn A. Targeting autophagy in cancer. Nat Rev Cancer. 2017;17(9):528–42. https://doi.org/10.1038/nrc.2017.53.
Shen C, Xuan B, Yan T, Ma Y, Xu P, Tian X, et al. m6A-dependent glycolysis enhances colorectal cancer progression. Mol Cancer. 2020;19(1):72. https://doi.org/10.1186/s12943-020-01190-w.
Lu S, Han L, Hu X, Sun T, Xu D, Li Y, et al. N6-methyladenosine reader IMP2 stabilizes the ZFAS1/OLA1 axis and activates the Warburg effect: implication in colorectal cancer. J Hematol Oncol. 2021;14(1):188. https://doi.org/10.1186/s13045-021-01204-0.
Wang Y, Lu J-H, Wu Q-N, Jin Y, Wang D-S, Chen Y-X, et al. LncRNA LINRIS stabilizes IGF2BP2 and promotes the aerobic glycolysis in colorectal cancer. Mol Cancer. 2019;18(1):174. https://doi.org/10.1186/s12943-019-1105-0.
Huang J, Sun W, Wang Z, Lv C, Zhang T, Zhang D, et al. FTO suppresses glycolysis and growth of papillary thyroid cancer via decreasing stability of APOE mRNA in an N6-methyladenosine-dependent manner. J Experimental Clin Cancer Research: CR. 2022;41(1):42. https://doi.org/10.1186/s13046-022-02254-z.
Weng H, Huang F, Yu Z, Chen Z, Prince E, Kang Y, et al. The m6A reader IGF2BP2 regulates glutamine metabolism and represents a therapeutic target in acute myeloid leukemia. Cancer Cell. 2022;40(12):1566–82e10. https://doi.org/10.1016/j.ccell.2022.10.004.
Xu Y, Zhou J, Li L, Yang W, Zhang Z, Zhang K, et al. FTO-mediated autophagy promotes progression of clear cell renal cell carcinoma via regulating SIK2 mRNA stability. Int J Biol Sci. 2022;18(15):5943–62. https://doi.org/10.7150/ijbs.77774.
Wang Q, Chen C, Ding Q, Zhao Y, Wang Z, Chen J, et al. METTL3-mediated m(6)a modification of HDGF mRNA promotes gastric cancer progression and has prognostic significance. Gut. 2020;69(7):1193–205. https://doi.org/10.1136/gutjnl-2019-319639.
Jiang L, Li Y, He Y, Wei D, Yan L, Wen H. Knockdown of m6A reader IGF2BP3 inhibited Hypoxia-Induced Cell Migration and Angiogenesis by regulating Hypoxia Inducible Factor-1alpha in stomach Cancer. Front Oncol. 2021;11:711207. https://doi.org/10.3389/fonc.2021.711207.
Maniotis AJ, Folberg R, Hess A, Seftor EA, Gardner LM, Pe’er J, et al. Vascular channel formation by human melanoma cells in vivo and in vitro: vasculogenic mimicry. Am J Pathol. 1999;155(3):739–52.
Liu X, He H, Zhang F, Hu X, Bi F, Li K, et al. m6A methylated EphA2 and VEGFA through IGF2BP2/3 regulation promotes vasculogenic mimicry in colorectal cancer via PI3K/AKT and ERK1/2 signaling. Cell Death Dis. 2022;13(5):483. https://doi.org/10.1038/s41419-022-04950-2.
Wan W, Ao X, Chen Q, Yu Y, Ao L, Xing W, et al. METTL3/IGF2BP3 axis inhibits tumor immune surveillance by upregulating N(6)-methyladenosine modification of PD-L1 mRNA in breast cancer. Mol Cancer. 2022;21(1):60. https://doi.org/10.1186/s12943-021-01447-y.
Kuai D, Zhu S, Shi H, Yang R, Liu T, Liu H, et al. Aberrant expression of m6A mRNA methylation regulators in colorectal adenoma and adenocarcinoma. Life Sci. 2021;273:119258. https://doi.org/10.1016/j.lfs.2021.119258.
Jiang Z, Chu PG, Woda BA, Rock KL, Liu Q, Hsieh C-C, et al. Analysis of RNA-binding protein IMP3 to predict metastasis and prognosis of renal-cell carcinoma: a retrospective study. Lancet Oncol. 2006;7(7):556–64.
Yang L, Wu S, Ma C, Song S, Jin F, Niu Y, et al. RNA m6A methylation regulators Subclassify Luminal subtype in breast Cancer. Front Oncol. 2020;10:611191. https://doi.org/10.3389/fonc.2020.611191.
Gong Y, Woda BA, Jiang Z. Oncofetal protein IMP3, a new cancer biomarker. Adv Anat Pathol. 2014;21(3):191–200. https://doi.org/10.1097/PAP.0000000000000021.
Zhou SL, Yue WB, Fan ZM, Du F, Liu BC, Li B, et al. Autoantibody detection to tumor-associated antigens of P53, IMP1, P16, cyclin B1, P62, C-myc, Survivn, and Koc for the screening of high-risk subjects and early detection of esophageal squamous cell carcinoma. Dis Esophagus. 2014;27(8):790–7. https://doi.org/10.1111/dote.12145.
Liu W, Wang P, Li Z, Xu W, Dai L, Wang K, et al. Evaluation of tumour-associated antigen (TAA) miniarray in immunodiagnosis of colon cancer. Scand J Immunol. 2009;69(1):57–63. https://doi.org/10.1111/j.1365-3083.2008.02195.x.
Zhang JY, Chan EK, Peng XX, Tan EM. A novel cytoplasmic protein with RNA-binding motifs is an autoantigen in human hepatocellular carcinoma. J Exp Med. 1999;189(7):1101–10.
Zhang J, Chan EKL. Autoantibodies to IGF-II mRNA binding protein p62 and overexpression of p62 in human hepatocellular carcinoma. Autoimmun Rev. 2002;1(3):146–53.
Li Y, Xiao J, Bai J, Tian Y, Qu Y, Chen X, et al. Molecular characterization and clinical relevance of m6A regulators across 33 cancer types. Mol Cancer. 2019;18(1):137. https://doi.org/10.1186/s12943-019-1066-3.
Li Y, Gu J, Xu F, Zhu Q, Chen Y, Ge D, et al. Molecular characterization, biological function, tumor microenvironment association and clinical significance of m6A regulators in lung adenocarcinoma. Brief Bioinform. 2021;22(4):bbaa225. https://doi.org/10.1093/bib/bbaa225.
Zhao H, Xu Y, Xie Y, Zhang L, Gao M, Li S, et al. m6A regulators is differently expressed and correlated with Immune Response of Esophageal Cancer. Front Cell Dev Biol. 2021;9:650023. https://doi.org/10.3389/fcell.2021.650023.
Wang L, Zhang S, Li H, Xu Y, Wu Q, Shen J, et al. Quantification of m6A RNA methylation modulators pattern was a potential biomarker for prognosis and associated with tumor immune microenvironment of pancreatic adenocarcinoma. BMC Cancer. 2021;21(1):876. https://doi.org/10.1186/s12885-021-08550-9.
Sun Z, Jing C, Xiao C, Li T, Wang Y. Prognostic risk signature based on the expression of three m6A RNA methylation regulatory genes in kidney renal papillary cell carcinoma. Aging. 2020;12(21):22078–94. https://doi.org/10.18632/aging.104053.
Huang W, Li Y, Zhang C, Zha H, Zhou X, Fu B, et al. IGF2BP3 facilitates cell proliferation and tumorigenesis via modulation of JAK/STAT signalling pathway in human bladder cancer. J Cell Mol Med. 2020;24(23):13949–60. https://doi.org/10.1111/jcmm.16003.
Ji G, Huang C, He S, Gong Y, Song G, Li X, et al. Comprehensive analysis of m6A regulators prognostic value in prostate cancer. Aging. 2020;12(14):14863–84. https://doi.org/10.18632/aging.103549.
Liu J, Zhou Z, Ma L, Li C, Lin Y, Yu T, et al. Effects of RNA methylation N6-methyladenosine regulators on malignant progression and prognosis of melanoma. Cancer Cell Int. 2021;21(1):453. https://doi.org/10.1186/s12935-021-02163-9.
Pan Z, Zhao R, Li B, Qi Y, Qiu W, Guo Q, et al. EWSR1-induced circNEIL3 promotes glioma progression and exosome-mediated macrophage immunosuppressive polarization via stabilizing IGF2BP3. Mol Cancer. 2022;21(1):16. https://doi.org/10.1186/s12943-021-01485-6.
Liu J, Li Z, Cheang I, Li J, Zhou C, RNA-Binding Protein. IGF2BP1 Associated with Prognosis and Immunotherapy Response in Lung Adenocarcinoma. Front Genet. 2022;13:777399. https://doi.org/10.3389/fgene.2022.777399.
Wang X, Guan D, Wang D, Liu H, Wu Y, Gong W, et al. Genetic variants in m6A regulators are associated with gastric cancer risk. Arch Toxicol. 2021;95(3):1081–8. https://doi.org/10.1007/s00204-020-02958-1.
Cui J, Zhu Y, Liu X, Wang W, Jiang X, Xia Y, et al. Comprehensive analysis of N6-methyladenosine regulators with the tumor immune landscape and correlation between the insulin-like growth factor 2 mRNA-binding protein 3 and programmed death ligand 1 in bladder cancer. Cancer Cell Int. 2022;22(1):72. https://doi.org/10.1186/s12935-022-02456-7.
Du J, Ji H, Ma S, Jin J, Mi S, Hou K, et al. m6A regulator-mediated methylation modification patterns and characteristics of immunity and stemness in low-grade glioma. Brief Bioinform. 2021;22(5):bbab013. https://doi.org/10.1093/bib/bbab013.
Liu H, Qin G, Ji Y, Wang X, Bao H, Guan X, et al. Potential role of m6A RNA methylation regulators in osteosarcoma and its clinical prognostic value. J Orthop Surg Res. 2021;16(1):294. https://doi.org/10.1186/s13018-021-02422-5.
Jin L, Chen C, Yao J, Yu Z, Bu L. The RNA N6-methyladenosine modulator HNRNPA2B1 is involved in the development of non-small cell lung cancer. Clin Exp Pharmacol Physiol. 2022;49(3):329–40. https://doi.org/10.1111/1440-1681.13608.
Chen L, Hu B, Song X, Wang L, Ju M, Li Z, et al. m6A RNA methylation regulators Impact Prognosis and Tumor Microenvironment in Renal Papillary Cell Carcinoma. Front Oncol. 2021;11:598017. https://doi.org/10.3389/fonc.2021.598017.
Li J, Cao J, Liang C, Deng R, Li P, Tian J. The analysis of N6-methyladenosine regulators impacting the immune infiltration in clear cell renal cell carcinoma. Med Oncol. 2022;39(4):41. https://doi.org/10.1007/s12032-021-01645-0.
Dahlem C, Abuhaliema A, Kessler SM, Kröhler T, Zoller BGE, Chanda S, et al. First small-molecule inhibitors targeting the RNA-Binding protein IGF2BP2/IMP2 for Cancer Therapy. ACS Chem Biol. 2022;17(2):361–75. https://doi.org/10.1021/acschembio.1c00833.
Mahapatra L, Andruska N, Mao C, Le J, Shapiro DJ. A novel IMP1 inhibitor, BTYNB, targets c-Myc and inhibits Melanoma and Ovarian Cancer Cell Proliferation. Transl Oncol. 2017;10(5):818–27. https://doi.org/10.1016/j.tranon.2017.07.008.
Mahapatra L, Mao C, Andruska N, Zhang C, Shapiro DJ. High-throughput fluorescence anisotropy screen for inhibitors of the oncogenic mRNA binding protein, IMP-1. J Biomol Screen. 2014;19(3):427–36. https://doi.org/10.1177/1087057113499633.
Wallis N, Oberman F, Shurrush K, Germain N, Greenwald G, Gershon T, et al. Small molecule inhibitor of Igf2bp1 represses Kras and a pro-oncogenic phenotype in cancer cells. RNA Biol. 2022;19(1):26–43. https://doi.org/10.1080/15476286.2021.2010983.
Roberts TC, Langer R, Wood MJA. Advances in oligonucleotide drug delivery. Nat Rev Drug Discov. 2020;19(10):673–94. https://doi.org/10.1038/s41573-020-0075-7.
Coulis CM, Lee C, Nardone V, Prokipcak RD. Inhibition of c-myc expression in cells by targeting an RNA-protein interaction using antisense oligonucleotides. Mol Pharmacol. 2000;57(3):485–94.
King DT, Barnes M, Thomsen D, Lee CH. Assessing specific oligonucleotides and small molecule antibiotics for the ability to inhibit the CRD-BP-CD44 RNA interaction. PLoS ONE. 2014;9(3):e91585. https://doi.org/10.1371/journal.pone.0091585.
Mehmood K, Akhtar D, Mackedenski S, Wang C, Lee CH. Inhibition of GLI1 expression by targeting the CRD-BP-GLI1 mRNA Interaction using a specific oligonucleotide. Mol Pharmacol. 2016;89(6):606–17. https://doi.org/10.1124/mol.115.102434.
Efferth T, Li PCH, Konkimalla VSB, Kaina B. From traditional chinese medicine to rational cancer therapy. Trends Mol Med. 2007;13(8):353–61.
Cui Y, Wu Y, Wang C, Wang Z, Li Y, Jiang Z, et al. Isoliquiritigenin inhibits non-small cell lung cancer progression via m6A/IGF2BP3-dependent TWIST1 mRNA stabilization. Phytomedicine. 2022;104:154299. https://doi.org/10.1016/j.phymed.2022.154299.
Zhang Y, Liu X, Yu M, Xu M, Xiao Y, Ma W, et al. Berberine inhibits proliferation and induces G0/G1 phase arrest in colorectal cancer cells by downregulating IGF2BP3. Life Sci. 2020;260:118413. https://doi.org/10.1016/j.lfs.2020.118413.
Li M, Zhang L, Ge C, Chen L, Fang T, Li H, et al. An isocorydine derivative (d-ICD) inhibits drug resistance by downregulating IGF2BP3 expression in hepatocellular carcinoma. Oncotarget. 2015;6(28):25149–60. https://doi.org/10.18632/oncotarget.4438.
Dai N, Christiansen J, Nielsen FC, Avruch J. mTOR complex 2 phosphorylates IMP1 cotranslationally to promote IGF2 production and the proliferation of mouse embryonic fibroblasts. Genes Dev. 2013;27(3):301–12. https://doi.org/10.1101/gad.209130.112.
Hüttelmaier S, Zenklusen D, Lederer M, Dictenberg J, Lorenz M, Meng X, et al. Spatial regulation of beta-actin translation by src-dependent phosphorylation of ZBP1. Nature. 2005;438(7067):512–5.
Lambrianidou A, Sereti E, Soupsana K, Komini C, Dimas K, Trangas T. mTORC2 deploys the mRNA binding protein IGF2BP1 to regulate c-MYC expression and promote cell survival. Cell Signal. 2021;80:109912. https://doi.org/10.1016/j.cellsig.2020.109912.
UniProt. The Universal protein knowledgebase in 2023. Nucleic Acids Res. 2023;51(D1):D523–D31. https://doi.org/10.1093/nar/gkac1052.
Acknowledgements
Not applicable.
Author information
Authors and Affiliations
Contributions
Tian-Yu Zhu designed the article and wrote the initial manuscript draft. Lian-Lian Hong revised and expanded the manuscript. Zhi-Qiang Ling was the overall principle investigator of this project, who designed the article and obtained financial support, was responsible for the manuscript design, oversaw the entire manuscript, and revised and synthesized the paper. All authors provided input into figures and tables and have seen and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Zhu, TY., Hong, LL. & Ling, ZQ. Oncofetal protein IGF2BPs in human cancer: functions, mechanisms and therapeutic potential. Biomark Res 11, 62 (2023). https://doi.org/10.1186/s40364-023-00499-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40364-023-00499-0