
Abstract
Haematologic malignancies account for a large proportion of cancers worldwide. The high occurrence and mortality of haematologic malignancies create a heavy social burden. Allogeneic haematopoietic stem cell transplantation is widely used in the treatment of haematologic malignancies. However, graft-versus-host disease and relapse after allogeneic haematopoietic stem cell transplantation are inevitable. An emerging treatment method, adoptive cellular therapy, has been effectively used in the treatment of haematologic malignancies. T cells, natural killer (NK) cells and tumour-infiltrating lymphocytes (TILs) all have great potential in therapeutic applications, and chimeric antigen receptor T (CAR-T) cell therapy especially has potential, but cytokine release syndrome and off-target effects are common. Efficient anticancer measures are urgently needed. In recent years, double-negative T cells (CD3+CD4−CD8−) have been found to have great potential in preventing allograft/xenograft rejection and inhibiting graft-versus-host disease. They also have substantial ability to kill various cell lines derived from haematologic malignancies in an MHC-unrestricted manner. In addition, healthy donor expanded double-negative T cells retain their antitumour abilities and ability to inhibit graft-versus-host disease after cryopreservation under good manufacturing practice (GMP) conditions, indicating that double-negative T cells may be able to be used as an off-the-shelf product. In this review, we shed light on the potential therapeutic ability of double-negative T cells in treating haematologic malignancies. We hope to exploit these cells as a novel therapy for haematologic malignancies.
Similar content being viewed by others


Background
Haematologic malignancies are cancers driven by genetic or epigenetic lesions in haematopoietic stem cells (HSCs) that lead to the development of cancer stem cells, and haematologic malignancies result in fewer functional haematologic cells and invasion into peripheral lymph organs [1, 2]. There are many types of haematologic malignancies, such as leukaemia, lymphoma, and myeloma. The global cancer statistics in 2021 produced by the International Agency for Research on Cancer showed a high incidence and death rate of haematologic malignancies, of which leukaemia ranked in the top ten in terms of mortality (the mortality of leukemia is 3.1%) [3]. Moreover, the latest cancer statistics published by Siegel et al. showed leukaemia was the most common childhood cancer, accounting for 28% of cases [4]. The high occurrence and mortality of haematologic malignancies indicate the inefficiency of therapy and poor prognosis, implying the urgent need for novel treatment methods.
Allogeneic haematopoietic stem cell transplantation (allo-HSCT) plays a crucial role in various haematologic malignancies. It usually begins with high doses of chemotherapy and radiation, followed by infusion of donor-specific haematopoietic stem cells derived from peripheral blood, bone marrow, or umbilical cord blood. The constitution of a new haematologic system is significantly related to the outcome. Graft-versus-host disease (GVHD) and relapse are two main causes of death after allo-HSCT [5]. Adoptive cellular therapy (ACT) is an emerging method in the treatment of haematologic cancers, involving the transfusion of cells with great antitumour capacity [6]. Many kinds of cells have been used in ACT and have demonstrated to be effective in the treatment of haematologic cancers, such as chimeric antigen receptor T (CAR-T) cells [7], chimeric antigen receptor natural killer (CAR-NK) cells [8], natural killer (NK) cells [9], and tumour-infiltrating-lymphocytes (TILs) [10], but CAR-T cell therapy is always accompanied by a series of side effects, such as serious cytokine release syndrome (CRS) and immune effector cell associated Neurotoxicity syndrome (ICANS) [11]. However, a newly identified type of cell, double-negative T cells (DNTs), have been found to be effective effector cells for ACT.
DNTs, also called natural suppressor (NS) cells, were first described by Strober for their suppressive activity and “null phenotype” without antigen specificity [12] (Fig. 1). Subsequently, they demonstrated inhibition of GVHD without major histocompatibility complex (MHC) restriction [13] and were identified to express the cell surface makers αβ-T cell receptor (TCR) and CD3, but without CD4 and CD8 [14]. In 2000, Zhang et al. [15] proposed DNTs for the first time, and these DNTs were characterized as abnormal regulatory T cells expressing αβ-TCR and CD25 but without CD4 or CD8 expression. Although DNTs account for only 1% ~ 5% of peripheral lymphocytes [16], many researchers have demonstrated the strong suppressive ability of DNTs towards CD8+ T cells, CD4+ T cells, B cells and NK cells in vitro and in vivo, which leads to xenograft or allograft transplantation tolerance and powerful prevention of GVHD [17,18,19]. In addition, DNTs possess strong antitumour characteristics without MHC restriction in vitro and in vivo and can be efficiently expanded ex vivo [20]. Therefore, we suggest that DNTs have great potential as off-the-shelf products and may have superior performance in the ACT field. Here, we review the functions and clinical application of DNTs to aid understanding.

The timeline of studies in development of DNTs function in immunoregulation and anticancer. Since the discovery of DNTs in 1984, a lot of research have been performed in DNTs development and function. DNTs, double negative T cells; GVHD, graft-versus-host disease; GVL, graft-versus-leukaemia
Development and subgroups of DNTs
Maturation of most T cells begins with migration of bone marrow-derived progenitor cells to the thymus, where they go through T cell receptor (TCR) rearrangement, positive selection and negative selection to acquire diverse TCR repertoires and tolerance to autoantigens [21]. However, DNTs, considered unconventional T cells, have controversial origins and maturation pathways due to their heterogeneous cell surface marker expression and differences in cytokine secretion [22]. Human T cells can mature into αβT cells and γδT cells, and the signal strength of the TCR plays a critical role in lineage differentiation [23]. The quality and magnitude of CD8+T-cell responses were important to overcome virus infections and kill tumour cells [24]. In addition, naïve CD4+ T cells could differentiate into effector Th cells that are characterized by the expression of distinct cytokines and transcription factors. These cells have differential immune functions [25,26,27,28]. What is more, in Hayes’s study, DN3 stage (a later double-negative (DN) stage in thymic T cell development) T cells converted into DNTs through strong signal transduction of the pre-TCR/CD3 complex [29]. Leonid and colleagues proposed that under TCR signalling, double-positive (DP) thymocytes are prevented from polarizing into conventional single-positive (SP) T cells, instead undergoing developmental diversion and then becoming DNTs. Subsequently, these DNTs could migrate to the intestine and differentiate into intraepithelial lymphocytes (IELs). Here, the features and properties of T cell subsets were summarized and shown in the form of table (Table 1).
Several studies have identified that activated peripheral CD4+ or CD8+ T cells can transform into DNTs, in the presence of different cytokines, such as IL-2 [31,32,33,34,35]. Fas-deficient or Fas ligand-deficient mice, which exhibit accumulation of DNTs in secondary lymphoid organs [36], are widely used in DNT research. Some researchers have proposed that the accumulation of DNTs in lpr mice results from Fas pathway deficiency, which impacts the normal clearance of DNTs rather than CD8+ or CD4+ T cells [37]. Zhang and coworkers also showed that DNTs can develop in athymic mice, and these DNTs exhibit more robust killing ability than those from mice with a thymus [38]. In addition, some researchers suggest that DNTs could be derived from the bone marrow [39], the appendix [40], the liver [41], the kidney [42, 43], nasal-associated lymphoid tissue(NALT) [44], or the female genital tract [45] and thus are thymus-independent. T-cell large granular lymphocyte(T-LGL) leukaemia is the most common cause of double-negative T-lymphocytosis, in which the peripheral DNTs increase a lot, accounting for 38.5%(median) of circulating lymphocytes [46]. Using a murine anaplastic large cell lymphoma (ALCL) model, they found DNTs (DN3/4 thymocytes) with aberrant TCR α/β chain could give rise to lymphoma stem cells and induce lymphoma [47]. In NP23-NHD13 double transgenic acute myelocytic leukaemia(AML) mice model, researchers verified the stemness of DNTs(DN1/2 thymocytes) [48]. These indicate a relationship between DNTs and leukemic stem cells and supply an idea for treatment of refractory leukaemia.
Based on our transcriptome data, DNTs can be divided into different subpopulations according to the distinction of phenotype and function. Using single-cell RNA sequencing, Lu Yang et al. [30] identified five naïve DNT (nDNT) subgroups from splenocytes of male C57BL/6 mice, including resting DNT (nDNT0), which was accounted for the majority (78.0%) of nDNTs, and contributed to synthetic proteins, keeping themselves alive, preparing for activation but not killing target cells; and helper DNT (nDNT1), which could secret cytokines, such as IL-17, intermediate DNT (nDNT2), cytotoxic DNT (nDNT3), as well as innate DNT (nDNT4) cells. In addition, Lu Yang et al. [30] demonstrated the transcriptome landscape of DNTs, and indicated that IKZF2 might be a distinguished transcriptional factor of DNTs. The characterization of functional DNT subsets into innate DNT, cytotoxic DNT, and helper DNT could help to better understand the function of DNT subsets, and may facilitate diagnosis and therapy. Therefore, DNTs play critical roles in the immune system, exploring functional interactions of DNTs and other immune cells is helpful for propelling clinical application in haematological malignancies.
Functions of DNTs
Antitumour functions of DNT cells
The fas/fasL pathway
An series of lymphocytes, such as CD8+ T cells, [49] TILs [50], NK cells [51] and γδT cells [52], have been employed as therapeutic effector cells in adoptive cell therapy (ACT) for their tumour cytotoxic ability, including in the treatment of haematologic malignancies. Fas-dependent pathway is one of the major mechanisms of T cell-mediated effects [53]. As unconventional Treg cells, DNTs have excellent antitumour capacity in haematologic malignancies. In Young et al.’s research, infusion of either primary DNTs or cloned DNTs prevented mice from onset and progression of lymphoma and lysed A20 lymphoma cells in vitro and in vivo without causing GVHD [54, 55]. A killing assay targeting lymphoma cells with high Fas expression and low Fas expression showed that DNT-mediated anti-leukaemia activity was dependent on the Fas pathway [20, 55]. Subsequently, researchers realized that ex vivo DNT expansion was sufficient for therapeutic needs, as discovered in acute myeloid leukaemia (AML) patients who achieved complete remission [20]. Of note, DNTs are cytotoxic to primary allogeneic blasts and autologous AML blasts [20]. In a recent study, Chen showed that pancreatic cancer cells cocultured with DNTs highly expressed Fas, caspase-8 and cleaved caspase-8, which are necessary in the Fas/FasL signalling pathway [56]. Therefore, they identified a Fas-dependent mechanism underlying the anti-pancreatic cancer effect of DNTs [57].
Innate lymphoid cell-like cytotoxic mechanisms
Innate lymphoid cells (ILCs), including NK cells, play a predominant role against cancer [58]. NK cells account for a major proportion of ILCs and act as common cytotoxic cells that induce antitumor cytotoxicity in a non-antigen-specific manner [59]. Similar to NK cells, DNTs exert innate killing ability in a non-antigen-specific manner. Using a patient-derived xenograft model of AML, Lee et al. demonstrated that DNTs showed notable antitumor effects both ex vivo and in vivo [60]. The researchers proved that DNTs encountering tumour cells expressed increased NKG2D and DNAM-1 [60], which are activating receptors and are expressed by most NK cells [61]. Binding with their ligands (MICA/B and ULBP1-6 for NKG2D and CD155 and CD112 for DNAM-1), which are expressed in tumour cells at a high level, induces effective antitumor activity [62]. Xu et al. [63] found elevated expression of NKG2D and MICA in vivo in a pancreatic carcinoma model. The researchers demonstrated a NKG2D-dependent mechanism of DNT antitumor activity [63]. Lee’s study [60] used a patient-derived AML xenograft model, and they observed high levels of NKG2D and DNAM-1, as well as their respective ligands, in DNTs and AML cells. Moreover, secretion of IFN-γ by DNTs after interaction with tumour cells massively increases the expression of ligands in AML cells and promotes killing activity [60]. In addition to NKG2D and DNAM-1, when DNTs are cocultured with haematologic malignancies cell lines, NKp30, KIR2DS4 and membrane TRAIL (mTRAIL), which are all actively involved in cytotoxicity, are also detected [64]. Interestingly, in the study, costimulatory molecules CD30, GITR, CD27 and CD28 expression of DNTs increase, while coinhibitory molecules ICOS, CTLA4, PD-1 and PD-1 ligands are low, suggesting well viability of DNTs [64].
Cytokines
The secretion of IFN-γ, TNF, perforin and granzyme B has been discovered in haematologic malignancies models. Genetically cloned DNTs (T4H2) secrete IFN-γ and TNF upon recognizing specific antigen-loaded melanoma cells [65]. When ex vivo-expanded DNTs recognize AML cells, they express IFN-γ and TNF at levels that are similar to or higher than those seen in bulk activated CD8+ T cells [20], and this phenomenon was also shown after recognition of lung cancer cells by DNTs [66]. Ponzetta et al. [67] proposed a novel antitumour pathway by which tumour-associated neutrophils can induce IL-12 secretion by macrophages and thus lead to the polarization of cytotoxic DNTs, ultimately achieving tumour inhibition in an IFN-γ-dependent manner. IFN-γ has been shown to play a critical antitumour role and to participate in cancer immunosurveillance and the upregulation of cytotoxic ligands such as TNF, FasL and TRAIL [68], enhancing anti-leukaemia capacity. Additionally, the perforin-dependent pathway is strongly related to DNT-mediated antitumour activity. Ex vivo-expanded DNTs exhibit an effective ability to target and kill allogeneic blasts and autologous leukaemic blasts in a perforin- and granzyme B-dependent manner, which was confirmed in Lee’s study [20, 60]. In a study of DNT-mediated killing of melanoma cells, DNTs showed high expression of perforin and granzyme B, indicating DNT activity [65], which was consistent with research on cytotoxic DNTs targeting NSCLC cells [66]. There may be other mechanisms by which DNTs enact their antitumour abilities, and discovering the interactions of the various involved pathways will deepen our understanding of DNT cellular immunotherapy.
Regulation of adaptive and innate immunity
Cell types with immunoregulatory functions, including CD4+ regulatory T (Treg) cells, CD8+ T cells and γδTCR+ T cells, play crucial roles in HSCT and tumour immunity [69]. DNTs were first demonstrated to possess regulatory functions in 2000 [15]. Accumulated evidence demonstrated that DNTs paly crucial role in both adaptive and innate immunity. Using a murine model, Zhang et al. proposed that DNTs could eliminate CD8+ T cells in a cell–cell contact-dependent manner [15]. The eradication of CD8+ T cells is Fas-dependent and antigen-specific and is closely related to trogocytosis [70], a phenomenon resulting in membrane acquisition by the majority of lymphocytes. DNTs can express MHC-peptide complexes to APCs, interact with syngenic CD8+ T cells through TCR-MHC recognition and subsequently kill the cells via the Fas-FasL pathway [71, 72]. In addition, Mackensen and coworkers studied human peripherally derived DNTs and found that they showed high expression of perforin and antigen-specific killing of CD4+/CD8+ T cells [65, 73]. DNTs from humans share similar regulatory functions with those from murine models [73, 74]. The cytotoxic activity of murine DNTs has also been demonstrated to be perforin- and granzyme B-dependent [34, 75]. The IL-7/Akt/mTOR signalling pathway was recently found to play a potential role in the regulatory functions of DNTs during their elimination of target cells. A recent study showed that DNTs restrain mTOR signalling in CD4+ T cells, resulting in the inhibition of glucose uptake, which suggests that DNTs can also suppress metabolic alterations [76]. DNTs have shown an ability to inhibit CD4+ and CD8+ T cells, and researchers have demonstrated that both ex vivo- and in vivo-induced DNTs can prevent immune rejection in an antigen-specific manner in allogeneic and xenogeneic skin and cardiac grafts in transgenic and wild-type mouse models [15, 18, 77, 78].
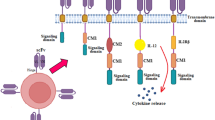
Of interest, accumulation of infiltrating DNTs was found in donor-specific skin grafts, and the interaction of CXCR5 expressed in DNTs and CXCL13 upregulated in cardiac graft after transplantation was essential for the migration of DNTs into the allografts [79]. What is more, CXCR5/CXCL13 interaction is important for DNTs home to lymph nodes and kidneys [29, 79, 80]. In addition, DNTs infiltrated in the allograft could kill syngeneic CD8 effector T cell and induce immune tolerance. This finding indicates that DNTs could play a role in the local graft [81]. Moreover, antibodies secreted by mature B cells hinder long-term tolerance in allografts and transplantation [82, 83]. In a xenograft model, adoptive transfer of xenoreactive DNTs lysed B cells via perforin- and granzyme B-dependent mechanisms and inhibited IgG subtypes, which resulted in alteration of rejection patterns from acute vascular rejection mainly mediated by antibodies to cell-mediated rejection [75, 84]. During this process, the TCR/MHC-peptide interaction may increase the killing activity of DNTs by effectively stimulating DNT proliferation [85]. In Hu’s study [86], DNTs interacted with B cells in a natural killer group 2-member D (NKG2D)-NKG2D ligand-dependent manner and released accumulated granzyme B, which accelerated B cell apoptosis and prevented the proliferation of B cells. Dendritic cells (DCs) plays an important role in innate immunity. Data have shown that DNTs highly expressing NKG2D tend to lyse DCs through the Fas/FasL pathway in an antigen-specific manner and downregulate the expression of CD80 and CD86 in DCs, which prevents them from interacting with T cells [87] (Fig. 2). Of note, infusion of autologous tolerogenic DCs is also able to induce immunosuppression by producing Epstein-Barr virus-induced gene 3 (EBI3), after which IFN-γ secretion by DNTs is increased [88].

The regulatory and antitumor functions of DNTs. The antitumor functions of DNTs depend on expression of FasL, NKG2D, DNAM-1, TRAIL and NKp30. Moreover, secretion of IFN-γ, TNF-α, granzyme B and perforin take an important place. In addition, DNTs can mediate many types of immunocyte and exert immunoregulatory functions. DNTs downregulate expression of CD80/CD86 in dendritic cell and impact their interaction with other cells. DNTs can express MHC-peptide complex which expressed in dendritic cell by trogocytosis way, then interact with T cell and kill them through Fas/Fas L manner. Granzyme B and perforin secretion by DNTs induce death of T cell and B cell. Alternatively, DNTs kill M1 macrophage by down-regulating TNF-α and IFN-γ secretion of M1. FasL, Fas ligand; TNF-α, tumor necrosis factor α; NKG2D, natural killer group 2-member D; TRAIL: tumour necrosis factor-related apoptosis-inducing ligand
Moreover, Lu Yang et al. reported that different naïve DNT subsets (resting DNT (nDNT0), helper DNT (nDNT1), intermediate DNT (nDNT2), cytotoxic DNT (nDNT3), and innate DNT (nDNT4)) also upregulated several genes associated with innate or adaptive immune responses, therefore, the functions of five identified DNT subgroups may span a wide spectrum from innate to adaptive immune responses [30].
Clinical application of DNTs
Relationship with GVHD and GVL
DNTs play an important role in anti-leukaemia and regulating immune response. In Lee’s study [64], DNTs expanded under good manufacturing practice (GMP) conditions were cytotoxic to myeloma, T-cell leukaemia, Burkitt lymphoma, and AML In light of the promising regulatory function and antitumour ability of DNTs, the transfer of DNTs could result in excellent antitumor effects without causing GVHD, which is a major obstacle in cellular immunotherapy [89].
Our data showed the expanded honor-derived DNTs effectively kill leukemic cell lines and primary cells derived from leukaemia patient without toxicity to CD34+ hematopoietic stem cells. What is more, DNTs suppressed the proliferation of CD4+ T cells in vitro. A first-in human clinical trial conducted by our transplantation center found that the in vivo infusion of DNTs is safe. Through RNA-seq, we demonstrated the increased expression of GZMA, GZMB and GZMK in CD8+ T cell cocultured with DNTs. Furthermore, we used reverse transcription-polymerase chain reaction (RT-qPCR) as well as flow cytometry, and confirmed the increased expression of GZMB and IFN-γ of CD8+ T cell. Therefore, we suggest allogenetic DNTs may preserve the GVL effect by increased expression of IFN-γ and GZM of CD8+ T cells.
In Ye’s research [90], the researchers detected DNT-cell reconstitution in 73 patients undergoing allo-HSCT at days 30, 60, 90 and 180 and demonstrated delayed reconstitution in patients who developed acute GVHD. The researchers demonstrated a reverse linear relationship between the reconstitution of DNTs at 60 days and the severity of GVHD [90]. Coincidentally, Hillhouse et al. proposed that decreases in the levels of DNTs significantly affect the occurrence of chronic GVHD by mediating B cells [91]. In addition, McIver et al. [92] investigated a cohort of 40 patients experiencing GVHD after HSCT and found a negative relationship between DNT number and grade of GVHD.
Novel clinical therapies
DNTs are potential ACT candidates for the treatment of haematopoietic malignancies due to their substantial graft-versus-leukaemia effect and GVHD suppression. Lee et al. [64] established a novel method to produce clinical-grade therapeutic DNTs. Off-the-shelf DNTs have outstanding killing ability in various cancers without causing GVHD and remain effective after cryopreservation [64]. To explore the safety and efficacy of DNTs, a first-in-human phase I/IIa clinical trial (ChiCTR1900023499) of the infusion of unrelated third-party donor DNTs for the treatment of AML patients who relapsed after allo-HSCT conducted by our transplantation centre was completed. After lymphodepleting preconditioning, 10 AML patients with relapsed or residual disease after allo-HSCT were treated with DNTs. No patients developed GVHD or ≥ grade 3 DNT treatment-related toxicity. An initial response was observed in 70.0% of evaluable patients. Five of the DNT-treated patients remained alive, and 4 remained in complete remission (CR) for 16.5 to 24.3 months after DNT treatment. The predicted 2-year overall survival after DNT treatment was 50.0%. This study supports the safety, feasibility, and potential efficacy of allo-DNTs as a novel ACT for relapsed AML after allo-HSCT. Using CRISPR, Soares [93] screened genes in DNT treatment-resistant AML and identified the expression of CD64 as a strongly related factor that can be used as a marker to predict the effect of DNT cellular therapy. Moreover, novel treatments for adoptive DNT combined with other treatments have been explored. Chen found that infusion of DNTs after treatment with conventional chemotherapy, such as cytosine arabinoside (Arac) and daunorubicin (DNR), has greater cytotoxic activity in chemotherapy-resistant AML [94]. The same outcome was obtained when DNTs and venetoclax, a bcl-2 inhibitor, and DNTs and PD-1 inhibitors were combined [95, 96]. These findings provide suggestions for combining DNT cellular therapy with other methods for curing refractory cancer.
Perspectives and conclusions
DNTs have been demonstrated to have great cytotoxic activity towards various haematopoietic cancers, such as myeloma, T cell leukaemia, Burkitt’s lymphoma, and AML, in vitro and to possess cytotoxicity against AML in vivo without HLA restriction [64]; in addition, they have been shown to prevent GVHD. Currently, healthy donor-derived DNTs can be expanded in vitro and maintain viability and cytotoxic ability after being cryopreserved under GMP conditions [64]. However, the characteristics and subtypes of DNTs are not fully understood.
Though DNT therapy is a novel form of adoptive cellular therapy (ACT) with promising result in treating relapsed or refractory AML in preclinical studies, the in vivo persistence and efficacy of DNTs need to be improved. Hyeonjeong Kang et al. [97] used an PI3Kδ inhibitor, idelalisib (Ide) and expanded DNTs. They verified that Ide treatment could improve the overall fitness of DNTs to promote better engraftment and longer persistence in vivo. Given that clinical application of both DNTs and Ide have been tried, application of these findings into clinic might contribute to enhancing the efficacy and durability of response [97]. What is more, DNTs therapy could significantly reduce lung tumor growth in patient-derived xenograft models but could not eradicate the full extent of the tumor cells [98].
After infusion of DNTs [60], ~ 30% of the primary AML cells remain.
DNT cells play important role both in regulatory functions and in protective immune responses, therefore, new studies are needed to elucidate mechanisms of their differentiation, regulation, effector activities, as well as the in vivo kinetics in patients. Clarifying these issues could not only provide knowledge of the pathogenic and protective mechanisms of human diseases but also further develop the potential use of DNT cells as therapeutic strategy.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- HSCs:
-
Hematopoietic stem cells
- allo-HSCT:
-
Allogeneic hematopoietic stem cell transplantation
- GVHD:
-
Graft-versus-host-disease
- ACT:
-
Adoptive cellular therapy
- CAR:
-
Chimeric antigen receptor
- NK:
-
Natural killer
- TIL:
-
Tumor-infiltrating-lymphocytes
- CRS:
-
Cytokine-release-syndrome
- ICANS:
-
Immune effector cell associated neurotoxicity syndrome
- DNTs:
-
Double negative T cells
- TCR:
-
T cell receptor
- MHC:
-
Major histocompatibility complex
- DP:
-
Double positive
- SP:
-
Single positive
- T-LGL:
-
T-cell large granular lymphocyte
- ALCL:
-
Anaplastic large cell lymphoma
- IELs:
-
Intraepithelial lymphocytes
- DCs:
-
Dendritic cells
- NALT:
-
Nasal-associated lymphoid tissue
- Treg cell:
-
Regulatory T cell
- DLI:
-
Donor lymphocyte infusion
- NKG2D:
-
Natural killer group 2-member D
- EBI3:
-
Epstein-Barr virus-induced gene 3
- ILC:
-
Innate lymphoid cell
- mTRAIL:
-
Membrane tumor necrosis factor-related apoptosis-inducing ligand
- sTRAIL:
-
Soluble tumor necrosis factor-related apoptosis-inducing ligand
- NSCLC:
-
Non-small-cell lung cancer
- RT-qPCR:
-
Reverse transcription-polymerase chain reaction
- Arac:
-
Cytosine arabinoside
- DNR:
-
Daunorubicin
- Ide:
-
Idelalisib
References
Mendez-Ferrer S, Bonnet D, Steensma DP, Hasserjian RP, Ghobrial IM, Gribben JG, et al. Bone marrow niches in haematological malignancies. Nat Rev Cancer. 2020;20(5):285–98.
Andersen M, Hasselbalch HC, Kjaer L, Skov V, Ottesen JT. Global dynamics of healthy and cancer cells competing in the hematopoietic system. Math Biosci. 2020;326:108372.
Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;71(3):209–49.
Siegel RL, Miller KD, Fuchs HE, Jemal A. Cancer statistics, 2022. CA Cancer J Clin. 2022;72(1):7–33.
D’Souza A, Fretham C, Lee SJ, Arora M, Brunner J, Chhabra S, et al. Current use of and trends in hematopoietic cell transplantation in the United States. Biol Blood Marrow Transplant. 2020;26(8):e177–82.
Brenner MK. Adoptive Cell Therapy: ACT-Up or ACT-Out? Mol Ther. 2019;27(4):693–4.
Larson RC, Maus MV. Recent advances and discoveries in the mechanisms and functions of CAR T cells. Nat Rev Cancer. 2021;21(3):145–61.
Xie G, Dong H, Liang Y, Ham JD, Rizwan R, Chen J. CAR-NK cells: A promising cellular immunotherapy for cancer. EBioMedicine. 2020;59:102975.
Caruso S, De Angelis B, Carlomagno S, Del Bufalo F, Sivori S, Locatelli F, et al. NK cells as adoptive cellular therapy for hematological malignancies: Advantages and hurdles. Semin Hematol. 2020;57(4):175–84.
Stevanovic S, Helman SR, Wunderlich JR, Langhan MM, Doran SL, Kwong MLM, et al. A phase ii study of tumor-infiltrating lymphocyte therapy for human papillomavirus-associated epithelial cancers. Clin Cancer Res. 2019;25(5):1486–93.
Schubert ML, Schmitt M, Wang L, Ramos CA, Jordan K, Muller-Tidow C, et al. Side-effect management of chimeric antigen receptor (CAR) T-cell therapy. Ann Oncol. 2021;32(1):34–48.
Oseroff A, Okada S, Strober S. Natural suppressor (NS) cells found in the spleen of neonatal mice and adult mice given total lymphoid irradiation (TLI) express the null surface phenotype. J Immunol. 1984;132(1):101–10.
Strober S, Palathumpat V, Schwadron R, Hertel-Wulff B. Cloned natural suppressor cells prevent lethal graft-vs-host disease. J Immunol. 1987;138(3):699–703.
Strober S, Dejbachsh-Jones S, Van Vlasselaer P, Duwe G, Salimi S, Allison JP. Cloned natural suppressor cell lines express the CD3+CD4-CD8- surface phenotype and the alpha, beta heterodimer of the T cell antigen receptor. J Immunol. 1989;143(4):1118–22.
Zhang ZX, Yang L, Young KJ, DuTemple B, Zhang L. Identification of a previously unknown antigen-specific regulatory T cell and its mechanism of suppression. Nat Med. 2000;6(7):782–9.
Juvet SC, Zhang L. Double negative regulatory T cells in transplantation and autoimmunity: recent progress and future directions. J Mol Cell Biol. 2012;4(1):48–58.
Zhang Z, Yang L, Young KJ, Zhang L. Suppression of alloimmune responses in vitro and in vivo by CD3(+)CD8(-)CD4(-)alphabeta(+) regulatory T cells. Transplant Proc. 2001;33(1–2):84–5.
Chen W, Ford MS, Young KJ, Cybulsky MI, Zhang L. Role of double-negative regulatory T cells in long-term cardiac xenograft survival. J Immunol. 2003;170(4):1846–53.
Young KJ, DuTemple B, Phillips MJ, Zhang L. Inhibition of graft-versus-host disease by double-negative regulatory t cells. J Immunol. 2003;171(1):134–41.
Merims S, Li X, Joe B, Dokouhaki P, Han M, Childs RW, et al. Anti-leukemia effect of ex vivo expanded DNT cells from AML patients: a potential novel autologous T-cell adoptive immunotherapy. Leukemia. 2011;25(9):1415–22.
Liu C, Lan Y, Liu B, Zhang H, Hu H. T cell development: old tales retold by single-cell RNA sequencing. Trends Immunol. 2021;42(2):165–75.
D’Acquisto F, Crompton T. CD3+CD4−CD8− (double negative) T cells: Saviours or villains of the immune response? Biochem Pharmacol. 2011;82(4):333–40.
Hayes SM, Shores EW, Love PE. An architectural perspective on signaling by the pre-, alphabeta and gammadelta T cell receptors. Immunol Rev. 2003;191:28–37.
Wang D, Fu B, Shen X, Guo C, Liu Y, Zhang J, et al. Restoration of HBV-specific CD8(+) T-cell responses by sequential low-dose IL-2 treatment in non-responder patients after IFN-alpha therapy. Signal Transduct Target Ther. 2021;6(1):376.
Dong C. Cytokine regulation and function in t cells. Annu Rev Immunol. 2021;39:51–76.
Zhou L, Chong MM, Littman DR. Plasticity of CD4+ T cell lineage differentiation. Immunity. 2009;30(5):646–55.
Basu A, Ramamoorthi G, Albert G, Gallen C, Beyer A, Snyder C, et al. Differentiation and regulation of TH Cells: A balancing act for cancer immunotherapy. Front Immunol. 2021;12:669474.
Wing JB, Tanaka A, Sakaguchi S. Human FOXP3 (+) regulatory T cell heterogeneity and function in autoimmunity and cancer. Immunity. 2019;50(2):302–16.
Hayes SM, Li L, Love PE. TCR signal strength influences alphabeta/gammadelta lineage fate. Immunity. 2005;22(5):583–93.
Yang L, Zhu Y, Tian D, Wang S, Guo J, Sun G, et al. Transcriptome landscape of double negative T cells by single-cell RNA sequencing. J Autoimmun. 2021;121:102653.
Crispin JC, Tsokos GC. Human TCR-alpha beta+ CD4- CD8- T cells can derive from CD8+ T cells and display an inflammatory effector phenotype. J Immunol. 2009;183(7):4675–81.
Rodríguez-Rodríguez N, Flores-Mendoza G, Apostolidis SA, Rosetti F, Tsokos GC, Crispín JC. TCR-α/β CD4(-) CD8(-) double negative T cells arise from CD8(+) T cells. J Leukoc Biol. 2020;108(3):851–7.
Hedrich CM, Rauen T, Crispin JC, Koga T, Ioannidis C, Zajdel M, et al. cAMP-responsive element modulator α (CREMα) trans-represses the transmembrane glycoprotein CD8 and contributes to the generation of CD3+CD4-CD8- T cells in health and disease. J Biol Chem. 2013;288(44):31880–7.
Zhang D, Yang W, Degauque N, Tian Y, Mikita A, Zheng XX. New differentiation pathway for double-negative regulatory T cells that regulates the magnitude of immune responses. Blood. 2007;109(9):4071–9.
Rensing-Ehl A, Volkl S, Speckmann C, Lorenz MR, Ritter J, Janda A, et al. Abnormally differentiated CD4+ or CD8+ T cells with phenotypic and genetic features of double negative T cells in human Fas deficiency. Blood. 2014;124(6):851–60.
Rossin A, Miloro G, Hueber AO. TRAIL and fasl functions in cancer and autoimmune diseases: towards an increasing complexity. Cancers (Basel). 2019;11(5):639.
Martina MN, Noel S, Saxena A, Rabb H, Hamad ARA. Double Negative (DN) αβ T Cells: misperception and overdue recognition. Immunol Cell Biol. 2014;93(3):305–10.
Ford MS, Zhang Z-X, Chen W, Zhang L. Double-negative t regulatory cells can develop outside the thymus and do not mature from CD8+T cell precursors. J Immunol. 2006;177(5):2803–9.
Palathumpat V, Dejbakhshjones S, Holm B, Strober S. Different subsets of t-cells in the adult-mouse bone-marrow and spleen induce or suppress acute graft-versus-host disease. J Immunol. 1992;149(3):808–17.
Yamagiwa S, Sugahara S, Shimizu T, Iwanaga T, Yoshida Y, Honda S, et al. The primary site of CD4- 8–B220+ alphabeta T cells in lpr mice: the appendix in normal mice. J Immunol. 1998;160(6):2665–74.
Ohteki T, Seki S, Abo T, Kumagai K. Liver is a possible site for the proliferation of abnormal CD3+4-8- double-negative lymphocytes in autoimmune MRL-lpr/lpr mice. J Exp Med. 1990;172(1):7–12.
Ascon DB, Ascon M, Satpute S, Lopez-Briones S, Racusen L, Colvin RB, et al. Normal mouse kidneys contain activated and CD3+CD4- CD8- double-negative T lymphocytes with a distinct TCR repertoire. J Leukoc Biol. 2008;84(6):1400–9.
Sadasivam M, Noel S, Lee SA, Gong J, Allaf ME, Pierorazio P, et al. Activation and Proliferation of PD-1(+) Kidney Double-Negative T Cells Is Dependent on Nonclassical MHC Proteins and IL-2. J Am Soc Nephrol. 2019;30(2):277–92.
Rharbaoui F, Bruder D, Vidakovic M, Ebensen T, Buer J, Guzmán CA. Characterization of a B220+ lymphoid cell subpopulation with immune modulatory functions in nasal-associated lymphoid tissues. J Immunol. 2005;174(3):1317–24.
Johansson M, Lycke N. A unique population of extrathymically derived alpha beta TCR+CD4-CD8- T cells with regulatory functions dominates the mouse female genital tract. J Immunol. 2003;170(4):1659–66.
Liapis K, Tsagarakis NJ, Panitsas F, Taparkou A, Liapis I, Roubakis C, et al. Causes of double-negative T-cell lymphocytosis in children and adults. J Clin Pathol. 2020;73(7):431–8.
Kreutmair S, Klingeberg C, Poggio T, Andrieux G, Keller A, Miething C, et al. Existence of reprogrammed lymphoma stem cells in a murine ALCL-like model. Leukemia. 2020;34(12):3242–55.
Kundu S, Park ES, Chung YJ, Walker RL, Zhu YJ, Negi V, et al. Thymic precursor cells generate acute myeloid leukemia in NUP98-PHF23/NUP98-HOXD13 double transgenic mice. Sci Rep. 2019;9(1):17213.
Effern M, Glodde N, Braun M, Liebing J, Boll HN, Yong M, et al. Adoptive T cell therapy targeting different gene products reveals diverse and context-dependent immune evasion in melanoma. Immunity. 2020;53(3):564-80 e9.
Nguyen LT, Saibil SD, Sotov V, Le MX, Khoja L, Ghazarian D, et al. Phase II clinical trial of adoptive cell therapy for patients with metastatic melanoma with autologous tumor-infiltrating lymphocytes and low-dose interleukin-2. Cancer Immunol Immunother. 2019;68(5):773–85.
Myers JA, Miller JS. Exploring the NK cell platform for cancer immunotherapy. Nat Rev Clin Oncol. 2021;18(2):85–100.
Sebestyen Z, Prinz I, Dechanet-Merville J, Silva-Santos B, Kuball J. Translating gammadelta (gammadelta) T cells and their receptors into cancer cell therapies. Nat Rev Drug Discov. 2020;19(3):169–84.
Kägi D, Vignaux F, Ledermann B, Bürki K, Depraetere V, Nagata S, et al. Fas and perforin pathways as major mechanisms of T cell-mediated cytotoxicity. Science. 1994;265(5171):528–30.
Young KJ, DuTemple B, Zhang Z, Levy G, Zhang L. CD4(-)CD8(-) regulatory T cells implicated in preventing graft-versus-host and promoting graft-versus-leukemia responses. Transplant Proc. 2001;33(1–2):1762–3.
Young KJ, Kay LS, Phillips MJ, Zhang L. Antitumor activity mediated by double-negative T cells. Cancer Res. 2003;63(22):8014–21.
Kaufmann T, Strasser A, Jost PJ. Fas death receptor signalling: roles of Bid and XIAP. Cell Death Differ. 2012;19(1):42–50.
Chen J, Hu P, Wu G, Zhou H. Antipancreatic cancer effect of DNT cells and the underlying mechanism. Pancreatology. 2019;19(1):105–13.
Artis D, Spits H. The biology of innate lymphoid cells. Nature. 2015;517(7534):293–301.
Chiossone L, Dumas PY, Vienne M, Vivier E. Natural killer cells and other innate lymphoid cells in cancer. Nat Rev Immunol. 2018;18(11):671–88.
Lee J, Minden MD, Chen WC, Streck E, Chen B, Kang H, et al. Allogeneic human double negative T cells as a novel immunotherapy for acute myeloid leukemia and its underlying mechanisms. Clin Cancer Res. 2018;24(2):370–82.
Zingoni A, Ardolino M, Santoni A, Cerboni C. NKG2D and DNAM-1 activating receptors and their ligands in NK-T cell interactions: role in the NK cell-mediated negative regulation of T cell responses. Front Immunol. 2012;3:408.
Molfetta R, Zingoni A, Santoni A, Paolini R. Post-translational Mechanisms Regulating NK Cell Activating Receptors and Their Ligands in Cancer: Potential Targets for Therapeutic Intervention. Front Immunol. 2019;10:2557.
Xu H, Zhu XX, Chen J. DNT cell inhibits the growth of pancreatic carcinoma via abnormal expressions of NKG2D and MICA in vivo. Biochem Biophys Res Commun. 2016;469(2):145–50.
Lee JB, Kang H, Fang L, D’Souza C, Adeyi O, Zhang L. Developing allogeneic double-negative T cells as a novel off-the-shelf adoptive cellular therapy for cancer. Clin Cancer Res. 2019;25(7):2241–53.
Voelkl S, Moore TV, Rehli M, Nishimura MI, Mackensen A, Fischer K. Characterization of MHC class-I restricted TCRalphabeta+ CD4- CD8- double negative T cells recognizing the gp100 antigen from a melanoma patient after gp100 vaccination. Cancer Immunol Immunother. 2009;58(5):709–18.
Yao J, Ly D, Dervovic D, Fang L, Lee JB, Kang H, et al. Human double negative T cells target lung cancer via ligand-dependent mechanisms that can be enhanced by IL-15. J Immunother Cancer. 2019;7(1):17.
Ponzetta A, Carriero R, Carnevale S, Barbagallo M, Molgora M, Perucchini C, et al. Neutrophils Driving Unconventional T Cells Mediate Resistance against Murine Sarcomas and Selected Human Tumors. Cell. 2019;178(2):346-60 e24.
Alspach E, Lussier DM, Schreiber RD. Interferon γ and its important roles in promoting and inhibiting spontaneous and therapeutic cancer immunity. Cold Spring Harb Perspect Biol. 2019;11(3):a028480.
Ferreira LMR, Muller YD, Bluestone JA, Tang Q. Next-generation regulatory T cell therapy. Nat Rev Drug Discov. 2019;18(10):749–69.
Joly E, Hudrisier D. What is trogocytosis and what is its purpose? Nat Immunol. 2003;4(9):815.
Young KJ, Zhang L. The nature and mechanisms of DN regulatory T-cell mediated suppression. Hum Immunol. 2002;63(10):926–34.
Ford McIntyre MS, Young KJ, Gao J, Joe B, Zhang L. Cutting edge: in vivo trogocytosis as a mechanism of double negative regulatory T cell-mediated antigen-specific suppression. J Immunol. 2008;181(4):2271–5.
Fischer K, Voelkl S, Heymann J, Przybylski GK, Mondal K, Laumer M, et al. Isolation and characterization of human antigen-specific TCRαβ+ CD4-CD8- double-negative regulatory T cells. Blood. 2005;105(7):2828–35.
Voelkl S, Gary R, Mackensen A. Characterization of the immunoregulatory function of human TCR-alphabeta+ CD4- CD8- double-negative T cells. Eur J Immunol. 2011;41(3):739–48.
Zhang ZX, Ma Y, Wang H, Arp J, Jiang J, Huang X, et al. Double-negative T cells, activated by xenoantigen, lyse autologous B and T cells using a perforin/granzyme-dependent Fas-Fas ligand-independent pathway. J Immunol. 2006;177(10):6920–9.
Haug T, Aigner M, Peuser MM, Strobl CD, Hildner K, Mougiakakos D, et al. Human Double-Negative Regulatory T-Cells Induce a Metabolic and Functional Switch in Effector T-Cells by Suppressing mTOR Activity. Front Immunol. 2019;10:883.
Zhang ZX, Stanford WL, Zhang L. Ly-6A is critical for the function of double negative regulatory T cells. Eur J Immunol. 2002;32(6):1584–92.
Chen W, Ford MS, Young KJ, Zhang L. Infusion of in vitro-generated DN T regulatory cells induces permanent cardiac allograft survival in mice. Transplant Proc. 2003;35(7):2479–80.
Lee BP, Chen W, Shi H, Der SD, Forster R, Zhang L. CXCR5/CXCL13 interaction is important for double-negative regulatory T cell homing to cardiac allografts. J Immunol. 2006;176(9):5276–83.
Wiener A, Schippers A, Wagner N, Tacke F, Ostendorf T, Honke N, et al. CXCR5 is critically involved in progression of lupus through regulation of B cell and double-negative T cell trafficking. Clin Exp Immunol. 2016;185(1):22–32.
Young KJ, Yang L, Phillips MJ, Zhang L. Donor-lymphocyte infusion induces transplantation tolerance by activating systemic and graft-infiltrating double-negative regulatory T cells. Blood. 2002;100(9):3408–14.
Loupy A, Lefaucheur C. Antibody-mediated rejection of solid-organ allografts. N Engl J Med. 2018;379(12):1150–60.
Gladstone DE, Bettinotti MP. HLA donor-specific antibodies in allogeneic hematopoietic stem cell transplantation: challenges and opportunities. Hematology Am Soc Hematol Educ Program. 2017;2017(1):645–50.
Ma Y, He KM, Garcia B, Min W, Jevnikar A, Zhang ZX. Adoptive transfer of double negative T regulatory cells induces B-cell death in vivo and alters rejection pattern of rat-to-mouse heart transplantation. Xenotransplantation. 2008;15(1):56–63.
Ford McIntyre MS, Gao JF, Li X, Naeini BM, Zhang L. Consequences of double negative regulatory T cell and antigen presenting cell interaction on immune response suppression. Int Immunopharmacol. 2011;11(5):597–603.
Hu SH, Zhang LH, Gao J, Guo JH, Xun XD, Xiang X, et al. NKG2D Enhances Double-Negative T Cell Regulation of B Cells. Front Immunol. 2021;12:650788.
Gao JF, McIntyre MS, Juvet SC, Diao J, Li X, Vanama RB, et al. Regulation of antigen-expressing dendritic cells by double negative regulatory T cells. Eur J Immunol. 2011;41(9):2699–708.
Hill M, Thebault P, Segovia M, Louvet C, Beriou G, Tilly G, et al. Cell therapy with autologous tolerogenic dendritic cells induces allograft tolerance through interferon-gamma and epstein-barr virus-induced gene 3. Am J Transplant. 2011;11(10):2036–45.
Chang YJ, Zhao XY, Huang XJ. Strategies for enhancing and preserving anti-leukemia effects without aggravating graft-versus-host disease. Front Immunol. 2018;9:3041.
Ye H, Chang Y, Zhao X, Huang X. Characterization of CD3+CD4-CD8- (double negative) T cells reconstitution in patients following hematopoietic stem-cell transplantation. Transpl Immunol. 2011;25(4):180–6.
Hillhouse EE, Thiant S, Moutuou MM, Lombard-Vadnais F, Parat R, Delisle JS, et al. Double-negative t cell levels correlate with chronic graft-versus-host disease severity. Biol Blood Marrow Transplant. 2019;25(1):19–25.
McIver Z, Serio B, Dunbar A, O’Keefe CL, Powers J, Wlodarski M, et al. Double-negative regulatory T cells induce allotolerance when expanded after allogeneic haematopoietic stem cell transplantation. Br J Haematol. 2008;141(2):170–8.
Soares F, Chen B, Lee JB, Ahmed M, Ly D, Tin E, et al. CRISPR screen identifies genes that sensitize AML cells to double-negative T-cell therapy. Blood. 2021;137(16):2171–81.
Chen B, Lee JB, Kang H, Minden MD, Zhang L. Targeting chemotherapy-resistant leukemia by combining DNT cellular therapy with conventional chemotherapy. J Exp Clin Cancer Res. 2018;37(1):88.
Lee JB, Khan DH, Hurren R, Xu M, Na Y, Kang H, et al. Venetoclax enhances T cell-mediated antileukemic activity by increasing ROS production. Blood. 2021;138(3):234–45.
Fang L, Ly D, Wang SS, Lee JB, Kang H, Xu H, et al. Targeting late-stage non-small cell lung cancer with a combination of DNT cellular therapy and PD-1 checkpoint blockade. J Exp Clin Cancer Res. 2019;38(1):123.
Kang H, Lee JB, Khatri I, Na Y, D’Souza C, Arruda A, et al. Enhancing therapeutic efficacy of double negative t cells against acute myeloid leukemia using idelalisib. Cancers (Basel). 2021;13(20):5039.
Li Y, Dong K, Fan X, Xie J, Wang M, Fu S, et al. DNT cell-based immunotherapy: progress and applications. J Cancer. 2020;11(13):3717–24.
Acknowledgements
The authors thank American Journal Experts for their assistance in editing this manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (grant # 81670165; # 82100230), International Cooperation Projects in Anhui Province (grant # 1804b06020352), and the Fundamental Research Funds for the Central Universities (grant # WK9110000060, WK9110000204 and WK9110000168).
Author information
Authors and Affiliations
Contributions
X.C. wrote the manuscript with D.W. X. Z. supervised the project, conceived the project and provided crucial ideas. The author(s) read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Chen, X., Wang, D. & Zhu, X. Application of double-negative T cells in haematological malignancies: recent progress and future directions. Biomark Res 10, 11 (2022). https://doi.org/10.1186/s40364-022-00360-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40364-022-00360-w