
Abstract
Background and objective
Studies have shown that Caveolin gene polymorphisms (CAV-1) are involved in chronic diseases, such as metabolic syndrome. Moreover, the dietary insulin index (DII) and dietary insulin load (DIL) have been shown to potentially elicit favorable effects on cardiovascular disease (CVD) risk. Therefore, this study sought to investigate the effect of DII DIL and CAV-1 interaction on CVD risk factors.
Methods
This cross-sectional study consisted of 333 overweight and obese women aged 18–48 years. Dietary intakes, DII, and DIL were evaluated using the 147-item food frequency questionnaire (FFQ). Serum profiles were measured by standard protocols. The CAV-1 rs 3,807,992 and anthropometric data were measured by the PCR–RFLP method and bioelectrical impedance analysis (BIA), respectively. Participants were also divided into three groups based on DII, DIL score, and rs3807992 genotype.
Results
This comparative cross-sectional study was conducted on 333 women classified as overweight or obese. Participants with A allele for the caveolin genotype and higher DII score showed significant interactions with high-density lipoprotein (HDL) (P for AA = 0.006 and P for AG = 0.019) and CRI-I (P for AA < 0.001 and P for AG = 0.024). In participants with AA genotype and greater DII score, interactions were observed in weight, systolic blood pressure (SBP), diastolic blood pressure (DBP), total cholesterol, CRI-II, fat-free mass (FFM), and skeletal muscle mass (SMM) (P < 0.079). Those with higher DIL scores and AA genotype had higher weight (P = 0.033), FFM (P = 0.022), and SMM (P = 0.024). In addition, DIL interactions for waist/hip ratio (WHR), waist circumference (WC), triglyceride (TG), CRI-I, and body fat mass (BFM) among individuals with AA genotype, while an HDL interaction was observed in individuals with AG and AA (P < 0.066).
Conclusion
The findings of the present study indicate that people who carry the caveolin rs3807992 (A) allele and have greater DII and DIL scores are at higher risk for several cardiovascular disease and metabolic syndrome biomarkers. These results highlight that diet, gene variants, and their interaction, should be considered in the risk evaluation of developing CVD.
Similar content being viewed by others

Introduction
Obesity is a chronic disease whose global prevalence has nearly tripled in the last four decades, affecting more women than men in developed and developing countries [1,2,3]. The obese population includes metabolically healthy and metabolically unhealthy individuals [4]. Indeed, it is not clear whether obesity causes metabolic syndrome as it is also observed in lean people [5]. However, obesity is related to high levels of oxidative stress, which plays an imperative role in the pathogenesis of numerous diseases [6,7,8], and obesity and insulin resistance (IR) are the factors of the metabolic syndrome that contribute the most its relationship with oxidative stress [6]. Increased IR and oxidative stress in obesity result from changes in several factors, including dietary patterns and genetic backgrounds [9,10,11,12]. Dietary patterns are key components associated with a higher incidence of obesity, particularly those containing fast-absorbing carbohydrates, which increase insulin concentrations attributed to hyperinsulinemia and IR [13]. Two proven methods to characterize higher insulin concentrations in serum are the dietary insulin load (DIL) and dietary insulin index (DII), which are also associated with obesity and metabolic syndrome risk factors, indicating that those with a higher score of the aforementioned indexes have greater insulin concentrations [14,15,16].
Regardless of carbohydrate intake, other macronutrients (proteins and fats) are also involved in insulin secretion [17]. Although they do not raise glucose concentrations as much, they increase fructose, some amino acids, and fatty acids, enhancing insulin secretion [18, 19]. Recently, the food insulin index (FII) has been introduced to measure the insulin response to foods [20]. Indeed, FII is a more accurate way to predict postprandial insulin secretion than other methods [20]. Using the FII, the insulin response to the diet indicated by DII and DIL can be calculated [21, 22]. This is important as high DII and DIL scores, especially in women, are associated with an increased incidence of obesity [14] and metabolic syndrome [16]. Research to date has also shown important associations between DII and DIL with certain glycemic, lipid, and inflammatory markers. For instance, the DIL score was positively associated with fasting blood glucose (FBG) and C-reactive protein (CRP) in older men [23]. In adolescents, a higher DII score was associated with higher IR and food cravings [24]. Moreover, both the DII and DIL are associated with plasma lipids (positively related to triglycerides and inversely related to HDL), especially in obese individuals [25]. The above-mentioned associations reinforce the importance of these dietary indices in populations with excess body fat and metabolic abnormalities.
Several genomes may have synergic effects with dietary patterns leading to a higher prevalence of obesity. Small pits 60–80 nm in diameter are found in the plasma membrane called caveola [26]. The caveola can be found in particular tissues such as adipocytes, vascular endothelial cells, muscle cells, epithelial cells, and fibroblasts [27]. Caveola has several functions, including regulating cholesterol and lipid metabolism, cellular signaling, mechanical protection, endocytosis, and a significant increase in cell surface area [28, 29]. The role of the caveolin gene on metabolic status can be explained by increasing the expression of CAV-1 mRNA in visceral and subcutaneous adipose tissue that is associated with obesity and its related disorders, such as type two diabetes (T2D) [30]. The association of CAV-1 rs3807992 and metabolic syndrome has been confirmed through the effects of CAV-1 on visceral fat and IR [31, 32]. CAV-1 and CAVIN-1 mRNA are highly expressed in visceral and subcutaneous adipose tissues, particularly among obese individuals, which may be associated with dyslipidemia [30], and atherosclerosis [33]. A high-fat diet increases caveolin expression [27], which can affect the insulin pathway [34, 35]. The relationship between CAV-1 and high-fat diets has been shown in knockout mice. The available investigations showed that mice with polymorphisms in this gene had resistance to weight gain, hyperinsulinemia, and accumulation of epididymal fat following a high-fat diet [36,37,38]. These findings indicate that caveolin gene polymorphisms are closely related to diets and metabolic factors in an obesity-promoting environment. Therefore, evaluating the associations between diet-dependent insulin response indices, metabolic components, and caveolin gene polymorphisms may be clinically relevant to employing successful prevention and treatment strategies in overweight and obese populations.
To date, no study has investigated the association of caveolin gene polymorphisms with DII and DIL. Given the role of caveolin in insulin signaling pathways, the present study aimed to investigate the interaction between caveolin genotypes with DII and DIL on metabolic components and body composition indices in overweight and obese women. The recent research underscores the importance of understanding the interplay between dietary inflammatory index (DII), caveolin-1 (CAV-1) expression, and metabolic markers. This study extends these insights by examining their relationships in a specific population, offering novel perspectives for dietary interventions. The integration of genetic factors like CAV-1 with dietary patterns provides a more comprehensive understanding of metabolic health in women, potentially guiding personalized dietary recommendations and management strategies for obesity and related metabolic disorders. Evaluation of several dietary macronutrients and micronutrients was also performed as these may be involved in insulin signaling and secretion.
Methods
Study population
A cross-sectional of 333 overweight and obese (body mass index [BMI]: 25–40 kg/m2) premenopausal (18–48 years) women who were referred to the health centers in Tehran were recruited as participants. Women who had a history of acute or chronic illnesses such as hypertension, cardiovascular diseases, diabetes, kidney disease, liver disease, cancers, and thyroid diseases, were pregnant or breastfeeding, smokers, regular alcohol consumers, and those who followed a diet with less than 800 kcal and more than 4,200 kcal were excluded from the study. Moreover, those who took medications or dietary supplements were also excluded. This study was conducted according to the Declaration of Helsinki. All procedures involving human participants were approved by the ethics committee of the Tehran University of Medical Sciences (IR.TUMS.VCR.REC.1398.142). Written informed consent was obtained from all participants.
The screening of potential participants took place between January 2019 and December 2020. We screened a total of 410 overweight and obese women. Of these, only 333 qualified for inclusion, as 77 individuals were excluded for reasons such as age (n = 7), pregnancy or lactation (n = 10), recent changes in diet (n = 8), and other chronic diseases (n = 29). Furthermore, subjects with missing data for biomarkers or covariates (n = 10), those who did not answer more than 70 questions (n = 3) on the FFQ, and those who reported an overall total daily energy intake outside the range of 800–4200 kcal (n = 10) were not included in the statistical analysis.
Anthropometric assessment and body composition
Anthropometrics and body composition measurements took place between 8–9 am after 12 h of overnight fasting. Participants were asked to avoid any strenuous physical activity for 72 h before the assessment. Furthermore, 30 min before the test, participants were asked to urinate (void) completely and avoid consuming water. Weight was measured using a digital scale (Seca 711; Seca, Hamburg, Germany) to the nearest 0.1 kg. Height was measured using a wall-mounted stadiometer (Seca 711; Seca, Hamburg, Germany). Waist circumference (WC) and hip circumference (HC) were determined at both the smallest and largest girths using standard anthropometric guidelines [39]. The waist-to-hip ratio (WHR) was obtained by dividing the waist circumference by the hip circumference. As a final point, we calculated BMI by dividing weight (kg) by the square of height (meters) [BMI = weight/height (kg/m2)]. Obesity and overweight were defined as BMI 30–40 kg/m2 and BMI 25–29.9 kg/m2, respectively. A multi-frequency bioelectrical impedance analyzer, InBody 770 scanner (test–re-test reliability: 0.980) according to manufacturer guidelines.
Biochemical assessments
Blood samples (10 mL) were taken following an overnight fast (12 h). The serum was separated by centrifuging and stored at a temperature of − 80 °C until the analysis was carried out. All measurements were taken at the nutrition laboratory of TUMS. Commercial kits (Pars Azmoon, Iran) were used to measure lipid profile and glucose. The enzyme-linked immunosorbent assay (ELISA) method (Human insulin ELISA kit, DRG Pharmaceuticals, GmbH, USA) was used to measure serum insulin concentrations.
Castelli risk indices I and II calculation
Castelli risk indices I (CRI-I) and Castelli risk indices II (CRI- II) were calculated by following the formula (TC/HDL-C) and (LDL-C / HDL-C), respectively [40].
Blood pressure measurement
Blood pressure was measured using an automated BP monitor (Omron) after ten minutes of sitting. Two measurements at 1-min intervals were collected and averaged.
Dietary measurements
To assess participants’ dietary intake and nutritional status over the past year, a semi-quantitative food frequency questionnaire (semi-FFQ) was used. This process was designed based on the Willett study that included 147 food items and standard serving sizes for each nutrient [41]. The reliability and validity of this FFQ from common Iranian foods have been previously described [42]. Household measures were used to convert the size of food consumption into grams [43]. Trained nutritionists filled out questionnaires. To find out the participants' daily intake of each food, the Nutritionist 4 software was used.
Calculation of DII and DIL
The FII was defined as The area under the curve, representing the food insulin response in a portion of 1000 kJ (239 kcal) of energy over 2 h divided by the area under the reference food curve over 2 h with the same amount of energy [18]. To determine the insulin index of each food in the semi-FFQ, we used the methods outlined in Bell's thesis [44]. For some foods that were exclusively in our questionnaire, we used the insulin index of similar foods described by the Bell study [44]. The insulin load of each food was first calculated separately by the following formula: Insulin index of each food × energy content of food/1 g × amount of food consumption (g/d). Insulin loads of the foods consumed in one day were added together, and the DIL was calculated. DII was obtained by dividing DIL by total energy consumption.
Genotyping
To determine the different genotypes of CAV-1 polymorphisms, DNA was extracted from serum, and then Polymerase Chain Reaction (PCR-R) Restriction Fragment Length Polymorphism of PCR products (RFLP) technique was used to examine cav 1 polymorphisms. Contrast primers were used in PCR: F:3′AGTATTGACCTGATTTGCCATG5′R:5′GTCTTCTGGAAAAAGCACATGA-3′0.1 μl of extracted DNA, 1 μl of Forwarding primers, 1 μl Revers primers, 7 μl of distilled water and 10 μl of Taq DNA polymerase Master Mix, making a total of 20 μl. PCR reactions in this solution were performed as follows: PCR was utilized to denature DNA templates for 3 min at 40 cycles, including one-minute denaturing at 94 °C, one-minute annealing at 42–50 °C, and angulation at 72 °C for two minutes. To separate the amplified DNA, we first use the Hin 1II(Nlalll) restriction enzyme at 37 °C overnight to digest it and then separate it by electrophoresis on an agarose gel (2%). The genotypes identified from the CAV-1 rs 3,807,992 variant are: uncut homozygous AA (213 bp), cut heterozygous AG (3 bands: 118 and 95 and 213 bp), and cut homozygous GG (2 bands: 118 and 95 bp) [45].
Physical activity assessment
The International Physical Activity Questionnaire (IPAC) was used to assess physical activity levels. This questionnaire evaluates the number of activities in leisure, housework, work, physical activity related to transportation, and sports in the last seven days. Using the data obtained from this questionnaire, we established metabolic equivalents (METS) and subsequently categorized the level of physical activity of the participants with the following classification: low, below 600 MET/h per week; moderate, 600 to 3000 MET/h per week; and high, more than 3000 MET/h per week [46].
Assessment of other variables
Economic status, a key covariate in our study, is assessed through a multi-faceted approach. This approach encompasses income levels, employment status, and educational attainment, reflecting a broad spectrum of socio-economic factors. The demographic information was gathered through a structured demographic questionnaire designed to systematically acquire pertinent personal details. Economic status, a focal point of the study, was meticulously assessed through inquiries encompassing annual income and property holdings. Participants were asked to disclose their employment status, educational attainment, marital status, smoking habits, medication history, and any previous experiences with significant trauma. This comprehensive approach ensured a thorough exploration of the participants’ socio-economic and personal backgrounds, contributing to the robustness of the collected data. By incorporating these varied aspects, our study offers a more nuanced analysis of how economic status intersects with dietary choices and metabolic health.
Statistical analysis
The data were analyzed using IBM SPSS version 23 software. We examined the normality of the data using the Kolmogorov–Smirnov test. Comparison between DII and DIL tertiles and different genotypes of the caveolin gene were evaluated using one-way analysis of variance (ANOVA) and analysis of covariance (ANCOVA). For the relationships, p < 0.05 was considered statistically significant. The general linear model (GLM) was employed to investigate the interaction between caveolin gene polymorphisms, DII and DIL. GG was considered as a reference. The genotypes were recorded and given GG, code 0, AG, code 1, and AA code 2. In Model 1. Interaction analysis data were adjusted for age, BMI, and physical activity, and in model 2, economic status, education level, marital status, and job were added. To detect the interaction significance, P < 0.1 was considered. The exact test was used for the Hardy–Weinberg Equilibrium (HWE) The Hardy–Weinberg equilibrium and comparison of categorical variables were assessed with the c2 test.
The selection of confounding variables was driven by a thoughtful consideration of factors that could influence the relationships under investigation. Recognizing the complexity of the interactions between dietary indices, genetic variants, and cardiometabolic risk factors, we specifically chose confounding variables that are established contributors to metabolic health [47, 48]. The age and BMI were included as essential covariates given their well-documented associations with both dietary patterns and cardiometabolic outcomes [49]. The total energy intake, a key determinant of nutritional status, was considered to control for variations in overall dietary consumption [50]. Additionally, physical activity was incorporated as a confounding variable, acknowledging its impact on metabolic health and potential to confound the observed associations [51]. This selection aimed to enhance the precision of our findings by accounting for the potential influence of these variables on the relationships of interest.
Results
Associations between DII and anthropometric indices, body composition, blood pressure, biochemical factors, and lipid ratios
Participants were divided into three groups based on DII scores: low, medium, and high. Marginally difference was indicated for HDL (P = 0.062), and CRI-I (P = 0.073). Those in the first tertile had higher HDL and lower CRI-I than those in the third tertile (48.27 ± 9.51 vs. 44.48 ± 10.65) and (3.88 ± 0.85 vs. 4.52 ± 2.24), respectively. After adjusting for age, BMI, total energy expenditure, and physical activity, a significant difference was observed in the job status of the participants (P = 0.031). (Table 1).
Associations between DIL and anthropometric indices, body composition, blood pressure, biochemical factors, physical activity, and lipid ratios:
Participants were divided into three groups based on DIL scores: low, medium, and high. A significant difference was observed in physical activity (P = 0.032). After adjusting for age, BMI, total energy expenditure, and physical activity, a significant difference was observed for diastolic blood pressure (DBP) (P = 0.014). Individuals in the first tertile had higher DBP than those in the third tertile (79.22 ± 9.2 vs. 77.10 ± 9.21) (Table 1).
Associations between anthropometric indices, body composition, blood pressure, biochemical factors, and lipid ratios with rs3807992 genotypes
In Table 2, the target population was divided based on different genotypes of the caveolin gene, which include GG (n = 88), AG (n = 78), and AA (n = 167). There was a significant difference between the three genotypes. Participants who carried AA in comparison with other genotypes had a higher height, CRI-I, and FFM (P < 0.05) in both the crude and model I (adjusted for age, BMI, total energy intake, and physical activity). In addition, insulin concentrations were higher in GG carriers compared to others (P = 0.042). Moreover, after adjusting for potential confounders, a significant link was observed between the genotypes and SMM (P = 0.023).
Associations between dietary intake components and DII tertile
As shown in Table 3, except for total fiber, glucose, galactose, fructose, sodium, calcium, and vitamin K, all of the other variables had significant associations among DII groups (P < 0.05). After adjusting for total energy, those who had higher DII also had greater intake of carbohydrates, SFA, iron, potassium, magnesium, phosphorus, vitamins E, C, B1, B6, and B9; while the intake of total fat, monounsaturated fatty acid (MUFA), polyunsaturated fatty acid (PUFA), vitamin A, D, B12, and caffeine was lower in compared with other tertiles (Table 3).
Associations between dietary intake components and DIL tertile
After adjusting for total energy intake, a significant association was observed between DIL and intake of carbohydrates, total fat, MUFA, PUFA, vitamin B1, K, E, zinc, magnesium, phosphorus, and iron (P < 0.05). This was positively associated with the intake of these components, wherein the individuals with a higher index of DIL had a higher intake (Table 3).
Interactions between DII and caveolin rs3807992 genotypes on Metabolic components
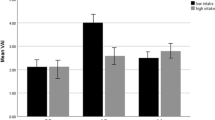
An Allele carrier showed significant interactions with HDL and CRI-I. Participants who scored higher for DII had lower HDL concentrations than the reference population (GG) (genotype AA (β = − 2.95, 95% CI (− 5.02, − 0.87), P = 0.006) and genotype AG (β = − 2.70, 95% CI (− 4.96, − 0.44), P = 0.019)). CRI-I indicated a significant interaction with DII in AA and AG genotypes, where the AA genotype (β = 0.67, 95% CI (0.34, 1.0), P ≤ 0.001), as well as the AG genotype (β = 0.41, 95% CI (0.05, 0.77), P = 0.024), had a high CRI-I index. In participants with AA genotype and those with higher DII score, significant interactions were observed in body mass (β = 1.86, 95% CI (− 0.24, 3.97), P = 0.078), systolic blood pressure (SBP) (β = 2.9, 95% CI (0.29, 5.52), P = 0.030), diastolic blood pressure (DBP) (β = 2.0, 95% CI (0.19, 3.82), P = 0.031), CRI-II (β = 0.14, 95% CI (0.01, 0.26), P = 0.030), FFM (β = 1.10, 95% CI (0.04, 2.17), P = 0.042), SMM (β = 0.66, 95% CI (0.03, 1.30), P = 0.040) and total cholesterol (β = 6.95, 95% CI (− 0.47, 4.37), P = 0.067). There was also a significant interaction for insulin in the AG genotype, β = − 0.07, 95% CI (− 0.14, − 0.03), and P = 0.042 (Table 4 and Fig. 1).


Interaction between Cav-1 genotype with dietary insulin index (DII) on: A Body Mass, B Systolic Blood Pressure(SBP), C Diastolic Blood Pressure(DBP), D Fat Free Mass(FFM), E Skeletal Muscle Mass(SMM), F Total Cholesterol (TC), G High-Density Lipoprotein (HDL), H CRI-I, I CRI-II, J insulin
Interactions between DIL and caveolin rs3807992 genotypes on metabolic components
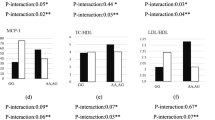
Those with higher DIL adherence and AA genotype had higher body mass (β = 2.37, 95% CI (0.19, 4.55), P = 0.033), FFM (β = 1.29, 95% CI (0.9, 2.39), P = 0.022), and SMM (β = 0.76, 95% CI (0.1, 1.41), P = 0.024). In addition, we observed interactions for WHR (β = 0.01, 95% CI (0.0, 0.02), P = 0.065), WC (β = 2.0, 95% CI (0.13, 3.86), P = 0.036), TG (β = 0.1, 95% CI (0.0, 0.20), P = 0.047) CRI-I (β = 0.36, 95% CI (0.01, 0.70), P = 0.029)and BFM (β = 1.41, 95% CI (− 0.11, 2.94), P = 0.069) in individuals with AA genotype, as well as for HDL in those with AG (β = − 2.21, 95% CI (− 4.52, 0.10), P = 0.061) and AA (β = − 2.38, 95% CI (− 4.52, − 0.24), P = 0.029) (Table 4 and Fig. 2).


Interaction between CAV-1 genotype with Dietary Insulin Load (DIL) on: A Body Mass, B Waist Hip Ratio (WHR), C Waist Circumference (WC), D Fat Free Mass (FFM), E Body Fat Mass(BFM), F High Density Lipoprotein (HDL), G Triglyceride (TG), H CRI-I, I Skeletal Muscle Mass (SMM)
Discussion
To our knowledge, this is the first study to investigate the interactions of caveolin gene polymorphisms with DII and DIL. Our results indicate that overweight and obese women with elevated DII had lower HDL and higher CRI-I. Moreover, those with higher DIL scores differed significantly in physical activity and DBP. An interaction was observed between DII and caveolin gene polymorphisms on HDL and CRI variables, while another interaction was detected between DII and caveolin on SBP, DBP, FFM, SMM, TC, and Insulin. DIL and rs 3,807,792 CAV-1 polymorphism had a significant interaction on BM, FFM, and SMM variables. In addition, an interaction was observed on WHR, WC, TG, CRI -I, and BFM.
The DII and DIL have recently received substantial attention as these indexes reflect the insulin response associated with different food groups [52, 53]. In this study, participants with higher DII showed lower HDL and higher CRI-I. Nimptsch et al. demonstrated that DII and DIL were not associated with glycemic control and inflammatory markers. At the same time, there were significant positive and negative relationships between TG and HDL with these dietary indexes, respectively. These relationships intensified with increasing body fat [54]. Also, DII and DIL showed a significant negative relationship with HDL among obese people [54]. Given that CRI-I is obtained from the ratio of TC to HDL, we can justify a significant increase in CRI-I as a result of a decrease in HDL.
We also found that individuals who were in the third DIL score tertile had significantly lower DBP than those in the first tertile (77.10 ± 9.21 vs. 79.22 ± 9.2). This may be related to differences in protein intake (Table 3.), as those who had a higher DIL score (third tertile) also reported a higher protein intake (111.91 ± 23.99 g/d vs. 64.65 ± 16.24 g/d). Based on the results of observational studies, high protein consumption is inversely related to blood pressure [55]. Moreover, the previous interventional studies showed that higher protein consumption is associated with a decrease in SBP and DBP [56, 57]. The mechanisms of action are likely related to increases in the intake of arginine (which is a precursor to the vasodilator nitric oxide), biologically active peptides like angiotensin-converting enzyme inhibitors, and foods that are correlated with higher protein consumption as well as lower blood pressure, such as isoflavonoids and soy protein [58].
Based on the previous studies, DII and DIL are associated with anthropometric and metabolic indices. The DONALD cohort (Dortmund Nutritional and Anthropometric Longitudinally Designed Study) measured the relationship between food intake recorded during adolescence and anthropometric indices in adulthood. This investigation found that higher DII and DIL during adolescence are associated with higher body fat percentage in adulthood [59]. However, no such relationship was detected with BMI, hyperinsulinemia, and insulin resistance [59]. In a cohort study with a population of 927 males and females, higher DII was associated with an elevated risk of insulin resistance during a three-year follow-up [15]. High DII stimulates insulin secretion in various ways, such as inhibiting fat oxidation, facilitating carbohydrate oxidation, increasing fat storage, and promoting obesity [60]. Studies have shown that diets that reduce insulin secretion can effectively control chronic diseases and improve body composition [15, 61]. In a study by Sadeghi et al., DII and DIL were associated with a higher risk of metabolic syndrome in women, while no association was observed in men [16]. In agreement with this study, Shoaa et al. concluded that DII was positively associated with abdominal obesity in women [14]. The current study was conducted on women only, and consequently, body composition and appetite may be attributed to gonadal steroids [62]. Indeed, changes in estrogen concentrations may alter hypothalamic pathways, subsequently affecting the production of various hormones that affect appetite [63].
According to our results, the consumption of macronutrients and micronutrients was significantly and positively associated with DII and DIL scores. Participants in our study with greater DII and DIL scores had a higher consumption of carbohydrates, protein, fat, vitamin B1, vitamin B6, magnesium, iron, etc., which was similar to the study of Sadeghi et al. [16]. Carbohydrates are well-established as the main factor in insulin secretion. However, evidence also supports the effects of proteins and fats in this process [17,18,19]. Indeed, these 3 macronutrients were significantly greater in those who had higher DII and DIL scores. Some minerals and vitamins, such as iron, magnesium, and phosphorus, which were also significant in our results, are also involved in insulin signaling [64]. Vitamin D plays a role in insulin secretion and signaling [65], but participants who had a higher DII score had a lower intake of vitamin D, which is probably due to the limited food sources of this vitamin in Iran [66].
The pathogenesis of the metabolic syndrome and its components may be influenced by interactions between genes and nutrients [67]. In the present study, those with the AA genotype had a significantly greater height, FFM, SMM, and CRI-I and lower plasma insulin concentrations than the population with the reference genotype (GG). Recently, certain genetic strains of CAV-1 have been associated with insulin resistance and hypertriglyceridemia [68]. Increased expression of CAV-1 is linked to the minor allele A [69], and reduced expression of CAV-1 can influence aldosterone and mineralocorticoid receptor signaling in various pathways connected to glycemia and dyslipidemia [70, 71]. Because of the key role of CAV-1 in pancreatic beta cells, it is well established that CAV-1 is involved in energy metabolism disorders such as insulin resistance and hypertriglyceridemia [72].
A significant interaction between HDL and CRI-I variables was observed in the present study between the rs 3,807,992 variant of the caveolin-1 gene and DII. For DIL, the interaction was significant between body composition variables such as body mass, SMM, and FFM. Abaj et al. found that overweight and obese women who carried the A allele had higher BMI, and lower TC, HDL, and LDL. The significant interactions we identified between DIL and the rs3807992 CAV-1 polymorphism about body composition variables, including BM, FFM, SMM, WHR, WC, TG, and BFM, underscore the importance of considering genetic influences when assessing the impact of dietary factors on body composition. Our findings highlight a significant interaction between dietary inflammation, as quantified by DII, and CAV-1 in modulating metabolic health. This interaction sheds light on the complex mechanisms underpinning metabolic disorders in overweight and obese women. The study's implications extend to clinical practice, suggesting that dietary interventions tailored to individual genetic profiles may enhance metabolic outcomes. Furthermore, our research paves the way for future studies exploring the gene-diet interface in metabolic health. In addition, those with higher adherence to a healthy diet pattern had higher HDL and lower hs-CRP concentrations [67], which is consistent with the results of our investigation. In another study by Abaj et al., it was found that consuming more PUFAs could weaken the association between rs 3,807,992 and metabolic syndrome while consuming saturated fatty acids reinforces this association [45]. Diet and nutrients can alter metabolic biomarkers by interacting with caveola-related cellular signaling [73]. Increasing the expression of CAV-1 by reducing the production of nitric oxide (NO) leads to long-term exposure to glucose, which plays an important role in strengthening the inflammatory pathways [74]. Diets with a lower inflammatory load can transport CAV-1 from the caveola to the cytoplasm and impede the inhibitory effects of CAV-1 on endothelial nitric oxide synthase (eNOS) and HDL receptors [75]. Sodium and potassium can also affect the binding of eNOS to the caveola membrane, so changing these two nutrients in diets can also affect the function of CAV-1 [76]. Pojoga et al. found that CAV-1 deficiency was associated with high blood pressure, hyperglycemia, and decreased vasoconstriction [77]. A study showed that CAV-1(+ / +) mice on a high-cholesterol diet had less TC and TG than CAV-1 (- / -) mice [78]. Thus, the expression of CAV-1 can be tightly associated with the intake of some macro and micronutrients in a dietary pattern which may also affect several metabolic components. The observed interactions between DII, CAV-1, and various metabolic parameters, such as HDL, CRI-I, SBP, DBP, FFM, SMM, TC, and insulin, provide insight into the interplay of dietary choices and genetic factors in shaping metabolic outcomes. The interaction between DII and CAV-1 highlights the potential role of dietary insulin response in modulating insulin sensitivity and related metabolic components. Elevated DII scores were associated with lower HDL and higher CRI-I, indicating that the dietary insulin response may play a crucial role in lipid profiles and cardiovascular risk.
CAV-1 is involved in insulin secretion, insulin resistance, and insulin signaling [74] by mediating insulin receptors [79]. These receptors are mainly located in areas of the plasma membrane rich in caveolae and cav-1 and play an important role in insulin signaling and secretion [80, 81]. A homozygous polymorphism in the CAV-1 gene can cause congenital generalized human lipodystrophy type 3 (CGL3), which causes severe IR [82]. CAV-1 variants are also associated with IR [83]. During IR, the insulin receptor detaches from cav-1, and insulin signaling is disrupted, leading to a decreased Glucose transporter type 4 (GLUT 4) transport to the membrane, which reduces insulin sensitivity and glucose uptake [81]. The CAV-1 gene is located in region 7q 31, area 7q 31, and its vicinity is associated with IR, blood pressure, and some vascular conditions [79,80,86]. The above may be a mechanism for the role of caveolin in hyperinsulinemia. Hyperinsulinemia causes a rise in DII and DIL scores, which logically explains our current outcomes.
In summary, our study contributes to a growing body of evidence highlighting the dynamic relationship between dietary indices, genetic factors, and metabolic markers. Understanding how DII and DIL interact with CAV-1 and impact metabolic outcomes is a crucial step toward more precise dietary interventions and prevention strategies for individuals at risk of metabolic abnormalities. Our findings underscore the potential clinical relevance of this gene-diet interaction and provide a foundation for further research in the field. This was the first study to examine gene interaction with DII and DIL among overweight and obese women. Despite its novelty, our study has certain limitations. Due to the cross-sectional design of the study, any causality cannot be argued. Further studies are needed to determine the exact interaction of the caveolin gene with DII and DIL. Performing clinical trials to establish the effects of diets with low DII and DIL on caveolin gene expression and metabolic components will help expand knowledge on this topic.
Conclusions
In summary, the results of this study demonstrate that overweight and obese women who had high DII and DIL scores, as well as those who were at risk for the caveolin gene allele (a), had higher body mass, FFM, SMM, TC, and CRI, as well as lower HDL concentrations. In addition, DII had a positive interaction with SBP, DBP, and CRI -II, and a negative interaction with insulin. A positive correlation with WHR, WC, BFM, and TG for DIL was also observed. These outcomes indicate that those who carry the caveolin rs3807992 (A) allele and have greater DII and DIL are at higher risk for cardiovascular disease and metabolic syndrome.
Availability of data and materials
The datasets analyzed during the current study available from the corresponding author on reasonable request.
References
Mauvais-Jarvis F. Sex differences in metabolic homeostasis, diabetes, and obesity. Biol Sex Differ. 2015;6:14. https://doi.org/10.1186/s13293-015-0033-y.
Blüher M. Obesity: global epidemiology and pathogenesis. Nat Rev Endocrinol. 2019;15:288–98. https://doi.org/10.1038/s41574-019-0176-8.
Ng M, Fleming T, Robinson M, Thomson B, Graetz N, Margono C, Mullany EC, Biryukov S, Abbafati C, Abera SF, et al. Global, regional, and national prevalence of overweight and obesity in children and adults during 1980–2013: a systematic analysis for the Global Burden of Disease Study 2013. Lancet (London, England). 2014;384:766–81. https://doi.org/10.1016/s0140-6736(14)60460-8.
Stefan N, Häring HU, Hu FB, Schulze MB. Metabolically healthy obesity: epidemiology, mechanisms, and clinical implications. Lancet Diabetes Endocrinol. 2013;1:152–62. https://doi.org/10.1016/s2213-8587(13)70062-7.
Lee SC, Hairi NN, Moy FM. Metabolic syndrome among non-obese adults in the teaching profession in Melaka, Malaysia. J Epidemiol. 2017;27:130–4. https://doi.org/10.1016/j.je.2016.10.006.
Jakubiak GK, Osadnik K, Lejawa M, Kasperczyk S, Osadnik T, Pawlas N. Oxidative stress in association with metabolic health and obesity in young adults. Oxid Med Cell Longev. 2021;2021:9987352. https://doi.org/10.1155/2021/9987352.
Dubois-Deruy E, Peugnet V, Turkieh A, Pinet F. Oxidative stress in cardiovascular diseases. Antioxidants (Basel, Switzerland). 2020. https://doi.org/10.3390/antiox9090864.
Hayes JD, Dinkova-Kostova AT, Tew KD. Oxidative stress in cancer. Cancer Cell. 2020;38:167–97. https://doi.org/10.1016/j.ccell.2020.06.001.
Gasmi A, Noor S, Menzel A, Doşa A, Pivina L, Bjørklund G. Obesity and insulin resistance: associations with chronic inflammation, genetic and epigenetic factors. Curr Med Chem. 2021;28:800–26. https://doi.org/10.2174/0929867327666200824112056.
Tan BL, Norhaizan ME. Effect of high-fat diets on oxidative stress, cellular inflammatory response and cognitive function. Nutrients. 2019. https://doi.org/10.3390/nu11112579.
Napoleão A, Fernandes L, Miranda C, Marum AP. Effects of calorie restriction on health span and insulin resistance: classic calorie restriction diet vs. ketosis-inducing diet. Nutrients. 2021. https://doi.org/10.3390/nu13041302.
Tettamanzi F, Bagnardi V, Louca P, Nogal A, Monti GS, Mambrini SP, Lucchetti E, Maestrini S, Mazza S, Rodriguez-Mateos A, et al. A high protein diet is more effective in improving insulin resistance and glycemic variability compared to a mediterranean diet-a cross-over controlled inpatient dietary study. Nutrients. 2021. https://doi.org/10.3390/nu13124380.
Liu S, Willett WC, Stampfer MJ, Hu FB, Franz M, Sampson L, Hennekens CH, Manson JE. A prospective study of dietary glycemic load, carbohydrate intake, and risk of coronary heart disease in US women. Am J Clin Nutr. 2000;71:1455–61. https://doi.org/10.1093/ajcn/71.6.1455.
Anjom-Shoae J, Keshteli AH, Sadeghi O, Pouraram H, Afshar H, Esmaillzadeh A, Adibi P. Association between dietary insulin index and load with obesity in adults. Eur J Nutr. 2020;59:1563–75. https://doi.org/10.1007/s00394-019-02012-6.
Mirmiran P, Esfandiari S, Bahadoran Z, Tohidi M, Azizi F. Dietary insulin load and insulin index are associated with the risk of insulin resistance: a prospective approach in tehran lipid and glucose study. J Diabetes Metab Disord. 2015;15:23. https://doi.org/10.1186/s40200-016-0247-5.
Sadeghi O, Hasani H, Mozaffari-Khosravi H, Maleki V, Lotfi MH, Mirzaei M. Dietary insulin index and dietary insulin load in relation to metabolic syndrome: the Shahedieh cohort study. J Acad Nutr Diet. 2020;120:1672-1686.e1674. https://doi.org/10.1016/j.jand.2020.03.008.
Moghaddam E, Vogt JA, Wolever TM. The effects of fat and protein on glycemic responses in nondiabetic humans vary with waist circumference, fasting plasma insulin, and dietary fiber intake. J Nutr. 2006;136:2506–11. https://doi.org/10.1093/jn/136.10.2506.
Nuttall FQ, Gannon MC. Plasma glucose and insulin response to macronutrients in nondiabetic and NIDDM subjects. Diabetes Care. 1991;14:824–38. https://doi.org/10.2337/diacare.14.9.824.
Ranganath L, Norris F, Morgan L, Wright J, Marks V. The effect of circulating non-esterified fatty acids on the entero-insular axis. Eur J Clin Invest. 1999;29:27–32. https://doi.org/10.1046/j.1365-2362.1999.00426.x.
Holt SH, Miller JC, Petocz P. An insulin index of foods: the insulin demand generated by 1000-kJ portions of common foods. Am J Clin Nutr. 1997;66:1264–76. https://doi.org/10.1093/ajcn/66.5.1264.
Bao J, de Jong V, Atkinson F, Petocz P, Brand-Miller JC. Food insulin index: physiologic basis for predicting insulin demand evoked by composite meals. Am J Clin Nutr. 2009;90:986–92. https://doi.org/10.3945/ajcn.2009.27720.
Anjom-Shoae J, Shayanfar M, Mohammad-Shirazi M, Sadeghi O, Sharifi G, Siassi F, Esmaillzadeh A. Dietary insulin index and insulin load in relation to glioma: findings from a case-control study. Nutr Neurosci. 2021;24:354–62. https://doi.org/10.1080/1028415x.2019.1631594.
Mozaffari H, Namazi N, Larijani B, Surkan PJ, Azadbakht L. Associations between dietary insulin load with cardiovascular risk factors and inflammatory parameters in elderly men: a cross-sectional study. Br J Nutr. 2019;121:773–81. https://doi.org/10.1017/s0007114518003872.
Bao Y, Nimptsch K, Wolpin BM, Michaud DS, Brand-Miller JC, Willett WC, Giovannucci E, Fuchs CS. Dietary insulin load, dietary insulin index, and risk of pancreatic cancer. Am J Clin Nutr. 2011;94:862–8. https://doi.org/10.3945/ajcn.110.011205.
Caferoglu Z, Hatipoglu N, Gokmen Ozel H. Does food insulin index in the context of mixed meals affect postprandial metabolic responses and appetite in obese adolescents with insulin resistance? A randomised cross-over trial. Br J Nutr. 2019;122:942–50. https://doi.org/10.1017/s0007114519001351.
Bastiani M, Parton RG. Caveolae at a glance. J Cell Sci. 2010;123:3831–6. https://doi.org/10.1242/jcs.070102.
Thorn H, Stenkula KG, Karlsson M, Ortegren U, Nystrom FH, Gustavsson J, Stralfors P. Cell surface orifices of caveolae and localization of caveolin to the necks of caveolae in adipocytes. Mol Biol Cell. 2003;14:3967–76. https://doi.org/10.1091/mbc.e03-01-0050.
Shvets E, Ludwig A, Nichols BJ. News from the caves: update on the structure and function of caveolae. Curr Opin Cell Biol. 2014;29:99–106. https://doi.org/10.1016/j.ceb.2014.04.011.
Parton RG, Simons K. The multiple faces of caveolae. Nat Rev Mol Cell Biol. 2007;8:185–94. https://doi.org/10.1038/nrm2122.
Catalán V, Gómez-Ambrosi J, Rodríguez A, Silva C, Rotellar F, Gil MJ, Cienfuegos JA, Salvador J, Frühbeck G. Expression of caveolin-1 in human adipose tissue is upregulated in obesity and obesity-associated type 2 diabetes mellitus and related to inflammation. Clin Endocrinol. 2008;68:213–9. https://doi.org/10.1111/j.1365-2265.2007.03021.x.
Abaj F, Saeedy SAG, Mirzaei K. Are caveolin-1 minor alleles more likely to be risk alleles in insulin resistance mechanisms in metabolic diseases? BMC Res Notes. 2021;14:185. https://doi.org/10.1186/s13104-021-05597-6.
Abaj F, Saeedy SAG, Mirzaei K. Mediation role of body fat distribution (FD) on the relationship between CAV1 rs3807992 polymorphism and metabolic syndrome in overweight and obese women. BMC Med Genomics. 2021;14:202. https://doi.org/10.1186/s12920-021-01050-6.
Méndez-Giménez L, Rodríguez A, Balaguer I, Frühbeck G. Role of aquaglyceroporins and caveolins in energy and metabolic homeostasis. Mol Cell Endocrinol. 2014;397:78–92. https://doi.org/10.1016/j.mce.2014.06.017.
Gómez-Ruiz A, de Miguel C, Campión J, Martínez JA, Milagro FI. Time-dependent regulation of muscle caveolin activation and insulin signalling in response to high-fat diet. FEBS Lett. 2009;583:3259–64. https://doi.org/10.1016/j.febslet.2009.09.016.
Tan Z, Zhou LJ, Mu PW, Liu SP, Chen SJ, Fu XD, Wang TH. Caveolin-3 is involved in the protection of resveratrol against high-fat-diet-induced insulin resistance by promoting GLUT4 translocation to the plasma membrane in skeletal muscle of ovariectomized rats. J Nutr Biochem. 2012;23:1716–24. https://doi.org/10.1016/j.jnutbio.2011.12.003.
Lillo Urzúa P, Núñez Murillo O, Castro-Sepúlveda M, Torres-Quintana MA, Lladser Caldera Á, Quest AF, Espinoza Robles C, Llanos Vidal P, Wehinger S. Loss of caveolin-1 is associated with a decrease in beta cell death in mice on a high fat diet. Int J Mol Sci. 2020. https://doi.org/10.3390/ijms21155225.
Razani B, Combs TP, Wang XB, Frank PG, Park DS, Russell RG, Li M, Tang B, Jelicks LA, Scherer PE, et al. Caveolin-1-deficient mice are lean, resistant to diet-induced obesity, and show hypertriglyceridemia with adipocyte abnormalities. J Biol Chem. 2002;277:8635–47. https://doi.org/10.1074/jbc.M110970200.
Cohen AW, Razani B, Wang XB, Combs TP, Williams TM, Scherer PE, Lisanti MP. Caveolin-1-deficient mice show insulin resistance and defective insulin receptor protein expression in adipose tissue. Am J Physiol Cell Physiol. 2003;285:C222-235. https://doi.org/10.1152/ajpcell.00006.2003.
Madden AM, Smith S. Body composition and morphological assessment of nutritional status in adults: a review of anthropometric variables. J Hum Nutr Diet. 2016;29:7–25. https://doi.org/10.1111/jhn.12278.
Cai G, Shi G, Xue S, Lu W. The atherogenic index of plasma is a strong and independent predictor for coronary artery disease in the Chinese Han population. Medicine. 2017;96: e8058. https://doi.org/10.1097/md.0000000000008058.
Price DD, McGrath PA, Rafii A, Buckingham B. The validation of visual analogue scales as ratio scale measures for chronic and experimental pain. Pain. 1983;17:45–56. https://doi.org/10.1016/0304-3959(83)90126-4.
Mirmiran P, Esfahani FH, Mehrabi Y, Hedayati M, Azizi F. Reliability and relative validity of an FFQ for nutrients in the Tehran lipid and glucose study. Public Health Nutr. 2010;13:654–62. https://doi.org/10.1017/s1368980009991698.
Ghaffarpour M, Houshiar-Rad A, Kianfar H. The manual for household measures, cooking yields factors and edible portion of foods. Tehran Nashre Olume Keshavarzy. 1999;7:42–58.
Bell K. Clinical application of the food insulin index to diabetes mellitus. 2014.
Abaj F, Mirzaei K. Caveolin-1 genetic polymorphism interacts with PUFA to modulate metabolic syndrome risk. Br J Nutr. 2021. https://doi.org/10.1017/s0007114521002221.
Craig CL, Marshall AL, Sjöström M, Bauman AE, Booth ML, Ainsworth BE, Pratt M, Ekelund U, Yngve A, Sallis JF, et al. International physical activity questionnaire: 12-country reliability and validity. Med Sci Sports Exerc. 2003;35:1381–95. https://doi.org/10.1249/01.mss.0000078924.61453.fb.
Karlsson T, Rask-Andersen M, Pan G, Höglund J, Wadelius C, Ek WE, Johansson Å. Contribution of genetics to visceral adiposity and its relation to cardiovascular and metabolic disease. Nat Med. 2019;25(9):1390–5.
Drewnowski A, Henderson SA, Cockroft JE. Genetic sensitivity to 6-n-propylthiouracil has no influence on dietary patterns, body mass indexes, or plasma lipid profiles of women. J Am Diet Assoc. 2007;107(8):1340–8.
Bakhtiyari M, Kazemian E, Kabir K, Hadaegh F, Aghajanian S, Mardi P, Ghahfarokhi NT, Ghanbari A, Mansournia MA, Azizi F. Contribution of obesity and cardiometabolic risk factors in developing cardiovascular disease: a population-based cohort study. Sci Rep. 2022;12(1):1544.
Willett WC, Howe GR, Kushi LH. Adjustment for total energy intake in epidemiologic studies. Am J Clin Nutr. 1997;65(4 Suppl):1220S-1228S.
Park SK, Larson JL. The relationship between physical activity and metabolic syndrome in people with chronic obstructive pulmonary disease. J Cardiovasc Nurs. 2014;29(6):499–507.
Sung KC, Lee MY, Kim YH, Huh JH, Kim JY, Wild SH, Byrne CD. Obesity and incidence of diabetes: effect of absence of metabolic syndrome, insulin resistance, inflammation and fatty liver. Atherosclerosis. 2018;275:50–7. https://doi.org/10.1016/j.atherosclerosis.2018.05.042.
Bell KJ, Petocz P, Colagiuri S, Brand-Miller JC. Algorithms to improve the prediction of postprandial insulinaemia in response to common foods. Nutrients. 2016;8:210. https://doi.org/10.3390/nu8040210.
Nimptsch K, Brand-Miller JC, Franz M, Sampson L, Willett WC, Giovannucci E. Dietary insulin index and insulin load in relation to biomarkers of glycemic control, plasma lipids, and inflammation markers. Am J Clin Nutr. 2011;94:182–90. https://doi.org/10.3945/ajcn.110.009555.
Obarzanek E, Velletri PA, Cutler JA. Dietary protein and blood pressure. JAMA. 1996;275:1598–603. https://doi.org/10.1001/jama.1996.03530440078040.
He J, Gu D, Wu X, Duan X, Whelton P. Soybean protein supplementation and blood pressure: a randomized, controlled clinical trialop 159. J Hypertens. 2004;22:S169.
Burke V, Hodgson JM, Beilin LJ, Giangiulioi N, Rogers P, Puddey IB. Dietary protein and soluble fiber reduce ambulatory blood pressure in treated hypertensives. Hypertension (Dallas, Tex : 1979). 2001;38:821–6. https://doi.org/10.1161/hy1001.092614.
Appel LJ. The effects of protein intake on blood pressure and cardiovascular disease. Curr Opin Lipidol. 2003;14:55–9. https://doi.org/10.1097/00041433-200302000-00010.
Xu H, Barnes GT, Yang Q, Tan G, Yang D, Chou CJ, Sole J, Nichols A, Ross JS, Tartaglia LA, et al. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J Clin Investig. 2003;112:1821–30. https://doi.org/10.1172/jci19451.
Kahn SE, Hull RL, Utzschneider KM. Mechanisms linking obesity to insulin resistance and type 2 diabetes. Nature. 2006;444:840–6. https://doi.org/10.1038/nature05482.
Tay J, Thompson CH, Luscombe-Marsh ND, Wycherley TP, Noakes M, Buckley JD, Wittert GA, Yancy WS Jr, Brinkworth GD. Effects of an energy-restricted low-carbohydrate, high unsaturated fat/low saturated fat diet versus a high-carbohydrate, low-fat diet in type 2 diabetes: A 2-year randomized clinical trial. Diabetes Obes Metab. 2018;20:858–71. https://doi.org/10.1111/dom.13164.
Lovejoy JC, Sainsbury A. Sex differences in obesity and the regulation of energy homeostasis. Obes Rev. 2009;10:154–67. https://doi.org/10.1111/j.1467-789X.2008.00529.x.
Clegg DJ, Brown LM, Zigman JM, Kemp CJ, Strader AD, Benoit SC, Woods SC, Mangiaracina M, Geary N. Estradiol-dependent decrease in the orexigenic potency of ghrelin in female rats. Diabetes. 2007;56:1051–8. https://doi.org/10.2337/db06-0015.
Derakhshanian H, Javanbakht MH, Zarei M, Djalali E, Djalali M. Vitamin D increases IGF-I and insulin levels in experimental diabetic rats. Growth Horm IGF Res. 2017;36:57–9. https://doi.org/10.1016/j.ghir.2017.09.002.
Orwoll E, Riddle M, Prince M. Effects of vitamin D on insulin and glucagon secretion in non-insulin-dependent diabetes mellitus. Am J Clin Nutr. 1994;59:1083–7. https://doi.org/10.1093/ajcn/59.5.1083.
Vatandost S, Jahani M, Afshari A, Amiri MR, Heidarimoghadam R, Mohammadi Y. Prevalence of vitamin D deficiency in Iran: a systematic review and meta-analysis. Nutr Health. 2018;24:269–78. https://doi.org/10.1177/0260106018802968.
Abaj F, Koohdani F, Rafiee M, Alvandi E, Yekaninejad MS, Mirzaei K. Interactions between Caveolin-1 (rs3807992) polymorphism and major dietary patterns on cardio-metabolic risk factors among obese and overweight women. BMC Endocr Disord. 2021;21:138. https://doi.org/10.1186/s12902-021-00800-y.
Mora-García G, Gómez-Camargo D, Alario Á, Gómez-Alegría C. A common variation in the caveolin 1 gene is associated with high serum triglycerides and metabolic syndrome in an admixed Latin American population. Metab Syndr Relat Disord. 2018;16:453–63. https://doi.org/10.1089/met.2018.0004.
Chen S, Wang X, Wang J, Zhao Y, Wang D, Tan C, Fa J, Zhang R, Wang F, Xu C, et al. Genomic variant in CAV1 increases susceptibility to coronary artery disease and myocardial infarction. Atherosclerosis. 2016;246:148–56. https://doi.org/10.1016/j.atherosclerosis.2016.01.008.
Baudrand R, Gupta N, Garza AE, Vaidya A, Leopold JA, Hopkins PN, Jeunemaitre X, Ferri C, Romero JR, Williams J, et al. Caveolin 1 modulates aldosterone-mediated pathways of glucose and lipid homeostasis. J Am Heart Assoc. 2016. https://doi.org/10.1161/jaha.116.003845.
Ramírez CM, Zhang X, Bandyopadhyay C, Rotllan N, Sugiyama MG, Aryal B, Liu X, He S, Kraehling JR, Ulrich V, et al. Caveolin-1 regulates atherogenesis by attenuating low-density lipoprotein transcytosis and vascular inflammation independently of endothelial nitric oxide synthase activation. Circulation. 2019;140:225–39. https://doi.org/10.1161/circulationaha.118.038571.
Pol A, Martin S, Fernandez MA, Ferguson C, Carozzi A, Luetterforst R, Enrich C, Parton RG. Dynamic and regulated association of caveolin with lipid bodies: modulation of lipid body motility and function by a dominant negative mutant. Mol Biol Cell. 2004;15:99–110. https://doi.org/10.1091/mbc.e03-06-0368.
Majkova Z, Toborek M, Hennig B. The role of caveolae in endothelial cell dysfunction with a focus on nutrition and environmental toxicants. J Cell Mol Med. 2010;14:2359–70. https://doi.org/10.1111/j.1582-4934.2010.01064.x.
Haddad D, Al Madhoun A, Nizam R, Al-Mulla F. Role of caveolin-1 in diabetes and its complications. Oxid Med Cell Longev. 2020;2020:9761539. https://doi.org/10.1155/2020/9761539.
Matveev S, Uittenbogaard A, van Der Westhuyzen D, Smart EJ. Caveolin-1 negatively regulates SR-BI mediated selective uptake of high-density lipoprotein-derived cholesteryl ester. Eur J Biochem. 2001;268:5609–16. https://doi.org/10.1046/j.1432-1033.2001.02496.x.
Oberleithner H, Kusche-Vihrog K, Schillers H. Endothelial cells as vascular salt sensors. Kidney Int. 2010;77:490–4. https://doi.org/10.1038/ki.2009.490.
Pojoga LH, Yao TM, Opsasnick LA, Garza AE, Reslan OM, Adler GK, Williams GH, Khalil RA. Dissociation of hyperglycemia from altered vascular contraction and relaxation mechanisms in caveolin-1 null mice. J Pharmacol Exp Ther. 2014;348:260–70. https://doi.org/10.1124/jpet.113.209189.
Frank PG, Lee H, Park DS, Tandon NN, Scherer PE, Lisanti MP. Genetic ablation of caveolin-1 confers protection against atherosclerosis. Arterioscler Thromb Vasc Biol. 2004;24:98–105. https://doi.org/10.1161/01.atv.0000101182.89118.e5.
Nevins AK, Thurmond DC. Caveolin-1 functions as a novel Cdc42 guanine nucleotide dissociation inhibitor in pancreatic beta-cells. J Biol Chem. 2006;281:18961–72. https://doi.org/10.1074/jbc.M603604200.
Gustavsson J, Parpal S, Karlsson M, Ramsing C, Thorn H, Borg M, Lindroth M, Peterson KH, Magnusson KE, Strâlfors P. Localization of the insulin receptor in caveolae of adipocyte plasma membrane. FASEB J. 1999;13:1961–71.
Cohen AW, Combs TP, Scherer PE, Lisanti MP. Role of caveolin and caveolae in insulin signaling and diabetes. Am J Physiol Endocrinol Metab. 2003;285:E1151-1160. https://doi.org/10.1152/ajpendo.00324.2003.
Kim CA, Delépine M, Boutet E, El Mourabit H, Le Lay S, Meier M, Nemani M, Bridel E, Leite CC, Bertola DR, et al. Association of a homozygous nonsense caveolin-1 mutation with Berardinelli-Seip congenital lipodystrophy. J Clin Endocrinol Metab. 2008;93:1129–34. https://doi.org/10.1210/jc.2007-1328.
Pojoga LH, Underwood PC, Goodarzi MO, Williams JS, Adler GK, Jeunemaitre X, Hopkins PN, Raby BA, Lasky-Su J, Sun B, et al. Variants of the caveolin-1 gene: a translational investigation linking insulin resistance and hypertension. J Clin Endocrinol Metab. 2011;96:E1288-1292. https://doi.org/10.1210/jc.2010-2738.
Xiang AH, Azen SP, Raffel LJ, Tan S, Cheng LS, Diaz J, Toscano E, Henderson PC, Hodis HN, Hsueh WA, et al. Evidence for joint genetic control of insulin sensitivity and systolic blood pressure in hispanic families with a hypertensive proband. Circulation. 2001;103:78–83. https://doi.org/10.1161/01.cir.103.1.78.
Hunt SC, Ellison RC, Atwood LD, Pankow JS, Province MA, Leppert MF. Genome scans for blood pressure and hypertension: the National Heart, Lung, and Blood Institute Family Heart Study. Hypertension. 2002;40:1–6. https://doi.org/10.1161/01.hyp.0000022660.28915.b1.
Williams TM, Lisanti MP. The Caveolin genes: from cell biology to medicine. Ann Med. 2004;36:584–95. https://doi.org/10.1080/078538904100.
Acknowledgements
We thank the school of Nutritional and Dietetics at Tehran University of medical sciences and participants in this investigation.
Funding
This study was supported by the Tehran University of Medical Sciences (Grant number: 97–03-161–41017).
Author information
Authors and Affiliations
Contributions
Conceptualization, KM and LS; methodology, and KM; formal analysis, KM writing—original draft preparation, RAK, LS, AM, and RB; writing—review and editing, RB, AW, KS, MHS, and LS; supervision, RB and MK; project administration. All authors have read and agreed to the published version of the manuscript.”
Corresponding authors
Ethics declarations
Ethical approval and consent to participate
This research has been supported by the Tehran University of Medical Sciences (TUMS) (Ethics No. IR.TUMS.VCR.REC.1398.142). All methods were performed following the relevant guidelines and regulations. All participants provided informed consent before the investigation.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Amiri khosroshahi, R., Mirzababaei, A., Setayesh, L. et al. Dietary Insulin Index (DII) and Dietary Insulin load (DIL) and Caveolin gene variant interaction on cardiometabolic risk factors among overweight and obese women: a cross-sectional study. Eur J Med Res 29, 74 (2024). https://doi.org/10.1186/s40001-024-01638-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40001-024-01638-5