
Abstract
Cow milk is a widely consumed liquid diet, and its fatty acid composition impacts its nutritional and biochemical properties. However, research on the anticancer efficacy of cow milk-derived fat in liver cancer cells is limited. This study investigated the antiproliferative and apoptotic effects of cow milk-derived fat in human hepatoma HepG2 cells. Additionally, the effects of cow milk-derived fat on cell cycle progression and Janus kinase (JAK)-2/signal transducer and activator of transcription (STAT)-3 signaling in HepG2 cells were assessed. Furthermore, the histone deacetylase inhibitory and 2,2-diphenyl-1-picrylhydrazyl radical scavenging potential of cow milk-derived fat were examined. The results demonstrate that cow milk-derived fat can exert antiproliferative and anticlonogenic effects and induce apoptosis in HepG2 cells dose-dependently. Furthermore, cow milk-derived fat induced cell cycle arrest, suppressed the levels of pJAK-2 and pSTAT-3, and inhibited the total histone deacetylase activity in HepG2 cells. The fatty acid profile of cow milk-derived fat revealed that palmitic, oleic, and linoleic acids were abundant as saturated, monounsaturated, and polyunsaturated fatty acids, respectively. Our findings provide a new scientific basis for the development of anticancer strategies that utilize cow milk-derived fat and its derivatives against liver cancer.
Similar content being viewed by others

Introduction
According to the Food and Agriculture Organization, cow milk and its products are the most commonly consumed forms of mammalian milk, accounting for nearly 81.26% of global milk production [1]. The consumption of cow milk has several health benefits, including bone strengthening, blood pressure reduction, and reducing the risk of some chronic diseases owing to its antimicrobial and antioxidant properties [2]. It is a good source of proteins, water-soluble vitamins (riboflavin and B12), fat-soluble vitamins (A and E), fat, and minerals, such as calcium, phosphorus, sodium, magnesium, potassium, and iodine [3]. Cow milk-derived fat contains more than 400 types of fatty acids, and it is present mainly as fat globules [4]. Although season, breed, climate, health, diet, and lactation stage can affect the fatty acid composition of cow milk, palmitic acid (16:0) is the most abundant saturated fatty acid, followed by stearic acid (18:0) and myristic acid (14:0) [5, 6]. Among the unsaturated fatty acids, oleic acid (18:1) is the most abundant, followed by linoleic (18:2) and α-linoleic (18:3) acids. In addition, cow milk contains odd-chain fatty acids produced by the cow’s ruminal bacteria [4, 7].
The anticancer effects of odd-chain amino acids have gained considerable attention in fatty acid biochemistry. Heptadecanoic acid (C17:0) exerts anticancer effects on non-small cell lung carcinoma cells [8]. Moreover, pentadecanoic acid (C15:0) has been shown to have anti-cancerous effects on breast cancer stem cells [9]. Valeric acid also exerts anticancer effects on liver cancer cells [10]. Notably, when breast cancer stem-like cells were exposed to a combination of pentadecanoic acid and tamoxifen, the cells re-expressed estrogen receptor-α [11]. A recent study identified a series of odd-chain fatty acids as novel inhibitors of histone deacetylase (HDAC) 6. These play a key role in tumorigenesis through histone deacetylation, thereby contributing to tumor cell survival and proliferation. This further strengthens the biochemical significance of fatty acids [12]. Several investigations have shown the anticancer properties of specific fatty acids [13,14,15,16,17,18,19,20]. However, research on the anticancer efficacy of cow milk-derived fat, which contains a diverse range of fatty acids, including high concentrations of saturated fatty acids, is very limited in liver cancer models.
The liver plays a major role in fatty acid metabolism. Fatty acids accumulate in the liver through uptake from the bloodstream and de novo biosynthesis [21]. Dietary fat is an important source of hepatic fatty acids. Following consumption, fats are hydrolyzed in the intestinal lumen. Once absorbed into the intestine, free fatty acids are reassembled into triglycerides, which are packaged into chylomicrons and transported to the muscle and adipose tissue. In hepatocytes, the remaining triglycerides found in remnants of the chylomicrons are then transported to the liver, where they undergo intracellular processing to release free fatty acids [21]. Liver cancer is a major global health concern. Current treatment modalities include radiotherapy, chemotherapy, surgery, and immunotherapy, alone or in combination. However, these methods have limitations that can reduce their effectiveness, highlighting the need for innovations in the treatment of liver cancer [22].
The Janus kinase/signal transducer and activator of transcription (JAK/STAT) pathway is an essential signal transduction mechanism that controls gene expression and cellular functioning in the presence of growth factors and cytokines, such as epidermal growth factor (EGF) family members, interferons, and interleukins [23]. Upon ligand-induced activation, JAKs phosphorylate the cytoplasmic portion of receptor tyrosine kinases, creating binding sites for STATs. After binding, JAKs phosphorylate STATs. JAK-2 is one of four members of the JAK family, while STAT-3 is one of seven members of the STAT-3 family in humans [23, 24]. The JAK/STAT pathway is one of the most frequently dysregulated signaling pathways in cancer, specifically in hepatocellular carcinoma. STAT-3 is a transcription factor that is constantly activated in up to 60% of liver cancer cases [25]. The activation of STAT-3 (functioning as a transcription factor) results in the expression of multiple genes that significantly contribute to the diverse characteristics of cancer. This emphasizes the crucial involvement of STAT-3 in the development of liver cancer [25, 26].
Given that the liver plays a crucial role in metabolizing and accumulating fatty acids and that cow milk is a rich source of fatty acids, we conducted this study to explore the potential anticancer properties of cow milk-derived fat on liver cancer cells in vitro.
Materials and methods
Cell culture
The human liver cancer cells (HepG2) used in the present investigation were purchased from the American Type Culture Collection (Rockville, MD, USA) and cultured in Dulbecco’s modified Eagle’s medium (DMEM) (Catalog number 11,965, Life Technologies Corporation, Grand Island, NY, USA) according to the supplier’s recommendations. The medium has increased nutrient concentration, particularly glucose (4.5 g/L d-Glucose), providing energy for cell metabolism. DMEM also includes a mixture of amino acids essential for protein synthesis, and it is supplemented with a broader range of vitamins and minerals necessary for cellular processes.
Cow milk sample collection
Fresh cow milk samples were purchased from a village farmer who sells cow milk for household uses in the Colombo District, Sri Lanka. According to the farmer, the samples are from a healthy, local breed (Batu cattle) who allowed to graze on natural pastures or cultivated forages and milk for this study was obtained from local cows that were milked twice a day and in the early stage of lactation. Given that Sri Lanka does not experience significant seasonal changes, the influence of seasons was not a factor considered in this study. The samples were carefully transported to the laboratory in sealed containers and used for fat extraction upon arrival at the laboratory.
Fat extraction from cow milk
Fat was extracted from cow milk samples using the Folch method with slight modifications [27]. First, 50 mL cow milk was combined with 60 mL chloroform and methanol at a 2:1 ratio (v/v) and vigorously mixed for 10–15 s. Subsequently, 10 mL water was introduced, mixed thoroughly, and left for 48 h to separate the organic and aqueous layers. The organic layer was transferred to a clean beaker, evaporated, and used for bioassays following the preparation of the stock solutions.
Cell viability assay
HepG2 cells were seeded in 96-well plates at a density of 5000 cells/well. After 24 h, the cells were treated with cow milk-derived fat (0, 0.5, 1, 1.5, 2, or 2.5 mg/mL) dissolved in dimethyl sulfoxide. The 3-[4,5-dimethylthiazol-2-yl]-2,5 diphenyl tetrazolium bromide (MTT) assay and its calculations were conducted according to our previous study [12].
2,2-diphenyl-1-picrylhydrazyl (DPPH) radical scavenging assay
The DPPH radical scavenging assay and its data analysis were conducted as previously described with minor modifications [28]. Cow milk-derived fat with concentrations of 5, 10, and 20 mg/mL (dissolved in dimethyl sulfoxide) was used for the assay with catechin as the positive control.
Cell cycle analysis
Flow cytometry was used to analyze the effects of cow milk-derived fat on the liver cancer cell cycle. BD FACSDiva™ Software (BD Biosciences, NJ, USA) was employed to conduct the flow cytometric analysis. Before the analysis, HepG2 cells (3 × 104 cells) were washed with phosphate-buffered saline and then fixed using 70% ethanol for 30 min at 37 °C. Then, the cells were treated with RNase A (25 ng/mL) and stained with propidium iodide (40 µg/mL) before the analysis.
HDAC inhibitory assay
The HDAC colorimetric activity assay kit (ab1432; Abcam, MA, USA) evaluates the activity of HDACs through a colorimetric method. The experiment followed the manufacturer’s protocol provided with the kit. The concentrations of the final cow milk-derived fat samples ranged from 1 to 4 mg/mL. Absorbance was measured at 405 nm using a microplate reader. Trichostatin A was used as a positive control. The results were expressed as the percentage of enzyme activity for each sample tested compared to the untreated controls.
Colony formation assay
HepG2 cells were seeded at a density of 400 cells/dish in 60 cm-diameter cell culture dishes and incubated for 24 h. The cells were then treated with different concentrations of cow milk-derived fat for 21 days, after which the colony formation assay was conducted according to our previous studies [12].
Analysis of free fatty acids
The fatty acid compositions in the extracted fat fractions of cow milk samples were analyzed using the Association of Official Analytical Collaboration (AOAC) 996.06 method [29]. The relative peak area percentage (peak area relative to the total peak area %) was calculated following the established standards set by the AOAC protocols to express the fatty acid composition.
Western blot
Western blotting was performed as previously described [12]. All primary antibodies, except the anti-β-Actin primary antibody (approximately 1:10000 dilution), were diluted 1:1000 in skim milk. The primary antibodies were purchased from Cell Signaling Technology (Beverly, MA, USA). Anti-rabbit immunoglobulin G secondary antibody (Vector Laboratories, Burlingame, CA, USA) was diluted to 1:5000 before use. The bands were visualized using the BS ECL Plus Kit (Biosesang, Seongnam, South Korea). Protein bands were quantified using ImageJ software (US National Institutes of Health, Bethesda, MD, USA).
Statistical analysis
Statistical analyses entailed one-way analyses of variance using GraphPad Prism software (version 7.0; GraphPad, USA). Statistical significance was considered when p < 0.05 and indicated by an asterisk (*). All experiments were performed in triplicate, and the results are presented as the mean ± standard deviation.
Results and discussion
Free fatty acid profile of cow milk-derived fat
Analysis of the fatty acid profiles revealed varying degrees of saturated, mono-, and poly-unsaturated fatty acids in cow milk-derived fat (Fig. 1). Palmitic acid (C16:0) is cow milk’s most abundant saturated fatty acid. Among monounsaturated fatty acids, oleic acid (C18:1) is the most abundant in cow milk-derived fat. Among the polyunsaturated fatty acids, linoleic acid (C18:2), is the most abundant. Pentadecylic (or pentadecanoic) acid (C15:0) and margaric (or heptadecanoic) acid (C17:0) were detected in cow milk-derived fat samples. Our findings corroborate those of previous reports highlighting the fatty acid profiles of cow milk-derived fat [4, 6, 7].

Fatty acid profile of cow milk-derived fat analyzed using the Association of Official Analytical Collaboration 996.06 method. Peak areas relative to the total peak area are shown as percentages
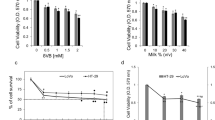
Cow milk-derived fat inhibits the proliferation and colony formation ability of liver cancer cells
Cancer cells are characterized by uncontrolled proliferation. Several studies have shown that natural and synthetic compounds can inhibit cancer cell proliferation in vitro and in vivo [9, 30, 31]. Following exposure to cow-milk fat, dose- and time-dependent inhibition of cell viability was observed with IC50 values 1.501 ± 0.053 mg/mL and 1.299 ± 0.046 mg/mL for 24 and 48 h, respectively (Fig. 2A). The colony formation assay revealed that cow milk significantly inhibited colony formation in HepG2 cells (Fig. 2B, C). Cow milk contains various components with potential anticancer effects. These components include fatty acids, ceramides, sphingosines, and specific proteins [32,33,34]. Fatty acids such as trans-10, cis-12-conjugated linoleic acid [13], propionic acid [14], butyric acid [14], linoleic acid [15], elaidic acid [16], oleic acid [17], γ-linolenic acid and dihomo-γ-linolenic acid [18], palmitic acid [19], and docosahexaenoic acid [20] have been reported to exert anticancer effects on a range of human cancer cell lines. Additionally, certain odd-chain fatty acids found in cow milk, such as C15:0 and C17:0, have been shown to inhibit cancer cell growth [8, 9], suggesting that the collective presence of these antiproliferative components in cow milk may result in synergistic effects against cancer cell proliferation. In addition to cow milk, camel colostrum exosomes exert antiproliferative effects on liver cancer cell lines HepaRG [35].

Inhibitory effects of cow milk-derived fat in HepG2 cells. A A 3-[4,5-dimethylthiazol-2-yl]-2,5 diphenyl tetrazolium bromide assay was used to assess the antiproliferative effects of cow milk-derived fat following 24 and 48 h incubations. B Effects of cow milk-derived fat on the clonogenic ability of HepG2 cells following 21 days exposure. C Statistical quantification of colonies. We tested for statistical significance using one-way analyses of variance with Dunnett’s post hoc tests. ****p < 0.0001
Cow milk-derived fat induces apoptosis in liver cancer cells
Cancer cells undergo apoptosis, characterized by specific morphological and biochemical changes. The activation of caspases plays a vital role in apoptosis. Specifically, caspase 3, an effector caspase, becomes active as apoptosis is initiated, promoting the transmission of apoptotic signals via poly ADP-ribose polymerase (PARP). Caspase 3 catalyzes the cleavage of PARP [36]. Hoechst 33,342 staining was employed to determine whether cow milk-derived fat mediates cytotoxic effects by inducing apoptosis. Chromatin condensation, a biochemical characteristic of apoptosis [37], was observed in HepG2 cells exposed to cow-milk fat (Fig. 3A). Western blotting results indicated that the cleaved form of caspase-3 was present in HepG2 cells exposed to cow milk-derived fat at the first two doses tested (Fig. 3B). Additionally, cow milk-derived fat significantly increased the levels of cleaved PARP in HepG2 cells (Fig. 3B). As shown in Fig. 3C, treatment with cow milk-derived fat caused a substantial increase in the sub-G1 population, from 6.78 ± 2.50% (0 mM) to 37.66 ± 3.92% (1.5 mM), indicating a potential cell cycle arrest in HepG2 cells exposed to cow-milk fat. Collectively, these findings suggest that cow milk-derived fat induces apoptosis and cell cycle arrest in HepG2 cells. Notably, a previous study reported that cow milk-derived fat enhances the effects of paclitaxel in vivo [38]. Furthermore, a recent study revealed that β-Casomorphin, a degradation product of β-casein, inhibits the metastasis of colorectal cancer [39]. Another recent study demonstrated that donkey milk induces apoptosis in triple-negative breast cancer cells [40], highlighting the apoptotic potential of milk. However, it is necessary to indicate that the consumption of dietary fat is associated with an increased risk of breast cancer [41].

Apoptotic potential of cow milk-derived fat and its effects on HepG2 cell cycle. A Hoechst 33,342 staining of HepG2 cells. Red arrows indicate condensed chromatin in Hoechst 33,342 stained HepG2 cells. B Western blot experiments showing the expression of apoptosis-associated proteins in HepG2 cells exposed to cow milk-derived fat for 24 h. Statistical quantification of C Cleaved-caspase 3 (C-Caspase 3)/parent caspase 3 and D Cleaved-poly ADP-ribose polymerase (C-PARP)/parent PARP. E Effects of cow milk-derived fat on the cell cycle of HepG2 cells. We tested for statistical significance using one-way analyses of variance with Dunnett’s post hoc tests (*p < 0.05 and **p < 0.01)
Total HDAC inhibition by cow milk-derived fat
HDAC inhibitors are a class of small molecules that can inhibit HDAC activity, resulting in cell cycle arrest, cell growth inhibition, and apoptosis in cancer cells [42]. We previously reported that fatty acids can inhibit HDAC6 expression [12]. In addition, we repurposed bempedoic acid, a United States Food and Drug Administration (FDA)-approved ATP citrate lyase inhibitor that mimics the structure of a dicarboxylic fatty acid, as an HDAC6 inhibitor [43]. These findings have generated interest in exploring the overall HDAC-inhibitory effects of fats extracted from cow milk. As shown in Fig. 4, cow milk-derived fat inhibited HDAC activity in a dose-dependent manner, highlighting the HDAC-inhibitory potential of cow milk-derived fat.

Total histone deacetylases (HDAC) inhibitory potential of cow-milk fat. We tested for statistical significance using a one-way analysis of variance and Dunnett’s post hoc test (**p < 0.01, and ***p < 0.001)
Cow milk-derived fat exerts antioxidant effects
The interest in antioxidants is growing rapidly, primarily because of their potential to combat the harmful effects of free radicals in the human body [44]. Several natural and synthetic compounds have been identified as free radical scavengers [44]. The results of the DPPH assay in this study indicated that cow milk-derived fat can scavenge DPPH free radicals in a dose-dependent manner with an EC50 value of 18.83 ± 1.27 (mg/mL) (Fig. 5). In addition to fatty acids, amino acids, milk-derived proteins, and minerals have been reported to contribute to the antioxidant activities of cow milk and its products [45,46,47].

The DPPH radical scavenging ability of cow-milk fat. We tested for statistical significance using a one-way analysis of variance and Dunnett’s post hoc test (****p < 0.0001)
Cow milk derived-fat suppresses the JAK-2/STAT-3 signaling in HepG2 cells
We recently reported that pentadecanoic acid can effectively suppress JAK-2/STAT-3 signaling in breast cancer stem cells [9]. This finding prompted us to investigate the potential of fatty acids as potent JAK-2/STAT-3 inhibitors. It motivated us to examine various food sources rich in fatty acids to identify natural inhibitors of JAK-2/STAT-3 signaling. We observed a significant decrease in the level of pSTAT-3/STAT-3 (Fig. 6A) and, although not statistically significant, a decrease in the level of pJAK-2/JAK-2 (Fig. 6B) in HepG2 cells when exposed to cow milk-derived fat at the tested dose of 1.5 mg/mL (Fig. 6A and B). This dose is close to the IC50 value obtained from the anti-proliferative assay after 24 h of exposure. These findings indicate that cow milk-derived fat has the potential to inhibit the proliferation of HepG2 cells, possibly by modulating the JAK-2/STAT-3 signaling pathway in liver cancer cells. It has been reported that silencing or inhibiting STAT-3 can induce apoptosis in vitro and in vivo [48,49,50]. In addition, STAT-3 has been reported to participate in cell proliferation, metastasis, and epithelial-mesenchymal transition (EMT) in liver cells, suggesting its pivotal oncogenic role in liver cancer cells [51, 52].

Western blot analysis showing the expression of signal transducers and activators of transcription Janus kinase (JAK)-2, pJAK-2 and signal tranducer and activator of transcription (STAT)-3, and pSTAT-3 in HepG2 cells exposed to cow milk-derived fat for 24 h. Statistical quantification of A pSTAT-3/STAT-3 and B pJAK-2/JAK-2. p represents phosphorylated forms. We tested for statistical significance using one-way analyses of variance with Dunnett’s post hoc tests (***p < 0.001)
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Abbreviations
- AOAC:
-
Association of Official Analytical Collaboration
- DPPH:
-
2,2-Diphenyl-1-picryhydrazyl
- EGF:
-
Epidermal growth factor
- FDA:
-
The United States Food and Drug Administration
- HDAC:
-
Histone deacetylase
- JAK:
-
Janus kinase
- MTT:
-
3-(4,5-Dimethylthiazol- 2-yl)-2,5-diphenyltetrazolium bromide
- PARP:
-
Poly ADP-ribose polymerase
- STAT:
-
Signal transducer and activator of transcription
References
FAOSTAT (2021) Food and Agriculture Organisation of the United Nations Statistical Database. Statistical Division; FAO: Rome, Italy. http://www.fao.org/statistics/en/
Claeys WL et al (2013) Raw or heated cow milk consumption: review of risks and benefits. Food Control 31(1):251–262
Gaucheron F (2005) The minerals of milk. Reprod Nutr Dev 45(4):473–483
Moate P et al (2007) Milk fatty acids. I. Variation in the concentration of individual fatty acids in bovine milk. J Dairy Sci 90(10):4730–4739
Rodríguez-Bermúdez R et al (2023) Factors affecting fatty acid composition of Holstein cow’s milk. Animals 13(4):574
Karrar E et al (2022) Fatty acid composition and stereospecificity and sterol composition of milk fat from different species. Int Dairy J 128:105313
Marangoni F et al (2019) Cow’s milk consumption and health: a health professional’s guide. J Am Coll Nutr 38(3):197–208
Xu C et al (2019) Heptadecanoic acid inhibits cell proliferation in PC-9 non-small-cell lung cancer cells with acquired gefitinib resistance. Oncol Rep 41(6):3499–3507
To NB et al (2020) Pentadecanoic acid, an odd-chain fatty acid, suppresses the stemness of MCF-7/SC human breast cancer stem-like cells through JAK2/STAT3 signaling. Nutrients 12(6):1663
Han R et al (2020) Valeric acid suppresses liver cancer development by acting as a novel HDAC inhibitor. Mol Therapy-Oncolytics 19:8–18
To NB et al (2022) Effects of combined pentadecanoic acid and tamoxifen treatment on tamoxifen resistance in MCF – 7/SC breast Cancer cells. Int J Mol Sci 23(19):11340
Ediriweera MK et al (2021) Odd-chain fatty acids as novel histone deacetylase 6 (HDAC6) inhibitors. Biochimie 186:147–156
Ediriweera MK, Tennekoon KH, Samarakoon SR (2019) Emerging role of histone deacetylase inhibitors as anti-breast-cancer agents. Drug Discovery Today 24(3):685–702
Sakai Y et al (2006) Conjugated linoleic acid reduced metastasized LL2 tumors in mouse peritoneum. Virchows Arch 449:341–347
Hague A et al (1995) Apoptosis in colorectal tumour cells: induction by the short chain fatty acids butyrate, propionate and acetate and by the bile salt deoxycholate. Int J Cancer 60(3):400–406
Lu X et al (2010) Linoleic acid suppresses colorectal cancer cell growth by inducing oxidant stress and mitochondrial dysfunction. Lipids Health Dis 9:1–11
Kishi S et al (2018) Pro–metastatic signaling of the trans fatty acid elaidic acid is associated with lipid rafts. Oncol Lett 15(4):4423–4426
DAILEY OD et al (2011) Anticancer activity of branched-chain derivatives of oleic acid. Anticancer Res 31(10):3165–3169
Xu Y, Qian SY (2014) Anti-cancer activities of ω-6 polyunsaturated fatty acids. Biomedical J 37(3):112
Zhu S et al (2021) Palmitic acid inhibits prostate cancer cell proliferation and metastasis by suppressing the PI3K/Akt pathway. Life Sci 286:120046
Pizato N et al (2018) Omega-3 docosahexaenoic acid induces pyroptosis cell death in triple-negative breast cancer cells. Sci Rep 8(1):1952
Alves-Bezerra M, Cohen DE (2017) Triglyceride metabolism in the liver. Compr Physiol 8(1):1
Foda ZH et al (2023) Detecting liver cancer using cell-free DNA fragmentomes. Cancer Discov 13(3):616–631
Stark GR, Darnell JE (2012) The JAK-STAT pathway at twenty. Immunity 36(4):503–514
Rah B et al (2023) JAK/STAT signaling and cellular iron metabolism in hepatocellular carcinoma: therapeutic implications. Clin Exp Med 23:1–11
Song X, Ma J (2022) SRRM1 promotes the proliferation, migration, and invasion of hepatocellular carcinoma cells by regulating the JAK/STAT signaling pathway. Tissue Cell 79:101954
Hin Tang JJ et al (2020) JAK/STAT signaling in hepatocellular carcinoma. Hepatic Oncol 7(1):HEP18
Folch J, Lees M, Sloane Stanley GH (1957) A simple method for the isolation and purification of total lipids from animal tissues. J biol Chem 226(1):497–509
Kim DK et al (2022) Antioxidant activity of banana flesh and antiproliferative effect on breast and pancreatic cancer cells. Food Sci Nutr 10(3):740–750
Cunniff P, Washington D (1997) Official methods of analysis of AOAC international. J AOAC Int 80(6):127A
Ediriweera MK et al (2020) 10-Gingerol targets lipid rafts associated PI3K/Akt signaling in radio-resistant triple negative breast cancer cells. Molecules 25(14):3164
Ediriweera MK, Tennekoon KH, Samarakoon SR (2019) In vitro assays and techniques utilized in anticancer drug discovery. J Appl Toxicol 39(1):38–71
Gill HS, Cross M (2000) Anticancer properties of bovine milk. Br J Nutr 84(S1):161–166
Leischner C et al (2021) Potential protective protein components of cow’s milk against certain tumor entities. Nutrients 13(6):1974
Parodi PW (1997) Cows’ milk fat components as potential anticarcinogenic agents. J Nutr 127(6):1055–1060
Shaban AM et al (2023) Antibacterial, antifungal, and anticancer effects of camel milk exosomes: an in vitro study. Veterinary Sci 10(2):124
Boulares AH et al (1999) Role of poly (ADP-ribose) polymerase (PARP) cleavage in apoptosis: caspase 3-resistant PARP mutant increases rates of apoptosis in transfected cells. J Biol Chem 274(33):22932–22940
Tixeira R et al (2017) Defining the morphologic features and products of cell disassembly during apoptosis. Apoptosis 22:475–477
Sun X et al (2011) Dairy milk fat augments paclitaxel therapy to suppress tumour metastasis in mice, and protects against the side-effects of chemotherapy. Clin Exp Metastasis 28:675–688
Mori S et al (2021) Enhancement of anti-tumoral immunity by β-casomorphin-7 inhibits cancer development and metastasis of colorectal cancer. Int J Mol Sci 22(15):8232
Li Q et al (2020) Donkey milk inhibits triple-negative breast tumor progression and is associated with increased cleaved-caspase-3 expression. Food Funct 11(4):3053–3065
Kroenke CH et al (2013) High-and low-fat dairy intake, recurrence, and mortality after breast cancer diagnosis. J Natl Cancer Inst 105(9):616–623
Ediriweera MK, Rajan P, Cho SK (2023) Repurposing bempedoic acid as a histone deacetylase 6 inhibitor. Int J Epigenetics 3(1):1–5
Fang Y-Z, Yang S, Wu G (2002) Free radicals, antioxidants, and nutrition. Nutrition 18(10):872–879
Khan IT et al (2019) Antioxidant properties of Milk and dairy products: a comprehensive review of the current knowledge. Lipids Health Dis 18:1–13
Śpitalniak-Bajerska K et al (2020) Effects of n-3 fatty acids on growth, antioxidant status, and immunity of preweaned dairy calves. J Dairy Sci 103(3):2864–2876
Lozano B et al (2014) Vitamins, fatty acids, and antioxidant capacity stability during storage of freeze-dried human milk. Int J Food Sci Nutr 65(6):703–707
Rajendran P et al (2012) Celastrol suppresses growth and induces apoptosis of human hepatocellular carcinoma through the modulation of STAT3/JAK2 signaling cascade in vitro and in vivo. Cancer Prev Res 5(4):631–643
Liu Y et al (2010) Inhibition of STAT3 signaling blocks the anti-apoptotic activity of IL-6 in human liver cancer cells. J Biol Chem 285(35):27429–27439
Zhang N et al (2015) STAT3 regulates the migration and invasion of a stem–like subpopulation through microRNA-21 and multiple targets in hepatocellular carcinoma. Oncol Rep 33(3):1493–1498
Wang B et al (2018) STAT3 aggravates TGF-β1-induced hepatic epithelial-to-mesenchymal transition and migration. Biomed Pharmacother 98:214–221
Wendt MK et al (2014) STAT3 and epithelial–mesenchymal transitions in carcinomas. Jak-stat 3(2):e28975
Acknowledgements
We would like to thank Editage (www.editage.co.kr) for English language editing.
Funding
The present study was supported by grants from the UNESCO-TWAS and the Swedish International Development Cooperation Agency (Grant number 22–140 RG/BIO/AS_I) and by Basic Science Research Program through the National Research Foundation of Korea funded by the Ministry of Education (2016R1A6A1A03012862).
Author information
Authors and Affiliations
Contributions
MKE, DMC and SKC designed the study. MKE and DMC performed the experiments, analyzed the data and wrote the manuscript. MKE, DMC and SKC confirm the authenticity of all the raw data.
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ediriweera, M.K., Cuong, D.M. & Cho, S.K. Cow milk derived-fat inhibits the proliferation of liver cancer-cells. Appl Biol Chem 67, 7 (2024). https://doi.org/10.1186/s13765-024-00860-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13765-024-00860-3