
Abstract
We propose and study a nonautonomous harvesting Lotka–Volterra commensalism model incorporating partial closure for the populations. By using the differential inequality theory we obtain sufficient conditions that ensure the extinction, partial survival, and permanence of the system. By applying the fluctuation lemma we establish sufficient conditions that ensure the extinction of one of the components and the stability of the the other one. For the permanent case, by constructing a suitable Lyapunov function we obtain some sufficient conditions for the globally attractivity of the positive solution of the system. Examples, together with their numeric simulations, show the feasibility of the main results. To ensure the stable coexistence of the two species, the harvesting area should be carefully restricted.
Similar content being viewed by others

1 Introduction
During the last decade, many scholars [1–11] investigated the dynamic behavior of the mutualism model, and many excellent results were obtained. For example, Chen, Xie, and Chen [1] showed that the stage structure of the species can lead to the extinction of the mutualism model, despite the cooperation between the species; Chen, Chen, and Li [3] showed that the feedback control variables have no influence on the persistent property of a kind of mutualism model, and in this direction, some similar results was established in [6, 8]; several scholars [2, 4, 7, 10, 11] investigated the stability property of the positive equilibrium of the cooperative system, Xie, Chen, and Xue [10] showed that if the harvesting effort is limited, then the cooperative system admits a unique positive equilibrium, which is globally attractive.
Commensalism, which describes a symbiotic interaction between two populations where one population gets benefit from the other while the other is neither harmed nor benefited due to the interaction with the previous species [12], has not arisen the attention of the scholars, since the model seems simple and can be seen as a particular case of the mutualism model. Only recently scholars paid attention to such a kind of relationship; see [12–20] and the references therein. Topics such as the existence of the positive periodic solution [17], the existence of a positive almost periodic solution [14], the existence and stability of the positive equilibrium [16], the influence of the impulsive [15] were investigated, and many excellent results were obtained. However, as was pointed out by Georgescu and Maxin [20], “One would think that the stability of the coexisting equilibria for two-species models of commensalism would follow immediately from the corresponding results for models of mutualism, when these results are available, …, However, this is not actually the case”. Hence, it is necessary to do some further works on commensalism model.
Sun and Sun [18] proposed the following commensalism system:
where \(r_{1}\), \(r_{2}\), \(K_{1}\), \(K_{2}\), α are all positive constants. By linearizing the system at equilibrium the authors investigated the local stability property of the equilibria of the system. They showed that the unique positive equilibrium of the system is locally asymptotically stable, whereas the other three boundary equilibria are all unstable.
Recently, Xue, Han, Yang et al. [21] argued that the nonautonomous model is more suitable, since the coefficients of the system vary with time. They proposed the following two species nonautonomous commensalism model:
The authors gave a set of sufficient conditions that ensure the existence of a unique globally attractive positive periodic solution.
On the other hand, to obtain the resource for the development of the human being, harvest of the species is necessary. During the last decades, many scholars investigated the influence of the harvesting to predator–prey or competition system; see [22–27] and the references therein. Chakraborty, Das, and Kar [24] argued that it is necessary to harvest the population but harvesting should be regulated, so that both the ecological sustainability and conservation of the species can be implemented in a long run. They proposed the following harvesting predator–prey model:
Some interesting results concerned with the boundedness of the system, existence of equilibria, and local and global stability of the positive equilibrium were obtained.
Stimulated by the works of Xue, Han, Yang et al. [21] and Chakraborty, Das, and Kar [24], in this paper, we propose the following nonautonomous nonselective harvesting Lotka–Volterra commensalism model incorporating partial closure for the populations:
where \(a(t)\), \(b(t)\), \(c(t)\), \(d(t)\), \(e(t)\), \(q_{1}(t)\), \(q_{2}(t)\), \(F(t)\), and \(m(t)\) are positive constants, and \(a(t)\), \(b(t)\), \(c(t)\), \(d(t)\), \(e(t)\) have the same meaning as in system (1.2); \(F(t)\) is the combined fishing effort used to harvest, and \(m(t)\) (\(0< m(t)<1\)) is the fraction of the stock available for harvesting.
As for as an ecosystem is concerned, there are the most important three topics: permanence, extinction, and global attractivity, which reflect the existence of the species in the long run, the extinction of the species, and the species maintained in a stable state. During the last decades, there are many excellent results on these three topics; see [28–37] and the references therein. For example, Shi, Li, and Chen [30] studied the extinction property of a competition system with infinite delay and feedback controls; Chen, Xie, and Li [31] investigated the partial extinction of the predator–prey model with stage structure; Chen, Chen, and Huang [32] investigated the extinction property of the nonlinear competition system with Beddington–DeAngelis functional response; Xie, Xue, Wu et al. [33] studied the extinction property of a nonlinear toxic substance competition system; Chen, Ma, and Zhang [34] showed that if the refuge is restricted to suitable area, then the Lotka–Volterra predato–prey system can admit a unique positive equilibrium, which is globally attractive. In this paper, we also focus our attention on the persistency, extinction, and stability of system (1.4).
The paper is arranged as follows. We will investigate the extinction, partial survival and persistency of system (1.4) in the next section. In Sect. 3, we investigate the global stability property of the solutions of the system. Two examples, together with their numeric simulations, are presented in Sect. 4 to show the feasibility of the main results. We end this paper by a brief discussion.
2 Extinction and persistency of the system
For the rest of the paper, for a bounded continuous function g defined on R, let
Lemma 2.1
([28])
If \(a>0\), \(b>0\), and \(\dot{x}\geq x(b-ax)\) for \(t\geq 0\) and \(x(0)>0\), then
If \(a>0\), \(b>0\), and \(\dot{x} \leq x(b-ax)\) for \(t\geq 0\) and \(x(0)>0\), then
Lemma 2.2
The domain \(R^{2}_{+}=\{(x,y)|x>0,y>0\}\) is invariant with respect to (1.4).
Proof
Since
where
the assertion of the lemma immediately follows for all \(t\in [0,+ \infty )\). □
Theorem 2.1
Let \((N_{1}(t),N_{2}(t))^{T}\) be any solution of system (1.4). Assume that
Then
that is, both species will be driven to extinction.
Proof
It follows from (2.1) that there exists a small enough \(\varepsilon >0\) such that
Let \((N_{1}(t),N_{2}(t))^{T}\) be any solution of system (1.4). From the second equation of system (1.4) it follows that
So
For \(\varepsilon >0\) as in (2.2), it follows from (2.4) that there exists a large enough \(T_{1}\) such that
From the first equation and (2.5) it follows that
So
This ends the proof of Theorem 2.1. □
Theorem 2.2
Let \((N_{1}(t),N_{2}(t))^{T}\) be any solution of system (1.4). Assume that
Then
that is, the first species is permanent, and the second species will be driven to extinction.
Proof
From the first inequality of (2.8), similarly to the analysis of (2.3)–(2.4), for any solution \((N_{1}(t),N_{2}(t))^{T}\) of system (1.4), we obtain
For \(\varepsilon >0\) small enough, it follows from (2.9) that there exists a large enough \(T_{2}\) such that
From the first equation and (2.10) it follows that
It follows from Lemma 2.1 and (2.11) that
Since \(\varepsilon >0\) is an arbitrary small positive constant, letting \(\varepsilon \rightarrow 0\) in (2.12) leads to
From the first equation we also have
It follows from Lemma 2.1 and (2.14) that
Relations (2.9), (2.13), and (2.15) show that the statement of Theorem 2.2 holds. This ends the proof of the theorem. □
Theorem 2.3
Let \((N_{1}(t),N_{2}(t))^{T}\) be any solution of system (1.4). Assume that
Then
that is, the second species is permanent, whereas the first species will be driven to extinction.
Proof
It follows from (2.16) that there exists a small enough \(\varepsilon >0\) such that
Let \((N_{1}(t),N_{2}(t))^{T}\) be any positive solution of system (1.4). From the second equation of system (1.4) it follows that
It follows from Lemma 2.1 and (2.18) that
For \(\varepsilon >0\) as in (2.17), it follows from (2.19) that there exists \(T_{3}>0\) such that
Again, from the second equation of system (1.4) we also have
It follows from Lemma 2.1 and (2.21) that
From the first equation and (2.20), for \(t\geq T_{3}\), it follows that
So
where
It immediately follows from (2.19), (2.22), and (2.24) that the statement of Theorem 2.3 holds. This ends the proof of the theorem. □
Theorem 2.4
Let \((N_{1}(t),N_{2}(t))^{T}\) be any solution of system (1.4). Assume that
Then the system is permanent, that is, there exist positive constants \(m_{i}\), \(M_{i}\), \(i=1,2\), independent of the solutions of (1.4), such that
where
Proof
It follows from (2.25) that, indeed, for enough small \(\varepsilon >0\), namely, for
we have the inequality
From (2.28) we can easily see that
Let \((N_{1}(t),N_{2}(t))^{T}\) be any solution of system (1.4). From the second equation of system (1.4), applying the first inequality of (2.25), similarly to the analysis of (2.18)–(2.22), we can show that
For any positive constant \(\varepsilon >0\) small enough, which satisfies (2.27) and \(\varepsilon < \frac{d^{L}-q_{2}^{M}F^{M}m^{M}}{e^{M}}\), there exists a large enough \(T_{4}>0\) such that
From the first equation of (1.4) and (2.31), for \(t\geq T_{4}\), it follows that
Applying Lemma 2.1 to (2.32) leads to
Setting \(\varepsilon \rightarrow 0\) in this inequality leads to
From the first equation of (1.4) and (2.31), for \(t\geq T_{4}\), we also have
Applying Lemma 2.1 to (2.34) leads to
Setting \(\varepsilon \rightarrow 0\) in this inequality leads to
Relations (2.30), (2.33), and (2.35) show that the statement of Theorem 2.4 holds. This ends the proof of the theorem. □
3 Global attractivity
In Sect. 2, we discussed the persistent or extinction property of the system, which means that the solutions of the system are bounded above and below by some positive constants or the species will be driven to extinction. One of the interesting problems is to give sufficient conditions to ensure the global attractivity of the positive solution of the system. Before we state the main results of this section, we need to introduce two useful lemmas.
Lemma 3.1
(Fluctuation lemma, [35, Lemma 4])
Let \(x(t)\) be a bounded differentiable function on \((\alpha , \infty )\). Then there exist sequences \(\tau_{n}\rightarrow \infty \) and \(\sigma_{n}\rightarrow \infty \) such that
For the logistic equation
from Lemma 2.1 of Zhao and Chen [36] we have the following:
Lemma 3.2
Suppose that \(r(t)\) and \(a(t)\) are bounded above and below by positive constants. Then any positive solutions of Eq. (3.1) are defined on \([0, +\infty )\), bounded above and below by positive constants, and globally attractive.
Theorem 3.1
Under the assumptions of Theorem 2.2, let \(N(t)=(N_{1}(t),N_{2}(t))^{T}\) be any positive solution of system (1.4). Then the species \(N_{2}\) will be driven to extinction, that is, \(N_{2}(t)\rightarrow 0 \) as \(t\rightarrow +\infty \), and \(N_{1}(t)\rightarrow N_{1}^{*}(t)\) as \(t\rightarrow +\infty \), where \(N_{1}^{*}(t)\) is any positive solution of
Proof
Let \(N(t)=(N_{1}(t),N_{2}(t))^{T}\) be any positive solution of system (1.4). By Theorem 2.2 the species \(N_{2}\) will be driven to extinction, that is, \(N_{2}(t)\rightarrow 0 \) as \(t\rightarrow +\infty \). To finish the proof of Theorem 3.1, it suffices to show that \(N_{1}(t)\rightarrow N_{1}^{*}(t)\) as \(t\rightarrow +\infty \), where \(N_{1}^{*}(t)\) is any positive solution of system (3.2). It follows from Theorem 2.2 and Lemma 3.2 that there exists \(T_{5}>0\) such that
and
where \(\eta_{i}\), \(i=1, 2\), are two positive constants independent of the solution of system (3.2). Let \(w(t)=(N_{1}(t))^{-1}\), \(w^{*}(t)=(N _{1}^{*}(t))^{-1}\), and \(z(t)=w(t)-w^{*}(t)\). Then
It follows that z satisfies
From this analysis, for \(t\geq T_{5}\), we have
and
Thus \(z(t)\) is a bounded differentiable function. By the fluctuation lemma (Lemma 3.1) there exist sequences \(\tau_{n}\rightarrow \infty \) and \(\sigma_{n}\rightarrow \infty \) such that \(z(\tau_{n})\rightarrow \overline{z}\), \(z^{\prime}(\tau_{n})\rightarrow 0\); \(z(\sigma_{n})\rightarrow \underline{z}\), \(z^{\prime}(\sigma_{n})\rightarrow 0\) as \(n\rightarrow \infty \). We will show that \(\overline{z}=\underline{z}=0\). From (3.3) we have
Noting that
and \(\lim_{t\rightarrow \infty } N_{2}(t)=0\), we can see that
Hence \(\overline{z}=\underline{z}=0\). Since
and both \(N_{1}(t)\) and \(N_{1}^{*}(t)\) are bounded functions, we have
as required. This completes the proof. □
Theorem 3.2
Under the assumptions of Theorem 2.3, let \(N(t)=(N_{1}(t),N_{2}(t))^{T}\) be any positive solution of system (1.4). Then the species \(N_{1}\) will be driven to extinction, that is, \(N_{1}(t)\rightarrow 0 \) as \(t\rightarrow +\infty \), and \(N_{2}(t)\rightarrow N_{2}^{*}(t)\) as \(t\rightarrow +\infty \), where \(N_{2}^{*}(t)\) is any positive solution of
Proof
Let \(N(t)=(N_{1}(t),N_{2}(t))^{T}\) be any positive solution of system (1.4). By Theorem 2.3 the species \(N_{1}\) will be driven to extinction, that is, \(N_{1}(t)\rightarrow 0 \) as \(t\rightarrow +\infty \). On the other hand, noting that the second equation of system (1.4) is independent of \(N_{1}(t)\), from Lemma 3.2 it immediately follows that \(N_{2}(t)\rightarrow N_{2}^{*}(t)\) as \(t\rightarrow + \infty \), where \(N_{2}^{*}(t)\) is any positive solution of system (3.4). This ends the proof of Theorem 3.2. □
Theorem 3.3
In addition to (2.25), assume further that
Let \(N(t)=(N_{1}(t),N_{2}(t))^{T}\) and \(N^{*}(t)=(N_{1}^{*}(t), N_{2} ^{*}(t))^{T}\) be any two positive solutions of system (1.4). Then
Proof
Let \(N(t)=(N_{1}(t),N_{2}(t))^{T}\) and \(N^{*}(t)=(N _{1}^{*}(t), N_{2}^{*}(t))^{T}\) be any two positive solutions of system (1.4). For any small enough positive constant \(\varepsilon >0\), it then follows from Theorem 2.4 that there exists a large enough \(T_{6}\) such that, for all \(t\geq T_{6}\),
Now let
Then, by (3.5), for \(t>T_{6}\), we have
Integrating both sides of (3.7) on the interval \([T_{6},t)\), we have
It follows from (3.8) that
Therefore \(V(t)\) is bounded on \([T_{6},+\infty )\), and also
By (3.6), \(\vert N_{1}(t)-N_{1}^{*}(t) \vert \) and \(\vert N_{2}(t)-N_{2}^{*}(t) \vert \) are bounded on \([T_{6},+\infty )\). On the other hand, it is easy to see that \(\dot{N_{1}}(t)\), \(\dot{N_{2}}(t)\), \(\dot{N}_{1}^{*}(t)\), and \(\dot{N}_{2}^{*}(t)\) are bounded for \(t\geq T_{6}\). Therefore \(\vert N_{1}(t)-N_{1}^{*}(t) \vert \), \(\vert N_{2}(t)-N_{2}^{*}(t) \vert \) are uniformly continuous on \([T_{6},+\infty )\). By the Barbălat lemma we conclude that
This ends the proof of Theorem 3.3. □
4 Numerical simulations
Example 4.1
Consider the following system:
Corresponding to system (1.4), here we take
We can easily verify that
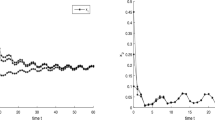
Hence, all the conditions of Theorem 2.1 hold, and it follows from Theorem 2.1 that both species will be driven to extinction. Numeric simulations (Figs. 1 and 2) also support this assertion.

Numeric simulations of the first component system (4.1) with initial conditions \((x(0), y(0))=(0.5,0.1), (0.8,0.5), (0.3, 0.7)\), and \((0.7,0.9)\)

Numeric simulations of the second component of system (4.1) with initial conditions \((x(0), y(0))=(0.5,0.1), (0.8,0.5), (0.3, 0.7)\), and \((0.7,0.9)\)
Example 4.2
Consider the following system:
Corresponding to system (1.4), here we take
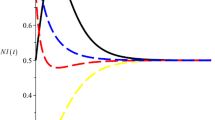
We can easily verify that all the conditions of Theorems 2.4 and 3.3 hold, and it follows from Theorems 2.4 and 3.3 that the system is permanent and that any positive solution of system (4.4) is globally attractive. Numeric simulations (Figs. 3 and 4) also support this assertion.

Numeric simulations of the first component of system (4.4) with initial conditions \((x(0), y(0))=(0.5,0.1), (0.8,0.5), (0.3, 0.7)\), and \((0.7,0.9)\)

Numeric simulations of the second component of system (4.4) with initial conditions \((x(0), y(0))=(0.5,0.1), (0.8,0.5), (0.3, 0.7)\), and \((0.7,0.9)\)
5 Discussion
Recently, many scholars [12–21] studied the dynamic behavior of the commensalism model; however, none of them consider the influence of harvesting. Stimulated by the recent works of Chakraborty, Das, and Kar [24], we propose a nonautonomous nonselective commensalism model incorporating partial closure to the population.
We focus our attention on the persistent and extinction property of the system, Theorems 2.1–2.4 show that, depending on the area that can be harvested, the the system may exhibit permanent, extinction, or partial survival phenomenon, that is, the introducing of harvesting makes the dynamic behavior of the system complicated. Theorem 2.4 shows that if the harvesting area is small enough (i.e., m is small enough), then two species can coexist in the long run. If we further assume that the intrinsic competition rate (\(e(t)\)) is larger than the cooperative between the two species (\(c(t)\)), then two species can coexist in a stable state. Such a result may help us in designing the reserve area of the species.
It seems interesting to incorporate the time delay to system (1.4) and study the influence of the time delay. We leave this for future study.
References
Chen, F.D., Xie, X.D., Chen, X.F.: Dynamic behaviors of a stage-structured cooperation model. Commun. Math. Biol. Neurosci. 2015, Article ID 4 (2015)
Chen, F.D., Xie, X.D.: Study on the Dynamic Behaviors of Cooperation Population Modeling. Science Press, Beijing (2014)
Chen, L.J., Chen, L.J., Li, Z.: Permanence of a delayed discrete mutualism model with feedback controls. Math. Comput. Model. 50, 1083–1089 (2009)
Yang, K., Miao, Z.S., Chen, F.D., Xie, X.D.: Influence of single feedback control variable on an autonomous Holling-II type cooperative system. J. Math. Anal. Appl. 435(1), 874–888 (2016)
Chen, L.J., Xie, X.D.: Permanence of an N-species cooperation system with continuous time delays and feedback controls. Nonlinear Anal., Real World Appl. 12, 34–38 (2001)
Chen, L.J., Xie, X.D., Chen, L.J.: Feedback control variables have no influence on the permanence of a discrete N-species cooperation system. Discrete Dyn. Nat. Soc. 2009, Article ID 306425 (2009)
Yang, K., Xie, X., Chen, F.: Global stability of a discrete mutualism model. Abstr. Appl. Anal. 2014, Article ID 709124 (2014)
Li, Y.K., Zhang, T.W.: Permanence of a discrete n-species cooperation system with time-varying delays and feedback controls. Math. Comput. Model. 53, 1320–1330 (2011)
Liu, Z.J., Wu, J.H., Tan, R.H., Chen, Y.P.: Modeling and analysis of a periodic delayed two-species model of facultative mutualism. Appl. Math. Comput. 217, 893–903 (2010)
Xie, X.D., Chen, F.D., Xue, Y.L.: Note on the stability property of a cooperative system incorporating harvesting. Discrete Dyn. Nat. Soc. 2014, Article ID 327823 (2014)
Xie, X.D., Chen, F.D., Yang, K., Xue, Y.L.: Global attractivity of an integrodifferential model of mutualism. Abstr. Appl. Anal. 2014, Article ID 928726 (2014)
Hari Prasad, B., Pattabhi Ramacharyulu, N.Ch.: Discrete model of commensalism between two species, I. J. Mod. Educ. Comput. Sci. 8, 40–46 (2012)
Wu, R.X., Li, L., Lin, Q.F.: A Holling type commensal symbiosis model involving Allee effect. Commun. Math. Biol. Neurosci. 2018, Article ID 6 (2018)
Xue, Y.L., Xie, X.D., Chen, F.D., et al.: Almost periodic solution of a discrete commensalism system. Discrete Dyn. Nat. Soc. 2015, Article ID 295483 (2015)
Miao, Z.S., Xie, X.D., Pu, L.Q.: Dynamic behaviors of a periodic Lotka–Volterra commensal symbiosis model with impulsive. Commun. Math. Biol. Neurosci. 2015, Article ID 3 (2015)
Wu, R.X., Lin, L., Zhou, X.Y.: A commensal symbiosis model with Holling type functional response. J. Math. Comput. Sci. 16, 364–371 (2016)
Lin, Q.F.: Dynamic behaviors of a commensal symbiosis model with non-monotonic functional response and non-selective harvesting in a partial closure. Commun. Math. Biol. Neurosci. 2018, Article ID 4 (2018)
Sun, G.C., Sun, H.: Analysis on symbiosis model of two populations. J. Weinan Normal Univ. 28(9), 6–8 (2013)
Zhu, Z.F., Chen, Q.L.: Mathematic analysis on commensalism Lotka–Volterra model of populations. J. Jixi Univ. 8(5), 100–101 (2008)
Georgescu, P., Maxin, D.: Global stability results for models of commensalism. Int. J. Biomath. 10(3), 1750037 (2017)
Xue, Y.L., Han, R.Y., Yang, L.Y., Chen, F.D.: On the existence and stability of positive periodic solution of a nonautonomous commensal symbiosis model of two populations. J. Shangming Univ. 32(2), 32–37 (2015)
Chen, B.G.: Dynamic behaviors of a non-selective harvesting Lotka–Volterra amensalism model incorporating partial closure for the populations. Adv. Differ. Equ. 2018, 111 (2018)
Chen, L., Chen, F.: Global analysis of a harvested predator–prey model incorporating a constant prey refuge. Int. J. Biomath. 3(02), 177–189 (2010)
Chakraborty, K., Das, S., Kar, T.K.: On non-selective harvesting of a multispecies fishery incorporating partial closure for the populations. Appl. Math. Comput. 221, 581–597 (2013)
Kar, T.K., Chaudhuri, K.S.: On non-selective harvesting of two competing fish species in the presence of toxicity. Ecol. Model. 161, 125–137 (2003)
Leard, B., Rebaza, J.: Analysis of predator–prey models with continuous threshold harvesting. Appl. Math. Comput. 217(12), 5265–5278 (2011)
Chakraborty, K., Jana, S., Kar, T.K.: Global dynamics and bifurcation in a stage structured prey–predator fishery model with harvesting. Appl. Math. Comput. 218(18), 9271–9290 (2012)
Chen, F., Li, Z., Huang, Y.J.: Note on the permanence of a competitive system with infinite delay and feedback controls. Nonlinear Anal., Real World Appl. 8, 680–687 (2007)
Chen, F.D., Xie, X.D., Miao, Z.S., et al.: Extinction in two species nonautonomous nonlinear competitive system. Appl. Math. Comput. 274, 119–124 (2016)
Shi, C., Li, Z., Chen, F.: Extinction in a nonautonomous Lotka–Volterra competitive system with infinite delay and feedback controls. Nonlinear Anal., Real World Appl. 13(5), 2214–2226 (2012)
Chen, F., Xie, X., Li, Z.: Partial survival and extinction of a delayed predator–prey model with stage structure. Appl. Math. Comput. 219(8), 4157–4162 (2012)
Chen, F., Chen, X., Huang, S.: Extinction of a two species non-autonomous competitive system with Beddington–DeAngelis functional response and the effect of toxic substances. Open Math. 14(1), 1157–1173 (2016)
Xie, X., Xue, Y., Wu, R., et al.: Extinction of a two species competitive system with nonlinear inter-inhibition terms and one toxin producing phytoplankton. Adv. Differ. Equ. 2016, Article ID 258 (2016)
Chen, F., Ma, Z., Zhang, H.: Global asymptotical stability of the positive equilibrium of the Lotka–Volterra prey–predator model incorporating a constant number of prey refuges. Nonlinear Anal., Real World Appl. 13(6), 2790–2793 (2012)
Montes De Oca, F., Vivas, M.: Extinction in two dimensional Lotka–Volterra system with infinite delay. Nonlinear Anal., Real World Appl. 7(5), 1042–1047 (2006)
Zhao, J.D., Chen, W.C.: The qualitative analysis of N-species nonlinear prey-competition systems. Appl. Math. Comput. 149, 567–576 (2004)
Lin, Q.X., Xie, X.D., et al.: Dynamical analysis of a logistic model with impulsive Holling type-II harvesting. Adv. Differ. Equ. 2018, 112 (2018)
Acknowledgements
The authors are grateful to anonymous referees for their excellent suggestions, which greatly improved the presentation of the paper.
Funding
This work is supported by the National Natural Science Foundation of China under Grant (11601085) and the Natural Science Foundation of Fujian Province (2017J01400).
Author information
Authors and Affiliations
Contributions
All authors contributed equally to the writing of this paper. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare that there is no conflict of interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Liu, Y., Xie, X. & Lin, Q. Permanence, partial survival, extinction, and global attractivity of a nonautonomous harvesting Lotka–Volterra commensalism model incorporating partial closure for the populations. Adv Differ Equ 2018, 211 (2018). https://doi.org/10.1186/s13662-018-1662-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13662-018-1662-3