
Abstract
Brain–heart interaction has raised up increasing attentions. Nuclear receptors (NRs) are abundantly expressed in the brain, and emerging evidence indicates that a number of these brain NRs regulate multiple aspects of cardiovascular diseases (CVDs), including hypertension, heart failure, atherosclerosis, etc. In this review, we will elaborate recent findings that have established the physiological relevance of brain NRs in the context of cardiovascular function. In addition, we will discuss the currently available evidence regarding the distinct neuronal populations that respond to brain NRs in the cardiovascular control. These findings suggest connections between cardiac control and brain dynamics through NR signaling, which may lead to novel tools for the treatment of pathological changes in the CVDs.
Similar content being viewed by others
Introduction
Nuclear receptors (NRs) contain a large superfamily of at least 48 DNA-binding transcription factors whose target genes are critical for many biological processes, including growth, differentiation, development, and metabolism [1, 2]. The NR superfamily, sharing common structure, can be divided into three subfamilies: (1) steroid hormone receptors, such as estrogen receptor (ER), androgen receptor (AR), glucocorticoid receptor (GR), mineralocorticoid receptor (MR), and progesterone receptor (PR); (2) non-steroid hormone receptors, such as thyroid hormone receptor (THR), retinoic acid receptor (RAR), and vitamin D receptor (VDR); (3) orphan receptors, such as testicular nuclear receptor (TR), retinoid X receptor (RXR), peroxisome proliferator-activated receptor (PPAR), liver X receptor (LXR), steroidogenic factor 1 (SF-1) [3].
NRs sense changing levels of nutrients and hormones and it has been well established that NRs play essential roles in the regulation of various diseases including dyslipidemia, diabetes, cancer, neurodegenerative disorders, infertility, and obesity [4]. More recently, the interaction of NRs and cardiovascular function has received increasing attentions [5, 6]. In this review, we will focus on the relationship between brain NRs and cardiovascular diseases (CVDs). NRs are found abundantly expressed in the brain [7,8,9,10]. Emerging evidence indicates that a number of these brain NRs regulate multiple aspects of CVDs, including hypertension, atherosclerosis, and heart failure [11,12,13,14]. Here, we classified the NRs as three groups: (1) NRs (ER, MR, and THR) that have direct evidence supports the brain NRs regulate cardiovascular functions; (2) NRs (AR, GR, PPAR, PR, RAR, RXR, VDR, etc.) that play critical roles in the regulation of cardiovascular functions but lack direct evidence to establish the relationship between brain NRs and cardiovascular functions; (3) NRs (LXR, SF-1, TR, etc.) that currently have no known roles in the regulation of cardiovascular or other functions.
An imbalanced brain–heart interaction has a negative impact on cardiovascular health [15, 16]. In this review, we summarize all the current and available evidence regarding the role of brain NRs in the regulation of cardiovascular function.
Brain NRs that regulate cardiovascular functions
Estrogen receptor (ER)
Estrogen exerts a vast range of biological effects in cardiovascular function, including vascular function, the inflammatory response, metabolism, insulin sensitivity, cardiac myocyte and stem cell survival, and the development of hypertrophy [17]. Estrogen, with its potential as a cardioprotective agent and as an immunomodulator of the inflammatory response in atherosclerosis, has been widely discussed during the past three decades [18,19,20]. CVDs are the leading cause of death among women [21]. Women of reproductive age have lower blood pressure than men and can be protected from atherosclerosis [22]; while the loss of endogenous estrogen production after menopause leads to a higher prevalence of hypertension and incidence of stroke than men [23]. While in postmenopausal women [24] and ovariectomized (OVX) rats [25], estrogen replacement decreases blood pressure. The studies [26, 27], investigating the sex differences in CVDs, emphasize the importance and urgent need to further understand of estrogen and estrogen receptors (ER) in cardiovascular function.
Estrogen signaling is mediated through two ERs, ERα and ERβ, both belonging to the NR family of transcription factors. Recent studies have demonstrated that activated ER may promote physiological functions via non-transcriptional mechanisms [12, 28]. These non-genomic effects involve steroid-induced modulation of cytoplasmic or cell membrane-bound regulatory proteins. Several biological actions of estrogen are associated with these non-genomic signaling, and intracellular regulatory cascades. For example, extracellular signal-regulated kinase/mitogen-activated protein kinases (ERK/MAPK) and tyrosine kinases or the modulation of G-protein-coupled receptors have been shown to be non-transcriptionally recruited by estrogen in diverse tissues. In the heart and vasculature, the nongenomic mechanisms underlie estrogen induced short-term arterial vasodilation, inhibition of atherosclerotic lesions, and amelioration of ischemia/reperfusion-induced cardiac injury [12, 28].
ER mRNA and protein have been detected in both peripheral tissues and central nervous system [7,8,9]. ERα and ERβ immunoreactivity (ir) were found primarily localized to cell nuclei within select regions of the brain, including the olfactory bulb, cerebral cortex, septum, preoptic area, bed nucleus of the stria terminalis, amygdala, paraventricular hypothalamic nucleus, thalamus, ventral tegmental area, substantia nigra, dorsal raphe, locus coeruleus, and cerebellum. Extranuclear-ir was detected in several areas, including fibers of the olfactory bulb, CA3 stratum lucidum, and CA1 stratum radiatum of the hippocampus and cerebellum. In detail, nuclear ERα-ir was the predominant subtype in the hippocampus, preoptic area, and most of the hypothalamus, whereas it was sparse or absent from the cerebral cortex and cerebellum [7,8,9]. ERα and ERβ have distinct tissue expression patterns in both humans and rodents [7, 8, 29, 30], and gene-targeted animal models lacking these receptors exhibit distinct phenotypes [9, 31].
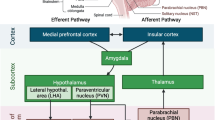
The rostral ventrolateral medulla (RVLM), where sympathetic premotor neurons are located, is a key region involved in regulation of blood pressure and hypertension [32]. ERα and ERβ are expressed in the RVLM [12, 13]. In rats, bilateral microinjection of 17β-estradiol in the RVLM elicited short-term cardiovascular depressive effects, while this effect was abolished by co-injection of ER antagonist [12]. This study further identified that ERβ, instead of ERα, induced significant decrease in systemic arterial pressure (SAP) and the power density of the vasomotor components of SAP signals [12]. Several studies have revealed the potential mechanisms in the 17β-estradiol-induced cardiovascular depressive responses, including the calcium current inhibition decrease in sympathetic tone [33] and the engagement of the inducible nitric oxide synthase (iNOS)-derived NO in the RVLM [12]. In addition to ERβ, extranuclear ERα also indirectly modulate the function of RVLM C1 bulbospinal neurons in selected afferents [33]. The high level of oxidative stress in the RVLM contributes to increased sympathetic outflow and hypertension [34], which is frequently seen in postmenopausal women. These studies emphasize that ERs in the RVLM is critical in the regulation of cardiovascular function (Fig. 1).

Schematic of the brain nuclear receptors regulate cardiovascular function. ER in the RVLM and the PVH decreases SAP and protects against aldosterone-induced hypertension; THR in the AH increases heart rate and blood pressure; MR in the NTS and the Amy increases salt appetite and signals salt satiation. MR-NTS neurons project to the PBN, PVH, and BNST, modulating sensory coding of salt taste and signaling salt satiation. ER estrogen receptor, RVLM rostral ventrolateral medulla, PVH paraventricular nucleus of the hypothalamus, SAP systolic arterial pressure, THR thyroid hormone receptor, AH anterior hypothalamus, MR mineralocorticoid receptor, NTS nucleus tractus solitarius, Amy amygdala, PBN parabrachial nucleus, BNST bed nucleus of the stria terminalis
The paraventricular nucleus of the hypothalamus (PVH) is another important site in the regulation of cardiovascular function [13, 35]. PVH neurons projecting to the RVLM play a regulatory role in the determination of the sympathetic outflow [35]. Using selective ER agonists and recombinant adeno-associated virus (AAV) carrying small interference (si) RNA to silence either ERα (AAV-siRNA-ERα) or ERβ (AAV-siRNA-ERβ) in the PVH and RVLM, researchers found that both PVH and RVLM ERβ, instead of ERα in these nuclei, contribute to the protective effects of estrogen against aldosterone-induced hypertension [13].
Mean arterial pressure and renal sympathetic nerve activities are significantly decreased following injection of estrogen into various brain regions including the nucleus tractus solitarius (NTS), RVLM, parabrachial nucleus (PBN), central nucleus of the amygdala (CNA) and the intrathecal space in estrogen-replaced rats [36]. Heart rate and vagal parasympathetic nerve activities are significantly decreased following injection of estrogen into the NTS, nucleus ambiguus (Amb), PBN and the intrathecal space [36]. This study underscores the importance of ER in other brain regions in the regulation of cardiovascular function. Further studies including using genetic rodent models are needed to further understand the regulatory role of ER in different brain regions in the regulation of cardiovascular function.
ERα is believed to be the primary ER that mediates estrogenic actions to prevent obesity and diabetes [37, 38], contributors to the prevalence of CVDs. The various brain regions responsible ERα’s regulatory effects on body weight and glucose homeostasis have been well studied. For example, ERα in amygdala neurons regulates body weight and ERα-expressing neurons in the ventrolateral subdivision of the ventromedial hypothalamic nucleus regulate glucose homeostasis [39,40,41]. However, the brain regions responsible for the direct and protective effects of ERα in cardiovascular function still need to be determined.
Mineralocorticoid receptor (MR)
Mineralocorticoids are a class of corticosteroids produced in the adrenal cortex, which influence salt and water balances (electrolyte balance and fluid balance). The primary mineralocorticoid is aldosterone [42, 43]. Increased levels of aldosterone tend to cause myocardial hypertrophy and remodeling, fibrosis, apoptosis, endothelial dysfunction, and impaired myocardial perfusion, which increasing the incidence of cardiovascular events [44].
The mineralocorticoid receptor (MR), also known as the aldosterone receptor, is a protein that in humans is encoded by the NR3C2 gene [45]. Aldosterone binding to MR results in reabsorption of sodium and water in exchange for potassium in various sites, causing an increased intravascular fluid retention and volume overload. This undesired impact in fluid balance further causes hypervolemia and promotes congestion in patients with chronic heart failure [42].
In addition, MR activation promotes hypertension, fibrosis, and inflammation [45, 46], while pharmacological blockade of the MR has protective effects in animal models of CVDs [47, 48] and preventive effects against mortality and morbidity in heart failure patients [49, 50]. Thus, a better understanding of the implications of the MR in the setting of CVDs is critical for refining treatments and improving patient care.
MR is considered as a ubiquitous transcription factor, which is expressed in peripheral tissues such as heart, kidney, salivary and sweat glands, skin, and also in central nervous system including hippocampus and hypothalamus [51]. In 1968, Bruce McEwen discovered that 3H-corticosterone administered to adrenalectomised rats is retained in neurons of hippocampus [52]. Since its discovery, a great body of studies were conducted to study the role of brain MR function in the cardiovascular regulation [53,54,55], especially the contributory role of salt intake [56, 57]. The NTS was found to be richly endowed with MR and the role of MR-NTS neurons in salt appetite regulation has been widely studied [58,59,60]. Neurons that abundantly express MR are located in the dorsal and ventral hippocampal CA1-4 pyramidal cell fields and the dentate gyrus, amygdala and lateral septum [53,54,55]. The MR-NTS neurons connect to the nearby area postrema, a periventricular region that is readily accessible to peptides and other compounds in the 4th ventricle. Intracerebroventricular (i.c.v.) infusion of the MR agonist aldosterone in the 4th ventricle increased salt appetite [59], while MR knockdown in the NTS prevented this aldosterone-induced salt appetite [60]. MR-NTS neurons project to the parabrachial nucleus, amygdala, paraventricular nucleus, hippocampus, and the bed nucleus of the striae terminalis (BNST), which subsequently signal salt satiation [58, 61, 62]. This neuronal network explained why interventions in the amygdala affected aldosterone regulation of salt appetite [63]. These studies highlight the importance of MR-NTS neurons in the regulation of cardiovascular function via regulation of salt appetite (Fig. 1).
Thyroid hormone receptor (THR)
Thyroid hormone (TH) has profound effects on cardiovascular function [64, 65]. TH increases myocardial inotropy and heart rate and dilates peripheral arteries to increase cardiac output (CO). Triiodothyronine (T3), the active form of TH, binds to NR proteins and mediates effects on the systemic vasculature include relaxation of vascular smooth muscle, resulting in decreased arterial resistance and diastolic blood pressure [65]. In patients with hyperthyroidism, cardiac contractility and cardiac output are enhanced and systemic vascular resistance is decreased, while in patients with hypothyroidism, the opposite phenotype is seen [66, 67]. In addition, the phenotype of the failing heart resembles that of the hypothyroid heart, both in cardiac physiology and in gene expression [68]. Changes in serum T3 levels in patients with chronic congestive heart failure are caused by alterations in TH metabolism suggesting that patients may benefit from T3 replacement in this setting [69]. The literature outlining the effect of TH replacement therapy for reducing the risk of CVDs have been comprehensively reviewed [66, 70,71,72,73]. Animal models showed that T3 deficiency in the brain influenced all aspects of brain development and physiology [74, 75], which urges the need of a complete understanding of T3 action in the brain.
TH receptors (THRs) have isoforms of THRs, THRα1, THRβ1 and THRβ2, which are predominantly responsible for mediating TH action [76, 77]. THRs are encoded by two genes called Thra and Thrb in mice, or THRA and THRB in humans [78]. THRs are transcription factors, which act mainly as heterodimers with RXRs. THR/RXR heterodimers bind to DNA at T3 response elements and regulate transcription initiation of target genes [78]. THRα1 mRNA is ubiquitous in the rodent brain [79], while THRβ1 mRNA only appears in the brain at a late developmental stage and in few cell types, predominantly in the hypothalamus, pituitary, retina, and cochlea [80]. Accordingly, most functional studies conclude that there is a predominant role of THRα1 in brain development, and a selective influence of THRβ1/2 in the development of sensory functions and the maturation of some specific neuronal populations [81].
In mice, THRα1 mRNA is predominantly expressed in the heart and brain, whereas THRβ1 is expressed in peripheral tissue including skeletal muscle, kidney, and liver [82]. THRα1 mRNA was widely expressed in the rat fetal neocortical plate, site of cortical neuronal differentiation, while THRβ1 transcripts were restricted in distribution, with prominent expression in zones of neuroblast proliferation such as the germinal trigone and the cortical ventricular layer [83].
The cardiovascular and metabolic effects of TH were previously thought to be mediated predominantly by THR isoforms expressed by peripheral tissues, including heart, skeletal muscle, and fat [84, 85]. However, recent studies suggest that TH can also modulate developmental, metabolic, and cardiovascular processes by acting on specific targets in the brain [86,87,88]. Mice expressing the mutant THRα1R384C displayed a mild bradycardia, accompanied by a reduced expression of nucleotide-gated potassium channel 2 mRNA in the heart [88]. Another study identified a previously unknown population of parvalbuminergic neurons in the anterior hypothalamus (AH) that requires THR signaling for proper cardiovascular function [14]. In this study, specific stereotaxic ablation of these cells in the mouse resulted in hypertension and temperature-dependent tachycardia [14], indicating a role in the central autonomic control of blood pressure and heart rate (Fig. 1). More studies are needed to address the role of brain THRs in the regulation of cardiovascular function.
NRs that regulate cardiovascular functions with unclear brain connection
Estrogen-related receptor (ERR)
Estrogen-related receptor (ERR), sharing sequence homology with ER, belongs to the subfamily of orphan NRs. ERRs have three family members, ERRα, ERRβ, and ERRγ [89,90,91]. ERRα has long been thought to not require an endogenous ligand, because it is activated by the transcriptional coactivator peroxisome proliferator-activated receptor γ (PPARγ) coactivator 1α (PGC-1α), which is necessary for postnatal cardiac mitochondrial biogenesis [92,93,94]. ERRα serves as a critical nodal point in the regulatory circuitry downstream of PGC-1α to direct the transcription of genes involved in mitochondrial energy-producing pathways in cardiac muscle [95]. One recent study showed that cholesterol is a potential endogenous ligand for ERRα that increased ERRα transcriptional activity [96]. This finding explained why ERRα is constitutively active because cholesterol is ubiquitous. Animal studies showed that knockdown the expression of ERRα and γ in heart after birth caused cardiomyopathy with an arrest in mitochondrial maturation [97], and ERRα null mice subjected to left ventricular pressure overload developed heart failure including chamber dilatation and reduced left ventricular fractional shortening [98]. In addition, using siRNA or inverse agonists to inhibit ERRα function impairs induction of mitochondrial gene expression and respiration by PGC-1α, which predisposes to cardiac remodeling [99, 100]. Disruption of ERRγ gene in mice blocked the high levels of expression in the fetal and postnatal mouse heart and resulted in lactatemia, electrocardiographic abnormalities, and death during the first week of life [101]. These studies indicate that ERRα and ERRγ are necessary for the maturation of both mitochondrial and structural processes during postnatal cardiac development and the adaptive bioenergetic response to hemodynamic stressors known to cause heart failure. Notably, ERRβ transcripts were increased in the fetal ERRα/γ knockdown mouse model, suggesting possibly as a compensatory regulatory response [97]. In addition, ERRβ has distinct functions in regulating energy metabolism in the adult cardiomyocyte, as shown that ERRβ is involved in maintaining maximal ATP generation in contracting cardiomyocytes [102]. ERRs have emerged as potential therapeutic targets for the treatment of heart failure in humans [103].
ERRα and ERRγ are both widely expressed in the brain [104, 105], while ERRβ expression is restricted to the hindbrain [106]. In the mouse brain, ERRα is abundantly expressed in several cortical regions, including orbitofrontal cortex, medial prefrontal cortex (mPFC), and cingulate cortex; highly expressed in the primary and secondary somatosensory cortex, piriform cortex, retrosplenial granular cortex, hippocampus, thalamus, mammillary nucleus, cerebellum, and several brainstem nuclei involved in transmission of sensory information; moderate levels of expression are observed in dorsolateral aspects of the striatum, septum, global pallidus, lateral hypothalamic area, and some midbrain structures including tegmentum and substantia nigra [104]. ERRγ transcripts were abundantly present in the isocortex, the olfactory system, cranial nerve nuclei and major parts of the coordination centers, e.g. reticular formation and major parts of the extrapyramidal motor systems. In addition, ERRγ expression was detected in trigeminal ganglion neurons [105]. There are a lot of studies investigating the functions of ERs and ERRs in the central regulation of energy homeostasis, learning and memory, and social behaviors [104], but the cardiovascular function is still largely unknown.
Androgen receptor (AR)
Low testosterone levels and androgen deprivation therapy are associated with increased atherosclerosis burden and increased risk of CVDs [107,108,109]. In men with heart failure, the testosterone replacement therapy is a significant predictor for an increase in peak oxygen consumption on exercise testing [110]. Acute and chronic administration of testosterone both demonstrate beneficial effects in the cardiovascular function [111, 112]. Intravenous administration of testosterone acutely increased cardiac output and reduced peripheral vascular resistance [111]. While during chronic testosterone replacement therapy, circulating levels of inflammatory mediators were reduced, potentially leading to a reduction of left ventricular muscle fibrosis [112].
The androgen receptor (AR) is activated by binding any of the androgenic hormones, including testosterone and dihydrotestosterone [113]. In male mice, the androgen-AR system regulates normal cardiac growth, cardiac adaptive hypertrophy and fibrosis during the process of cardiac remodeling under hypertrophic stress [113, 114]. Global knockout of AR in mice results in reduced heart size, impaired contraction, exacerbation of angiotensin II-induced cardiac fibrosis [114]. Depletion of ARs on the apolipoprotein E-deficient background increases atherosclerosis burden in male mice [115].
Besides the prostate, ARs are expressed in several other tissues including the brain [10, 116]. AR neurons are basically localized in limbic areas such as the hypothalamus [10]. In the rodent, brain AR distribution was distributed in the bed nucleus of the stria terminalis, lateral septum, medial amygdala, medial preoptic area, and ventromedial hypothalamic nucleus [117]. The neuron-AR knockout mice generated using the Nestin-Cre showed that central AR is required in testosterone-induced regulation of male-typical behaviors and gonadotrope and somatotropic axes [118]. While using the Synapsin-Cre, another group reported that neuronal AR deficiency leads to reduced insulin sensitivity in middle-aged mice [119]. Female neuron-AR knockout mice showed impaired kisspeptin/GnRH/LH cascade leading to compromised ovarian follicle dynamics [120]. However, whether the neuron-AR also regulates cardiovascular function is unknown yet.
Glucocorticoid receptor (GR)
Glucocorticoids are another class of corticosteroids, which are under regulation by the hypothalamic–pituitary–adrenal gland axis [121]. Research in both humans and animals have unraveled that glucocorticoid and their actions via binding to glucocorticoid receptor (GR) are highly involved in the genesis and development of CVDs [122,123,124]. Synthetic glucocorticoids are commonly prescribed in diverse CVDs, including infectious conditions such as rheumatic fever and myocarditis, structural conditions such as conduction defects and cardiomyopathy, and vascular conditions such as angina and acute myocardial infarction [125]. However, the severe side effects in diabetes, abdominal obesity, and hypertension, all of which are risk factors for CVDs, limit the therapeutic benefits of (synthetic) glucocorticoids [125]. Thus, it is essential to have comprehensive insight into the specific functions of glucocorticoids in the cardiovascular system.
The GR is activated either by the endogenous steroid hormone cortisol or by exogenous glucocorticoids and acts within the cardiovascular system via both genomic and non-genomic pathways. Polymorphisms of the GR are also reported to influence the progress and prognosis of CVDs [126]. The GR exerts numerous functions in the whole body including the brain [127,128,129]. In the brain, it has been implicated in feedback regulation of the hypothalamic–pituitary–adrenal axis, with potential deficits during aging and in depression. GRs are abundantly expressed in the hippocampus of rodent and human, indicating that this region can form an important target for corticosteroid effects [127]. The NTS abundantly expresses the GR and is a key brain region for processing autonomic and endocrine stress responses mediated by the HPA axis [128, 129]. One recent study found that ablated of GR in the hindbrain of mouse resulted in stress-related behavior and elevated circulating corticosterone and severe adrenal cortex disruption [130]. However, the direct role of brain GR in the regulation of cardiovascular function is still unclear.
Peroxisome proliferator-activated receptor (PPAR)
Peroxisomes are unique subcellular organelles that sequester diverse oxidative reactions [131]. Peroxisome proliferator-activated receptors (PPARs) are transcription factors of NR superfamily which also include RAR, ER, TR, GR, vitamin D receptor, and several other proteins involved in xenobiotic metabolism. PPARs act on DNA response elements as heterodimers with the retinoid X receptor (RXR). The PPARs, comprising of three subtypes: PPARα, PPARγ, and PPARβ/δ, have overlapping as well as different expression patterns and distinct functions in the metabolic control [4, 132, 133]. Besides expression in various peripheral tissues including liver, heart, kidney, adipose tissue, skeletal muscle and [132], PPAR mRNA and protein distribution in the brain were also investigated [134,135,136,137,138]. The expression of PPAR mRNA and protein in coarse brain regions as well as caudate putamen, hippocampus, hypothalamus, and thalamus were identified in rats [134, 135]. One recent study in mice showed that PPARα, PPARγ, and PPARβ/δ mRNA and protein were found to express in prefrontal cortex, nucleus accumbens, amygdala and ventral tegmental area [136]. Among the isotypes, PPARβ/δ was most widely and highly expressed across brain regions [137, 138] and PPARα was more highly expressed than PPARγ in all brain regions except the prefrontal cortex [136].
Lipid-derived substrates are their natural activating ligands. Omega-3 fatty acids are one kind of the natural ligands of PPARα [139], and their beneficial effects in cardiovascular functions are considered to be conveyed via PPARα [140]. Omega-3 fatty acids are also used to treat hyperlipidemia and hypertension and can significantly reduce the risk for sudden death caused by cardiac arrhythmias and all-cause mortality in patients with known coronary heart disease [141]. Like PPARα, PPARβ/δ improves cardiac function and ameliorates the pathological progression of cardiac hypertrophy, heart failure, cardiac oxidative damage, ischemia–reperfusion injury, lipotoxic cardiac dysfunction and lipid-induced cardiac inflammation [142, 143]. Furthermore, PPARβ/δ activation protects against diabetic cardiomyopathy [144, 145]. Previous studies showed that PPARβ/δ expression was decreased in cardiac muscle during the hyperglycemic state in diabetes mellitus, while in contrast, overexpression of this PPARβ/δ in cardiac cells diminished lipid accumulation and increased glucose metabolism [144, 145]. During the past decade, a large body of studies have demonstrated that PPARγ plays an important role in the cardiovascular system [146, 147]. Thiazolidinediones (TZDs) such as rosiglitazone and pioglitazone, are ligands for PPARγ, have long been used to treat type 2 diabetes and serve as a therapeutic target for CVDs [148]. Besides, PPARγ activation has a role against inflammation and suppresses the inducible nitric oxide synthase (iNOS) upregulation and reactive oxygen species (ROS) production [149].
The mRNA expression of PPARγ in hypothalamus is several folds higher than PPARα and PPARβ/δ [150]. PPARγ immunoreactivity was primarily observed in several brain regions including ARH, PVH, hypothalamic ventromedial neurons (VMH), the lateral hypothalamic area, and tyrosine hydroxylase-containing neurons in the ventral tegmental area [151]. Mice with neuron specific PPARγ knockout (PPARγ BKO) had reduced food intake and weight gain during high fat diet (HFD) feeding. When treated with rosiglitazone, these mice were resistant to rosiglitazone-induced hyperphagia and weight gain [152]. These findings suggest the critical effects of brain PPARγ on food intake, energy expenditure, and insulin sensitivity. However, the question of whether brain PPARs directly regulates cardiovascular function is still unanswered.
Progesterone receptor (PR)
Accumulated evidence has shown that progesterone exhibits beneficial effects on the cardiovascular function such as lowers blood pressure, inhibits coronary hyperactivity and has powerful vasodilatory and natriuretic effects [153]. Progesterone receptors (PRs), like other sex steroid hormone receptors, are members of a superfamily of ligand-activated transcription factors that regulate gene expression following hormone binding [154]. PRs are expressed in vascular cells [155, 156], and PR expression in vascular tissues is induced by estrogen [156]. One recent study found that ovariectomized female PR knockout mice had significantly greater vascular medial hypertrophy and vascular smooth muscle cell proliferation in response to vascular injury than did wildtype mice [154]. These findings indicate that an important role for the PR in regulating the response to vascular injury and vascular smooth muscle cell proliferation. Novel evidence showed that membrane PRs (mPRs), in addition to nuclear PRs, are also candidates for the rapid and cell-surface initiated progesterone actions in endothelial and smooth muscle vascular cells. Three mPR gene isoforms mPRα (PAQR VII), mPRβ (PAQR VIII), and mPRγ (PAQR V) identified in humans [157], are putative G protein-coupled receptors (GPCRs) for progesterone [158]. mPRβ human mRNA was found in the brain including spinal cord, cerebral cortex, cerebellum, pituitary, thalamus, and caudate nucleus [157], while mPRα and mPRβ rodent mRNA were seen hypothalamus [159, 160]. Several studies have examined the role of hypothalamic PR in the regulation of reproductive functions [161,162,163]. Microinjection of the PR antagonist, RU486, into the hypothalamic arcuate nucleus shortened LH pulse interval in the progesterone treated rats, while into the hypothalamic anteroventral periventricular nucleus reversed the prolonged cycle length and rescued the progesterone blockade LH surge [161]. Knockout of PR in kisspeptin neurons (enriched hypothalamic anteroventral periventricular and arcuate nuclei) in mice causes loss of LH surges, irregular estrous cycle, and infertility [162, 163]. Compared to the extensive studies regarding PR in reproductive function, the brain PRs in cardiovascular regulation are still largely unknown.
Retinoid acid receptor (RAR)
Retinoic acid (RA), the derivatives including all-trans RA, 13-cis RA, or 9-cis RA, carries out essential roles in heart development and after birth in the heart’s remodeling response to injury and disease [164, 165]. During early stages of cardiogenesis, alterations in RA levels are often associated with congenital heart defects such as abnormal folding and septation of the outflow tract and cardiac chambers [166]. Furthermore, RA increases myocyte cross-sectional area and induces cardiac hypertrophy in a dose-dependent way [167]. All-trans RA exerts its actions mainly through binding to RA receptor (RAR), while its enantiomer 9-cis RA binds to RAR or retinoid X receptor (RXR) [168].
RAR and RXR each has three different isoforms, termed α, β, and γ, which have overlapping as well as distinct expression patterns and functions during embryogenesis [169]. In vitro study showed that activation of RAR/RXR signaling protects cardiomyocytes from hyperglycemia, by reducing oxidative stress and inhibition of the renin-angiotensin system [170]. In addition, the selective RARβ2 agonist AC261066 reduced oxidative stress in an ex vivo murine model of ischemia/reperfusion [171] and later the same group performed in vivo study to show that RARβ2 agonist attenuated the development of contractile dysfunction and maladaptive remodeling in post-ischemic heart failure [172]. These studies identified that RARβ2 agonist as a potential target in the treatment of heart failure. However, there is lack of direct evidence to establish the relationship between brain RAR and cardiovascular regulation.
Retinoid X receptor (RXR)
The three RXR isoforms show tissue-specific expression, while their functions are partially overlapped. RXRα is predominantly expressed in metabolically active systems, such as liver, muscle, lung, kidney, and heart; RXRβ is ubiquitously expressed; whereas RXRγ is abundantly expressed in brain and skeletal muscle [173, 174].
RXR can form heterodimers with many other NRs including the RAR, THR, VDR, liver X receptor (LXR), PPARs, farnesoid X receptor, pregnane X receptor, and CAR [175]. RAR/RXR form heterodimers and regulate RA-induced expression of genes that control cell fate specification, proliferation, and differentiation [176]. RAR/RXR knockout mice developed cardiovascular dysfunction including heart malformations, defects in the conduction system and heart failure [177]. Overexpression of RAR or RXR induces dilated cardiomyopathy, depressed cardiac function, and congestive heart failure [178]. Due to the complex dimerization pattern, it is difficult to conclude whether a particular defect in RXR null mice is attributed to impaired RAR/RXR signaling or to an impairment is signaling by other heterodimeric partners of RXR or by RXR homodimers [176].
RXR mediates brain clean-up after stroke [179]. Deleted RXRα in myeloid phagocytes demonstrated worsened late functional recovery and developed brain atrophy that was larger in size than that seen in control mice [180]. In rats, the RXR agonist bexarotene attenuated neuroinflammation and improved neurological deficits after subarachnoid hemorrhage [181]. These studies highlight the important and protective role of RXR in cerebrovascular function, which further raises up the interest of how brain RXR in the regulation cardiovascular function.
Retinoic acid-related orphan receptor (ROR)
The RAR-related orphan receptor (ROR) belongs to a subfamily of the THR, which is a subfamily of the NRs [182]. The ROR subfamily contains three members: RORα (NR1F1), RORβ, and RORγ.
RORα plays an essential role in the regulation of cardiovascular function including circadian rhythm, development, and metabolism [183,184,185]. One human retrospective study found that RORα mRNA expression levels in patients with acute myocardial infarction is significantly higher than that in patients with stable coronary artery disease [186]. Serum antibody levels of Claudin domain containing 1 (CLDND1)-derived peptides are elevated in patients with cerebral infection, CVDs or diabetes mellitus as compared to healthy controls. RORα overexpression in human brain endothelial cells enhanced CLDND1 transcript expression, while in contrast, siRNA-mediated knockdown of RORα significantly decreased CLDND1 transcription [187].
RORγt, has been shown to contribute to the development of hypertension in spontaneously hypertensive rat and may be potential therapeutic targets [188]. Mice with RORα deficiency demonstrated significantly augmented diastolic dysfunction and cardiac remodeling, which were associated with aggravated myocardial apoptosis, autophagy dysfunction, and oxidative stress, while restoration of cardiac RORα levels significantly improved cardiac functional and structural parameters [189]. Consistent with genetic manipulation, pharmacological activation of RORα by melatonin and SR1078 (a synthetic agonist) showed beneficial effects against diabetic cardiomyopathy, while the RORα inhibitor SR3335 significantly exacerbated cardiac impairments in diabetic mice [189]. Collectively, these findings suggest that cardiac-targeted manipulation of nuclear melatonin receptor RORα may hold promise for delaying diabetic cardiomyopathy development.
RORα is widely expressed in many tissues, including cerebellar Purkinje cells, the liver, thymus, skeletal muscle, skin, lung, adipose tissue, and kidney [190, 191]. RORγ has a similar broad pattern of expression but is observed at very high levels within the thymus. RORβ has a more restricted pattern of expression relative to the other RORs and is found in regions of the central nervous system (CNS) that are involved in the processing of sensory information, the retina and the pineal gland [192].
Previous studies reported that RORs in the circadian rhythm, metabolism, immune function, and tumorigenesis [185, 193, 194]. Mice with a loss-of-function mutation in RORα had alterations in the circadian oscillator, indicating an essential role for this receptor in normal circadian function [195]. Circadian misalignment causes adverse metabolic and cardiovascular consequences [196]. Current studies imply that brain RORs may play critical roles in the regulation of cardiovascular functions, directly or indirectly, but further studies are needed.
Vitamin D receptor (VDR)
Vitamin D has a protective role in CVDs and vitamin D deficiency is associated with increased risk of hypertension, atherosclerosis, and heart failure [197]. It is noticed that a seasonality in patients suffering from CVDs could be attributed to the low 25-hydroxyvitamin D levels patients had in winter [198]. Vitamin D deficiency is associated with arterial stiffness, hypertension, left-ventricular hypertrophy, and endothelial dysfunction in patients with chronic kidney disease, as well as in normal subjects [199,200,201]. However, recent clinical intervention studies failed to prove the causal relationship between vitamin D supplementation and beneficial effects on cardiovascular health. In the RECORD (Randomised Evaluation of Calcium and/OR vitamin D) trial, daily vitamin D (800 IU) or calcium supplementation did not affect mortality, vascular disease, cancer mortality, or cancer incidence [202]. Another randomized, placebo-controlled trial found that inflammatory milieu was reduced in patients suffering from congestive heart failure, but left ventricular function remained unchanged after administration of 2000 IU vitamin D for nine months [203].
The vitamin D receptor (VDR), a member of the NR/steroid hormone receptor superfamily, functions as ligand-activated, transcriptional regulatory proteins. Thus, VDR selectively binds to 1,25-(OH)2-vitamin D3, the active form of vitamin D, and then forms a heterodimer with the RXR [204]. Previous immunohistochemical evidence showed that VDR was present in both neurons and glia of the human [205] and the rodent brains [206]. VDR has been noted in key dopaminergic brain regions in both the human brain [205] and the rat brain [206]. One study further showed that VDR is present in the nucleus of tyrosine hydroxylase-positive neurons in both the human and rat substantia nigra [207]. Emerging evidence indicates that brain VDR is critical in the vulnerability to central nervous system diseases including Parkinson’s Disease [208] and multiple sclerosis [209]. However, whether brain VDR regulates cardiovascular function is unclear.
REV-ERB
The NRs REV-ERBα and -β regulate circadian rhythm and modulate glucose and lipid metabolism and inflammatory response [210,211,212]. REV-ERBs lack C-terminal helix, the canonical NR activation domain, and thus function as a transcriptional repressor [213].
A great number of studies highlight the importance of REV-ERB in circadian rhythm and metabolic functions including glucose homeostasis and energy homeostasis [212, 214,215,216,217,218], while emerging studies point out the important role of REV-ERBα/β in cardiovascular function [219,220,221]. Mice with cardiac-specific deletion of REV-ERBα/β and inducible ablation of REV-ERBα/β in adult hearts both displayed progressive dilated cardiomyopathy and lethal heart failure [219]. This study further identified that circadian disruption, which was implicated in heart diseases, was seen in the knockout mice [219]. Moreover, altered temporal patterns of cardiac REV-ERB gene expression was associated with the cardiac dilation severity in human hearts with dilated cardiomyopathy [219]. RER-ERB agonists, SR9009 and SR9011, were shown to improve hyperglycemia, dyslipidemia, and skeletal muscle oxidative capacity through modulation of mitochondrial number and oxidative function [211]. Recently, long-term treatment with SR9009 was reported to reduce atherosclerotic plaque in low-density lipoprotein receptor-deficient mice fed a Western diet [220]. Moreover, REV-ERB agonist SR9009 treatment significantly reduced post-myocardial infarction mortality and improved cardiac function through modulating inflammation and remodeling process [221]. These findings highlight the critical role of REV-ERBα/β in CVDs, especially the progression of myocardial infarction and heart failure.
REV-ERBα mRNA and protein were found in different brain regions including hippocampus, cortex, cerebellum, nigra, striatum, and hypothalamus in mice [222]. Accumulating evidence implies that the circadian NR REV-ERBα is involved in mood regulation. Altered midbrain REV-ERBα induces mania-like behavior and a hyperdopaminergic state and defective hippocampal function and memory [223, 224]. Most recently, the specific deletion of both REV-ERBs in the tuberal hypothalamic nuclei (including the ARC, VMH, and DMH) revealed the crucial role of REV-ERB in hypothalamic control of food intake and diurnal leptin sensitivity in diet-induced obesity [225]. Further studies are needed to explore whether central REV-ERB directly regulate cardiovascular function.
Farnesoid X receptor (FXR)
The Farnesoid X receptor (FXR), also known as bile acid receptor, is a NR activated by bile acids. FXR regulates bile acid synthesis, conjugation, and transport, as well as various aspects of lipid and glucose metabolism [226]. Recent evidence shows that FXR is expressed in cardiomyocytes and endothelial cells and acts as a novel functional receptor in cardiac tissue, regulates apoptosis in cardiomyocytes, and contributes to myocardial ischemia/reperfusion injury [227]. Pharmacological inhibition or genetic ablation of FXR significantly reduced myocardial apoptosis, decreased infarct size, and improved cardiac function in ischemic/reperfused myocardium [227].
FXR expression was found in mouse and human brain [228, 229]. Further, multiple bile acids could be detected in the central nervous system [229,230,231], which may regulate FXR function in the brain. Recent studies have shown that FXR expressed in frontal cortex and hippocampus of mouse brain is involved in the pathogenesis of some neurological disorders, such as depression and Alzheimer's disease [230, 232, 233]. Besides, hippocampal FXR may help improving the insulin sensitivity in the Alzheimer's disease model rats, while FXR in the dorsal vagal complex (DVC) of the brain is involved in the insulin resistance in rats [230, 234]. It was shown that short-term HFD induces taurochenodeoxycholic acid (TCDCA, a FXR ligand) in the plasma and subsequently in the DVC of the brain induces insulin resistance in rats, while genetic knockdown or chemical inhibition of FXR in the DVC of HFD rats reversed insulin resistance [234]. More importantly, hippocampal FXR was found to regulate brain derived neurotrophic factor (BDNF) biosynthesis, which suggests that central FXR may play an important role in the regulation of metabolism considering the critical role of BDNF in the energy homeostasis [233, 235]. However, there is yet direct evidence to show that central FXR regulates cardiovascular function.
Liver X receptor (LXR)
The liver X receptors α and β (LXRα and LXR β) are members of the NR family of proteins that play a crucial role in the regulation of cholesterol and lipid homeostasis [236, 237]. In the nucleus, LXRs form obligate heterodimers with the RXR and are bound to LXR response elements in regulatory regions of target genes. LXRα and LXR β are expressed in an overlapping but nonidentical pattern. LXRα is abundantly expressed in the liver, adipose tissue, heart, skeletal muscle, kidney, and lung while LXRβ is ubiquitously expressed [238]. The endogenous ligands for the LXRs are oxysterols, the oxidized derivatives of cholesterol. LXRs function as whole body cholesterol sensors [238].
Besides their established role in metabolic homeostasis and disease, there is mounting evidence suggests that LXRs have beneficial effects in the heart. The emerging role of LXRs in cardiac pathophysiology and heart failure was previously summarized [239]. LXRs prevents pathogenic accumulation of cholesterol in macrophages by enhancing the rate of cholesterol efflux [240]. Thus, LXRs exerts protective effects in the development of atherosclerosis and subsequent myocardial infarction and systolic heart failure. LXR agonist significantly reduces atherosclerosis in mouse models [241]. On the other hand, LXRs prevent against the presence of co-morbidities such as hypertension, atrial fibrillation, and diabetes [239]. These observations support the notion that LXRs may represent a novel therapeutic target for the treatment of human CVDs.
Both LXR isoforms are expressed in the brain [135] and play neuroprotective roles in the CNS. LXRs have been widely involved in the development of CNS and implicated in the regulation of brain function, such as anxiety [242] and neurodegenerative diseases [237, 243]. More importantly, LXRs are expressed in the hypothalamus, which suggested that LXR may exert metabolic regulation in the brain [244]. Recent studies found the knock-down LXRs in vivo or in LXRαβ−/− mice activated TSH-releasing hormone (TRH) signaling in the PVH increased the activity of the hypothalamic-pituitary-thyroid axis, ultimately leading to the enhanced browning of subcutaneous adipose tissue thereby ameliorating obesity outcomes [245, 246]. LXRs also show neuroprotective roles in different experimental stroke models: stabilizing the integrity of brain capillaries, improving stroke outcome, and suppressing neuro-inflammation; and LXR activation protects hippocampal microvasculature through improving the microvascular architecture in Alzheimer Disease models [247,248,249,250]. These studies suggest the potentially preventative role of central LXR in the regulation of cardiovascular function.
Small heterodimer partner (SHP)
Like DAX1, small heterodimer partner (SHP) is also an atypical orphan NR and acts as a negative regulator of receptor-dependent signaling pathways [251].
SHP is highly expressed in liver, heart, brain, pancreas, and small intestine [252]. In vitro study found that SHP upregulation upon high-fat feeding leads to lipid accumulation, insulin resistance and inflammation in cardiomyocytes [253]. In vivo study showed that SHP-null mice showed a cardiac hypertrophy. In addition, the metformin-induced antihypertrophic effect was diminished in SHP-null mice or by SHP small interfering RNA in cardiomyocytes [254]. SHP gene is regulated by several NRs, including FXR, liver receptor homolog-1, ERRγ, ERα, LXR, and SF-1 [252, 255, 256]. SHP itself is also an important modulator of nuclear signaling pathways by acting as a repressor antagonizing the activities of many NRs. Like DAX-1, SHP inhibits interaction with ER and its function [257, 258]. SHP mediates the anti-inflammatory actions of LXRs through differential regulation of receptor SUMOylation specifically in astrocytes, thereby revealing potential avenues for therapeutic development in diseases associated with brain inflammation [259]. Other functions in the CNS have not been reported yet.
Constituitive androstane receptor (CAR)
The constitutive androstane receptor (CAR) is defined as xenobiotic NR that expressed in hepatocytes and activates genes involved in drug disposition, lipid homeostasis, and cell proliferation [260,261,262]. The metabolic function of CAR has been studied, which has a therapeutic potential in the prevention and treatment of type 2 diabetes and obesity [263, 264]. CAR was reported to interact with hypoxia inductile factor to regulate the hypoxia signaling for the development of CVDs [265]. Recent human [266, 267] and animal [268, 269] studies reported that mRNA and protein expression of CAR in the brain, including the cerebral cortex, hippocampus, amygdala, hypothalamus, and the basal ganglia. However, the role of brain CAR in the regulation of cardiovascular function is unknown.
Steroidogenic factor 1 (SF-1)
Steroidogenic factor-1 (SF-1, Ad4BP) is a pivotal determinant of endocrine function within the hypothalamic-pituitary–gonadal axis and adrenal cortex and an essential factor in sex differentiation [270]. SF-1 transcripts were originally detected in adrenocortical cells, testicular Leydig cells, and ovarian theca and granulosa cells, and recently found in the dorsomedial part of ventromedial hypothalamic nucleus (VMH) neurons [39, 271, 272]. These findings raised the intriguing possibility that SF-1 playes critical roles that extend beyond the maintenance of steroidogenic capacity within the primary steroidogenic tissues.
Following studies proved this possibility. Embryonic deletion of SF-1 results in perinatal lethality and malformation of the VMH, suggesting that SF-1 is essential for the normal development of the VMH [271, 273, 274]. The VMH has been demonstrated to modulate the sympathetic nervous system (SNS) activity in the periphery. Rats with lesions in the VMH showed reduced sympathetic outflow and heart rate [275]. In contrast, VMH activation by electronic stimulation or drug injection could increase the SNS activity [276, 277]. One study showed that activation of the VMH, using the chemogenetic tool, the Designer Receptors Exclusively Activated Designer Drugs (DREADDs), exacerbated cardiac remodeling and increased systolic blood pressure in hypertension rats via increased the sympathetic nervous system activity [278]. VMH is an important nucleus in responding to emotional stress, which is also a risk factor for CVDs [279]. SF-1 neurons in VMH play a pivotal role in the regulation of glucose and energy homeostasis [39, 280]. But currently there are no studies to examine the role of SF-1 neurons in VMH in the regulation of cardiovascular function.
Many studies have shown that the deletion of various key factors specifically in the SF-1 neurons, such as leptin receptor, ERα, phosphoinositide 3-kinase and AMPK, etc. involved in the regulation of energy homeostasis and obesity [272, 281,282,283,284,285,286]. VMH may be the origin of leptin-mediated sympathoexcitation which contributes to obesity-related hypertension [287], but no cardiovascular effects were reported in these mutant mouse models.
Liver receptor homolog‐1 (LRH‐1)
The orphan NR liver receptor homolog‐1 (LRH‐1) is highly expressed in liver, pancreas, and ovary [288]. Recent evidence further shows nuclear LRH-1 immunoreactivity in the heart, brown adipose tissue, and also ARH and PVH of the brain [289].
LRH-1 is constitutively active, which indicating its function occurs via interactions with co-activators and corepressors, as well as through posttranslational modifications, phosphorylation, and sumoylation [290]. Co-activators include PGC-1α, CBP, FXR, MBF-1, SRC-1/3, and phospholipids, while co-repressors include Prox1, SMRT, SHP, DAX1 [290]. FXR and SHP are NRs as discussed above.
A growing number of studies showing that LRH-1 plays critical roles in the regulation of development, cholesterol transport, bile acid homeostasis, and steroidogenesis [291], which are associated with cardiovascular functions. It has been shown that in vitro ligand of LHR-1, dilauroyl phosphatidylcholine, protects against hepatic steatosis and insulin resistance in mice fed with high-fat in an LRH-1–dependent manner [292]. Further study also suggested that LRH-1 is a critical component of the hepatic glucose-sensing system and integrates glucose and lipid homeostasis in the postprandial phase [293]. Recently, researchers also found that eliminating LRH-1 in the hepatocyte of the adult mice can profoundly affect the hepatic lipid metabolism and the glucose tolerance [294]. Heterozygous deletion of LRH-1 only causes mild body weight gain in HFD-fed mice [295]. In addition, using RNA interference and adenovirus-mediated overexpression, one study reported that LRH-1 directly regulates Apolipoprotein A1 (ApoA1) gene transcription [296]. ApoA1 is the primary protein component of high-density lipoprotein. Elevated ApoB and decreased ApoA1 are associated with increased risk of CVDs [297, 298]. Further studies will be needed to test whether LRH-1 directly regulates cardiovascular functions.
One recent study showed that LRH-1 is expressed in kisspeptin neurons in the arcuate nucleus but not in the anteroventral periventricular of the hypothalamus in female mice [299]. Kisspeptin is a neuropeptide, encoded by kisspeptin 1 (Kiss1) gene, which primarily acts as the regulator of reproductive functions [300]. Deletion of LRH-1 from kisspeptin neurons in mice led to decreased Kiss1 expression in the arcuate nucleus, associated with decreased serum FSH levels, impaired follicle maturation, and prolongation of the estrous cycle [299]. It was recently reported that kisspeptin could change the morphology and structure of myocardial cells and regulate the expression of certain genes and proteins related to heart diseases [301]. LRH-1’s precise roles in the CNS and furthermore whether it directly regulates cardiovascular function remain to be elucidated.
NRs without known cardiovascular functions
Nerve growth factor 1B (NGF1B)-like receptors: Nur77, Nurr1, NOR-1
Nerve growth factor IB (NGF1B)-like receptors (nuclear hormone receptor 4A, NR4A) are a group of orphan NRs, comprising three members Nur77, Nurr1, NOR-1 [302, 303]. Since the endogenous ligand has not been identified, NGF1B-like receptors are considered to be constitutively active and the transcriptional activity is regulated mainly by the expression levels, and also by intracellular localization, modifications and protein–protein interactions [304].
All three members of the NR4A are expressed in CNS and peripheral tissues with high metabolic activity such as skeletal muscle, liver, heart and adipose tissues [302]. In the brain, the expression of NR4A in different brain area may show different functions, for example, in the hippocampus, NR4A contributes to memory formation [305], and in the hypothalamus, Nur77 mediates the anorexigenic effects of leptin [306]. Decreased expression of Nur77 in the hypothalamus has been identified in obese mice, which suggest that decreased hypothalamic Nur77 may contribute to development of obesity. Consistent with this, global Nur77 knockout mice develop obesity associated with increased food intake and decreased energy expenditure and are insensitive to the anorexigenic effects of leptin. More importantly, knockdown of Nur77 specifically in the hypothalamus blunts the actions of leptin on feeding and body weight [306]. Therefore, hypothalamic Nur77 may function as a positive modulator in leptin signaling and can contribute to the regulation of body weight balance. NOR-1 may also promote food intake and body weight gain through its actions in the brain, as the injection of a siRNA oligonucleotide against NOR-1 into the third cerebral ventricle markedly suppressed food intake and body weight in mice [307].
NR4A also regulates the dopamine neurons activity in the midbrain. Activation of the SNS leads to the release of its effectors, the catecholamines (Norepinephrine and epinephrine), from post-ganglionic cardiac sympathetic neurons. Norepinephrine and epinephrine are synthesized from the tyrosine and dopamine, in which tyrosine hydroxylase (TH) is the rate-limiting enzyme. In the brain of Nur77-knockout mice, TH mRNA expression and levels of the dopamine precursor are higher than WT mice, suggesting the enhanced TH activity [308]. Nurr1 is also essential for the development of midbrain dopamine neurons. Nurr1-knockout mice die soon after birth due to complete lack of midbrain dopaminergic neurons, and Nurr1 heterozygous mice have decreased TH expression and catecholamine levels in the midbrain [309, 310]. Therefore, Nurr1 is essential for both survival and final differentiation of dopaminergic neurons.
However, whether the deletion of NR4A in these neurons will induce any cardiovascular phenotype still need further investigation. Most recently, it is reported that the old Nurr1 knockout mice (both male and female) have higher heart rate without alteration in systolic blood pressure; young female Nurr1 knockout mice showed an increased heart rate in comparison to their WT littermates [311], but the molecular mechanism is still unclear.
The germ cell nuclear factor (GCNF)
The orphan NR germ cell nuclear factor (GCNF), also known as RTR (retinoid receptor-related testis-associated receptor), is essential for embryonic development and reproduction [312,313,314,315]. In the mouse, GCNF is expressed in the developing nervous system, placenta, embryonic gonads, and adult ovaries and testes [316]. In vitro study established and characterized a GCNF−/− embryonic stem cell line and reported that GCNF plays a central role in repressing the embryonic stem cell phenotype during RA-induced differentiation [317]. Loss of GCNF in the oocyte of female mice caused impaired fertility, which was due to prolonged diestrus of estrus cycle and aberrant steroidogenesis [315]. However, GCNF’s function in the development of obesity and CVDs is unknown.
Testicular receptor (TR)
There are two forms of the testicular receptor (TR), TR2 and TR4. TR2 is robustly expressed in the developing olfactory epithelium and in more caudal regions of the brain, including the cortex, ventral forebrain structures, and thalamus [318]. TR4 is implicated in multiple pathological processes, including cerebellar development [319, 320], reproductive function [321, 322], and cancer development [323, 324]. Currently, there are no studies of TRs in the regulation of cardiovascular function.
Pregnane X receptor (PXR)
Pregnane X receptor (PXR) is an important component of the body’s adaptive defense mechanism against toxic substances including foreign chemicals (xenobiotics) [325]. PXR is activated by many endogenous and exogenous chemicals including steroids, antibiotics, antimycotics, bile acids, hyperforin, and other compounds such as meclizine, cafestol and forskolin [326, 327]. One recent study reported that Bisphenol A significantly increased atherosclerotic lesion area in the aortic root and brachiocephalic artery in PXR-Humanized ApoE deficient mice [328]. This study may suggest the role of PXR in the development of atherosclerosis, but currently there are no studies regarding the brain PXR in cardiovascular control.
Chicken ovalbumin upstream promoter transcription factor (COUP-TF)
Chicken ovalbumin upstream promoter transcription factors (COUP-TFs) are essential for the development of multiple tissues and organs, such as COUP-TFII is required for angiogenesis and heart development and COUP-TFI functions in neural development [329, 330]. COUP-TFII is a NR classified as an orphan due to the lack of a known natural ligand. COUP-TFII affects mitochondrial function, impairs metabolic remodeling, and has a key role in dilated cardiomyopathy [331]. Increased COUP-TFII expression is associated to brain arteriovenous malformations in humans [331]. COUP-TFII is important for migration of cortical interneurons from caudal ganglionic eminence (CGE) to the neocortex in rodents and human fetal brain [332, 333]. By using focal in utero labeling of the preoptic area (POA), one study [334] showed that switching on/off the transcription factor COUP-TFII and the receptor Neuropilin-2 (Nrp2) directs the POA-derived neurons toward either the amygdala or cortex. This study revealed an essential role of COUP-TFII/Nrp2 expression dynamics in the development of the amygdala and cortex. However, how brain COUP-TFII regulates cardiovascular function is unknown.
DAX-1
DAX-1 [dosage-sensitive sex reversal-adrenal hypoplasia congenita critical region on the X chromosome, gene 1] is an atypical orphan NR. DAX1 is expressed in the adrenal gland, gonad, pituitary gland, and hypothalamus [335].
DAX-1 emerges as a global negative regulator of genes involved in steroid hormone production and metabolism in steroidogenic tissues [336,337,338,339,340]. Previous study in Ahch (the mouse DAX-1 homologue) knockout mice indicates that DAX-1 may play a direct role in spermatogenesis [341]. Recent results obtained in patients of adrenal hypoplasia congenita suggest that DAX-1 is also required for spermatogenesis in humans, independent of gonadotropin and testosterone production [342]. Impaired spermatogenesis is a risk factor for CVDs [343]. It has been reported that DAX-1 can inhibit SF-1 transcriptional activity [337, 338]. In addition, DAX1 has been shown to repress the transcriptional transactivation of ER [339], AR [344] and PR [340]. Considering that all these NRs, especially ER, AR and PR, play key roles in the regulation of cardiovascular functions as described previously, DAX-1 may also be involved in this process which is unknown yet.
Tailless (TLX)
The orphan NR tailless (TLX) is exclusively expressed in the brain [345]. TLX has been identified as a fundamental regulator of adult neural stem cells and neurogenesis which involved in the learning and behavior [346, 347]. It has been reported that the protein level of TLX in the striatum, cortex, and hippocampus of mice was attenuated when fed with HFD [348]. However, the function of TLX in the development of obesity and CVD is not clear.
The photoreceptor cell-specific nuclear receptor (PNR)
The photoreceptor cell-specific NR (PNR) is a member of the nuclear receptor super family of intracellular transcription factors. PNR is exclusively expressed in the retina [349] and plays a role in the regulation of signaling pathways intrinsic to the photoreceptor cell function [349]. There is no report about effects of PNR on obesity and CVD.
Hepatocyte nuclear Factor‐4 (HNF4)
Hepatocyte nuclear factor‐4 (HNF4α) is a highly conserved member of the NR superfamily. HNF4α is expressed in liver, kidney, small intestine, colon, and pancreas [350, 351]. The essential fatty acid linoleic acid has been identified as the endogenous ligand for HNF4 [352]. Like LXR, HNF-4 emerged recently as another key regulators in reverse cholesterol transport, which prevents lipid accumulation and represents a promising target of new therapeutic agents for the prevention and treatment of metabolic diseases and atherosclerosis [353]. However, the studies in HNF-4 are much less than LXR and its central role is not clear.
Summary
Given the great number of NRs, the abundant expression of NRs expressed in many brain regions and peripheral tissues, and their broad roles in the regulation of cardiovascular functions, here we summarized the current pharmaceutical and genetic evidence, brain expression, functional brain regions and effects on cardiovascular functions in Table 1. Numerous studies demonstrated the abundant expression of the NRs in almost all brain regions including cerebral cortex, cerebellum, hypothalamus, thalamus, pituitary gland, pineal gland, amygdala, hippocampus, and midbrain [7,8,9, 14, 39, 53,54,55, 104, 134, 135, 205,206,207, 271, 272, 289, 305, 306, 318]. However, the functional studies from the perspective of brain NRs in the regulation of cardiovascular functions are only available for a few NRs [12,13,14, 58,59,60,61,62, 88]. Briefly, in this review we encapsulated that the ER in RVLM and PVH decreasing SAP and protecting against hypertension, the MR in NTS and projection neurons increasing salt appetite and satiation, and the THR in parvalbuminergic neurons of anterior hypothalamus augmenting heart rate and blood pressure in Fig. 1. In consideration of the many other NRs with unknown brain connections but have clear cardiovascular regulations, we call more efforts in this field. One of the reasons for these efforts is that NRs could be suitable therapeutic targets for CVDs. Raloxifene, a selective estrogen receptor modulator (SERM) was reported to favorably reduce serum levels of low-density lipoprotein (LDL) cholesterol in postmenopausal women in a randomized, placebo-controlled clinical trial [354]. One contemporary primary prevention cohort study showed that elevated LDL cholesterol had the highest absolute risk of CVDs including myocardial infarction and atherosclerosis [355]. Moreover, pravastatin, a medication used to lower cholesterol, was found to reduce the incidence of fatal coronary event [356]. Besides SERM, PPARγ agonist is another example to imply the potential of NRs in treatment of CVDs. Clinical trials have found that PPARγ agonists pioglitazone and rosiglitazone could reduce systolic and/or diastolic blood pressure, respectively [357, 358]. Long-term reduction of blood pressure might be expected to decrease the risk of congestive heart failure [359]. These studies underscored the importance of NRs in the treatment of CVDs and the necessity to improve our understanding of the mechanisms of NRs in the cardiovascular control, especially via the brain connection. Overall, a better understanding of the cardiovascular function of brain NRs may facilitate the development of novel targeted therapies in future.
The emerging role of central NRs in the regulation of cardiovascular functions have gradually drawn our attention. Some NRs show great potential to regulate brain–heart interactions, yet we must admit that we are in the very early phase to establish the causative role of brain NRs in the etiology of the CVDs. The function of NRs in the brain is highly dependent on the brain region, cell type, or developmental stages, thus future studies using animal models with temporally controlled genetic manipulations in a specific brain region, cell type, or a circuit-related neuron population would help clarify their functions.
Availability of data and materials
Not applicable.
References
Robinson-Rechavi M, Escriva Garcia H, Laudet V. The nuclear receptor superfamily. J Cell Sci. 2003;116(pt 4):585–6. https://doi.org/10.1242/jcs.00247.
Popov VM, et al. The functional significance of nuclear receptor acetylation. Steroids. 2007;72(2):221–30.
Aranda A, Pascual A. Nuclear hormone receptors and gene expression. Physiol Rev. 2001;81(3):1269–304.
Tyagi S, et al. The peroxisome proliferator-activated receptor: a family of nuclear receptors role in various diseases. J Adv Pharm Technol Res. 2011;2(4):236–40.
Huss JM, Kelly DP. Nuclear receptor signaling and cardiac energetics. Circ Res. 2004;95(6):568–78.
Knowlton AA, Lee AR. Estrogen and the cardiovascular system. Pharmacol Ther. 2012;135(1):54–70.
Mitra SW, et al. Immunolocalization of estrogen receptor beta in the mouse brain: comparison with estrogen receptor alpha. Endocrinology. 2003;144(5):2055–67.
Simerly RB, et al. Distribution of androgen and estrogen receptor mRNA-containing cells in the rat brain: an in situ hybridization study. J Comp Neurol. 1990;294(1):76–95.
Speirs V, et al. Distinct expression patterns of ER alpha and ER beta in normal human mammary gland. J Clin Pathol. 2002;55(5):371–4.
Fernandez-Guasti A, et al. Sex differences in the distribution of androgen receptors in the human hypothalamus. J Comp Neurol. 2000;425(3):422–35.
Dampney RA. Central neural control of the cardiovascular system: current perspectives. Adv Physiol Educ. 2016;40(3):283–96.
Shih CD. Activation of estrogen receptor beta-dependent nitric oxide signaling mediates the hypotensive effects of estrogen in the rostral ventrolateral medulla of anesthetized rats. J Biomed Sci. 2009;16:60.
Xue B, et al. Estrogen receptor-beta in the paraventricular nucleus and rostroventrolateral medulla plays an essential protective role in aldosterone/salt-induced hypertension in female rats. Hypertension. 2013;61(6):1255–62.
Mittag J, et al. Thyroid hormone is required for hypothalamic neurons regulating cardiovascular functions. J Clin Invest. 2013;123(1):509–16.
Battaglini D, et al. Brain–heart interaction after acute ischemic stroke. Crit Care. 2020;24(1):163.
Chen Z, et al. Brain–heart interaction: cardiac complications after stroke. Circ Res. 2017;121(4):451–68.
Murphy E. Estrogen signaling and cardiovascular disease. Circ Res. 2011;109(6):687–96.
Mendelsohn ME, Karas RH. The protective effects of estrogen on the cardiovascular system. N Engl J Med. 1999;340(23):1801–11.
Mendelsohn ME. Protective effects of estrogen on the cardiovascular system. Am J Cardiol. 2002;89(12A):12E-17E; discussion 17E-18E.
Iorga A, et al. The protective role of estrogen and estrogen receptors in cardiovascular disease and the controversial use of estrogen therapy. Biol Sex Differ. 2017;8(1):33.
Mozaffarian D, et al. Heart disease and stroke statistics–2015 update: a report from the American Heart Association. Circulation. 2015;131(4):e29-322.
Reckelhoff JF. Gender differences in the regulation of blood pressure. Hypertension. 2001;37(5):1199–208.
Stoberock K, et al. Gender differences in patients with carotid stenosis. Vasa. 2016;45(1):11–6.
Kawecka-Jaszcz K, et al. The effect of hormone replacement therapy on arterial blood pressure and vascular compliance in postmenopausal women with arterial hypertension. J Hum Hypertens. 2002;16(7):509–16.
Hao F, et al. Estrogen replacement reduces oxidative stress in the rostral ventrolateral medulla of ovariectomized rats. Oxid Med Cell Longev. 2016;2016:2158971.
Connelly PJ, et al. The importance of gender to understand sex differences in cardiovascular disease. Can J Cardiol. 2021;37(5):699–710.
Regitz-Zagrosek V, Kararigas G. Mechanistic pathways of sex differences in cardiovascular disease. Physiol Rev. 2017;97(1):1–37.
Simoncini T, et al. Novel non-transcriptional mechanisms for estrogen receptor signaling in the cardiovascular system. Steroids. 2002;67(12):935–9. https://doi.org/10.1016/S0039-128X(02)00040-5.
Mueller SO, Korach KS. Estrogen receptors and endocrine diseases: lessons from estrogen receptor knockout mice. Curr Opin Pharmacol. 2001;1(6):613–9.
Couse JF, et al. Tissue distribution and quantitative analysis of estrogen receptor-alpha (ERalpha) and estrogen receptor-beta (ERbeta) messenger ribonucleic acid in the wild-type and ERalpha-knockout mouse. Endocrinology. 1997;138(11):4613–21.
Couse JF, Korach KS. Estrogen receptor null mice: what have we learned and where will they lead us? Endocr Rev. 1999;20(3):358–417.
Guyenet PG. The sympathetic control of blood pressure. Nat Rev Neurosci. 2006;7(5):335–46.
Wang G, et al. Evidence that estrogen directly and indirectly modulates C1 adrenergic bulbospinal neurons in the rostral ventrolateral medulla. Brain Res. 2006;1094(1):163–78.
Subramanian M, et al. Chronic estradiol-17beta exposure increases superoxide production in the rostral ventrolateral medulla and causes hypertension: reversal by resveratrol. Am J Physiol Regul Integr Comp Physiol. 2011;300(6):R1560–8.
Lee SK, Ryu PD, Lee SY. Estrogen replacement modulates voltage-gated potassium channels in rat presympathetic paraventricular nucleus neurons. BMC Neurosci. 2013;14:134.
Saleh TM, Connell BJ. 17beta-estradiol modulates baroreflex sensitivity and autonomic tone of female rats. J Auton Nerv Syst. 2000;80(3):148–61.
Handgraaf S, et al. Prevention of obesity and insulin resistance by estrogens requires ERalpha activation function-2 (ERalphaAF-2), whereas ERalphaAF-1 is dispensable. Diabetes. 2013;62(12):4098–108.
Mauvais-Jarvis F. Estrogen and androgen receptors: regulators of fuel homeostasis and emerging targets for diabetes and obesity. Trends Endocrinol Metab. 2011;22(1):24–33.
Xu Y, O’Malley BW, Elmquist JK. Brain nuclear receptors and body weight regulation. J Clin Invest. 2017;127(4):1172–80.
Xu P, et al. Estrogen receptor-alpha in medial amygdala neurons regulates body weight. J Clin Invest. 2015;125(7):2861–76.
He Y, et al. Estrogen receptor-alpha expressing neurons in the ventrolateral VMH regulate glucose balance. Nat Commun. 2020;11(1):2165.
Weber KT. Aldosterone in congestive heart failure. N Engl J Med. 2001;345(23):1689–97.
Brown NJ. Eplerenone: cardiovascular protection. Circulation. 2003;107(19):2512–8.
Struthers AD. The clinical implications of aldosterone escape in congestive heart failure. Eur J Heart Fail. 2004;6(5):539–45.
Buonafine M, Bonnard B, Jaisser F. Mineralocorticoid receptor and cardiovascular disease. Am J Hypertens. 2018;31(11):1165–74.
Mohammed SF, et al. Mineralocorticoid accelerates transition to heart failure with preserved ejection fraction via “nongenomic effects.” Circulation. 2010;122(4):370–8.
Herrada AA, et al. Aldosterone promotes autoimmune damage by enhancing Th17-mediated immunity. J Immunol. 2010;184(1):191–202.
Fraccarollo D, et al. Immediate mineralocorticoid receptor blockade improves myocardial infarct healing by modulation of the inflammatory response. Hypertension. 2008;51(4):905–14.
Pitt B, et al. Eplerenone, a selective aldosterone blocker, in patients with left ventricular dysfunction after myocardial infarction. N Engl J Med. 2003;348(14):1309–21.
Zannad F, et al. Eplerenone in patients with systolic heart failure and mild symptoms. N Engl J Med. 2011;364(1):11–21.
Viengchareun S, et al. The mineralocorticoid receptor: insights into its molecular and (patho)physiological biology. Nucl Recept Signal. 2007;5: e012.
McEwen BS, Weiss JM, Schwartz LS. Selective retention of corticosterone by limbic structures in rat brain. Nature. 1968;220(5170):911–2.
Young MJ, Clyne CD. Mineralocorticoid receptor actions in cardiovascular development and disease. Essays Biochem. 2021;65(6):901–11.
Gomez-Sanchez EP. Brain mineralocorticoid receptors: orchestrators of hypertension and end-organ disease. Curr Opin Nephrol Hypertens. 2004;13(2):191–6.
Huang BS, Leenen FH. Mineralocorticoid actions in the brain and hypertension. Curr Hypertens Rep. 2011;13(3):214–20.
Cappuccio FP. Cardiovascular and other effects of salt consumption. Kidney Int Suppl (2011). 2013;3(4):312–5.
Kong YW, et al. Sodium and its role in cardiovascular disease—the debate continues. Front Endocrinol (Lausanne). 2016;7:164.
Geerling JC, Kawata M, Loewy AD. Aldosterone-sensitive neurons in the rat central nervous system. J Comp Neurol. 2006;494(3):515–27.
Formenti S, et al. Hindbrain mineralocorticoid mechanisms on sodium appetite. Am J Physiol Regul Integr Comp Physiol. 2013;304(3):R252–9.
Koneru B, et al. Mineralocorticoid receptor in the NTS stimulates saline intake during fourth ventricular infusions of aldosterone. Am J Physiol Regul Integr Comp Physiol. 2014;306(1):R61–6.
Krause EG, Sakai RR. Richter and sodium appetite: from adrenalectomy to molecular biology. Appetite. 2007;49(2):353–67.
Geerling JC, Loewy AD. Aldosterone in the brain. Am J Physiol Renal Physiol. 2009;297(3):F559–76.
Sakai RR, et al. The amygdala: site of genomic and nongenomic arousal of aldosterone-induced sodium intake. Kidney Int. 2000;57(4):1337–45.
Gomberg-Maitland M, Frishman WH. Thyroid hormone and cardiovascular disease. Am Heart J. 1998;135(2 Pt 1):187–96.
Danzi S, Klein I. Thyroid hormone and the cardiovascular system. Minerva Endocrinol. 2004;29(3):139–50.
Klein I, Danzi S. Thyroid disease and the heart. Circulation. 2007;116(15):1725–35.
Vale C, et al. The role of thyroid hormones in heart failure. Cardiovasc Drugs Ther. 2019;33(2):179–88.
Udovcic M, et al. Hypothyroidism and the heart. Methodist Debakey Cardiovasc J. 2017;13(2):55–9.
Gerdes AM, Iervasi G. Thyroid replacement therapy and heart failure. Circulation. 2010;122(4):385–93.
Razvi S. Novel uses of thyroid hormones in cardiovascular conditions. Endocrine. 2019;66(1):115–23.
Razvi S, et al. Thyroid hormones and cardiovascular function and diseases. J Am Coll Cardiol. 2018;71(16):1781–96.
Jabbar A, et al. Thyroid hormones and cardiovascular disease. Nat Rev Cardiol. 2017;14(1):39–55.
Danzi S, Klein I. Thyroid disease and the cardiovascular system. Endocrinol Metab Clin N Am. 2014;43(2):517–28.
Berbel P, Navarro D, Roman GC. An evo-devo approach to thyroid hormones in cerebral and cerebellar cortical development: etiological implications for autism. Front Endocrinol (Lausanne). 2014;5:146.
Bernal J. Thyroid hormone receptors in brain development and function. Nat Clin Pract Endocrinol Metab. 2007;3(3):249–59.
Hahin R. Removal of inactivation causes time-invariant sodium current decays. J Gen Physiol. 1988;92(3):331–50.
Ortiga-Carvalho TM, Sidhaye AR, Wondisford FE. Thyroid hormone receptors and resistance to thyroid hormone disorders. Nat Rev Endocrinol. 2014;10(10):582–91.
Flamant F, Gauthier K, Richard S. Genetic investigation of thyroid hormone receptor function in the developing and adult brain. Curr Top Dev Biol. 2017;125:303–35.
Bradley DJ, Young WS 3rd, Weinberger C. Differential expression of alpha and beta thyroid hormone receptor genes in rat brain and pituitary. Proc Natl Acad Sci U S A. 1989;86(18):7250–4.
Jones I, et al. The thyroid hormone receptor beta gene: structure and functions in the brain and sensory systems. Thyroid. 2003;13(11):1057–68.
Nunez J, et al. Multigenic control of thyroid hormone functions in the nervous system. Mol Cell Endocrinol. 2008;287(1–2):1–12.
Wallis K, et al. The thyroid hormone receptor alpha1 protein is expressed in embryonic postmitotic neurons and persists in most adult neurons. Mol Endocrinol. 2010;24(10):1904–16.
Bradley DJ, Towle HC, Young WS 3rd. Spatial and temporal expression of alpha- and beta-thyroid hormone receptor mRNAs, including the beta 2-subtype, in the developing mammalian nervous system. J Neurosci. 1992;12(6):2288–302.
Kahaly GJ, Dillmann WH. Thyroid hormone action in the heart. Endocr Rev. 2005;26(5):704–28.
Silva JE. Thermogenic mechanisms and their hormonal regulation. Physiol Rev. 2006;86(2):435–64.
Fliers E, Klieverik LP, Kalsbeek A. Novel neural pathways for metabolic effects of thyroid hormone. Trends Endocrinol Metab. 2010;21(4):230–6.
Lopez M, et al. Hypothalamic AMPK and fatty acid metabolism mediate thyroid regulation of energy balance. Nat Med. 2010;16(9):1001–8.
Mittag J, et al. Adaptations of the autonomous nervous system controlling heart rate are impaired by a mutant thyroid hormone receptor-alpha1. Endocrinology. 2010;151(5):2388–95.
Giguere V, et al. Identification of a new class of steroid hormone receptors. Nature. 1988;331(6151):91–4.
Heard DJ, et al. Human ERRgamma, a third member of the estrogen receptor-related receptor (ERR) subfamily of orphan nuclear receptors: tissue-specific isoforms are expressed during development and in the adult. Mol Endocrinol. 2000;14(3):382–92.
Hong H, Yang L, Stallcup MR. Hormone-independent transcriptional activation and coactivator binding by novel orphan nuclear receptor ERR3. J Biol Chem. 1999;274(32):22618–26.
Lehman JJ, et al. Peroxisome proliferator-activated receptor gamma coactivator-1 promotes cardiac mitochondrial biogenesis. J Clin Invest. 2000;106(7):847–56.
Lai L, et al. Transcriptional coactivators PGC-1alpha and PGC-lbeta control overlapping programs required for perinatal maturation of the heart. Genes Dev. 2008;22(14):1948–61.
Lehman JJ, et al. The transcriptional coactivator PGC-1alpha is essential for maximal and efficient cardiac mitochondrial fatty acid oxidation and lipid homeostasis. Am J Physiol Heart Circ Physiol. 2008;295(1):H185–96.
Huss JM, et al. Estrogen-related receptor alpha directs peroxisome proliferator-activated receptor alpha signaling in the transcriptional control of energy metabolism in cardiac and skeletal muscle. Mol Cell Biol. 2004;24(20):9079–91.
Ghanbari F, Mader S, Philip A. Cholesterol as an endogenous ligand of ERRalpha promotes ERRalpha-mediated cellular proliferation and metabolic target gene expression in breast cancer cells. Cells. 2020. https://doi.org/10.3390/cells9081765.
Sakamoto T, et al. A critical role for estrogen-related receptor signaling in cardiac maturation. Circ Res. 2020;126(12):1685–702.
Huss JM, et al. The nuclear receptor ERRalpha is required for the bioenergetic and functional adaptation to cardiac pressure overload. Cell Metab. 2007;6(1):25–37.
Mootha VK, et al. Erralpha and Gabpa/b specify PGC-1alpha-dependent oxidative phosphorylation gene expression that is altered in diabetic muscle. Proc Natl Acad Sci U S A. 2004;101(17):6570–5.
Schreiber SN, et al. The estrogen-related receptor alpha (ERRalpha) functions in PPARgamma coactivator 1alpha (PGC-1alpha)-induced mitochondrial biogenesis. Proc Natl Acad Sci U S A. 2004;101(17):6472–7.
Alaynick WA, et al. ERRgamma directs and maintains the transition to oxidative metabolism in the postnatal heart. Cell Metab. 2007;6(1):13–24.
Cunningham KF, et al. Increased expression of estrogen-related receptor beta during adaptation of adult cardiomyocytes to sustained hypoxia. Am J Cardiovasc Dis. 2016;6(2):46–54.
Huss JM, Garbacz WG, Xie W. Constitutive activities of estrogen-related receptors: transcriptional regulation of metabolism by the ERR pathways in health and disease. Biochim Biophys Acta. 2015;1852(9):1912–27.
Cui H, et al. Behavioral disturbances in estrogen-related receptor alpha-null mice. Cell Rep. 2015;11(3):344–50.
Lorke DE, et al. Differential expression of the estrogen receptor-related receptor gamma in the mouse brain. Brain Res Mol Brain Res. 2000;77(2):277–80.
Byerly MS, et al. Estrogen-related receptor beta deficiency alters body composition and response to restraint stress. BMC Physiol. 2013;13:10.
Kelly DM, Jones TH. Testosterone: a vascular hormone in health and disease. J Endocrinol. 2013;217(3):R47-71.
Ohlsson C, et al. High serum testosterone is associated with reduced risk of cardiovascular events in elderly men. The MrOS (Osteoporotic Fractures in Men) study in Sweden. J Am Coll Cardiol. 2011;58(16):1674–81.
Araujo AB, et al. Clinical review: endogenous testosterone and mortality in men: a systematic review and meta-analysis. J Clin Endocrinol Metab. 2011;96(10):3007–19.
Jankowska EA, et al. Reduction in circulating testosterone relates to exercise capacity in men with chronic heart failure. J Card Fail. 2009;15(5):442–50.
Pugh PJ, Jones TH, Channer KS. Acute haemodynamic effects of testosterone in men with chronic heart failure. Eur Heart J. 2003;24(10):909–15.
Malkin CJ, et al. The effect of testosterone replacement on endogenous inflammatory cytokines and lipid profiles in hypogonadal men. J Clin Endocrinol Metab. 2004;89(7):3313–8.
Davey RA, Grossmann M. Androgen receptor structure, function and biology: from bench to bedside. Clin Biochem Rev. 2016;37(1):3–15.
Ikeda Y, et al. Androgen receptor gene knockout male mice exhibit impaired cardiac growth and exacerbation of angiotensin II-induced cardiac fibrosis. J Biol Chem. 2005;280(33):29661–6.
Bourghardt J, et al. Androgen receptor-dependent and independent atheroprotection by testosterone in male mice. Endocrinology. 2010;151(11):5428–37.
Prins GS, Birch L, Greene GL. Androgen receptor localization in different cell types of the adult rat prostate. Endocrinology. 1991;129(6):3187–99.
Fernandez-Guasti A, Swaab D, Rodriguez-Manzo G. Sexual behavior reduces hypothalamic androgen receptor immunoreactivity. Psychoneuroendocrinology. 2003;28(4):501–12.
Raskin K, et al. Conditional inactivation of androgen receptor gene in the nervous system: effects on male behavioral and neuroendocrine responses. J Neurosci. 2009;29(14):4461–70.
Yu IC, et al. Neuronal androgen receptor regulates insulin sensitivity via suppression of hypothalamic NF-kappaB-mediated PTP1B expression. Diabetes. 2013;62(2):411–23.
Walters KA, et al. The role of central androgen receptor actions in regulating the hypothalamic-pituitary-ovarian axis. Neuroendocrinology. 2018;106(4):389–400.
Herman JP, et al. Regulation of the hypothalamic-pituitary-adrenocortical stress response. Compr Physiol. 2016;6(2):603–21.
Gomez-Sanchez CE, Gomez-Sanchez EP. Editorial: cardiac steroidogenesis—new sites of synthesis, or much ado about nothing? J Clin Endocrinol Metab. 2001;86(11):5118–20.
Lee SR, et al. Non-genomic effect of glucocorticoids on cardiovascular system. Pflugers Arch. 2012;464(6):549–59.
Taves MD, Gomez-Sanchez CE, Soma KK. Extra-adrenal glucocorticoids and mineralocorticoids: evidence for local synthesis, regulation, and function. Am J Physiol Endocrinol Metab. 2011;301(1):E11-24.
Nussinovitch U, et al. Glucocorticoids and the cardiovascular system: state of the art. Curr Pharm Des. 2010;16(32):3574–85.
Liu B, et al. The glucocorticoid receptor in cardiovascular health and disease. Cells. 2019. https://doi.org/10.3390/cells8101227.
Wang Q, et al. Glucocorticoid receptor protein expression in human hippocampus; stability with age. Neurobiol Aging. 2013;34(6):1662–73.
Ghosal S, et al. Glucocorticoid receptors in the nucleus of the solitary tract (NTS) decrease endocrine and behavioral stress responses. Psychoneuroendocrinology. 2014;45:142–53.
Smith SM, Vale WW. The role of the hypothalamic-pituitary-adrenal axis in neuroendocrine responses to stress. Dialogues Clin Neurosci. 2006;8(4):383–95.
Gannon AL, et al. Ablation of glucocorticoid receptor in the hindbrain of the mouse provides a novel model to investigate stress disorders. Sci Rep. 2019;9(1):3250.
Michalik L, Wahli W. Involvement of PPAR nuclear receptors in tissue injury and wound repair. J Clin Invest. 2006;116(3):598–606.
Wang Y, et al. PPARs as metabolic regulators in the liver: lessons from liver-specific PPAR-null mice. Int J Mol Sci. 2020. https://doi.org/10.3390/ijms21062061.
Grygiel-Gorniak B. Peroxisome proliferator-activated receptors and their ligands: nutritional and clinical implications—a review. Nutr J. 2014;13:17.
Hall MG, Quignodon L, Desvergne B. Peroxisome proliferator-activated receptor beta/delta in the brain: facts and hypothesis. PPAR Res. 2008;2008: 780452.
Gofflot F, et al. Systematic gene expression mapping clusters nuclear receptors according to their function in the brain. Cell. 2007;131(2):405–18.
Warden A, et al. Localization of PPAR isotypes in the adult mouse and human brain. Sci Rep. 2016;6:27618.
Moreno S, Farioli-Vecchioli S, Ceru MP. Immunolocalization of peroxisome proliferator-activated receptors and retinoid X receptors in the adult rat CNS. Neuroscience. 2004;123(1):131–45.
Woods JW, et al. Localization of PPARdelta in murine central nervous system: expression in oligodendrocytes and neurons. Brain Res. 2003;975(1–2):10–21.
Sheu SH, et al. Exploring the binding site structure of the PPAR gamma ligand-binding domain by computational solvent mapping. Biochemistry. 2005;44(4):1193–209.
Neschen S, et al. Contrasting effects of fish oil and safflower oil on hepatic peroxisomal and tissue lipid content. Am J Physiol Endocrinol Metab. 2002;282(2):E395-401.
Jain AP, Aggarwal KK, Zhang PY. Omega-3 fatty acids and cardiovascular disease. Eur Rev Med Pharmacol Sci. 2015;19(3):441–5.
Palomer X, et al. PPARbeta/delta and lipid metabolism in the heart. Biochim Biophys Acta. 2016;1861(10):1569–78.
Ngala RA, et al. A new, highly selective murine peroxisome proliferator-activated receptor delta agonist increases responsiveness to thermogenic stimuli and glucose uptake in skeletal muscle in obese mice. Diabetes Obes Metab. 2011;13(5):455–64.
Yu BC, et al. Decrease of peroxisome proliferator-activated receptor delta expression in cardiomyopathy of streptozotocin-induced diabetic rats. Cardiovasc Res. 2008;80(1):78–87.
Burkart EM, et al. Nuclear receptors PPARbeta/delta and PPARalpha direct distinct metabolic regulatory programs in the mouse heart. J Clin Invest. 2007;117(12):3930–9.
Chen YE, et al. Peroxisome proliferator-activated receptors and the cardiovascular system. Vitam Horm. 2003;66:157–88.
Marx N, et al. Peroxisome proliferator-activated receptors and atherogenesis: regulators of gene expression in vascular cells. Circ Res. 2004;94(9):1168–78.
Touyz RM, Schiffrin EL. Peroxisome proliferator-activated receptors in vascular biology-molecular mechanisms and clinical implications. Vascul Pharmacol. 2006;45(1):19–28.
Tontonoz P, Spiegelman BM. Fat and beyond: the diverse biology of PPARgamma. Annu Rev Biochem. 2008;77:289–312.
Diano S, et al. Peroxisome proliferation-associated control of reactive oxygen species sets melanocortin tone and feeding in diet-induced obesity. Nat Med. 2011;17(9):1121–7.
Sarruf DA, et al. Expression of peroxisome proliferator-activated receptor-gamma in key neuronal subsets regulating glucose metabolism and energy homeostasis. Endocrinology. 2009;150(2):707–12.
Lu M, et al. Brain PPAR-gamma promotes obesity and is required for the insulin-sensitizing effect of thiazolidinediones. Nat Med. 2011;17(5):618–22.
Thomas P, Pang Y. Protective actions of progesterone in the cardiovascular system: potential role of membrane progesterone receptors (mPRs) in mediating rapid effects. Steroids. 2013;78(6):583–8.
Karas RH, et al. A complex role for the progesterone receptor in the response to vascular injury. J Clin Invest. 2001;108(4):611–8.
Lee WS, et al. Progesterone inhibits arterial smooth muscle cell proliferation. Nat Med. 1997;3(9):1005–8.
Lin AL, McGill HC, Shain SA. Hormone receptors of the baboon cardiovascular system. Biochemical characterization of aortic and myocardial cytoplasmic progesterone receptors. Circ Res. 1982;50(5):610–6. https://doi.org/10.1161/01.RES.50.5.610.
Zhu Y, Bond J, Thomas P. Identification, classification, and partial characterization of genes in humans and other vertebrates homologous to a fish membrane progestin receptor. Proc Natl Acad Sci U S A. 2003;100(5):2237–42.
Kasubuchi M, et al. Membrane progesterone receptor beta (mPRbeta/Paqr8) promotes progesterone-dependent neurite outgrowth in PC12 neuronal cells via non-G protein-coupled receptor (GPCR) signaling. Sci Rep. 2017;7(1):5168.
Nutu M, et al. Distribution and hormonal regulation of membrane progesterone receptors beta and gamma in ciliated epithelial cells of mouse and human fallopian tubes. Reprod Biol Endocrinol. 2009;7:89.
Cai Z, Stocco C. Expression and regulation of progestin membrane receptors in the rat corpus luteum. Endocrinology. 2005;146(12):5522–32.
He W, et al. Hypothalamic effects of progesterone on regulation of the pulsatile and surge release of luteinising hormone in female rats. Sci Rep. 2017;7(1):8096.
Gal A, et al. Loss of fertility in the absence of progesterone receptor expression in Kisspeptin neurons of female mice. PLoS ONE. 2016;11(7): e0159534.
Stephens SB, et al. Absent progesterone signaling in kisspeptin neurons disrupts the LH surge and impairs fertility in female mice. Endocrinology. 2015;156(9):3091–7.
Masters M, Riley PR. The epicardium signals the way towards heart regeneration. Stem Cell Res. 2014;13(3 Pt B):683–92.
Stefanovic S, Zaffran S. Mechanisms of retinoic acid signaling during cardiogenesis. Mech Dev. 2017;143:9–19.
Wang S, et al. Alterations in retinoic acid signaling affect the development of the mouse coronary vasculature. Dev Dyn. 2018;247(8):976–91.
Silva RAC, et al. Cardiac remodeling induced by all-trans retinoic acid is detrimental in normal rats. Cell Physiol Biochem. 2017;43(4):1449–59.
Allenby G, et al. Retinoic acid receptors and retinoid X receptors: interactions with endogenous retinoic acids. Proc Natl Acad Sci U S A. 1993;90(1):30–4.
Mangelsdorf DJ, et al. Characterization of three RXR genes that mediate the action of 9-cis retinoic acid. Genes Dev. 1992;6(3):329–44.
Guleria RS, et al. Retinoic acid receptor-mediated signaling protects cardiomyocytes from hyperglycemia induced apoptosis: role of the renin-angiotensin system. J Cell Physiol. 2011;226(5):1292–307.
Marino A, et al. A retinoic acid beta2-receptor agonist exerts cardioprotective effects. J Pharmacol Exp Ther. 2018;366(2):314–21.
Tang XH, et al. A retinoic acid receptor beta 2 agonist improves cardiac function in a heart failure model. J Pharmacol Exp Ther. 2021;379(2):182–90.
Watanabe, M. and H. Kakuta, Retinoid X Receptor Antagonists. Int J Mol Sci, 2018. 19(8).
Mascrez B, et al. A transcriptionally silent RXRalpha supports early embryonic morphogenesis and heart development. Proc Natl Acad Sci U S A. 2009;106(11):4272–7.
Shao M, et al. The multi-faceted role of retinoid X receptor in cardiovascular diseases. Biomed Pharmacother. 2021;137: 111264.
Wang S, Moise AR. Recent insights on the role and regulation of retinoic acid signaling during epicardial development. Genesis. 2019;57(7–8): e23303.
Mendelsohn C, et al. Function of the retinoic acid receptors (RARs) during development (II). Multiple abnormalities at various stages of organogenesis in RAR double mutants. Development. 1994;120(10):2749–71.
Colbert MC, et al. Cardiac compartment-specific overexpression of a modified retinoic acid receptor produces dilated cardiomyopathy and congestive heart failure in transgenic mice. J Clin Invest. 1997;100(8):1958–68.
Wood H. Retinoid X receptor mediates brain clean-up after stroke. Nat Rev Neurol. 2020;16(3):128–9.
Ting SM, et al. Brain cleanup as a potential target for poststroke recovery: the role of RXR (retinoic X receptor) in phagocytes. Stroke. 2020;51(3):958–66.
Zuo Y, et al. Activation of retinoid X receptor by bexarotene attenuates neuroinflammation via PPARgamma/SIRT6/FoxO3a pathway after subarachnoid hemorrhage in rats. J Neuroinflamm. 2019;16(1):47.
Jetten AM, et al. Retinoic acid-related Orphan Receptor gamma (RORgamma): connecting sterol metabolism to regulation of the immune system and autoimmune disease. Curr Opin Toxicol. 2018;8:66–80.
Caballero B, et al. Favorable effects of a prolonged treatment with melatonin on the level of oxidative damage and neurodegeneration in senescence-accelerated mice. J Pineal Res. 2008;45(3):302–11.
Kim EJ, et al. Transcriptional activation of HIF-1 by RORalpha and its role in hypoxia signaling. Arterioscler Thromb Vasc Biol. 2008;28(10):1796–802.
Kojetin DJ, Burris TP. REV-ERB and ROR nuclear receptors as drug targets. Nat Rev Drug Discov. 2014;13(3):197–216.
Meng H, et al. High retinoic acid receptor-related orphan receptor A gene expression in peripheral blood leukocytes may be related to acute myocardial infarction. J Int Med Res. 2021;49(6):3000605211019663.
Matsuoka H, et al. The retinoic acid receptor-related orphan receptor alpha positively regulates tight junction protein claudin domain-containing 1 mRNA expression in human brain endothelial cells. J Biochem. 2017;161(5):441–50.
Singh MV, et al. Abnormal CD161(+) immune cells and retinoic acid receptor-related orphan receptor gammat-mediate enhanced IL-17F expression in the setting of genetic hypertension. J Allergy Clin Immunol. 2017;140(3):809-821 e3.
Zhao Y, et al. Novel protective role of the circadian nuclear receptor retinoic acid-related orphan receptor-alpha in diabetic cardiomyopathy. J Pineal Res. 2017. https://doi.org/10.1111/jpi.12378.
Hamilton BA, et al. Disruption of the nuclear hormone receptor RORalpha in staggerer mice. Nature. 1996;379(6567):736–9.
Steinmayr M, et al. staggerer phenotype in retinoid-related orphan receptor alpha-deficient mice. Proc Natl Acad Sci U S A. 1998;95(7):3960–5.
Aubert ML, et al. Metabolic control of sexual function and growth: role of neuropeptide Y and leptin. Mol Cell Endocrinol. 1998;140(1–2):107–13.
Fan J, et al. Retinoic acid receptor-related orphan receptors: critical roles in tumorigenesis. Front Immunol. 2018;9:1187.
Lee JM, Kim H, Baek SH. Unraveling the physiological roles of retinoic acid receptor-related orphan receptor alpha. Exp Mol Med. 2021;53(9):1278–86.
Sato TK, et al. A functional genomics strategy reveals Rora as a component of the mammalian circadian clock. Neuron. 2004;43(4):527–37.
Scheer FA, et al. Adverse metabolic and cardiovascular consequences of circadian misalignment. Proc Natl Acad Sci U S A. 2009;106(11):4453–8.
Latic N, Erben RG. Vitamin D and cardiovascular disease, with emphasis on hypertension, atherosclerosis, and heart failure. Int J Mol Sci. 2020. https://doi.org/10.3390/ijms21186483.
Scragg R. Seasonality of cardiovascular disease mortality and the possible protective effect of ultra-violet radiation. Int J Epidemiol. 1981;10(4):337–41.
Al Mheid I, et al. Vitamin D status is associated with arterial stiffness and vascular dysfunction in healthy humans. J Am Coll Cardiol. 2011;58(2):186–92.
Lee JH, et al. Prevalence of vitamin D deficiency in patients with acute myocardial infarction. Am J Cardiol. 2011;107(11):1636–8.
London GM, et al. Mineral metabolism and arterial functions in end-stage renal disease: potential role of 25-hydroxyvitamin D deficiency. J Am Soc Nephrol. 2007;18(2):613–20.
Avenell A, et al. Long-term follow-up for mortality and cancer in a randomized placebo-controlled trial of vitamin D(3) and/or calcium (RECORD trial). J Clin Endocrinol Metab. 2012;97(2):614–22.
Schleithoff SS, et al. Vitamin D supplementation improves cytokine profiles in patients with congestive heart failure: a double-blind, randomized, placebo-controlled trial. Am J Clin Nutr. 2006;83(4):754–9.
Ma Y, et al. Identification and characterization of noncalcemic, tissue-selective, nonsecosteroidal vitamin D receptor modulators. J Clin Invest. 2006;116(4):892–904.
Eyles DW, et al. Distribution of the vitamin D receptor and 1 alpha-hydroxylase in human brain. J Chem Neuroanat. 2005;29(1):21–30.
Prufer K, et al. Distribution of 1,25-dihydroxyvitamin D3 receptor immunoreactivity in the rat brain and spinal cord. J Chem Neuroanat. 1999;16(2):135–45.
Cui X, et al. The vitamin D receptor in dopamine neurons; its presence in human substantia nigra and its ontogenesis in rat midbrain. Neuroscience. 2013;236:77–87.
Lee PW, et al. Neuron-specific vitamin D signaling attenuates microglia activation and CNS autoimmunity. Front Neurol. 2020;11:19.
Smolders J, et al. Expression of vitamin D receptor and metabolizing enzymes in multiple sclerosis-affected brain tissue. J Neuropathol Exp Neurol. 2013;72(2):91–105.
Bass J, Lazar MA. Circadian time signatures of fitness and disease. Science. 2016;354(6315):994–9.
Woldt E, et al. Rev-erb-alpha modulates skeletal muscle oxidative capacity by regulating mitochondrial biogenesis and autophagy. Nat Med. 2013;19(8):1039–46.
Cho H, et al. Regulation of circadian behaviour and metabolism by REV-ERB-alpha and REV-ERB-beta. Nature. 2012;485(7396):123–7.
Burke L, et al. Transcriptional repression by the orphan steroid receptor RVR/Rev-erb beta is dependent on the signature motif and helix 5 in the E region: functional evidence for a biological role of RVR in myogenesis. Nucleic Acids Res. 1996;24(18):3481–9.
Preitner N, et al. The orphan nuclear receptor REV-ERBalpha controls circadian transcription within the positive limb of the mammalian circadian oscillator. Cell. 2002;110(2):251–60.
Yin L, et al. Rev-erbalpha, a heme sensor that coordinates metabolic and circadian pathways. Science. 2007;318(5857):1786–9.
Solt LA, et al. Regulation of circadian behaviour and metabolism by synthetic REV-ERB agonists. Nature. 2012;485(7396):62–8.
Bugge A, et al. Rev-erbalpha and Rev-erbbeta coordinately protect the circadian clock and normal metabolic function. Genes Dev. 2012;26(7):657–67.
Gerhart-Hines Z, et al. The nuclear receptor Rev-erbalpha controls circadian thermogenic plasticity. Nature. 2013;503(7476):410–3.
Song S, et al. Myocardial Rev-erb-Mediated diurnal metabolic rhythm and obesity paradox. Circulation. 2022;145(6):448–64.
Sitaula S, et al. Suppression of atherosclerosis by synthetic REV-ERB agonist. Biochem Biophys Res Commun. 2015;460(3):566–71.
Stujanna EN, et al. Rev-erb agonist improves adverse cardiac remodeling and survival in myocardial infarction through an anti-inflammatory mechanism. PLoS ONE. 2017;12(12): e0189330.
Zhang T, et al. Dysregulation of REV-ERBalpha impairs GABAergic function and promotes epileptic seizures in preclinical models. Nat Commun. 2021;12(1):1216.
Jager J, et al. Behavioral changes and dopaminergic dysregulation in mice lacking the nuclear receptor Rev-erbalpha. Mol Endocrinol. 2014;28(4):490–8.
Chung S, et al. Impact of circadian nuclear receptor REV-ERBalpha on midbrain dopamine production and mood regulation. Cell. 2014;157(4):858–68.
Adlanmerini M, et al. Hypothalamic REV-ERB nuclear receptors control diurnal food intake and leptin sensitivity in diet-induced obese mice. J Clin Invest. 2021. https://doi.org/10.1172/JCI140424.
Claudel T, Staels B, Kuipers F. The Farnesoid X receptor: a molecular link between bile acid and lipid and glucose metabolism. Arterioscler Thromb Vasc Biol. 2005;25(10):2020–30.
Pu J, et al. Cardiomyocyte-expressed farnesoid-X-receptor is a novel apoptosis mediator and contributes to myocardial ischaemia/reperfusion injury. Eur Heart J. 2013;34(24):1834–45.
Huang C, et al. Identification of functional farnesoid X receptors in brain neurons. FEBS Lett. 2016;590(18):3233–42.
McMillin M, et al. Bile acid signaling is involved in the neurological decline in a murine model of acute liver failure. Am J Pathol. 2016;186(2):312–23.
Bazzari FH, Abdallah DM, El-Abhar HS. Chenodeoxycholic acid ameliorates AlCl3-induced Alzheimer’s disease neurotoxicity and cognitive deterioration via enhanced insulin signaling in rats. Molecules. 2019. https://doi.org/10.3390/molecules24101992.
Higashi T, et al. Unconjugated bile acids in rat brain: analytical method based on LC/ESI-MS/MS with chemical derivatization and estimation of their origin by comparison to serum levels. Steroids. 2017;125:107–13.
Chen WG, et al. Hippocampal FXR plays a role in the pathogenesis of depression: a preliminary study based on lentiviral gene modulation. Psychiatry Res. 2018;264:374–9.
Chen Q, et al. Farnesoid X receptor (FXR) aggravates amyloid-beta-triggered apoptosis by modulating the cAMP-response element-binding protein (CREB)/brain-derived neurotrophic factor (BDNF) pathway in vitro. Med Sci Monit. 2019;25:9335–45.
Zhang SY, et al. FXR in the dorsal vagal complex is sufficient and necessary for upper small intestinal microbiome-mediated changes of TCDCA to alter insulin action in rats. Gut. 2021;70(9):1675–83.
Yang Y, Xu Y. The central melanocortin system and human obesity. J Mol Cell Biol. 2020;12(10):785–97.
Kouidhi S, Clerget-Froidevaux MS. Integrating thyroid hormone signaling in hypothalamic control of metabolism: crosstalk between nuclear receptors. Int J Mol Sci. 2018. https://doi.org/10.3390/ijms19072017.
Courtney R, Landreth GE. LXR regulation of brain cholesterol: from development to disease. Trends Endocrinol Metab. 2016;27(6):404–14.
Peet DJ, Janowski BA, Mangelsdorf DJ. The LXRs: a new class of oxysterol receptors. Curr Opin Genet Dev. 1998;8(5):571–5.
Cannon MV, van Gilst WH, de Boer RA. Emerging role of liver X receptors in cardiac pathophysiology and heart failure. Basic Res Cardiol. 2016;111(1):3.
Naik SU, et al. Pharmacological activation of liver X receptors promotes reverse cholesterol transport in vivo. Circulation. 2006;113(1):90–7.
Joseph SB, et al. Synthetic LXR ligand inhibits the development of atherosclerosis in mice. Proc Natl Acad Sci U S A. 2002;99(11):7604–9.
Tan XJ, et al. Anxiety in liver X receptor beta knockout female mice with loss of glutamic acid decarboxylase in ventromedial prefrontal cortex. Proc Natl Acad Sci U S A. 2012;109(19):7493–8.
Ben Aissa M, et al. Discovery of nonlipogenic ABCA1 inducing compounds with potential in Alzheimer’s disease and Type 2 diabetes. ACS Pharmacol Transl Sci. 2021;4(1):143–54.
Kruse MS, Suarez LG, Coirini H. Regulation of the expression of LXR in rat hypothalamic and hippocampal explants. Neurosci Lett. 2017;639:53–8.
Miao Y, et al. Liver X receptor beta controls thyroid hormone feedback in the brain and regulates browning of subcutaneous white adipose tissue. Proc Natl Acad Sci U S A. 2015;112(45):14006–11.
Ghaddab-Zroud R, et al. Liver X receptor regulation of thyrotropin-releasing hormone transcription in mouse hypothalamus is dependent on thyroid status. PLoS ONE. 2014;9(9): e106983.
Sandoval-Hernandez AG, et al. LXR activation protects hippocampal microvasculature in very old triple transgenic mouse model of Alzheimer’s disease. Neurosci Lett. 2016;621:15–21.
Morales JR, et al. Activation of liver X receptors promotes neuroprotection and reduces brain inflammation in experimental stroke. Circulation. 2008;118(14):1450–9.
Sironi L, et al. Treatment with LXR agonists after focal cerebral ischemia prevents brain damage. FEBS Lett. 2008;582(23–24):3396–400.
ElAli A, Hermann DM. Liver X receptor activation enhances blood-brain barrier integrity in the ischemic brain and increases the abundance of ATP-binding cassette transporters ABCB1 and ABCC1 on brain capillary cells. Brain Pathol. 2012;22(2):175–87.
Seol W, Choi HS, Moore DD. An orphan nuclear hormone receptor that lacks a DNA binding domain and heterodimerizes with other receptors. Science. 1996;272(5266):1336–9.
Lee YS, et al. Structure and function of the atypical orphan nuclear receptor small heterodimer partner. Int Rev Cytol. 2007;261:117–58.
Rodriguez-Calvo R, et al. Small heterodimer partner (SHP) contributes to insulin resistance in cardiomyocytes. Biochim Biophys Acta Mol Cell Biol Lipids. 2017;1862(5):541–51.
Nam YS, et al. Small heterodimer partner blocks cardiac hypertrophy by interfering with GATA6 signaling. Circ Res. 2014;115(5):493–503.
Chiang JY. Bile acid regulation of gene expression: roles of nuclear hormone receptors. Endocr Rev. 2002;23(4):443–63.
Kim JY, et al. Orphan nuclear receptor small heterodimer partner, a novel corepressor for a basic helix-loop-helix transcription factor BETA2/neuroD. Mol Endocrinol. 2004;18(4):776–90.
Seol W, et al. Inhibition of estrogen receptor action by the orphan receptor SHP (short heterodimer partner). Mol Endocrinol. 1998;12(10):1551–7.
Wang L, Liangpunsakul S. Circadian clock control of hepatic lipid metabolism: role of small heterodimer partner (Shp). J Investig Med. 2016;64(7):1158–61.
Lee JH, et al. Small heterodimer partner SHP mediates liver X receptor (LXR)-dependent suppression of inflammatory signaling by promoting LXR SUMOylation specifically in astrocytes. Sci Signal. 2016;9(439):ra78.
Locker J, et al. A common set of immediate-early response genes in liver regeneration and hyperplasia. Hepatology. 2003;38(2):314–25.
Reising K, et al. Biomechanical testing of an innovative fixation procedure to stabilize olecranon osteotomy. Proc Inst Mech Eng H. 2014;228(11):1146–53.
Columbano A, et al. Gadd45beta is induced through a CAR-dependent, TNF-independent pathway in murine liver hyperplasia. Hepatology. 2005;42(5):1118–26.
Gao J, et al. The constitutive androstane receptor is an anti-obesity nuclear receptor that improves insulin sensitivity. J Biol Chem. 2009;284(38):25984–92.
Dong B, et al. Activation of nuclear receptor CAR ameliorates diabetes and fatty liver disease. Proc Natl Acad Sci U S A. 2009;106(44):18831–6.
Abe H, Semba H, Takeda N. The roles of hypoxia signaling in the pathogenesis of cardiovascular diseases. J Atheroscler Thromb. 2017;24(9):884–94.
Lamba JK, et al. Expression of constitutive androstane receptor splice variants in human tissues and their functional consequences. J Pharmacol Exp Ther. 2004;311(2):811–21.
Dutheil F, et al. Xenobiotic-metabolizing enzymes and transporters in the normal human brain: regional and cellular mapping as a basis for putative roles in cerebral function. Drug Metab Dispos. 2009;37(7):1528–38.
Kajta M, et al. Triclocarban disrupts the epigenetic status of neuronal cells and induces AHR/CAR-mediated apoptosis. Mol Neurobiol. 2019;56(5):3113–31.
Litwa E, et al. RXRalpha, PXR and CAR xenobiotic receptors mediate the apoptotic and neurotoxic actions of nonylphenol in mouse hippocampal cells. J Steroid Biochem Mol Biol. 2016;156:43–52.
Parker KL, Schimmer BP. Steroidogenic factor 1: a key determinant of endocrine development and function. Endocr Rev. 1997;18(3):361–77.
Ikeda Y, et al. The nuclear receptor steroidogenic factor 1 is essential for the formation of the ventromedial hypothalamic nucleus. Mol Endocrinol. 1995;9(4):478–86.
Dhillon H, et al. Leptin directly activates SF1 neurons in the VMH, and this action by leptin is required for normal body-weight homeostasis. Neuron. 2006;49(2):191–203.
Dellovade TL, et al. Disruption of the gene encoding SF-1 alters the distribution of hypothalamic neuronal phenotypes. J Comp Neurol. 2000;423(4):579–89.
Budefeld T, Tobet SA, Majdic G. Altered position of cell bodies and fibers in the ventromedial region in SF-1 knockout mice. Exp Neurol. 2011;232(2):176–84.
Valensi P, et al. Cardiovascular vagosympathetic activity in rats with ventromedial hypothalamic obesity. Obes Res. 2003;11(1):54–64.
Gauthier P, Reis DJ, Nathan MA. Arterial hypertension elicited either by lesions or by electrical stimulations of the rostral hypothalamus in the rat. Brain Res. 1981;211(1):91–105.
King BM. The rise, fall, and resurrection of the ventromedial hypothalamus in the regulation of feeding behavior and body weight. Physiol Behav. 2006;87(2):221–44.
Zhou Y, et al. Ventromedial hypothalamus activation aggravates hypertension myocardial remodeling through the sympathetic nervous system. Front Cardiovasc Med. 2021;8: 737135.
Kivimaki M, Steptoe A. Effects of stress on the development and progression of cardiovascular disease. Nat Rev Cardiol. 2018;15(4):215–29.
Labbe SM, et al. Hypothalamic control of brown adipose tissue thermogenesis. Front Syst Neurosci. 2015;9:150.
Martinez-Sanchez N, et al. Hypothalamic AMPK-ER stress-JNK1 axis mediates the central actions of thyroid hormones on energy balance. Cell Metab. 2017;26(1):212-229 e12.
Xu Y, et al. PI3K signaling in the ventromedial hypothalamic nucleus is required for normal energy homeostasis. Cell Metab. 2010;12(1):88–95.
Xu Y, et al. Distinct hypothalamic neurons mediate estrogenic effects on energy homeostasis and reproduction. Cell Metab. 2011;14(4):453–65.
Seoane-Collazo P, et al. SF1-specific AMPKalpha1 deletion protects against diet-induced obesity. Diabetes. 2018;67(11):2213–26.
Castillo-Armengol J, et al. Hypothalamic CDK4 regulates thermogenesis by modulating sympathetic innervation of adipose tissues. EMBO Rep. 2020;21(9): e49807.
Rouabhi M, et al. BBSome ablation in SF1 neurons causes obesity without comorbidities. Mol Metab. 2021;48: 101211.
Lim K, et al. Origin of aberrant blood pressure and sympathetic regulation in diet-induced obesity. Hypertension. 2016;68(2):491–500.
Mendelson CR, et al. Transcriptional regulation of aromatase in placenta and ovary. J Steroid Biochem Mol Biol. 2005;95(1–5):25–33.
Higashiyama H, Kinoshita M, Asano S. Expression profiling of liver receptor homologue 1 (LRH-1) in mouse tissues using tissue microarray. J Mol Histol. 2007;38(1):45–52.
Nadolny C, Dong X. Liver receptor homolog-1 (LRH-1): a potential therapeutic target for cancer. Cancer Biol Ther. 2015;16(7):997–1004.
Fayard E, Auwerx J, Schoonjans K. LRH-1: an orphan nuclear receptor involved in development, metabolism and steroidogenesis. Trends Cell Biol. 2004;14(5):250–60.
Lee JM, et al. A nuclear-receptor-dependent phosphatidylcholine pathway with antidiabetic effects. Nature. 2011;474(7352):506–10.
Oosterveer MH, et al. LRH-1-dependent glucose sensing determines intermediary metabolism in liver. J Clin Invest. 2012;122(8):2817–26.
Miranda DA, et al. LRH-1 regulates hepatic lipid homeostasis and maintains arachidonoyl phospholipid pools critical for phospholipid diversity. JCI Insight. 2018. https://doi.org/10.1172/jci.insight.96151.
Hattori T, et al. LRH-1 heterozygous knockout mice are prone to mild obesity. Endocr J. 2014;61(5):471–80.
Delerive P, et al. Identification of liver receptor homolog-1 as a novel regulator of apolipoprotein AI gene transcription. Mol Endocrinol. 2004;18(10):2378–87.
Lu M, et al. ApoB/apoA1 is an effective predictor of coronary heart disease risk in overweight and obesity. J Biomed Res. 2011;25(4):266–73.
Rahim S, et al. Serum Apo A-1 and its role as a biomarker of coronary artery disease. Cureus. 2016;8(12): e941.
Atkin SD, et al. Nuclear receptor LRH-1 induces the reproductive neuropeptide kisspeptin in the hypothalamus. Mol Endocrinol. 2013;27(4):598–605.
Lee JH, et al. KiSS-1, a novel human malignant melanoma metastasis-suppressor gene. J Natl Cancer Inst. 1996;88(23):1731–7.
Zhang Y, et al. The effects of kisspeptin-10 on serum metabolism and myocardium in rats. PLoS ONE. 2017;12(7): e0179164.
Pearen MA, Muscat GE. Minireview: nuclear hormone receptor 4A signaling: implications for metabolic disease. Mol Endocrinol. 2010;24(10):1891–903.
Ranhotra HS. The NR4A orphan nuclear receptors: mediators in metabolism and diseases. J Recept Signal Transduct Res. 2015;35(2):184–8.
Kurakula K, et al. NR4A nuclear receptors are orphans but not lonesome. Biochim Biophys Acta. 2014;1843(11):2543–55.
Hawk JD, et al. NR4A nuclear receptors support memory enhancement by histone deacetylase inhibitors. J Clin Invest. 2012;122(10):3593–602.
Chen Y, et al. Enhancement of hypothalamic STAT3 acetylation by nuclear receptor Nur77 dictates leptin sensitivity. Diabetes. 2015;64(6):2069–81.
Nonogaki K, et al. Serotonin 5-HT2C receptor-independent expression of hypothalamic NOR1, a novel modulator of food intake and energy balance, in mice. Biochem Biophys Res Commun. 2009;386(2):311–5.
Gilbert F, et al. Nur77 gene knockout alters dopamine neuron biochemical activity and dopamine turnover. Biol Psychiatry. 2006;60(6):538–47.
Zetterstrom RH, et al. Dopamine neuron agenesis in Nurr1-deficient mice. Science. 1997;276(5310):248–50.
Saucedo-Cardenas O, et al. Nurr1 is essential for the induction of the dopaminergic phenotype and the survival of ventral mesencephalic late dopaminergic precursor neurons. Proc Natl Acad Sci U S A. 1998;95(7):4013–8.
Montarolo F, et al. NURR1-deficient mice have age- and sex-specific behavioral phenotypes. J Neurosci Res. 2022. https://doi.org/10.1002/jnr.25067.
Chung AC, et al. Loss of orphan receptor germ cell nuclear factor function results in ectopic development of the tail bud and a novel posterior truncation. Mol Cell Biol. 2001;21(2):663–77.
Fuhrmann G, et al. Mouse germline restriction of Oct4 expression by germ cell nuclear factor. Dev Cell. 2001;1(3):377–87.
Lan ZJ, et al. Expression of the orphan nuclear receptor, germ cell nuclear factor, in mouse gonads and preimplantation embryos. Biol Reprod. 2003;68(1):282–9.
Lan ZJ, et al. GCNF-dependent repression of BMP-15 and GDF-9 mediates gamete regulation of female fertility. EMBO J. 2003;22(16):4070–81.
Bizkarguenaga M, et al. Increase of germ cell nuclear factor expression in globozoospermic Gopc(-/-) knockout mice. Andrology. 2019;7(3):319–28.
Gu P, et al. Orphan nuclear receptor GCNF is required for the repression of pluripotency genes during retinoic acid-induced embryonic stem cell differentiation. Mol Cell Biol. 2005;25(19):8507–19.
Baker JL, et al. Testicular receptor 2, Nr2c1, is associated with stem cells in the developing olfactory epithelium and other cranial sensory and skeletal structures. Gene Expr Patterns. 2016;20(1):71–9.
Chen YT, et al. The roles of testicular orphan nuclear receptor 4 (TR4) in cerebellar development. Cerebellum. 2008;7(1):9–17.
Chen YT, et al. Abnormal cerebellar cytoarchitecture and impaired inhibitory signaling in adult mice lacking TR4 orphan nuclear receptor. Brain Res. 2007;1168:72–82.
Chen LM, et al. Subfertility with defective folliculogenesis in female mice lacking testicular orphan nuclear receptor 4. Mol Endocrinol. 2008;22(4):858–67.
Collins LL, et al. Growth retardation and abnormal maternal behavior in mice lacking testicular orphan nuclear receptor 4. Proc Natl Acad Sci U S A. 2004;101(42):15058–63.
Lin SJ, et al. TR4 nuclear receptor functions as a tumor suppressor for prostate tumorigenesis via modulation of DNA damage/repair system. Carcinogenesis. 2014;35(6):1399–406.
Chen B, et al. The role of testicular nuclear receptor 4 in chemo-resistance of docetaxel in castration-resistant prostate cancer. Cancer Gene Ther. 2014;21(10):411–5.
Kliewer SA, Goodwin B, Willson TM. The nuclear pregnane X receptor: a key regulator of xenobiotic metabolism. Endocr Rev. 2002;23(5):687–702.
Ding X, Staudinger JL. Induction of drug metabolism by forskolin: the role of the pregnane X receptor and the protein kinase a signal transduction pathway. J Pharmacol Exp Ther. 2005;312(2):849–56.
Ricketts ML, et al. The cholesterol-raising factor from coffee beans, cafestol, as an agonist ligand for the farnesoid and pregnane X receptors. Mol Endocrinol. 2007;21(7):1603–16.
Sui Y, et al. Bisphenol A increases atherosclerosis in pregnane X receptor-humanized ApoE deficient mice. J Am Heart Assoc. 2014;3(2): e000492.
Lin FJ, et al. Coup d’Etat: an orphan takes control. Endocr Rev. 2011;32(3):404–21.
Chen CA, et al. Nr2f1 heterozygous knockout mice recapitulate neurological phenotypes of Bosch-Boonstra-Schaaf optic atrophy syndrome and show impaired hippocampal synaptic plasticity. Hum Mol Genet. 2020;29(5):705–15.
Wu SP, et al. Increased COUP-TFII expression in adult hearts induces mitochondrial dysfunction resulting in heart failure. Nat Commun. 2015;6:8245.
Reinchisi G, et al. COUP-TFII expressing interneurons in human fetal forebrain. Cereb Cortex. 2012;22(12):2820–30.
Alzu’bi A, et al. The transcription factors COUP-TFI and COUP-TFII have distinct roles in arealisation and GABAergic interneuron specification in the early human fetal telencephalon. Cereb Cortex. 2017;27(10):4971–87.
Kanatani S, et al. The COUP-TFII/Neuropilin-2 is a molecular switch steering diencephalon-derived GABAergic neurons in the developing mouse brain. Proc Natl Acad Sci U S A. 2015;112(36):E4985–94.
Yu RN, et al. The role of DAX-1 in reproduction. Trends Endocrinol Metab. 1998;9(5):169–75.
Lalli E, Sassone-Corsi P. DAX-1, an unusual orphan receptor at the crossroads of steroidogenic function and sexual differentiation. Mol Endocrinol. 2003;17(8):1445–53.
Suzuki T, et al. LXXLL motifs in Dax-1 have target specificity for the orphan nuclear receptors Ad4BP/SF-1 and LRH-1. Endocr Res. 2002;28(4):537.
Ito M, Yu R, Jameson JL. DAX-1 inhibits SF-1-mediated transactivation via a carboxy-terminal domain that is deleted in adrenal hypoplasia congenita. Mol Cell Biol. 1997;17(3):1476–83.
Zhang H, et al. DAX-1 functions as an LXXLL-containing corepressor for activated estrogen receptors. J Biol Chem. 2000;275(51):39855–9.
Agoulnik IU, et al. Repressors of androgen and progesterone receptor action. J Biol Chem. 2003;278(33):31136–48.
Yu RN, et al. Role of Ahch in gonadal development and gametogenesis. Nat Genet. 1998;20(4):353–7.
Tabarin A, et al. A novel mutation in DAX1 causes delayed-onset adrenal insufficiency and incomplete hypogonadotropic hypogonadism. J Clin Invest. 2000;105(3):321–8.
Omu AE. Sperm parameters: paradigmatic index of good health and longevity. Med Princ Pract. 2013;22(Suppl 1):30–42.
Holter E, et al. Inhibition of androgen receptor (AR) function by the reproductive orphan nuclear receptor DAX-1. Mol Endocrinol. 2002;16(3):515–28.
Yu RT, et al. Relationship between Drosophila gap gene tailless and a vertebrate nuclear receptor Tlx. Nature. 1994;370(6488):375–9.
Zhang CL, et al. A role for adult TLX-positive neural stem cells in learning and behaviour. Nature. 2008;451(7181):1004–7.
Murai K, et al. Nuclear receptor TLX stimulates hippocampal neurogenesis and enhances learning and memory in a transgenic mouse model. Proc Natl Acad Sci U S A. 2014;111(25):9115–20.
Song J, et al. Adiponectin receptor-mediated signaling ameliorates cerebral cell damage and regulates the neurogenesis of neural stem cells at high glucose concentrations: an in vivo and in vitro study. Cell Death Dis. 2015;6: e1844.
Kobayashi M, et al. Identification of a photoreceptor cell-specific nuclear receptor. Proc Natl Acad Sci U S A. 1999;96(9):4814–9.
Drewes T, et al. Human hepatocyte nuclear factor 4 isoforms are encoded by distinct and differentially expressed genes. Mol Cell Biol. 1996;16(3):925–31.
Jiang S, et al. Expression and localization of P1 promoter-driven hepatocyte nuclear factor-4alpha (HNF4alpha) isoforms in human and rats. Nucl Recept. 2003;1:5.
Yuan X, et al. Identification of an endogenous ligand bound to a native orphan nuclear receptor. PLoS ONE. 2009;4(5): e5609.
Crestani M, et al. LXR (liver X receptor) and HNF-4 (hepatocyte nuclear factor-4): key regulators in reverse cholesterol transport. Biochem Soc Trans. 2004;32(Pt 1):92–6.
Walsh BW, et al. Effects of raloxifene on serum lipids and coagulation factors in healthy postmenopausal women. JAMA. 1998;279(18):1445–51.
Mortensen MB, Nordestgaard BG. Elevated LDL cholesterol and increased risk of myocardial infarction and atherosclerotic cardiovascular disease in individuals aged 70–100 years: a contemporary primary prevention cohort. Lancet. 2020;396(10263):1644–52.
Sacks FM, et al. The effect of pravastatin on coronary events after myocardial infarction in patients with average cholesterol levels. Cholesterol and recurrent events trial investigators. N Engl J Med. 1996;335(14):1001–9.
Dormandy JA, et al. Secondary prevention of macrovascular events in patients with type 2 diabetes in the PROactive Study (PROspective pioglitAzone Clinical Trial In macroVascular Events): a randomised controlled trial. Lancet. 2005;366(9493):1279–89.
Lachin JM, et al. Renal function in type 2 diabetes with rosiglitazone, metformin, and glyburide monotherapy. Clin J Am Soc Nephrol. 2011;6(5):1032–40.
Haider AW, et al. Systolic blood pressure, diastolic blood pressure, and pulse pressure as predictors of risk for congestive heart failure in the Framingham Heart Study. Ann Intern Med. 2003;138(1):10–6.
Acknowledgements
The figure was generated using BioRender with the assistance of Fei Peng.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
M.W. and Y.Y. contributed to the search and collation of literature. M.W. and Y.Y. contributed to the manuscript preparation. M.W. and Y.X. contributed to the revision of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Wang, M., Yang, Y. & Xu, Y. Brain nuclear receptors and cardiovascular function. Cell Biosci 13, 14 (2023). https://doi.org/10.1186/s13578-023-00962-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13578-023-00962-3