
Abstract
Porcine edema disease (ED) is an enterotoxaemia that frequently occurs in 4–12 week-old piglets and results in high mortality. ED is caused by Shiga toxin 2e (Stx2e), produced by host-adapted Shiga toxin-producing Escherichia coli (STEC) strains. We constructed a recombinant protein in which the B subunit of Stx2e (Stx2eB) was linked to Cartilage Oligomeric Matrix Protein (COMP)’s pentameric domain to enhance antigenicity to induce neutralizing antibodies against Stx2e. We evaluated the efficacy of this antigen as a vaccine on the farm where ED had occurred. The suckling piglets were divided into two groups. The pigs in the vaccinated group were intramuscularly immunized with the vaccine containing 30 µg/head of Stx2eB-COMP at 1 and 4 weeks of age. The control pigs were injected with saline instead of the vaccine. The neutralizing antibody titer to Stx2e, mortality, clinical score, and body weight was evaluated up to 11 weeks after the first vaccination. In the vaccinated group, the Stx2e neutralizing antibody was detected 3 weeks after the first vaccination, its titer increased during the following weeks. The antibody was not detected in the control group during the test period. The STEC gene was detected in both groups during the test period, but a typical ED was observed only in control pigs; the mortality and clinical score were significantly lower in the vaccinated group than in the control group. These data indicate that the pentameric B subunit vaccine is effective for preventing ED and offers a promising tool for pig health control.
Similar content being viewed by others
Introduction
ED is an enterotoxaemia caused by Stx2e that frequently occurs in 4–12 week-old piglets. It is clinically characterized by swelling of the eyelids, paralysis, unusual voice, neurologic signs such as ataxia, lateral recumbency, and high mortality (50–90%) [1]. Therefore, ED causes great economic losses to pig farming.
ED effected pigs (or piglets) are treated with antibiotics, but the efficacy is poor because Stx2e has already been absorbed in the blood stream when clinical signs become apparent. Certain antibiotics such as quinolones, trimethoprim, and furazolidone could induce SOS response, by increasing the transcription of Stx2 genes [2]. In addition, as multidrug-resistant bacteria have been reported, antibiotics need to be selected appropriately [1]. As an alternative method, high amounts of zinc oxide may be added to feed [3], but there are concerns about the increase of drug-resistant bacteria and environmental pollution by zinc. Therefore, the exploitation of other control tools for ED, such as vaccines, is desired.
Stx2e is categorized in the Stx2 group of Shiga toxins. It has been reported as a developmental factor and protective antigen of ED vaccines containing Stx2e toxoid or genetically detoxified Stx2e by mutating the active toxin sites [4,5,6,7,8]. Although the vaccines have been confirmed to be effective against ED, they have shortcomings, such as a risk of side reactions due to the remaining toxin activity in the toxoid. Also, the yields of both vaccines were too low for commercial use [7,8,9,10].
ED is caused by STEC colonizing the small intestine [1]. Shiga toxin is an AB5 toxin protein (Figure 1A) and is transferred from the intestine into the bloodstream. It binds to a receptor on the surface of vascular endothelial cells via the B subunits. Then, the A subunit is internalized by receptor-mediated endocytosis. The A subunit having rRNA N-glycosidase activity induces cell death by inhibiting ribosomal protein synthesis [11].

Structure of Shiga toxin and Stx2eB-COMP. A All Shiga toxins have the AB5 toxin structure. A subunit is cleaved into A1 and A2 (held together by disulfide bonds). The five small B subunits surround the A2 region. B A structural diagram of Stx2eB-COMP. It was produced by fusion with COMP, also a pentamer, to stabilize the pentamer structure of the B subunit.
We hypothesized that a vaccine could be developed to prevent the onset of ED by blocking the binding of Stx2e to the cell surface receptor. Therefore, we used the B subunit having cell receptor binding ability as a vaccine antigen. Furthermore, as described above, since the Shiga toxin has the structure of AB5, we considered it important to maintain the B subunits as an immunizing antigen. Therefore, we created a recombinant protein (Stx2eB-COMP, Figure 1B) by fusing a Stx2e B subunit and a coiled-coil forming unit derived from COMP. COMP is a pentameric structure of oligomeric glycoprotein synthesized by chondrocytes [12]. In this study, to induce neutralizing antibodies against Stx2e, we developed a vaccine containing Stx2eB-COMP antigen as an oil-in-water emulsion; and evaluated the efficacy of this vaccine on a farm where ED had occurred.
Materials and methods
Construction of Stx2eB-COMP expressing vector and preparation of Stx2eB-COMP expressing E. coli
The nucleotide sequence for Stx2eB was artificially synthesized based on a Stx2eB gene (GenBank: NC_003356.1) which optimized the codons for expression in E. coli and yeast. First, the synthetic nucleotide was digested with NdeI and XhoI, and ligated into a plasmid pET-22b (Merck KGaA) to generate a plasmid pStx2eB. Next, the nucleotide sequence for matured Stx2eB containing no secretory signal sequence was amplified by PCR using a plasmid pStx2eB and primers (sense, 5′-CATGCCATGGATTGCGCGAAAGGCAAAATTG-3′ and antisense, 5′-CCGCTCGAGATTGAATTTCACCTGCGCAAAAC-3′). Finally, the amplified fragment was digested with NcoI and XhoI, and ligated into a plasmid pET-21d (Merck KGaA) to generate a pSTXB plasmid.
The nucleotide sequence for a fusion protein of (GP)2GH6(G4S)3 linker and COMP was amplified by PCR using a plasmid pB-spacer which includes nucleotide sequence for (G4S)3-COMP [13] and primers (sense, 5′-GGGCTCGAGGGTCCAGGACCTGGACATC-3′ and antisense, 5′-GGGCTCGAGTCAGCCCGGGGTACGGGCC-3′). The amplified fragment and a pSTXB plasmid were treated with XhoI and joined to generate a plasmid pSTXC. The pSTXC plasmid is a vector for expressing a fusion protein of Stx2eB, the (GP)2GH6(G4S)3 linker, and COMP. E. coli BL21(DE3) strain transfected with pSTXC, generates an E. coli strain STXC which expresses Stx2eB-COMP protein.
Cultivation of E. coli strain STXC and purification of expressed protein
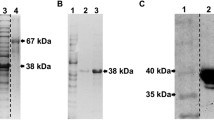
E. coli strain STXC was cultured in YE broth containing 50 µg/mL of ampicillin for 6 h at 37 °C. After adding isopropyl-β-d-thiogalactoside to express Stx2eB-COMP protein, it was further cultured for 40 h. After centrifugation, the harvested cells were washed with cell wash buffer (50 mM Tris–HCl (pH 8.0) containing 1 mM EDTA and 100 mM NaCl). The cells were subjected to high-pressure crushing, and the recovered centrifugal sediment was washed with protein-wash buffer (50 mM Tris–HCl (pH 8.0) containing 100 mM EDTA, 2% Triton X-100, and 2 M Urea) and distilled water. The obtained insoluble protein was solubilized with an unfolding buffer (20 mM Na2HPO4 (pH 8.2) containing 6 M guanidine hydrochloride (GuHCl) for 24 h at 37 °C, and then refolded with refolding buffer (20 mM Na2HPO4 (pH 8.2) containing 1 M arginine hydrochloride and 2 mM l-cystine) for 24 h at 25 °C. The protein solution was concentrated and the medium was replaced with PBS (pH 8.0) by ultrafiltration and then filtered using a 0.22 μm filter. The obtained Stx2eB-COMP protein was used as a vaccine antigen. The Stx2eB-COMP protein concentration was determined by densitometry (Quantity one ver.4.6, BIO-RAD, CA, US) after gel electrophoresis.
Vaccine
An oil-in-water type emulsion vaccine containing 30 µg of the Stx2eB-COMP protein in 1 mL per dose was prepared according to the following procedure. Liquid paraffin (Kaneda Co., Ltd, Tokyo, Japan) adjuvant and emulsifiers of Polysorbate 80 (NOF Corporatin, Tokyo, Japan) and sorbitan monooleate (NOF Corporatin, Tokyo, Japan) were mixed with PBS and emulsified using a homogenizer. One volume of Stx2eB-COMP antigen was mixed with two volumes of emulsified adjuvant and used as a trial vaccine.
Farm trial and its ethical statement
The test was conducted on a farm where ED had occurred. The 159 healthy crossbred suckling piglets (Landrace, Large-White, Duroc) at 1 week of age were divided into two groups without weight and gender bias. The vaccinated group (male: 40, female: 39) was injected with 1 mL of vaccine into the neck muscle at a 3-week interval, and the control group (male: 40, female: 40) was injected with 1 mL of saline as the vehicle. After weaning at 4 weeks of age, efficacy was evaluated until 11 weeks after the first vaccination (12 weeks old). Piglets were bred during the test period in a facility with constant feeding and water supply. This clinical trial was conducted in compliance with the “Act on Welfare and Management of Animals” [14] and following the KYODOKEN Institute guidelines for animal studies.
Detection of STEC
STEC was identified by PCR targeting the stx2e gene. Small intestinal contents and mesenteric lymph nodes collected at the time of death, and fecal swabs were used as test samples. The fecal swabs were collected from piglets that indicated fecal abnormalities during the test period.
Purification of Stx2e
STEC was cultured in YE broth overnight at 37 °C. The STEC culture suspension was centrifuged, and the precipitate was suspended in 20 mM phosphate buffer (pH7.0). The harvested cells were sonicated and centrifuged, the supernatant was purified by affinity chromatography (Cellfine™ Sulfate, JNC Corporation, Tokyo, Japan), and then dialyzed with PBS. Purified Stx2e was filtered with a 0.22 µm filter and stored frozen.
Stx2e neutralizing antibody assay
Sera were treated for 30 min at 56 °C before use, and then serially diluted twofold with supplemented Eagle minimal essential medium (E-MEM, Nissui Pharmaceutical Co., Ltd, Tokyo, Japan) containing 5% fetal bovine serum, 3.0 mg/mL trypticase soy broth (Becton, Dickinson and Company, Tokyo, Japan), 1.1 mg/mL sodium bicarbonate (FUJIFILM Wako Pure Chemical Corporation. Osaka, Japan), 2 mM l-glutamine (FUJIFILM Wako Pure Chemical Corporation. Osaka, Japan), 100 U/mL penicillin (Meiji Seika Pharma Co., Ltd, Tokyo, Japan), and 100 µg/mL streptomycin (Meiji Seika Pharma Co., Ltd, Tokyo, Japan). Then, 10 CD50 (median cytotoxic dose) of purified Stx2e were added to each serum sample and the toxin-antiserum mixtures were incubated at 37 °C for 1 h. Toxin-antiserum mixture (50 µL) was added to 96-well plates containing 100 µL of supplemented E-MEM, followed by 50 µL (4 × 105 cells/mL) of Vero cells (ATCC CCL-81) added to each well, and the plates were incubated at 37 °C in 5% CO2 for 5 days. The cell viability was assessed using the Cell Counting Kit-8 (CCK-8, DOJINDO LABORATORIES, Kumamoto, Japan). CCK-8 solution (50 µL) diluted 11 times with supplemented E-MEM was added into each well. The cultures were incubated at 37 °C in 5% CO2 for 2 h. The absorbance of each well was measured at 450 nm using a microplate reader (SUNRISE RAINBOW THERMO, Tecan Japan Co., Ltd., Kanagawa, Japan). When the value obtained by dividing the absorbance value of the sera by that of the control well (no toxin addition) was more than or equal to 0.5, the neutralizing effect of Stx2e was deemed positive. The neutralizing antibody titer was the reciprocal of the highest dilution of serum which had a Stx2e neutralizing effect.
Assessment of vaccine efficacy
Stx2e neutralizing antibody titer, mortality, clinical score, and body weight of the groups were compared to evaluate the vaccine efficacy. Sera were collected from 30 heads of the vaccinated group and 10 heads of the control group at 0, 3, 7, and 11 weeks after the first vaccination. The Stx2e neutralizing antibodies in serum were detected by the Stx2e neutralizing antibody assay. Clinical signs were scored between 0 and 11 weeks after the first vaccination using the criteria shown in Table 1 [15]. The body weights of all animals were measured at weeks 0, 3, 7, and 11 after the first vaccination.
Statistical analysis
Mortality was tested using the Fisher exact test. The body weight was subjected to an F test followed by the Student t-test or Aspin-Welch t-test. The clinical scores were evaluated using the Mann–Whitney U test. The significance level was ˂ 5% in all cases.
Results
Mortality
Seven out of eighty piglets showed clinical signs such as palpebral edema, intestinal edema/hyperemia, and mesenteric lymphadenopathy and died between days 38 and 67 in the control group (Table 2). The symptoms observed in these piglets were characteristic of edema disease. In the vaccinated group, seventy-nine piglets showed significantly lower clinical signs (scores) compared to the control group, and no piglets died. The mortality rates (%) were 0.0 (0/79) in the vaccinated group and 8.8 (7/80) in the control group, showing a significant difference in mortality between the groups.
Clinical score
As described in the sub-section “Mortality”, only control pigs, but no vaccinated pigs died. The clinical score increased in both groups, but the score of the vaccinated group remained lower than that of the control group (Figure 2). There were significant differences in clinical scores between the groups from weeks 3 to 11 after the first vaccination. In the vaccinated group, the clinical score of feces were significantly lower during 3 to 4 weeks after the first vaccination, and from week 5 onwards, other clinical scores were significantly lower than those of the control group.

No vaccinated piglets died and showed low clinical scores. The total clinical score (mean ± SEM) at 0, 2, 3, 4, 5, 7, 9, 11 weeks after 1st vaccination and the number of deaths during the test period are shown. *: There are significant differences (P < 0.05) in the clinical scores of the two groups.
Body weight gain
Body weight was not significantly different between the two groups at 3 weeks after the first vaccination. However, it was significantly higher in the vaccinated group than in the control group at 7 (18.8 kg vs. 17.4 kg) and 11 weeks (35.7 kg vs. 31.8 kg) after the first vaccination (Table 3).
Detection of STEC
STEC was detected in fecal swabs collected from the control group between 23–41 days after the first vaccination and from the mesenteric lymph nodes of dead piglets during the test period (Tables 2 and 4). In the vaccinated group, STEC was only detected in fecal swabs collected on day 34 after the first vaccination. The degree of abnormality in the fecal samples varied from clinical score 1 (loose stool) to 3 (severe diarrhea).
Stx2e neutralizing antibody
The Stx2e neutralizing antibody titer increased from the week 3 after the first vaccination and showed a peak value of 88.4 at week 7 after the first vaccination. On the contrary, the titer of the control group was negative during the test period despite the onset of symptoms and subsequent death.
Discussion
We developed a trial vaccine consisting of a purified Stx2eB-COMP protein in an oil-in-water emulsion and evaluated its efficacy in a trial on a farm where ED had occurred. The neutralizing antibody titer increased significantly in the vaccinated group. The group showed a lower clinical score and higher body weight than the control group. STEC was detected from fecal swabs of both groups, meaning that STEC was spread throughout the farm during the test period. However, the mortality in the vaccinated group was 0%, while that in the control group was 8.8%. These data indicate that this vaccine candidate can prevent the onset of ED. Moreover, no serious adverse reactions were observed after administering this trial vaccine.
This study focused on the Stx2e B subunit as a vaccine antigen to block toxin binding. We confirmed the vaccine efficacy due to the antibody to the B subunit induced by immunization with the Stx2eB-COMP protein that inhibited the binding of Stx2e to the receptor on the surface of vascular endothelial cells. Meanwhile, it has been reported that the B subunit alone induces little of the Stx2e neutralizing antibodies compared to the holotoxin [16]. In addition, the sera obtained by immunizing Stx2 B subunit monomers could not recognize Stx2 [17]. In our previous study, we compared neutralizing antibody induction by Stx2eB alone and Stx2eB-COMP using 7-week-old BALB/c mice. The result confirmed that the Stx2eB-COMP antigen induces neutralizing antibodies against Stx2e in mice, while no increase in neutralizing antibodies was observed in the Stx2eB alone injection group [18]. These reports and data suggest that the multimeric structure of the B subunit is important for the maintenance of neutralizing epitopes. Therefore, we considered that the pentameric structure of the B subunit was important for inducing Stx2e neutralizing antibodies efficiently; therefore, we produced the Stx2eB-COMP.
Clinical scores except those of feces were significantly inhibited at 7 weeks post-vaccination. The difference in clinical scores between vaccinated and control groups was the highest at this time point. These clinical symptoms, such as vitality, appetite, respiration, palpebral edema, neurologic impairment, and piloerection are considered to be due to Stx2e-induced toxemia. This inhibitory effect is presumed to be due to the neutralization of the toxin by the induced antibodies, suggesting that the induction of neutralizing antibodies against Stx2e is important for ED vaccines.
Although STEC were isolated from the control group, the Stx2e neutralizing antibody remained negative in this group during the test period. Gannon et al. showed previously that no Stx2e neutralizing antibody was detected in swine herds suffering from ED [19], confirmed in this study since no neutralizing antibody was detected even after STEC infection. Additionally, although the animal species are different, when a STEC oral infection experiment was conducted in cattle, the antibody against Stx1 increased rapidly in adult cattle (steered cattle), but gradually in calves. Moreover, no antibody response to Stx2 was developed [20]. It is inferred from these data that Stx2 has an extremely weak antibody-inducing ability in living organisms. Therefore, we believe that our vaccine is promising because it can induce neutralizing antibodies that are normally difficult to produce in STEC infection in piglets.
In conclusion, our trial vaccine induced Stx2e neutralizing antibodies efficiently using only the B subunits of Stx2e and effectively prevented ED. The result indicates that the A subunit is not essential for ED prevention. Since the antibodies against Stx2e were not able to inhibit STEC from developing, the clinical signs shown in some of vaccinated piglet might be caused by exceeded levels of un-neutralised Stx2e. These results indicate that this vaccine can be a new tool for prevention of ED exacerbation by Stx2e.
Availability of data and materials
The datasets of this clinical study are available from the corresponding author upon reasonable request.
References
Casanova NA, Redondo LM, Dailoff GC, Arenas D, Fernández Miyakawa ME (2018) Overview of the role of Shiga toxins in porcine edema disease pathogenesis. Toxicon 148:149–154
Kimmitt PT, Harwood CR, Barer MR (2000) Toxin gene expression by shiga toxin-producing Escherichia coli: the role of antibiotics and the bacterial SOS response. Emerg Infect Dis 6:458–465
Uemura R, Katsuge T, Sasaki Y, Goto S, Sueyoshi M (2017) Effects of zinc supplementation on Shiga toxin 2e-producing Escherichia coli in vitro. J Vet Med Sci 79:1637–1643
Gordon VM, Whipp SC, Moon HW, O’Brien AD, Samuel JE (1992) An enzymatic mutant of Shiga-like toxin II variant is a vaccine candidate for edema disease of swine. Infect Immun 60:485–490
Bosworth BT, Samuel JE, Moon HW, O’Brien AD, Gordon VM, Whipp SC (1996) Vaccination with genetically modified Shiga-like toxin IIe prevents edema disease in swine. Infect Immun 64:55–60
Johansen M, Andresen LO, Jorsal SE, Thomsen LK, Waddell TE, Gyles CL (1997) Prevention of edema disease in pigs by vaccination with verotoxin 2e toxoid. Can J Vet Res 61:280–285
Oanh TK, Nguyen VK, De Greve H, Goddeeris BM (2012) Protection of piglets against Edema disease by maternal immunization with Stx2e toxoid. Infect Immun 80:469–473
MacLeod DL, Gyles CL (1991) Immunization of pigs with a purified Shiga-like toxin II variant toxoid. Vet Microbiol 29:309–318
MacLeod DL, Gyles CL (1990) Purification and characterization of an Escherichia coli Shiga-like toxin II variant. Infect Immun 58:1232–1239
Acheson DW, Jacewicz M, Kane AV, Donohue-Rolfe A, Keusch GT (1993) One step high yield affinity purification of shiga-like toxin II variants and quantitation using enzyme linked immunosorbent assays. Microb Pathog 14:57–66
Doughari JH, Ndakidemi PA, Human IS, Bennade S (2009) Shiga toxins (Verocytotoxins). Afr J Microbiol Res 3:681–693
Oldberg A, Antonsson P, Lindblom K, Heinegård D (1992) COMP (cartilage oligomeric matrix protein) is structurally related to the thrombospondins. J Biol Chem 267:22346–22350
Arakawa T, Harakuni T (2014) Cholera toxin B subunit-five-stranded α-helical coiled-coil fusion protein: “five-to-five” molecular chimera displays robust physicochemical stability. Vaccine 32:5019–5026
Act on Welfare and Management of Animals (Act No. 105 of October 1, 1973) https://www.env.go.jp/nature/dobutsu/aigo/1_law/files/aigo_kanri_1973_105_en.pdf. Accessed 12 Sept 2022
Tsukahara T, Nakanishi N, Nakayama K, Matsubara N, Ushida K (2005) Experimental infection of enterotoxemic Escherichia coli associated with porcine edema disease and its pathologic characteristics in the intestine. J Vet Med Sci 67:1167–11671
Sato T, Matsui T, Takita E, Kadoyama Y, Makino S, Kato K, Sawada K, Hamabata T (2013) Evaluation of recombinant forms of the Shiga toxin variant Stx2eB subunit and non-toxic mutant Stx2e as vaccine candidates against porcine edema disease. J Vet Med Sci 75:1309–1315
Acheson DWK, De Breucker SA, Jacewicz M, Lincicome LL, Donohue-Rolfe A, Kane AV, Keusch GT (1995) Expression and purification of Shiga-like toxin II B subunits. Infect Immun 63:301–308
Yokogawa K, Waki T, Honda Y, Uefuji H, Sewaki T, Arakawa T, Harakuni T and Miyata T. Vaccine for preventing porcine edema disease. Patent: PCT/JP2013/078305. https://patents.google.com/patent/WO2014065210A1/en. Accessed 12 Sept 2022
Gannon VPJ, Gyles CL, Friendship RW (1988) Characteristics of verotoxigenic Escherichia coli from pigs. Can J Vet Res 52:331–337
Johnson RP, Cray WC Jr, Johnson ST (1996) Serum antibody responses of cattle following experimental infection with Escherichia coli O157:H7. Infect Immun 64:1879–1883
Acknowledgements
This work was supported by JECTAS INNOVATORS Co. Ltd. (#705 1-29-15 Tsuboya, Naha 902-0065, Okinawa, Japan) and Kyodoken Institute Co. Ltd. (585 Shimoitabashi, Kyoto 612-8073, Japan). The authors also wish to thank Mr. Kenichi Michishita and Dr Tomoyuki Tsuda for scientific discussion and editorial assistance in the preparation of this paper.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Author information
Authors and Affiliations
Contributions
RT, KS, and TW designed this study; YH, EY, RT, and TW manufactured the trial vaccine; MY, KS, and TW analyzed and interpreted the data; MY and TW prepared the main body of this manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This clinical trial was conducted following the GCP Guidelines, in compliance with the “Act on Welfare and Management of Animals”, and according to the guidelines of the KYODOKEN Institute (Approval number: CS142071, KYODOKEN 3169) for animal studies. In addition, the trial was conducted with the consent of the owner of Asagiri Farm, who owns the animals.
Competing interests
The authors declare that they have no competing interests.
Additional information
Handling editor: Tina S. Dalgaard
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Yoshimura, M., Honda, Y., Yonemitsu, E. et al. Induction of antitoxic antibody and preventive effect against porcine edema disease by the pentameric Stx2eB subunit vaccine. Vet Res 54, 29 (2023). https://doi.org/10.1186/s13567-023-01161-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13567-023-01161-1