
Abstract
Glutamine has a positive effect on ameliorating reproductive failure caused by porcine circovirus type 2 (PCV2). However, the mechanism by which glutamine affects PCV2 replication remains unclear. This study was conducted to investigate the effects of glutamine on PCV2 replication and its underlying mechanisms in vitro. The results show that glutamine promoted PK-15 cell viability. Surprisingly, glutamine starvation significantly increased PCV2 replication. The promotion of PCV2 replication by glutamine starvation disappeared after fresh media with 4 mM glutamine was added. Likewise, promotion of PCV2 was observed after adding buthionine sulfoximine (BSO). Glutamine starvation or BSO treatment increased the level of p38 MAPK phosphorylation and PCV2 replication in PK-15 cells. Meanwhile, p38 MAPK phosphorylation and PCV2 replication significantly decreased in p38-knockdown PK-15 cells. Promotion of PCV2 replication caused by glutamine starvation could be blocked in p38-knockdown PK-15 cells. Therefore, glutamine starvation increased PCV2 replication by promoting p38 MAPK activation, which was associated with the down regulation of intracellular glutathione levels. Our findings may contribute toward interpreting the possible pathogenic mechanism of PCV2 and provide a theoretical reference for application of glutamine in controlling porcine circovirus-associated diseases.
Similar content being viewed by others


Introduction
Porcine circovirus type 2 (PCV2) has been associated with porcine circovirus-associated diseases (PCVD), which collectively include the post-weaning multi-systemic wasting syndrome (PMWS), the porcine dermatitis and nephropathy syndrome (PDNS), porcine respiratory and reproductive disorders, proliferative and necrotizing pneumonia (PNP), congenital tremors (CT), and enteric disease [1]. Infection with PCV2 is necessary but not sufficient for pigs to develop PCVD. Infection with PCV2 alone does not generally cause overt clinical disease [2]. Most of the available data indicate that appropriate host, management, co-infection, immunostimulation, and nutrition are crucial for disease progression to PCVD [3]. However, the pathogenic mechanism of PCV2 remains poorly understood.
Glutamine (Gln) is the most abundant free amino acid in serum; this amino acid is important for the regulation of metabolism, cell integrity, protein synthesis, redox potential, gene expression, and intracellular signaling pathways [4]. Glutamine can be produced in sufficient quantities under normal physiological conditions, but becomes an essential amino acid during pathological or stress conditions [5]. Dietary l-glutamine supplementation can reportedly ameliorate the reproductive failure caused by PCV2 [6] and enhance the immune functions in PCV2-infected mice [7]. Dietary glutamine supplementation may confer a positive effect on the improvement of pregnancy outcomes in PCV2-infected mice by enhancing the immune response and the ability to clear PCV2 [6]. However, the mechanism by which glutamine affects PCV2 replication remains unclear.
Glutamine has an important role in cell culture in vitro. This amino acid is required by all mammalian and invertebrate cell lines as an energy substrate and a precursor for nucleotide, glutamate, and glutathione synthesis [8]. Furthermore, previous studies have suggested that glutamine affects the replication of viruses through different mechanisms within host cells. Dietary glutamine supplementation can protect the host from inflammation and infection [9] by stimulating glutathione synthesis in animal cells [10], which may lead to the activation of p38 mitogen-activated protein kinase (MAPK) [11,12]. Glutamine-deficient medium has been demonstrated to increase psittacosis virus multiplication [13], whereas Sendai virus proliferation in BHK cells is suppressed by a lack of glutamine [14]. In addition, glutamine deprivation enhances the plating efficiency of a herpes simplex virus type 1 ICP0-null mutant [15]. During human cytomegalovirus infection, glutamine is essential for virion production in cells [16]. Glutamine deficiency triggers the decrease in cellular glutathione (GSH) levels and promotes oxidative stress in HuH-7 cells [17]. In addition, GSH supplementation decreases DV2 production in HepG2 cells [18]. However, to the best of our knowledge, the influence of glutamine on PCV2 replication in PK-15 cells has not been reported to date.
Consequently, the present study investigated the effects of glutamine on PCV2 replication and its underlying mechanisms.
Methods
Cell culture and virus propagation
Dulbecco’s modified Eagle’s medium (DMEM) was used as the base medium for cell culture. The PCV1-free PK-15 continuous cell line was propagated and maintained at 37 °C in 5% CO2 in DMEM (Wisent, Nanjing, China) supplemented with 10% fetal calf serum, 100 U/mL penicillin, 0.1 mg/mL streptomycin, and 4 mM glutamine (hereafter referred to as complete medium) [19]. The wild-type PCV2 (PCV2NJ2002) used in the experiment was originally isolated from a kidney tissue sample of a pig with naturally occurring PMWS. The PCV type was determined by sequencing (TaKaRa Co., Dalian, China). PK-15 cells were infected with a PCV2 lysate at a multiplicity of infection (MOI) of 1. At 72 h after infection, approximately 26% of the PK-15 cells in the 96-well plates could be infected, as measured by indirect All experiments were undertaken following the guidelines of the Nanjing Agricultural University Ethics Committee, China.
Reagents and antibodies
Rabbit antibodies specific for p38 and the phosphorylated form of p38 (p-p38) were obtained from Cell Signaling Technology. The porcine anti-PCV2 antibody for immunofluorescence was purchased from Univ Biotech Co. Ltd., Shanghai, China. The FITC-linked secondary antibody (rabbit anti-pig IgG) was purchased from Sigma, St. Louis, USA.
Cell viability assay
PK-15 cells (2 × 104 cells/well) were plated onto 96-well plates in the presence of glutamine (0, 0.1, 0.5, 1, 2, 4, 8, and 16 mM). After 24 h of incubation, the cells were subjected to colorimetric 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay (Sigma) according to the protocol of the manufacturer. The absorbance of each well was measured at 550 nm, with a reference wavelength of 690 nm.
Quantification of PCV2 DNA by real-time PCR
The presence of newly synthesized viral DNA in the PK-15 cells was assayed by real-time PCR with the TaKaRa SYBR Green qPCR Kit (TaKaRa), as described elsewhere [20]. Briefly, the PCV2-infected PK-15 cells were harvested at 72 h post-inoculation, and DNA extraction was performed with a TaKaRa DNA Mini kit (TaKaRa). The purified DNA was used as a template for real-time PCR amplification. A 117 bp region was amplified from ORF2 of the PCV2 gene with a pair of PCV2-specific primers (forward primer: 5′-TAGTATTCAAAGGGCACAG-3′; reverse primer: 5′-AAGGCTACCACAGTCAG-3′). Quantitative real-time PCR was performed with the ABI PRISM 7300 Detection System (Applied Biosystems, Foster, USA). A recombinant pMD19 plasmid vector (TaKaRa) containing PCV2 genome inserts was used as the standard. Tenfold serial plasmid dilutions were tested and used to construct the standard curve by plotting the logarithm of the plasmid copy number against the measured CT values. The generated standard curve covered a linear range of 1 × 104–1 × 1010 copies/mL for PCV2. Regression analysis of the linear portion of the curve produced a slope coefficient of −0.326, whereas the linear correlation (R 2) between the logarithm of the plasmid copy number and CT was 0.998, with a Y-intercept of 45.087. The number of PCV2 DNA copies was calculated from the number of cells, specifically, the average amount of PCV2 DNA copies per 106 cells, according to previous publications [21,22].
Indirect immunofluorescence of PCV2-infected cells
At 72 h post-inoculation, the PK-15 cells were washed with phosphate-buffered saline (PBS) and fixed in methanol. The fixed cells were incubated in PBS containing 2% bovine serum albumin (BSA) at room temperature for 1 h, followed by incubation with the porcine anti-PCV2 antibody (Univ Biotech; diluted to 1:50 in PBS containing 1% BSA) at 37 °C for 1 h. After washing with PBS containing 0.1% Tween-20, cells were incubated with the FITC-conjugated rabbit anti-pig antibody (diluted to 1:100 in PBS containing 1% BSA) at 37 °C for 1 h. After washing in PBS, the cells were examined under a fluorescence microscope. Cells with the fluorescent stain (judged to be positive for the PCV2 viral antigen) were counted in six fields of view for each concentration in three replicated cultures. The calculation of the relative proportions of infected cells was based on the total amount of cells in each field.
Western blot analysis
The cell lysates were suspended in RIPA lysis buffer (Beyotime, Haimen, China). PMSF was added to the cell suspensions, followed by centrifugation at 8000 × g for 10 min at 4 °C. The supernatant fluids were stored at −80 °C before use. For each sample, 50 μg was subjected to SDS-PAGE, and the separated proteins were transferred onto a polyvinylidene difluoride membrane (Bio-Rad, Hercules, USA). The membranes were blocked for 1 h at room temperature in Tris-buffered saline (TBS) containing 5% BSA and 0.1% Tween 20. Subsequently, the membranes were incubated with the respective specific primary antibodies against p-p38, p38, and β-actin at 4 °C overnight. The membranes were washed thrice with TTBS buffer and incubated with the respective secondary antibodies for 1 h at room temperature. The Western blots were visualized with the SuperSignal ECL kit (Thermo, Waltham, USA).
SiRNA transfection
Double-stranded RNA sequences were designed with Invitrogen BlockiT RNAi Designer according to the Sus scrofa p38 mRNA sequence (Genbank accession number XM_003356616.1); the designed sequences were then synthesized by Invitrogen. The p38-specific siRNA sequence was 5′-GCAGGAGCUGAACAAGACATT-3′. The control siRNA had the following sequence: 5′-UUCUCCGAACGUGUCACGUTT-3′. The duplexes were resuspended in DEPC-treated water to obtain 20 μM solutions prior to use. PK-15 cells in DMEM with 10% fetal bovine serum (FBS) but without antibiotics were seeded into 24-well plates at a cell density of 5 × 104 cells/well. The cells were incubated for 24 h at 37 °C in a 5% CO2 atmosphere. When the cells were 30%–50% confluent, siRNA was introduced via the X-tremeGene siRNA transfection reagent (Roche, Mannheim, Germany), according to the manufacturer’s protocol. Subsequently, 2.5 μL transfection reagent and 0.5 μg siRNA were added to each well, and the cells were incubated for 4 h. Finally, the cells were washed with DMEM and cultured for another 20 h in DMEM containing 2% FBS before the subsequent experiments.
Determination of GSH and MDA in PK-15 cells
Cell extracts were prepared by sonication in ice-cold PBS. After sonication, the cell lysates were centrifuged at 10 000 × g for 20 min to remove the cell debris. The amount of reduced glutathione and maliondehyde (MDA) in the supernatants were measured with commercially available kits (Jiancheng Co., Nanjing, China), as previously described [23,24]. GSH was spectrophotometrically measured (412 nm) via a reaction with 5,5′-dithiobis(2-nitrobenzoic acid); the GSH concentrations were expressed as the micromoles of GSH per gram of protein. MDA was spectrophotometrically assayed at 548 nm via the thiobarbituric acid reaction; the MDA concentrations were expressed as micromoles per gram of protein. The total protein concentration was determined with a Bradford Protein Assay Kit (Beyotime). All samples were assayed in triplicate.
Statistical analysis
The experimental data was analyzed by one-way analysis of variance (ANOVA). The least-significant difference test was used to compare the differences among the groups. The results are presented as the mean ± SD of replicates. Differences among groups were considered significant when P < 0.05.
Results
Glutamine promotes the viability of PK-15 cells
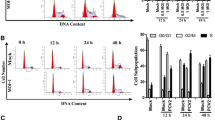
To assess the effect of glutamine on the growth of PK-15 cells with or without PCV2 infection, PK-15 cells were grown in 96-well plates with DMEM containing 4 mM glutamine for 24 h until 50% confluence was reached. Subsequently, the medium was changed to fresh DMEM with various concentrations of glutamine (0–16 mM). The cells were incubated for an additional 48 h before the cell viability was determined. As shown in Figures 1A and 1B, the significant promotion of cell viability was observed when glutamine was present at 4 mM or higher as compared with groups subjected to glutamine starvation (P < 0.05). The cell viability was significantly decreased by 24%, 50%, 81%, and 87% in non-infected groups incubated with glutamine at concentrations of 1, 0.5, 0.1, and 0 mM, respectively, as compared with the controls with 4 mM glutamine treatment. The relative cell viability of the infected cells was 93%, 68%, 49%, 28%, and 23% when glutamine was supplemented at concentrations of 2, 1, 0.5, 0.1, and 0 mM, respectively. The promotion of PK-15 cell viability by glutamine was concentration dependent within the range of 0–2 mM glutamine. Our data indicate that glutamine plays an important role in PK-15 cell viability. Glutamine starvation inhibited the growth of PK-15 cells when its concentration was less than 4 mM in the medium.

Effects of glutamine on viability of PK- 15 cells. PK-15 cells without (A) or with (B) PCV2 infection were grown in 96-well plates with DMEM containing 4 mM glutamine for 24 h until 50% confluence was reached. Subsequently, the medium was changed to fresh DMEM with various concentrations of glutamine (0–16 mM). The cells were incubated for an additional 48 h before the cell viability was determined by MTT assay. Values are given as mean ± SD from three independent experiments. Groups were compared by a 1-way ANOVA followed by a least-significant difference test (*P < 0.05, **P < 0.01).
Effect of glutamine starvation on PCV2 replication in PK-15 cells
To establish the viral infection and ensure that PK-15 cells were able to survive in glutamine-free medium in our experiment, the cells were infected with PCV2 at a MOI of 1 and incubated in the complete medium for 24 h. Subsequently, PK-15 cells were grown in media containing various concentrations of glutamine. After 48 h, the PK-15 cells were harvested to determine the number of PCV2 DNA copies and the relative proportion of PCV2-infected cells by real-time PCR and immunofluorescence microscopy, respectively. The PCV2 log 10 DNA copies per 106 cells (Figure 2A) and the proportion of PCV2 infected cells (Figures 2B and 2C) were significantly increased when the glutamine concentration was below 2 mM as compared with cells grown with >4 mM glutamine (P < 0.05). The number of PCV2 log 10 DNA copies per 106 cells was significantly increased by 2%, 4%, 5%, and 6% in groups with glutamine starvation at concentrations of 1, 0.5, 0.1, and 0 mM, respectively, as compared with the controls treated with 4 mM glutamine. The relative proportions of the PCV2-infected cells were 116%, 125%, 138%, and 145% when glutamine was supplemented at concentrations of 1, 0.5, 0.1, and 0 mM, respectively. Our results show that glutamine starvation increases PCV2 replication.

Effects of different concentrations of glutamine on PCV2 replication. PK-15 cells were infected with PCV2 at an MOI of 1 and incubated in the complete medium for 24 h. Subsequently, PK-15 cells were grown in medium containing various concentrations of glutamine. After 48 h, the PK-15 cells were harvested to determine the number of PCV2 log 10 DNA copies per 106 cells (A) and the relative proportion of PCV2-infected cells (B and C) by real-time PCR and immunofluorescence microscopy, respectively. Values shown are means ± SD mean from three independent experiments. Asterisks indicate groups statistically significantly different from control by a 1-way ANOVA followed by least-significant difference test (**P < 0.01).
Another set of experiments was performed to determine whether glutamine supplementation blocks the positive effect of glutamine starvation on PCV2 replication. First, PK-15 cells were infected with PCV2 at an MOI of 1 in complete medium for 24 h. These PK-15 cells were subsequently grown in medium containing various concentrations of glutamine for 48 h before the media of all treatment groups were changed to fresh media with 4 mM glutamine and the cells were incubated for another 48 h. Finally, the number of PCV2 DNA copies and the relative proportion of PCV2-infected cells were calculated. As shown in Figure 3, the shift to fresh media with 4 mM glutamine at 72 h and the additional incubation for 48 h did not significantly alter the PCV2 replication in all groups (P > 0.05). The results indicate that the addition of fresh media with 4 mM glutamine blocked the promotion of PCV2 replication by glutamine starvation.

Glutamine supplementation blocks the promotion of PCV2 replication by glutamine starvation. PK-15 cells were infected with PCV2 at an MOI of 1 in complete medium for 24 h. These PK-15 cells were subsequently grown in medium containing various concentrations of glutamine for 48 h before the media of all treatment groups were changed to fresh media with 4 mM glutamine. The cells were incubated for another 48 h and then harvested to determine the number of PCV2 log 10 DNA copies per 106 cells (A) and the relative proportion of PCV2 infected cells (B and C). Groups were compared by a 1-way ANOVA followed by a least-significant difference test. Significant changes are indicated by *(P < 0.05) in comparison with control.
GSH levels decrease as the MDA concentration increases in PK-15 cells with glutamine starvation
Glutamine can reportedly act as a precursor for glutathione synthesis [25]. Given the findings of previous studies [26,27], we hypothesized that the increased replication of PCV2 in PK-15 cells during glutamine starvation may be attributed to decreased GSH levels. To test this hypothesis, intracellular GSH levels were measured in PCV2-infected cells grown with various glutamine concentrations. After 24 h of PCV2 infection in complete medium, fresh glutamine-free medium with various glutamine concentrations was added to replace the original medium, and the cells were cultured for another 72 h. As shown in Figure 4, the intracellular GSH levels were significantly decreased by 16%, 23%, 28%, and 29% in PK-15 cells with glutamine starvation at concentrations of 1, 0.5, 0.1, and 0 mM, respectively, as compared with the control cells with 4 mM glutamine (Figure 4A; P < 0.05). The MDA concentration was significantly increased by 82%, 191%, and 211% in PK-15 cells with glutamine starvation at concentrations of 0.5, 0.1, and 0 mM, respectively (Figure 4B; P < 0.01). These results suggest that glutamine starvation induced oxidative stress by decreasing the GSH level and increasing the MDA concentration in PK-15 cells.

Effect of glutamine starvation on GSH levels, MDA concentration and PCV2 replication. After 24 h of PCV2 infection in complete medium, fresh glutamine-free medium with various concentrations of glutamine and BSO was added to replace the original medium. The cells were cultured for another 48 h before the determining of GSH levels (A and C), MDA concentration (B), the number of PCV2 log 10 DNA copies per 106 cells (D), and the proportion of PCV2 infected cells (E and F). Values are shown as mean ± SD from three independent experiments. Asterisks indicate groups statistically significantly different from control by a 1-way ANOVA followed by a least-significant difference test (*P < 0.05, **P < 0.01).
PCV2 replication increases in PK-15 cells after GSH synthesis is inhibited by BSO
To further confirm the role of decreased GSH levels in the promotion of PCV2 replication induced by glutamine starvation, BSO was used; this compound is a well-known specific inhibitor of GSH synthesis [28]. After 24 h of PCV2 infection, the complete medium was removed, and the infected cells were cultured for another 48 h in the absence of glutamine or in the presence of glutamine and BSO. Compared with the control group grown with 4 mM glutamine, the intracellular GSH concentrations were significantly reduced in the PK-15 cells treated with glutamine and BSO (P < 0.01; Figure 4C). Therefore, exogenous BSO attenuated the intracellular GSH levels in the presence of glutamine. The number of PCV2 DNA copies were increased (Figure 4D) and the relative proportion of infected cells (Figures 4E and 4 F) were enhanced by 36.2% and 51.0% in the presence of 4 mM glutamine after treatment with 20 and 50 μM BSO, respectively, as compared with the controls (P < 0.01). These results showed that PCV2 replication was increased in PK-15 cells after GSH synthesis was inhibited by BSO. Therefore, GSH may play an important role in the promotion of PCV2 replication during glutamine starvation.
Glutamine starvation or BSO treatment increase the level of p38 MAPK phosphorylation
p38 MAPK has been confirmed to have an important role in the efficient replication of PCV2 in PK-15 cells [29]. Therefore, the effect of glutamine starvation on PCV2 replication may be associated with the phosphorylation of p38 MAPK, which is induced by attenuated GSH synthesis. To address this question, we calculated the rate of p38 MAPK phosphorylation in PK-15 cells by measuring the p38 mRNA and protein levels via real-time PCR and Western blot analysis, respectively. After 24 h of PCV2 infection, the complete medium was removed, and fresh medium with various concentrations of glutamine was added for an additional 48 h of incubation. As shown in Figure 5, the levels of p38 MAPK phosphorylation increased by 1.53-, 2.21-, 2.12-, and 2.19-fold after supplementation with 1, 0.5, 0.1 and 0 mM glutamine, respectively, as compared with the controls incubated with 4 mM glutamine (Figure 5; P < 0.01). However, the relative p38 mRNA levels were not significantly different among groups with various concentrations of glutamine (data not shown). These results indicate that glutamine starvation increased the level of p38 MAPK phosphorylation.

Glutamine starvation increases the level of p38 MAPK phosphorylation. After 24 h of PCV2 infection, the complete medium was removed, and fresh medium with various concentrations of glutamine was added for an additional 48 h of incubation. The protein levels of p38 phosphorylation were measured by Western blot. Values are shown as mean ± SD from three independent experiments. Asterisks indicate groups statistically significantly different from control by a 1-way ANOVA followed by a least-significant difference test (**P < 0.01).
To further confirm whether the phosphorylation of p38 MAPK was induced by glutamine starvation via the reduced GSH synthesis, PK-15 cell lysates were extracted to measure the rate of p38 phosphorylation after incubation for 72 h in the absence of glutamine or in the presence of glutamine and BSO. As shown in Figure 6, glutamine starvation increased the levels of p38 MAPK phosphorylation by 2.07-fold as compared with the controls treated with 4 mM glutamine (P < 0.01). In the presence of 4 mM glutamine, the levels of p38 MAPK phosphorylation in the groups with 20 or 50 μM BSO treatment increased by 2.42- and 2.50-fold, respectively, as compared with the controls (P < 0.01). The total p38 protein levels were not significantly different among all groups (P > 0.05). These results suggest that glutamine starvation and BSO treatment increased the level of p38 MAPK phosphorylation.

Glutamine starvation increases the phosphorylation of p38 MAPK via the reduced GSH synthesis. PK-15 cells lysates were extracted to measure the protein levels of p38 phosphorylation by Western blot after incubation for 72 h in the absence of glutamine or in the presence of glutamine and BSO (A and B). Values are shown as mean ± SD from three independent experiments. Asterisks indicate groups statistically significantly different from control by a 1-way ANOVA followed by least-significant difference test (**P < 0.01).
Knockdown of p38 MAPK decreased PCV2 replication but did not affect the level of GSH and MDA in PK-15 cells
The extent of p38 MAPK knockdown was evaluated by determining the relative p38 mRNA and protein levels after infected PK-15 cells were transfected with p38-specific or control siRNA. As shown in Figure 7, the transfection of p38-specific siRNA into PK-15 cells decreased the relative p38 mRNA and protein levels as compared with the controls. The relative p38 mRNA, p38 protein, and p-p38 protein levels were decreased by 57%, 50%, and 58%, after transfection (Figures 7A, 7B, and 7C; P < 0.01). Transfection of PK-15 cells with anti-p38 siRNA decreased PCV2 replication as measured by the number of PCV2 DNA copies and the proportion of infected cells. A significant decrease of approximately 11% in the PCV2 log 10 DNA copies per 106 cells (Figure 7D) and 25% in the proportion of infected cells (Figure 7E) was observed in p38-knockdown cells (P < 0.01). As shown in Figures 7F and 7G, the knockdown of p38 had no effect on the intracellular GSH levels and the MDA concentration (P > 0.05). Our results demonstrate that p38 MAPK knockdown decreases PCV2 replication.

Knockdown of p38 MAPK decreases PCV2 replication but does not affect the level of GSH and MDA in PK-15 cells. PCV2-infected cells were transfected with p38-specific or control siRNA. After 5 h of transfection treatment, the medium was removed, and fresh basal medium was added. After transfected-cells were cultured for another 72 h, the relative mRNA levels of p38 (A), the relative proteins levels of p38 and p-p38 (B and C), the number of PCV2 log 10 DNA copies per 106 cells (D), the relative proportion of infected cells (E), the level of GSH (F) and MDA (G) were assayed as described in Methods. Values are given as mean ± SD from three independent experiments. Groups were compared by a 1-way ANOVA followed by least-significant difference test. Significant changes are indicated by **(P < 0.01) and ## (P < 0.01) in comparison with controls.
PCV2 replication promoted by glutamine starvation could be blocked by p38 knockdown in PK-15 cells
The influence of glutamine starvation on PCV2 replication in p38-knockdown cells was evaluated to further investigate the role of p38 in the promotion of PCV2 replication by glutamine starvation. A significant increase of approximately 6% and 5% in the number of PCV2 log 10 DNA copies per 106 cells as well as approximately 33% and 25% in the proportion of infected cells in the groups treated with non-siRNA and control siRNA (P < 0.01; Figure 8A), respectively, but no increase was detected in groups treated with p38-specific siRNA (P > 0.05), as compared with the controls without glutamine starvation. Between the groups with glutamine starvation and siRNA treatment, the number of PCV2 log 10 DNA copies per 106 cells and the proportion of infected cells in groups treated with p38-specific siRNA decreased to normal levels as compared with groups treated with control siRNA. These results show that the promotion of PCV2 replication caused by glutamine starvation could be blocked in p38-knockdown cells.

PCV2 replication promoted by glutamine starvation could be blocked by p38 knockdown in PK-15 cells. PCV2-infected cells were transfected with p38-specific, control or non siRNA. After 5 h of transfection treatment, the medium was removed, and fresh basal medium with various concentrations of glutamine was added. After transfected-cells were cultured for another 72 h, the number of PCV2 log 10 DNA copies per 106 cells (A) and the relative proportion of infected cells (B) were assayed by real-time PCR and immunofluorescence microscopy, respectively. Values are given as mean ± SD from three independent experiments. Groups were compared by a 1-way ANOVA followed by least-significant difference test. Significant changes are indicated by **(P < 0.01) and # (P < 0.05) in comparison with controls.
Discussion
PCV2 infects pigs worldwide; this virus has been linked to several pathological conditions collectively named PCVD [30-32]. Not all pigs infected with PCV2 develop PCVD, although PCV2 is recognized as an essential infectious agent of PCVD [3]. The infection of pigs with PCV2 and other unknown triggers are required for PCVD to occur [3]. The evaluation of the role of these triggers is essential to understand the observed incidence of PCVD. Dietary glutamine supplementation may have an inhibitory effect on PCV2 infection [6,7]. The initial purpose of this study was to investigate the effects of glutamine supplementation on PCV2 replication and the underlying mechanisms in vitro. However, glutamine supplementation at concentrations of 4–16 mM did not affect PCV2 replication in PK-15 cells. Surprisingly, glutamine starvation significantly increased PCV2 replication (Figure 2; P < 0.05). Further studies targeting the effect of glutamine starvation on PCV2 replication in vitro and in vivo may improve the treatment of PCVD and the production of PCV2 vaccines.
Previous studies investigated the effect of glutamine on virus growth. Human cytomegalovirus-infected cells failed to produce viruses under glutamine deprivation [16]. Likewise, the growth of HVJ (Sendai virus) was significantly suppressed in glutamine-starved BHK cells, but the VSV and NDV viruses were not markedly affected by glutamine starvation [14]. In the present study, the effect of glutamine on PCV2 replication in PK-15 cells differed from that of the above mentioned viruses. Glutamine starvation induced the significant increase of PCV2 (P < 0.05), but the addition of fresh media with 4 mM glutamine blocked the PCV2 replication promoted by glutamine starvation. The results of the present study were consistent with our previous study, wherein the reduction of GSH levels and the induction of oxidative stress both increased PCV2 replication in PK-15 cells [26,27,33].
Glutamine has an essential role in promoting and maintaining cell growth and function in rapidly dividing cells because this amino acid is an important precursor of proteins, amino sugars, purines, and pyrimidines [34]. Glutamine metabolism is crucial to produce ATP and glutamate, which are necessary for glutathione synthesis [35]. Glutathione is a tripeptide that contains glutamate, cysteine, and glycine; this particularly important antioxidant protects cells from free radical injury [36]. In Jurkat T cells, glutamine increases intracellular GSH and decreases oxidative stress [37]. Glutamine deficiency triggers the decrease of cellular GSH levels and the promotion of oxidative stress in HuH-7 cells [17]. In addition, GSH supplementation decreases DV2 production in HepG2 cells [18]. In the present study, glutamine deprivation decreased the GSH levels but increased the MDA concentration and PCV2 replication (P < 0.05; Figure 4). Once GSH synthesis is inhibited by BSO, there is also a significant increase of PCV2 replication in PK-15 cells without glutamine starvation (Figure 4; P < 0.01). These results indicate that glutamine starvation may increase PCV2 replication by reducing intracellular GSH levels in PK-15 cells.
The activation of p38 MAPK is essential for efficient PCV2 replication [29]. Intracellular GSH has a regulatory role in p38 MAPK activation [11,12]. A complex relationship may exist between glutamine starvation, GSH levels, p38 MAPK activation, and PCV2 replication. We demonstrate that glutamine starvation and BSO treatment did not change the level of p38 mRNA (data not shown). However, the lack of glutamine caused the reduced GSH levels, increased p38 MAPK phosphorylation, and increased PCV2 replication in PK-15 cells (Figures 4 and 5). p38 MAPK phosphorylation and PCV2 replication was significantly decreased by p38 knockdown in PK-15 cells without glutamine starvation (P < 0.01). Moreover, the promotion of PCV2 replication by glutamine starvation could be blocked by p38 knockdown in PK-15 cells (Figures 7 and 8). These results indicate that glutamine starvation increased PCV2 replication through intracellular GSH downregulation, which was associated with the promotion of p38 MAPK activation.
Recent studies have revealed that glutamine starvation causes the activation of p38 MAPK and the deactivation of mTOR, which then induces autophagy [38-40]. Moreover, PCV2 induces autophagy by repressing mTOR in a cascade of phosphorylated proteins involving TSC2, ERK1/2, and AMPK [41] and enhances the replication of autophagic machinery in PK-15 cells [42]. Therefore, the role of autophagy in PCV2 replication during glutamine starvation is an interesting topic for future studies. The results obtained in the present study improve the understanding of PCV2 pathogenesis.
In the present study, glutamine starvation affected PK-15 cell survival. To elucidate the exact mechanism by which glutamine starvation influences PCV2 replication, a different cell line would be preferred, whose survival is not affected by glutamine starvation. However, only a few available cell lines are permissive to PCV2 infection. Additionally, glutamine starvation affects PK-15 cells and other cell lines because glutamine is required for the growth and survival of almost all cells [43,44]. A cell line that is not affected by glutamine starvation is difficult to find. Therefore, we believe that a cell line that is suitable for PCV2 infection but not affected by glutamine starvation would be impossible to find. The present results are reasonable because previous studies have reported that glutamine supplementation can decrease PCV2 infection [6,7]. In addition, glutamine starvation can induce oxidative stress [25,35], whereas oxidative stress can improve PCV2 replication [26,27,33].
In conclusion, our results show that PCV2 replication increases under conditions of lower glutamine concentration (less than 4 mM). Glutamine starvation enhances PCV2 replication in PK-15 cells via the phosphorylation of p38 MAPK, which is related to the reduction of intracellular GSH levels. Our findings contribute towards the interpretation of the possible pathogenic mechanism of PCV2 and provide a theoretical reference for the application of glutamine in the control of PCVD.
References
Segalés J, Allan GM, Domingo M (2005) Porcine circovirus diseases. Anim Health Res Rev 6:119–142
Opriessnig T, Meng XJ, Halbur PG (2007) Porcine circovirus type 2 associated disease: update on current terminology, clinical manifestations, pathogenesis, diagnosis, and intervention strategies. J Vet Diagn Invest 19:591–615
Grau-Roma L, Fraile L, Segalés J (2011) Recent advances in the epidemiology, diagnosis and control of diseases caused by porcine circovirus type 2. Vet J 187:23–32
Curi R, Newsholme P, Procopio J, Lagranha C, Gorjão R, Pithon-Curi TC (2007) Glutamine, gene expression, and cell function. Front Biosci 12:344–357
Lacey JM, Wilmore DW (2009) Is glutamine a conditionally essential amino acid? Nutr Rev 48:297–309
Ren W, Li Y, Yu X, Luo W, Liu G, Shao H, Yin Y (2013) Glutamine modifies immune responses of mice infected with porcine circovirus type 2. Br J Nutr 110:1053–1060
Ren W, Luo W, Wu M, Liu G, Yu X, Fang J, Li T, Yin Y, Wu G (2013) Dietary L-glutamine supplementation improves pregnancy outcome in mice infected with type-2 porcine circovirus. Amino Acids 45:479–488
Roth E, Oehler R, Manhart N, Exner R, Wessner B, Strasser E, Spittler A (2002) Regulative potential of glutamine—relation to glutathione metabolism. Nutrition 18:217–221
Fang YZ, Yang S, Wu G (2002) Free radicals, antioxidants, and nutrition. Nutrition 18:872–879
Haynes TE, Li P, Li X, Shimotori K, Sato H, Flynn NE, Wang J, Knabe DA, Wu G (2009) L-Glutamine or L-alanyl-L-glutamine prevents oxidant- or endotoxin-induced death of neonatal enterocytes. Amino Acids 37:131–142
Nadeem A, Siddiqui N, Alharbi NO, Alharbi MM, Imam F (2014) Acute glutathione depletion leads to enhancement of airway reactivity and inflammation via p38MAPK-iNOS pathway in allergic mice. Int Immunopharmacol 22:222–229
Jang HJ, Kim YM, Tsoyi K, Park EJ, Lee YS, Kim HJ, Lee JH, Joe Y, Chung HT, Chang KC (2012) Ethyl pyruvate induces heme oxygenase-1 through p38 mitogen-activated protein kinase activation by depletion of glutathione in RAW 264.7 cells and improves survival in septic animals. Antioxid Redox Signal 17:878–889
Bader JP, Morgan HR (1958) Latent viral infection of cells in tissue culture. VI. Role of amino acids, glutamine, and glucose in psittacosis virus propagation in L cells. J Exp Med 108:617–630
Ito Y, Kimura Y, Nagata I, Kunii A (1974) Effects of l-glutamine deprivation on growth of HVJ (Sendai virus) in BHK cells. J Virol 13:557–566
Bringhurst RM, Dominguez AA, Schaffer PA (2008) Glutamine deprivation causes enhanced plating efficiency of a herpes simplex virus type 1 ICP0-null mutant. J Virol 82:11472–11475
Chambers JW, Maguire TG, Alwine JC (2010) Glutamine metabolism is essential for human cytomegalovirus infection. J Virol 84:1867–1873
Xu Y, Nguyen Q, Lo DC, Czaja MJ (1997) c-myc-dependent hepatoma cell apoptosis results from oxidative stress and not a deficiency of growth factors. J Cell Physiol 170:192–199
Tian Y, Jiang W, Gao N, Zhang J, Chen W, Fan D, Zhou D, An J (2010) Inhibitory effects of glutathione on dengue virus production. Biochem Biophys Res Commun 397:420–424
Wirén M, Magnusson KE, Larsson J (1998) The role of glutamine, serum and energy factors in growth of enterocyte-like cell lines. Int J Biochem Cell Biol 30:1331–1336
Pan Q, Huang K, He K, Lu F (2008) Effect of different selenium sources and levels on porcine circovirus type 2 replication in vitro. J Trace Elem Med Biol 22:143–148
Valentin A, Rosati M, Patenaude DJ, Hatzakis A, Kostrikis LG, Lazanas M, Wyvill KM, Yarchoan R, Pavlakis GN (2002) Persistent HIV-1 infection of natural killer cells in patients receiving highly active antiretroviral therapy. Proc Natl Acad Sci U S A 99:7015–7020
Whitney JB, Hill AL, Sanisetty S, Penaloza-MacMaster P, Liu J, Shetty M, Parenteau L, Cabral C, Shields J, Blackmore S, Smith JY, Brinkman AL, Peter LE, Mathew SI, Smith KM, Borducchi EN, Rosenbloom DI, Lewis MG, Hattersley J, Li B, Hesselgesser J, Geleziunas R, Robb ML, Kim JH, Michael NL, Barouch DH (2014) Rapid seeding of the viral reservoir prior to SIV viraemia in rhesus monkeys. Nature 512:74–77
Rahman I, Kode A, Biswas SK (2006) Assay for quantitative determination of glutathione and glutathione disulfide levels using enzymatic recycling method. Nat Protoc 1:3159–3165
Gavino VC, Miller JS, Ikharebha SO, Milo GE, Cornwell DG (1981) Effect of polyunsaturated fatty acids and antioxidants on lipid peroxidation in tissue cultures. J Lipid Res 22:763–769
Guerin PJ, Furtak T, Eng K, Gauthier ER (2006) Oxidative stress is not required for the induction of apoptosis upon glutamine starvation of Sp2/0-Ag14 hybridoma cells. Eur J Cell Biol 85:355–365
Chen X, Ren F, Hesketh J, Shi X, Li J, Gan F, Huang K (2012) Selenium blocks porcine circovirus type 2 replication promotion induced by oxidative stress by improving GPx1 expression. Free Radic Biol Med 53:395–405
Chen X, Ren F, Hesketh J, Shi X, Li J, Gan F, Hu Z, Huang K (2013) Interaction of porcine circovirus type 2 replication with intracellular redox status in vitro. Redox Rep 18:186–192
Wu X, Bishopric NH, Discher DJ, Murphy BJ, Webster KA (1996) Physical and functional sensitivity of zinc finger transcription factors to redox change. Mol Cell Biol 16:1035–1046
Wei L, Zhu Z, Wang J, Liu J (2009) JNK and p38 mitogen-activated protein kinase pathways contribute to porcine circovirus type 2 infection. J Virol 83:6039–6047
Allan GM, Ellis JA (2000) Porcine circoviruses: a review. J Vet Diagn Invest 12:3–14
Allan GM, McNeilly F, Kennedy S, Daft B, Clarke EG, Ellis JA, Haines DM, Meehan BM, Adair BM (1998) Isolation of porcine circovirus-like viruses from pigs with a wasting disease in the USA and Europe. J Vet Diagn Invest 10:3–10
Gillespie J, Opriessnig T, Meng XJ, Pelzer K, Buechner-Maxwell V (2009) Porcine circovirus type 2 and porcine circovirus-associated disease. J Vet Intern Med 23:1151–1163
Chen X, Ren F, Hesketh J, Shi X, Li J, Gan F, Huang K (2012) Reactive oxygen species regulate the replication of porcine circovirus type 2 via NF-kappaB pathway. Virology 426:66–72
Smith RJ (1990) Glutamine metabolism and its physiologic importance. J Parenter Enteral Nutr 14(4 Suppl):40S–44S
Klimberg VS, McClellan JL (1996) Glutamine, cancer, and its therapy. Am J Surg 172:418–424
Mytilineou C, Kramer BC, Yabut JA (2002) Glutathione depletion and oxidative stress. Parkinsonism Relat Disord 8:385–387
Chang W-K, Yang KD, Chuang H, Jan J-T, Shaio M-F (2002) Glutamine protects activated human T cells from apoptosis by up-regulating glutathione and Bcl-2 levels. Clin Immunol 104:151–160
Chen R, Zou Y, Mao D, Sun D, Gao G, Shi J, Liu X, Zhu C, Yang M, Ye W, Hao Q, Li R, Yu L (2014) The general amino acid control pathway regulates mTOR and autophagy during serum/glutamine starvation. J Cell Biol 206:173–182
Harnett CC, Guerin PJ, Furtak T, Gauthier ER (2013) Control of late apoptotic events by the p38 stress kinase in L-glutamine-deprived mouse hybridoma cells. Cell Biochem Funct 31:417–426
Lin TC, Chen YR, Kensicki E, Li AY, Kong M, Li Y, Mohney RP, Shen HM, Stiles B, Mizushima N, Lin LI, Ann DK (2012) Autophagy: resetting glutamine-dependent metabolism and oxygen consumption. Autophagy 8:1477–1493
Zhu B, Zhou Y, Xu F, Shuai J, Li X, Fang W (2012) Porcine circovirus type 2 induces autophagy via the AMPK/ERK/TSC2/mTOR signaling pathway in PK-15 cells. J Virol 86:12003–12012
Zhu B, Xu F, Li J, Shuai J, Li X, Fang W (2012) Porcine circovirus type 2 explores the autophagic machinery for replication in PK-15 cells. Virus Res 163:476–485
Larson SD, Li J, Chung DH, Evers BM (2007) Molecular mechanisms contributing to glutamine-mediated intestinal cell survival. Am J Physiol Gastrointest Liver Physiol 293:G1262–G1271
Crespo JL, Powers T, Fowler B, Hall MN (2002) The TOR-controlled transcription activators GLN3, RTG1, and RTG3 are regulated in response to intracellular levels of glutamine. Proc Natl Acad Sci U S A 99:6784–6789
Acknowledgments
This work was funded by the National Natural Science Foundation of China (NFSC) (31272627 and 31472252), the Natural Science Foundation of Jiangsu Province (BK20131323), and the Priority Academic Program Development of Jiangsu Higher Education Institutions (Jiangsu, China). The authors declare no financial conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
XC carried out some of the experiments, participated in the analysis of the data and drafted the manuscript. XS carried out most of the experiments. FG was involved in the Western blot preparation and analysis. XS and FG participated in the design of some experiments and helped to draft the manuscript. DH participated in the analysis of the data and revised the manuscript. KH participated in its design and coordination, analyzed the data and revised the manuscript. All authors read and approved the final manuscript.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

Cite this article
Chen, X., Shi, X., Gan, F. et al. Glutamine starvation enhances PCV2 replication via the phosphorylation of p38 MAPK, as promoted by reducing glutathione levels. Vet Res 46, 32 (2015). https://doi.org/10.1186/s13567-015-0168-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13567-015-0168-1