
Abstract
Background
Angiogenesis is of utmost importance for tissue regeneration and repair. Human dental pulp stromal cells (hDPSCs) possess angiogenic potential, as they secrete paracrine factors that may alter the host microenvironment. However, more insight into how hDPSCs guide endothelial cells (ECs) in a paracrine fashion is yet to be obtained. Therefore, the current study aimed to investigate the effect(s) of conditioned medium derived from hDPSCs (hDPSC-CM) on EC behavior in vitro.
Methods
hDPSCs were harvested from third molars scheduled for surgical removal under informed consent. The angiogenic profile of hDPSC-CM was identified using human angiogenesis antibody array and enzyme-linked immunosorbent assay (ELISA). Using real-time reverse transcription-polymerase chain reaction (RT-PCR) and ELISA, the mRNA and protein expression level of specific angiogenic biomarkers was determined in human umbilical vein endothelial cells (HUVECs) exposed to hDPSC-CM. The effect of hDPSC-CM on HUVEC attachment, proliferation and migration was evaluated by crystal violet staining, MTT, transwell migration along with real-time cell monitoring assays (xCELLigence; ACEA Biosciences, Inc.). A Matrigel assay was included to examine the influence of hDPSC-CM on HUVEC network formation. Endothelial growth medium (EGM-2) and EGM-2 supplemented with hDPSC-CM served as experimental groups, whereas endothelial basal medium (EBM-2) was set as negative control.
Results
A wide range of proangiogenic and antiangiogenic factors, including vascular endothelial growth factor, tissue inhibitor of metalloproteinase protein 1, plasminogen activator inhibitor (serpin E1), urokinase plasminogen activator and stromal cell-derived factor 1, was abundantly detected in hDPSC-CM by protein profiling array and ELISA. hDPSC-CM significantly accelerated the adhesion phases, from sedimentation to attachment and spreading, the proliferation rate and migration of HUVECs as shown in both endpoint assays and real-time cell analysis recordings. Furthermore, Matrigel assay demonstrated that hDPSC-CM stimulated tubulogenesis, affecting angiogenic parameters such as the number of nodes, meshes and total tube length.
Conclusions
The sustained proangiogenic and promaturation effects of hDPSC-CM shown in this in vitro study strongly suggest that the trophic factors released by hDPSCs are able to trigger pronounced angiogenic responses, even beyond EGM-2 considered as an optimal culture condition for ECs.
Similar content being viewed by others


Background
The dental pulp, whose main function is to maintain tooth homeostasis, is often susceptible to trauma and bacterial insults [1]. The integrity of the pulp tissue is threatened during pulpal damage and inflammation, which in turn can negatively affect healing and tissue repair in this low-compliant microenvironment lacking collateral blood supply [2].
Regenerative endodontics, an emerging domain of tissue engineering, aims at renewing damaged/lost pulp tissue, implementing principles of cell biology and engineering rather than conventional endodontic treatments [3]. The discovery of postnatal dental pulp progenitor cells, exhibiting similar properties to bone marrow mesenchymal stem cells (MSCs) [4], has encouraged attempts to regenerate the damaged pulp tissue and/or maintain its functional integrity [2, 3]. These precursor cells have high regenerative potential when activated in the damaged pulp and exert paracrine/trophic effects that can alter the host microenvironment [5].
Successful pulp tissue regeneration relies on rapid development of a local microvascular network, providing sufficient oxygen and nutrients to the cells, particularly after tissue injury [6, 7]. The blood vessel network in dental pulp, as elsewhere in the body, is dependent on highly orchestrated angiogenic signaling mechanisms under normal as well as pathological conditions [8]. Angiogenesis, defined as sprouting of new capillaries from preexisting blood vessels [9], plays a pivotal role in wound healing and tissue repair [10]. Angiogenesis is a complex dynamic process, regulated by a sequence of molecular and cellular interactions involving basement membrane and extracellular matrix (ECM) degradation, endothelial cell proliferation, migration, tube formation and maturation into functional blood vessels. A balanced interplay of proangiogenic and antiangiogenic signaling cues, such as matrix metalloproteinases, growth factors, enzymes, cytokines, chemokines, and adhesion molecules, is required during blood vessel formation and development [9].
Human dental pulp stromal cells (hDPSCs) are highly angiogenic and have the potential to induce tissue vascularization via at least two distinct mechanisms; either by secreting angiogenic factors (paracrine effect) that enhance vascularization by local endothelial cells, or by differentiating into vascular endothelium via a process mimicking developmental vasculogenesis [11]. Recently, a growing number of studies have reported that DPSCs release proangiogenic and antiangiogenic proteins under different culture conditions, affecting different steps in the angiogenic process [12]. hDPSCs are also capable of promoting tube formation in human umbilical vein endothelial cells (HUVECs) both in a paracrine fashion and in a coculture system in vitro [5]. In agreement with these studies, we have shown that local administration of secretome from hDPSCs, grown under hypoxic conditions, promoted bone healing during distraction osteogenesis and reduced healing time through blood vessel formation [13].
Despite the previously demonstrated proangiogenic effects of human dental pulp cells (hDPCs) both in vitro and in vivo, the results are to some extent contradictory. Detailed cellular mechanisms by which hDPSCs exert their paracrine angiogenic effects and their potential therapeutic applications in dental tissue engineering are yet to be elucidated. As the angiogenesis is dependent on coordinated signaling by several soluble factors and the effects of growth factors that exert little or no action alone can be greatly augmented by other growth factors [14, 15], studies concerned with the effects of single soluble factors are insufficient to fully address angiogenesis. Therefore, the present study evaluated the angiogenic properties of hDPSC conditioned medium (hDPSC-CM), as a cocktail of soluble factors, by identifying its angiogenic protein profile and ability to affect functional aspects of angiogenesis in vitro.
Methods
Cell isolation and maintenance
hDPSCs were isolated as described previously [4]. Briefly, human adult third molars, scheduled for routine extraction, were collected from healthy patients aged 18–25 years under informed consent, in accordance with the protocol approved by the Ethical Research Committee at the University of Bergen, Norway (2009/610REK vest). hDPSCs were isolated from pulp tissue by enzymatic dissociation. Single-cell suspension was cultured and maintained in Dulbecco’s modified Eagle’s medium (DMEM; PAA, GE Healthcare, Little Chalfont, UK) supplemented with 10% fetal bovine serum (FBS), 4 mM l-glutamine, 100 U/ml penicillin and 100 μg/ml streptomycin. Cells at passages 3–8 were used for all experiments. hDPSCs from different donors were phenotypically characterized using flow cytometry as described previously by Al-Sharabi et al. [16].
Human umbilical vein endothelial cells (HUVECs) were purchased from Lonza (Clonetics, Walkersville, MD, USA) and expanded in Endothelial Growth Medium-2 (EGM-2) (Lonza). Cells below passage 5 were used for further experiments.
Collection of conditioned medium from hDPSCs
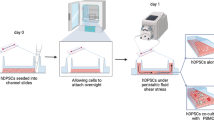
hDPSCs were seeded at a density of 5000–6000 cells/cm2 in culture flasks and grown until approximately 70–80% confluent. Cells were washed three to five times with PBS and incubated with freshly added serum-free DMEM containing penicillin–streptomycin for 48 h at 37 °C in 5% CO2. Supernatant was then collected, centrifuged at 4 °C at 3000 × g for 3 min followed by 5 min at 1500 × g, filtered through 0.2-μm filters and stored in aliquots at −80 °C as hDPSC-CM [13].
Angiogenic protein profile of CM
hDPSC-CM derived from at least three donors was screened for the relative expression of 55 different angiogenic proteins by Human Angiogenesis Antibody Array kit (Proteome profiler™; R&D Systems, Minneapolis, MN, USA). Briefly, hDPSC-CM was mixed with a detection antibody cocktail and then incubated with a nitrocellulose membrane spotted with capture antibodies in duplicate. Protein detection antibodies were visualized using streptavidin–HRP and chemiluminescent detection reagents. The Gel Doc EZ System (Bio-Rad Laboratories, CA, USA) was used for imaging. Positive signals (mean dot pixel density) on developed membranes were quantified using Quality One image analysis software version 4.7 (Bio-Rad Laboratories).
The expression of selected angiogenic factors in collected hDPSC-CM was quantified using ELISA. Vascular endothelial growth factor A (VEGF-A), stromal cell-derived factor 1 (SDF-1) and insulin-like growth factor 1 (IGF-1) ELISA kits were used according to the manufacturer’s protocol (R&D Systems). Absorbance was measured with a microplate reader (BMG Labtech, Ortenberg, Germany).
Cell morphology and attachment
Endothelial growth medium (EGM-2) was, based on pilot experiments, set as the optimal culture condition for HUVECs. A dilution of 25% and 50% v/v hDPSC-CM in EGM-2 (referred to as 25% CM and 50% CM) was used to evaluate its angiogenic properties. Endothelial basal medium (EBM-2) containing 2% FBS and GA-1000 (Gentamicin, Amphotericin-B) served as negative control in all experiments.
HUVECs were seeded in 96-well plates and allowed to attach for 15, 30, 45 and 60 min at 37 °C in 5% CO2. Attached cells were fixed with 4% paraformaldehyde (PFA) and stained with 0.1% crystal violet. Cell morphology was examined with an inverted optical microscope (Nikon Eclipse Ti, Tokyo, Japan). The cell number and cell surface area (freehand drawing) were quantified using ImageJ software (NIH, Bethesda, MD, USA) [17].
Adhesion kinetics and proliferation
After cell titration, the optimum cell density was chosen as 3000 cells/well. Thereafter, the effect of hDPSC-CM on viability, adhesion and proliferation of endothelial cells was assessed in real-time by the xCELLigence system E-plate 16 (ACEA Biosciences, Inc.) for 72 h at 15 min intervals and the cell status was expressed as the cell index (CI), as described previously [18]. Cell adhesion kinetics and proliferative responses were determined by calculating the mean CI value and slope of the line between two defined time points.
A 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT; Sigma Chemicals) assay was performed to validate the previous results in an endpoint experiment. Briefly, HUVECs were seeded at a density of approximately 9000 cells/cm2 in flat-bottom 96-well plates in four different culture media. At 24, 48 and 72 h, cells were washed with PBS and incubated with 100 μl MTT working solution (Sigma Chemicals) for 4 h at 37 °C. Then an equal volume of DMSO containing 6.25% (v/v) 0.1 M NaOH was added to dissolve the formed formazan for 20 min at room temperature. The optical density was measured at a wavelength of 570 nm with a FLUOstar Omega reader (BMG Labtech).
Real-time reverse transcriptase polymerase chain reaction
Total RNA was extracted from cultures at 2, 3 and 7 days using a Cell RNA isolation kit (Maxwell®; Promega, Madison, WI, USA) and reverse transcribed using a high-capacity cDNA reverse transcription kit (Applied Biosystems, Carlsbad, CA, USA). TaqMan gene expression assays (Applied Biosystems) were used to detect mRNA levels of glyceraldehyde-3-phosphate dehydrogenase (GAPDH), antigen Ki67 (Ki67), proliferating cell nuclear antigen (PCNA), VEGF-A, angiopoietin 1 (Ang-1), angiopoietin 2 (Ang-2), platelet endothelial cell adhesion molecule 1 (PECAM-1/CD31), von Willebrand factor (vWF), platelet-derived growth factor (PDGF) and fibroblast growth factor (FGF) (Table 1). Data were analyzed by a comparative CT method with GAPDH as endogenous control.
Enzyme-linked immunosorbent assay
To further evaluate the real-time reverse transcriptase polymerase chain reaction (RT-PCR) results at the protein level, cell supernatants from each experimental group were collected at 2, 3 and 7 days. The expression of human VEGF and Ang-1 in supernatants was then quantified using ELISA kits according to the manufacturer’s protocol (R&D Systems). Absorbance was measured at 450 nm with a FLUOstar microplate reader (BMG Labtech).
Migration and microvascular network formation
Cell migration was monitored in real-time by the xCELLigence system CIM-plate 16 (ACEA Biosciences, Inc.) [18]. Each well of the lower chamber was filled with 25% CM or 50% CM as experimental groups, whereas EBM-2 containing 2% FBS and EGM-2 containing 10% FBS served as negative and positive controls, respectively. A total number of 30,000 HUVECs was added to each well of the upper chamber (PET microporous membrane, 96-well format) in EBM-2 containing 2% FBS. Using real-time cell analysis (RTCA) software, a run of 24 h with automatic readings every 15 min was programmed. CI curves were analyzed by calculating the mean value and slope between specific time points.
Endothelial cell migration was also evaluated in a Transwell system. The wells of flat-bottom 24-well plates were filled with experimental and control groups, as already described. Eventually, HUVECs were seeded at a density of 150,000 cells/cm2 on cell culture inserts (8-μm pore size, PET track-etched membrane, 24-well format; Falcon, BD, USA) in EBM-2 containing 2% FBS. Following 24 h of incubation, transmigrated HUVECs were fixed with 4% PFA and stained with DAPI. Five representative (fields) images per insert were taken with an inverted phase-contrast microscope (Nikon Eclipse Ti, Tokyo, Japan). The number of positively stained DAPI nuclei was counted manually and by ImageJ software (NIH) [17].
The effect of hDPSC-CM on tubulogenesis was examined in a tube formation assay [13]. HUVECs were cultured in four different culture conditions, at a density of 4.5 × 105/cm2 in flat-bottom 96-well plates coated with growth factor-reduced Matrigel basement membrane matrix (BD Biosciences, San Jose, CA, USA). Two representative pictures (40× magnification) per well were taken after 6–8 h with an inverted phase-contrast microscope (Nikon TS100, Tokyo, Japan). The number of nodes, meshes, segments and total tube length were quantified using ImageJ analyzer.
Statistical analysis
Continuous data are presented as the mean with standard error of the mean (SEM). Multiple comparisons were tested by one-way analysis of variance (ANOVA) followed by a post-hoc Tukey test using the SPSS Statistics version 21.0 (IBM). For real-time analyses of proliferation and migration, a generalized linear model, assuming normal distributed data for comparisons of mean levels and slope, was set up. These analyses were performed using Stata version 14 (StataCorp), and robust variance estimates, adjusting for the repeated/clustered nature of the data, were applied. Statistical significance was determined at p < 0.05. All experiments were performed three times in four replicates and verified by at least three hDPSC donors.
Results
Proangiogenic and antiangiogenic proteins secreted by hDPSCs
Thirteen proangiogenic and antiangiogenic proteins were identified in hDPSC-CM (Fig. 1a, b). Tissue inhibitor of metalloproteinase 1 (TIMP-1), plasminogen activator inhibitor (PAI-1)/serpin E1, urokinase plasminogen activator (uPA), insulin-like growth factor binding protein 2 and 3 (IGFBP-2 and 3), pentraxin 3, serpin F-1 and VEGF (among all) were abundantly secreted in hDPSC-CM. No positive detection was found in DMEM, serving as control (data not shown). ELISA revealed high expression of VEGF-A and SDF-1 in hDPSC-CM, whilst IGF-1 was not detected (Fig. 1c). The expression level of detected proteins was highest in hDPSC-CM derived from passage 4 and declined gradually up to passage 8 (data not shown). Consequently, hDPSC-CM from passages 4–6 was used in all experiments.

Angiogenic protein profile of human dental pulp stromal cell conditioned medium (hDPSC-CM). a Proangiogenic and antiangiogenic proteins detected by human angiogenesis antibody array, marked 1–13 on the array membrane. b Histogram generated by quantification of mean spot pixel density using image analysis software. Number on each column corresponds to the protein spotted on the membrane in (a). c ELISA results showing concentration of vascular endothelial growth factor (VEGF), stromal cell-derived factor 1 (SDF-1) and insulin-like growth factor 1 (IGF-1) in hDPSC-CM. Data presented as mean ± standard deviation. ND not detected, HGF hepatocyte growth factor, IGFBP-2,3 insulin-like growth factor binding protein 2 and 3, MMP-9 matrix metalloproteinase 9, TIMP-1 tissue inhibitor of metalloproteinase 1, uPA urokinase plasminogen activator
hDPSC-CM enhanced HUVEC adhesion and proliferation
Morphological changes, including initial bond formations and subsequent spreading, were observed to happen faster in response to hDPSC-CM compared with other groups (Fig. 2a). The number of adhered cells as well as the cell spreading area remained higher in the CM groups at all time points, with significant values measured at 45 and 60 min, as shown in crystal violet staining assay (Fig. 2b–d). The CI curves based on real-time xCELLigence readings also confirmed that adhesion steps in HUVECs incubated with CM proceeded more than twice as fast as the controls within the first hour of cell monitoring (Fig. 2e–g).

Human umbilical vein cord endothelial cell (HUVEC) morphology, attachment, spreading and adhesion kinetics. a Representative crystal violet staining images of HUVECs grown under different culture conditions and time points. b–d Graphs from image analysis of attached HUVECs showing cell number (b), mean cell area (c) and total cell area (d). e Cell adhesion kinetics monitored in real time using xCELLigence system reveal faster increase in mean cell index (CI) in HUVECs exposed to human dental pulp stromal cell conditioned medium (hDPSC-CM). The effect was independent of concentration and happened within minutes after seeding. f, g Representative graphs comparing HUVEC attachment, showing difference in mean CI value (f) and slope of the line (g) between each selected time period (0–1, 1–3 and 3–6 h). Endothelial growth medium (EGM-2) group served as reference for comparison. Data presented as mean ± SEM. *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001, n = 4. EBM-2 Endothelial Basal Medium-2, CM conditioned medium, EGM-2 Endothelial Growth Medium-2
RTCA data, collected over a period of 72 h, showed that HUVECs proliferated significantly faster when grown in the presence of hDPSC-CM compared with controls (Fig. 3a–c). Consistently, CM significantly enhanced the proliferation rate of HUVECs at 72 h as shown by MTT assay (Fig. 3d). However, there was no difference in growth patterns and cell behavior among the CM groups.

Human umbilical vein cord endothelial cell (HUVEC) proliferation. a Cell proliferative activity monitored in real time using xCELLigence system. b, c Representative graphs showing difference in mean cell index value (b) and slope of the line (c) between each selected time period (12–24, 24–48 and 48–72 h). HUVECs treated with human dental pulp stromal cell conditioned medium (hDPSC-CM), independent of concentration, proliferated significantly faster than other groups at all time periods. d MTT assay showing significantly enhanced proliferation of HUVECs in hDPSC-CM compared with EGM-2 at 3 days. e, f mRNA expression of the proliferative biomarkers antigen KI-67 (Ki67) and proliferating cell nuclear antigen (PCNA) in HUVECs at 2, 3 and 7 days. Endothelial growth medium (EGM-2) group served as reference for comparison. Data presented as mean ± SEM. *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001, n = 4. EBM-2 Endothelial Basal Medium-2, CM conditioned medium, EGM-2 Endothelial Growth Medium-2, MTT 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide, d days
At the gene level, the mRNA expression of genes associated with proliferation (Ki67 and PCNA) was significantly upregulated in HUVECs exposed to CM (independent of CM concentration) at 2 and 7 days (Fig. 3e, f).
hDPSC-CM altered angiogenic gene expression in HUVECs
The expression of genes encoding vascular development, including VEGF-A, Ang-2, FGF and PECAM-1 was generally decreased at all time points in HUVECs exposed to hDPSC-CM compared with EGM-2 (the higher the concentration of CM, the lower the mRNA expression) (Fig. 4a–d). In contrast, the mRNA expression levels of Ang-1, vWF and PDGF were increased in CM groups compared with EGM-2 at 2 and 3 days, whereas a significant downregulation for the same genes was observed in CM groups at 7 days (Fig. 4e–g).

Real-time reverse transcription-polymerase chain reaction (RT-PCR). a–g Representative graphs showing expression of the angiogenesis-related genes vascular endothelial growth factor A (VEGF-A) (a), angiopoietin 2 (Ang-2) (b), platelet endothelial cell adhesion molecule 1 (PECAM-1) (c), fibroblast growth factor (FGF) (d), angiopoietin 1 (Ang-1) (e), von Willebrand factor (vWF) (f) and platelet-derived growth factor (PDGF) (g) in human umbilical vein cord endothelial cells (HUVECs) grown under different culture conditions at 2, 3 and 7 days. Data normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) as housekeeping gene and presented as mean ± SEM. *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001, n = 4. CM conditioned medium, EGM-2 Endothelial Growth Medium-2, d days
hDPSC-CM altered angiogenic protein expression in HUVECs
The expression of VEGF was reduced in the groups exposed to CM at 2, 3 and 7 days, following the same pattern as found at gene level (Fig. 5a). Ang-1 expression was consistently and significantly higher in CM groups compared to EGM-2, with 50% CM showing the highest expression level at all time points (Fig. 5b).

Enzyme linked immunosorbent assay (ELISA). a, b Representative graph showing expression of vascular endothelial growth factor A (VEGF) (a) and angiopoietin 1 (Ang-1) (b). Data presented as mean ± standard deviation. *p ≤ 0.05, ***p ≤ 0.001, n = 4. d days
hDPSC-CM stimulated HUVEC migration and microvascular network formation
The CI curves obtained from the xCELLigence platform indicated that HUVECs migrated almost twice as fast toward the hDPSC-CM compared with EGM-2 (Fig. 6a–c). Transwell migration assay also showed that more (although not significant) HUVECs migrated toward CM compared with EGM-2 at 24 h. As expected, positive control containing 10% FBS attracted a significantly higher number of cells through the inserts (Fig. 6d, e).

Human umbilical vein cord endothelial cell (HUVEC) migration and microvascular network formation. a Representative graph based on real-time measurements showing HUVEC migration rate. b, c Graphs showing mean cell index value (b) and slope (c) between each selected time period (1–3, 6–12, 12–24 h). HUVECs transmigrated significantly faster toward human dental pulp stromal cell conditioned medium (hDPSC-CM) compared with endothelial growth medium (EGM-2), independent of time period and concentration. d DAPI staining of transmigrated HUVECs toward different culture conditions at 24 h (10% FBS EGM-2 used as positive control). e Graph representing mean migrated cell number. f Representative photomicrographs of HUVEC tube formation 6–8 h after incubation in the different culture conditions. g–j Graphs demonstrating tube formation parameters, including number of nodes (g), number of meshes (h), number of segments (i) and total tube length (j). Data presented as mean ± SEM. *p ≤ 0.05, **p ≤ 0.01, ***p ≤ 0.001, n = 4. Scale bar 200 μm. EBM-2 Endothelial Basal Medium-2, CM conditioned medium, EGM-2 Endothelial Growth Medium-2, FBS fetal bovine serum
hDPSC-CM and EGM-2 demonstrated increased endothelial tube formation (Fig. 6f), and network formation parameters were significantly higher compared with EBM-2. Although not significant, a slightly higher number of nodes, segments, meshes and total tube length was quantified in response to hDPSC-CM (Fig. 6g–j).
Discussion
Targeting therapeutic angiogenesis for pulp tissue regeneration and repair requires stimuli that favor an angiogenic environment. Angiogenesis is tightly regulated in a spatiotemporal manner, with a rigorous interplay between a variety of proangiogenic and antiangiogenic molecules [8, 19]. The role of hDPSC-CM in promotion or inhibition of vessel formation, including its potential effects on EC behavior, is of high importance for optimizing dental pulp regeneration. It is well documented that cultured MSCs secrete trophic and immunomodulatory factors including angiogenic, mitogenic, antiapoptotic, immunomodulatory, chemoattractant and extracellular matrix proteins [20, 21]. However, the secreted molecules and their concentrations differ according to cell type, culture condition and duration [22]. In line with previous reports, the present study clearly demonstrated that a wide range of angiogenic proteins were expressed in hDPSC-CM. Interestingly, regardless of concentration, hDPSC-CM showed the ability to promote EC adhesion, proliferation, migration and tubular network formation, indicating angiogenic stimulation.
Currently used endpoint assays for evaluating angiogenesis do not provide continuous real-time data for cell adhesion, proliferation and migration, and demand high user input [23]. In order to obtain more detailed information we used the highly accurate RCTA platform (xCELLigence RTCA DP, ACEA), facilitating a non-invasive label-free real-time monitoring of ECs dynamics. The interaction of cells with the microelectrodes integrated in the bottom of an E-plate generates an impedance response measured as the cell index (CI). The CI is an arbitrary value proportional to the number of cells, cell morphology and cell adhesion dynamics [18, 24]. In the current study, an exponential increase in the CI readout was observed within minutes when HUVECs were grown in the presence of CM derived from hDPSCs. This remained significantly higher during the first hour after cell seeding, suggesting that CM contains biofactors that enhance HUVEC adhesion kinetics. Cell adhesion and spreading were also visually evaluated and quantified using phase-contrast microscopy after crystal violet staining. Quantitative data showed significantly higher number and surface area for the cells treated with CM compared with cells in EGM-2 alone. Furthermore, changes in cellular morphology within the initial steps of the adhesion process happened earlier than and twice as fast as in the EGM-2 group. As ECM composition can alter endothelial cell spreading dynamics, including the rate and morphology changes [25], it is suggested that the soluble ECM proteins present in CM are responsible for the effects observed. Previous studies have shown that cells on high densities of ECM proteins tend to spread isotropically [26], and that MSC-CM promotes the adhesion ability of HUVECs, possibly through SDF-1 expressed by MSCs [27].
In an attempt to address whether hDPSC-CM could promote HUVEC proliferation and growth, a series of experiments were performed. HUVECs treated with CM showed a significant increase in CI over a period of 72 h, which correlated well with a statistical increase in cell proliferation. The mean CI and the slope of growth curves were calculated for all time periods and the CI for CM-treated cells, irrespective of concentration, remained higher than that for untreated cells. The metabolic activity levels, as measured by MTT, were also significantly higher in the CM groups at 72 h. In parallel, the mRNA levels of the cell proliferation markers Ki67 and PCNA were significantly upregulated in the cells exposed to CM at 48 h, suggesting that the increase in proliferative activity of HUVECs from genome to transcriptome happened in a coordinated temporal manner. The enhanced proliferation rate might be explained by the abundant release of mitogens and antiapoptotic factors, such as VEGF [28], angiopoietins [19], TIMP-1 [29] and uPA [30], and their possible synergistic effect(s) in hDPSC-CM. As shown previously, DPSCs secrete VEGF, a key regulatory factor of vascular permeability and angiogenesis, which enhances the survival and differentiation of ECs [31]. VEGF signaling also promotes EC proliferation, filopodial extensions and ECM degradation [9]. uPA enhances endothelial permeability through intracellular signaling pathways shared with VEGF [30, 32]. TIMP-1 has a well-established antiapoptotic activity in many different cell types including HUVECs and possesses a novel MMP-independent cytokine-like activity in the regulation of cell proliferation [29]. In addition, MSCs derived from bone marrow and adipose tissue have the capacity to increase EC proliferation [33,34,35]. In contrast, other studies have shown no effect of CM from hDPSCs and BM-MSCs on EC growth [12, 36]. A possible explanation could be insufficient serum concentration for cell survival in these experimental setups.
There is a body of evidence that ECs sense small alterations in the microenvironment and have a remarkable capacity to adjust to the local requirements, which in turn determines the direction of differentiation [37]. Vascular development is initiated by Ang-2 and VEGF promoting proliferation, migration and sprouting of ECs, whereas establishment of functional and stabilized vessels is facilitated by Ang-1 in a competitive Tie2 receptor binding with Ang-2 [38]. The expression of endothelial specific markers was altered in the presence of CM, with a downregulation of VEGF-A, Ang-2 and FGF at all time points. As further confirmed by ELISA, the extracellular VEGF secretion followed the same pattern as the gene expression. The substantial proliferation and survival of HUVECs for up to 3 days in culture might suggest a high level of antiapoptotic and mitogenic proteins present in CM, suppressing excessive gene expression in a feedback loop mechanism. In addition, increased permeability and nascent phenotype of ECs could be attributed to the downregulation of PECAM-1 in CM-treated cells. Proliferation and differentiation of cells are two processes that do not occur simultaneously. In order for nascent vessels to mature into tubes with controlled permeability, the proliferation of ECs should subside [39]. As discussed earlier, CM induced EC proliferation for a prolonged period. Thus, it is logic to speculate that the differentiation of cells was delayed accordingly. An upregulation of Ang-1 expression, observed at day 2, is possibly mediated through competitive receptor binding with Ang-2. Nevertheless, the significant increase in Ang-1 protein expression in HUVECs exposed to CM compared to EGM-2 is indicative of EC maturation or vascular stabilization at 3 and 7 days. The translational process and protein turnover can take some time, independent of the translation/degradation rate and half-life of different proteins, as the cells need to adapt to microenvironmental changes [40, 41]. This delay might explain possible discrepancies between gene and protein expression at 7 days for Ang-1.
A further key step in angiogenesis is the migration of proliferating ECs along a gradient of chemotactic stimuli through the disintegrated basement membrane and ECM in the remodeled perivascular space. Data from the RTCA chemotactically driven migration setup showed that HUVECs migrated significantly faster through microporous inserts toward the media derived from hDPSCs than EGM-2. Higher concentrations of CM (50% CM) provided the strongest chemotactic stimuli for the recruitment of HUVECs. Despite a higher number of migrated HUVECs toward CM than EGM-2 alone, as shown in the 24 h endpoint migration assay, the difference was not found to be significant. This suggests that RTCA provides a more accurate platform as the electrodes sense passing cells in real time by recording CI changes. On the other hand, abundant secretion of stromal derived factor 1 (SDF-1) and VEGF in hDPSC-CM in our experiments might be the key chemotactic stimuli for HUVEC migration [12, 42, 43]. SDF-1 is involved in hematopoietic and nonhematopoietic stem/progenitor cell migration and homing [43], and the SDF-1α–CXCR4 axis plays a crucial role in the recruitment of CXCR4-positive DPSCs toward the damaged sites [44]. VEGF is (partly) responsible for the chemotactic activities of hDPSCs, as shown by neutralizing VEGF in the CM from hDPSCs [12].
Tube-like structure and lumen formation by migrated ECs is critically important in healing and regeneration. The Matrigel tube formation assay indicated a higher vessel density and larger lumen areas in CM groups than those in EGM-2. Although the number of nodes, meshes and tube length were higher in groups exposed to CM, the results were not significant. However, studies have previously reported that CM from different sources of MSCs or their side populations have the ability to promote tube formation [5, 45, 46], mostly through VEGF signaling that plays a pivotal role in EC sprouting and tubular network formation. This was evidenced by a reduction in the tubular perimeters after addition of VEGF and/or FGF neutralizing antibodies [10].
Conclusion
It is important to note that hDPSCs, also defined as dental pulp fibroblasts, are a heterogeneous cell population that can regulate the pericellular microenvironment by secreting ECM, growth factors and growth factor binding proteins. The angiogenic/vasculogenic properties of hDPSCs are independent of direct contact and are mainly due to the paracrine activity of the secreted soluble factors. This in vitro study strongly demonstrated that CM released from hDPSCs is able to trigger pronounced angiogenic effects, even beyond EGM-2 considered to be an optimal culture condition for ECs. Therefore, the valuable effect of hDPSC-CM on angiogenesis either alone or in combination with EGM-2 might be a result of the additive or synergistic action of several growth factors. This synergistic effect of growth factors in CM is likely to be important in therapeutic applications requiring angiogenesis. As a better understanding of the dynamic and mechanisms underlying the effects of CM develops, a clearer role of CM in vascular development and maintenance evolves. However, thorough proteomic analyses to detect detailed concentrations of identified angiogenic proteins in hDPSC-CM and novel in vivo studies are warranted to strengthen this conclusion, and to evaluate the potential effect of CM in healing and regeneration.
Abbreviations
- Ang-1:
-
Angiopoietin 1
- Ang-2:
-
Angiopoietin 2
- CI:
-
Cell index
- CM:
-
Conditioned medium
- DMEM:
-
Dulbecco’s Modifies Eagle’s Medium
- DMSO:
-
Dimethyl sulfoxide
- EBM-2:
-
Endothelial Basal Medium-2
- EC:
-
Endothelial cell
- ECM:
-
Extracellular matrix
- EGM-2:
-
Endothelial Growth Medium-2
- ELISA:
-
Enzyme-linked immunosorbent assay
- FBS:
-
Fetal bovine serum
- FGF:
-
Fibroblast growth factor
- GAPDH:
-
Glyceraldehyde-3-phosphate dehydrogenase
- hDPSC:
-
Human dental pulp stromal cell
- HUVEC:
-
Human umbilical vein endothelial cell
- IGF-1:
-
Insulin-like growth factor 1
- IGFBP-2:
-
3
Insulin-like growth factor binding protein 2 and 3
- Ki67:
-
Antigen KI-67
- MMP-9:
-
Matrix metalloproteinase 9
- MSC:
-
Mesenchymal stem cell
- PAI-1:
-
Plasminogen activator inhibitor
- PCNA:
-
Proliferating cell nuclear antigen
- PDGF:
-
Platelet-derived growth factor
- PECAM-1:
-
Platelet endothelial cell adhesion molecule 1
- PFA:
-
Paraformaldehyde
- RTCA:
-
Real-time cell analysis
- RT-PCR:
-
Real-time reverse transcriptase polymerase chain reaction
- SDF-1:
-
Stromal-derived factor 1
- TIMP-1:
-
Tissue inhibitor of metalloproteinase 1
- uPA:
-
Urokinase plasminogen activator
- VEGF:
-
Vascular endothelial growth factor
- vWF:
-
Von Willebrand factor
References
Nakashima M, Iohara K. Regeneration of dental pulp by stem cells. Adv Dent Res. 2011;23:313–9.
Huang GT. Dental pulp and dentin tissue engineering and regeneration: advancement and challenge. Front Biosci (Elite Ed). 2011;3:788–800.
Gong T, Heng BC, Lo EC, Zhang C. Current advance and future prospects of tissue engineering approach to dentin/pulp regenerative therapy. Stem Cells Int. 2016;2016:1–13.
Gronthos S, Mankani M, Brahim J, Robey PG, Shi S. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc Natl Acad Sci U S A. 2000;97:13625–30.
Tran-Hung L, Mathieu S, About I. Role of human pulp fibroblasts in angiogenesis. J Dent Res. 2006;85:819–23.
Laschke MW, Harder Y, Amon M, Martin I, Farhadi J, Ring A, Torip-Padron N, Schramm R, Rucker M, Junker D, et al. Angiogenesis in tissue engineering: breathing life into constructed tissue substitutes. Tissue Eng. 2006;12:2093–104.
Rombouts C, Giraud T, Jeanneau C, About I. Pulp vascularization during tooth development, regeneration, and therapy. J Dent Res. 2017;96:137–44.
Bronckaers A, Hilkens P, Martens W, Gervois P, Ratajczak J, Struys T, Lambrichts I. Mesenchymal stem/stromal cells as a pharmacological and therapeutic approach to accelerate angiogenesis. PharmacolTher. 2014;143:181–96.
Herbert SP, Stainier DY. Molecular control of endothelial cell behaviour during blood vessel morphogenesis. Nat Rev Mol Cell Biol. 2011;12:551–64.
Tran-Hung L, Laurent P, Camps J, About I. Quantification of angiogenic growth factors released by human dental cells after injury. Arch Oral Biol. 2008;53:9–13.
Sieveking DP, Ng MK. Cell therapies for therapeutic angiogenesis: back to the bench. Vasc Med. 2009;14:153–66.
Bronckaers A, Hilkens P, Fanton Y, Struys T, Gervois P, Politis C, Martens W, Lambrichts I. Angiogenic properties of human dental pulp stem cells. PLoS One. 2013;8:e71104.
Fujio M, Xing Z, Sharabi N, Xue Y, Yamamoto A, Hibi H, Ueda M, Fristad I, Mustafa K. Conditioned media from hypoxic-cultured human dental pulp cells promotes bone healing during distraction osteogenesis. J Tissue Eng Regen Med. 2015;11:2116–26.
Castellon R, Hamdi HK, Sacerio I, Aoki AM, Kenney MC, Ljubimov AV. Effects of angiogenic growth factor combinations on retinal endothelial cells. Exp Eye Res. 2002;74:523–35.
Kim SK, Lee J, Song M, Kim M, Hwang SJ, Jang H, Park Y. Combination of three angiogenic growth factors has synergistic effects on sprouting of endothelial cell/mesenchymal stem cell-based spheroids in a 3D matrix. J Biomed Mater Res B Appl Biomater. 2016;104:1535–43.
Al-Sharabi N, Xue Y, Fujio M, Ueda M, Gjerde C, Mustafa K, Fristad I. Bone marrow stromal cell paracrine factors direct osteo/odontogenic differentiation of dental pulp cells. Tissue Eng Part A. 2014;20:3063–72.
Carpentier G. ImageJ contribution: Angiogenesis Analyzer. ImajeJ News. 2012; http://image.bio.methods.free.fr/ImageJ/?Angiogenesis-Analyzer-for-ImageJ. Accessed 3 Nov 2017.
Dowling CM, Ors CH, Kiely PA. Using real-time impedance-based assays to monitor the effects of fibroblast-derived media on the adhesion, proliferation, migration and invasion of colon cancer cells. Biosci Rep. 2014;34:415–U127. https://doi.org/10.1042/BSR20140031.
Distler JH, Hirth A, Kurowska-Stolarska M, Gay RE, Gay S, Distler O. Angiogenic and angiostatic factors in the molecular control of angiogenesis. Q J Nucl Med. 2003;47:149–61.
Sorrell JM, Baber MA, Caplan AI. Influence of adult mesenchymal stem cells on in vitro vascular formation. Tissue Eng Part A. 2009;15:1751–61.
Meirelles Lda S, Fontes AM, Covas DT, Caplan AI. Mechanisms involved in the therapeutic properties of mesenchymal stem cells. Cytokine Growth Factor Rev. 2009;20:419–27.
Dowling P, Clynes M. Conditioned media from cell lines: a complementary model to clinical specimens for the discovery of disease-specific biomarkers. Proteomics. 2011;11:794–804.
Staton CA, Stribbling SM, Tazzyman S, Hughes R, Brown NJ, Lewis CE. Current methods for assaying angiogenesis in vitro and in vivo. Int J Exp Pathol. 2004;85:233–48.
Abbasi Y. Label-free and dynamic monitoring of cell-based assays. Biochemica. 2008;3:8–11.
Khalili AA, Ahmad MR. A review of cell adhesion studies for biomedical and biological applications. Int J Mol Sci. 2015;16:18149–84.
Reinhart-King CA, Dembo M, Hammer DA. The dynamics and mechanics of endothelial cell spreading. Biophys J. 2005;89:676–89.
Zheng F, Li X, Zhang L, Kong X, Guo LY, Yang JY, Huang YZ, Tang JM, Wang JN. Effect of conditioned medium of mesenchymal stem cells on proliferation, migration and adhesion of human umbilical vein endothelial cells. Zhongguo Shi Yan Xue Ye Xue Za Zhi. 2012;20:154–8.
Beckermann BM, Kallifatidis G, Groth A, Frommhold D, Apel A, Mattern J, Salnikov AV, Moldenhauer G, Wagner W, Diehlmann A, et al. VEGF expression by mesenchymal stem cells contributes to angiogenesis in pancreatic carcinoma. Br J Cancer. 2008;99:622–31.
Chirco R, Liu XW, Jung KK, Kim HR. Novel functions of TIMPs in cell signaling. Cancer Metastasis Rev. 2006;25:99–113.
Stepanova V, Jayaraman PS, Zaitsev SV, Lebedeva T, Bdeir K, Kershaw R, Holman KR, Parfyonova YV, Semina EV, Beloglazova IB, et al. Urokinase-type plasminogen activator (uPA) promotes angiogenesis by attenuating proline-rich homeodomain protein (PRH) transcription factor activity and de-repressing vascular endothelial growth factor (VEGF) receptor expression. J Biol Chem. 2016;291:15029–45.
Grando Mattuella L, Poli de Figueiredo JA, Nor JE, de Araujo FB, Medeiros Fossati AC. Vascular endothelial growth factor receptor-2 expression in the pulp of human primary and young permanent teeth. J Endod. 2007;33:1408–12.
Behzadian MA, Windsor LJ, Ghaly N, Liou G, Tsai NT, Caldwell RB. VEGF-induced paracellular permeability in cultured endothelial cells involves urokinase and its receptor. FASEB J. 2003;17:752–4.
Rehman J, Traktuev D, Li J, Merfeld-Clauss S, Temm-Grove CJ, Bovenkerk JE, Pell CL, Johnstone BH, Considine RV, March KL. Secretion of angiogenic and antiapoptotic factors by human adipose stromal cells. Circulation. 2004;109:1292–8.
Potapova IA, Gaudette GR, Brink PR, Robinson RB, Rosen MR, Cohen IS, Doronin SV. Mesenchymal stem cells support migration, extracellular matrix invasion, proliferation, and survival of endothelial cells in vitro. Stem Cells. 2007;25:1761–8.
Kinnaird T, Stabile E, Burnett MS, Lee CW, Barr S, Fuchs S, Epstein SE. Marrow-derived stromal cells express genes encoding a broad spectrum of arteriogenic cytokines and promote in vitro and in vivo arteriogenesis through paracrine mechanisms. Circ Res. 2004;94:678–85.
Gruber R, Kandler B, Holzmann P, Vogele-Kadletz M, Losert U, Fischer MB, Watzek G. Bone marrow stromal cells can provide a local environment that favors migration and formation of tubular structures of endothelial cells. Tissue Eng. 2005;11:896–903.
Alberts B, Johnson A, Lewis J, Raff M, Robert K, Walter P. Specialized tissues, stem cells, and tissue renewal; blood vessel, lymphatics, and endothelial cells. In: Anderson M, Granum S, editors. Molecular biology of the cell. New York: Garland Science; 2002. p. 1445–50.
Gerhardt H. VEGF and endothelial guidance in angiogenic sprouting. Organ. 2008;4:241–6.
Kutcher ME, Herman IM. The pericyte: cellular regulator of microvascular blood flow. Microvasc Res. 2009;77:235–46.
Edfors F, Danielsson F, Hallström BM, Käll L, Lundberg E, Pontén F, Forsström B, Uhlén M. Gene-specific correlation of RNA and protein levels in human cells and tissues. Mol Syst Biol. 2016;12:883.
Vogel C, Marcotte EM. Insights into the regulation of protein abundance from proteomic and transcriptomic analyses. Nat Rev Genet. 2012;13:227–32.
Burlacu A, Grigorescu G, Rosca AM, Preda MB, Simionescu M. Factors secreted by mesenchymal stem cells and endothelial progenitor cells have complementary effects on angiogenesis in vitro. Stem Cells Dev. 2013;22:643–53.
Kucia M, Reca R, Miekus K, Wanzeck J, Wojakowski W, Janowska-Wieczorek A, Ratajczak J, Ratajczak MZ. Trafficking of normal stem cells and metastasis of cancer stem cells involve similar mechanisms: pivotal role of the SDF-1-CXCR4 axis. Stem Cells. 2005;23:879–94.
Jiang L, Zhu YQ, Du R, Gu YX, Xia L, Qin F, Ritchie HH. The expression and role of stromal cell-derived factor-1alpha-CXCR4 axis in human dental pulp. J Endod. 2008;34:939–44.
Ishizaka R, Hayashi Y, Iohara K, Sugiyama M, Murakami M, Yamamoto T, Fukuta O, Nakashima M. Stimulation of angiogenesis, neurogenesis and regeneration by side population cells from dental pulp. Biomaterials. 2013;34:1888–97.
Fanton Y, Houbrechts C, Willems L, Daniels A, Linsen L, Ratajczak J, Bronckaers A, Lambrichts I, Declercq J, Rummens JL, et al. Cardiac atrial appendage stem cells promote angiogenesis in vitro and in vivo. J Mol Cell Cardiol. 2016;97:235–44.
Acknowledgements
The authors acknowledge financial support by the University of Bergen, Norway.
Funding
The study was funded by the University of Bergen, Norway.
Availability of data and materials
Please contact author for data requests.
Author information
Authors and Affiliations
Contributions
MAG, YX, KM, SAL and IF conceived and designed the experiments. MAG performed the experiments. MAG, YX and SAL analyzed data. MAG, YX, KM and IF wrote the paper. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The protocol was reviewed and approved by the Ethical Research Committee at the University of Bergen, Norway (2009/610REK vest). The purpose of the study was explained to the patients and written informed consent was obtained.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Gharaei, M.A., Xue, Y., Mustafa, K. et al. Human dental pulp stromal cell conditioned medium alters endothelial cell behavior. Stem Cell Res Ther 9, 69 (2018). https://doi.org/10.1186/s13287-018-0815-3
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13287-018-0815-3