
Abstract
Purpose
Balanophora polyandra Griff. is a holoparasitic medicinal plant that produces compounds with antibacterial and antioxidant activities. Plant endophytic fungi are an abundant reservoir of bioactive metabolites for medicinal exploitation, and an increasing number of novel bioactive compounds are being isolated from endophytic fungi. The present study investigated the diversity of culturable endophytic fungi from the roots of holoparasite B. polyandra to explore active strains and metabolites. In addition, the antibacterial and antioxidant activities of 22 strains cultured from B. polyandra were also evaluated.
Methods
The endophytic fungi were identified according to their colony morphology and ITS-5.8S rDNA sequencing. TLC-MTT-Bioautography assays and DPPH radical scavenging assays were employed to assess the antibacterial and antioxidant activities of ethyl acetate extracts of the endophytic fungi.
Results
One hundred and twenty-five endophytic strains were isolated from the roots of B. polyandra, including 70 from female samples and 55 from male samples. Of them, twenty-two distinct isolates representing 15 genera and 22 species based on their ITS-rDNA genomic sequence were successfully identified from female and male samples of B. polyandra. The genus Calonectria was the most prevalent genus, with a CF% of 18.3, followed by the genera Clonostachys and Botryosphaeria, with CF% values of 13.4 and 10.0, respectively. Interestingly, the fungal extracts exhibited broad-spectrum antibacterial activities against gram-positive and gram-negative bacteria, as well as potential antioxidant activities with IC50 values ranging from 0.45 to 6.90 mg/mL. Among them, endophytes Bpf-10 (Diaporthe sp.) and Bpf-11 (Botryosphaeria sp.) showed the strongest biological activities and more abundant secondary metabolites.
Conclusions
This study reported the diversity of endophytic fungi from the roots of B. polyandra and the antibacterial and antioxidant activities of the crude extracts for the first time. The results revealed that B. polyandra contains diverse culturable endophytic fungi that potentially produce natural antibacterial and antioxidant compounds with great value to the agriculture and pharmaceutical industries.
Similar content being viewed by others


Background
Plant endophytic fungi are microorganisms that grow inside plant tissues without causing negative symptoms to the host and produce biologically active substances (Sheik and Chandrashekar 2018). They have been regarded as a novel source of natural bioactive compounds with tremendous applications in medicine, agriculture, and the food industry. In the past few years, many valuable bioactive compounds with anticancer, insecticidal, antimicrobial, and cytotoxic activities have been successfully isolated from endophytic fungi (Ascêncio et al. 2014; Jia et al. 2016; Atiphasaworn et al. 2017; Bedi et al. 2017). Endophytic fungi produce bioactive compounds similar to the host plant (Venieraki et al. 2017). Thus, endophytic fungi can be used to isolate active metabolites and reduce the large-scale utilization of plants as a method to protect the environment.
Balanophora polyandra Griff., belonging to the family Balanophoraceae, is a natural medicinal parasitic plant that lives in the root system of many Fagaceae plants and is mainly distributed in southern China, Japan, Nepal, India, and Burma (Wang et al. 2006; Tao et al. 2009). The whole plant has been used as a folk medicine due to its antipyretic, antidotal, and hemostatic properties. Moreover, B. polyandra has also been used as a traditional Chinese medicine, especially for treating gonorrhea, syphilis, wounds, and bleeding of the alimentary tract (Wang et al. 2013). Previous studies have shown the presence of potentially active metabolites in B. polyandra with antioxidant, immunosuppressive, hypoglycemic, antitumor, and antibacterial activities (Wang et al. 2013; Ouyang et al. 2017). However, no reports have discussed endophytic fungi and their biological activities. Therefore, the diversity of endophytic fungi of B. polyandra with biological effects must be elucidated.
The threat of drug-resistant pathogens has become a major concern worldwide (Nafis et al. 2018). Ralstonia solanacearum is a soil-borne bacterium that causes bacterial wilt in eucalyptus plantations worldwide, and efficient control measures are still limited (Mao et al. 2021). Similarly, the oxidative stress caused by free radicals is known to be involved in pathophysiological events, especially in some human diseases, such as diabetes mellitus, aging, atherosclerosis, Alzheimer’s disease, and Parkinson’s disease (Gunasekaran et al. 2017). The main characteristic of antioxidant compounds is capturing and stabilizing free radicals. The development of effective and safe drugs to combat human, animal, and plant diseases is now urgently needed (Patil et al. 2016). One approach to solving these problems is to search for new antibacterial and antioxidant metabolites. Endophytic fungi broaden the scope of new antibiotics, chemotherapeutic agents, and agrochemicals with high efficiency and low toxicity (Hateet 2016; Das et al. 2017; Zhong et al. 2017).
A promising future for developing a new drug exists by exploiting and utilizing endophytic fungi resources from medicinal plants. Hence, this study aims to screen potential endophytic fungi with significant antibacterial and antioxidant activities from the roots of B. polyandra. Molecular and morphological approaches were used for the isolation, characterization, and analysis of the diversity of endophytic fungi. Furthermore, the antibacterial and antioxidant activities of endophytic fungal extracts were assessed using TLC-MTT-Bioautography assays and DPPH radical scavenging assays. Finally, the chemical compositions of the crude extracts that had significant biological activities were analyzed using HPLC.
Results
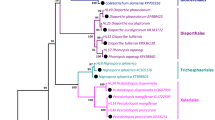
Isolation, identification, and phylogenetic analysis of endophytic fungi isolated from B. polyandra
A total of 125 endophytic fungi were isolated from 60 samples of B. polyandra (30 female samples and 30 male samples). Seventy fungal isolates were obtained from female samples, and fifty-five were obtained from male samples. According to their colony morphology (shape of conidia, mycelial growth rate, colony color and texture, etc.), 22 distinct fungal isolates (Bpf-1~Bpf-22) were selected for further molecular and microscopic identification. The colonies of Bpf-1~Bpf-22 grown on PDA medium are shown in Fig. 1. The obtained ITS sequences were compared with those in GenBank to identify the fungi. They were identified as members of fifteen genera, including Clonostachys (Bpf-1 and Bpf-15), Gliocladiopsis (Bpf-2), Calonectria (Bpf-3, Bpf-5, Bpf-9, Bpf-15 and Bpf-18), Gliocephalotrichum (Bpf-4), Pestalotiopsis (Bpf-6), Botryosphaeria (Bpf-7 and Bpf-11), Trichoderma (Bpf-8), Diaporthe (Bpf-10), Myrothecium (Bpf-12), Cylindrocladium (Bpf-13), Fusarium (Bpf-14 and Bpf-16), Colletotrichum (Bpf-17 and Bpf-20), Mucor (Bpf-19), Lasiodiplodia (Bpf-20), and Neofusicoccum (Bpf-22) (Table 1). Calonectria was the most prevalent genus, with a colonization frequency (CF) of 18.3%, followed by Clonostachys and Botryosphaeria with CF values of 13.4% and 10.0%, respectively. The genetic identities of 22 isolates were greater than 98%. The obtained ITS sequences of 22 isolates were submitted to GenBank to obtain their accession numbers (MH378888 and MH378889; MH397479-MH397498) and the closest related species were identified from a BLASTn analysis. The identified fungi with their accession numbers, the closest related species, the percentage of identity, and colonization frequency are presented in Table 1.

Front views of the colonies of endophytic fungi isolated from B. polyandra. a~v Bpf-1 to Bpf-22, respectively
Assessment of endophytic fungal diversity
The detailed results of the analysis of endophytic fungal diversity in male and female samples associated with B. polyandra are listed in Table 2. Larger values indicated the richness of the endophytic fungi in the samples. According to the Margalef abundance index (D′), female samples (2.354) had a higher value than male samples (2.246), showing the richness of the endophytic fungi in female samples compared to male samples. Moreover, Simpson’s (D) and Shannon’s (H') diversity indices in female samples were relatively higher than those in male samples (D = 0.881 and 0.869; H' = 2.247 and 2.126, respectively), suggesting that these endophytic fungi preferentially colonize female samples. However, the Pielou species evenness index (J) of female samples (0.923) was similar to that of male samples (0.937), indicating a uniform species composition across both hosts. The values of D′, D, H', and J in the whole tissue were 2.900, 0.898, 2.469, and 0.912, respectively. These values indicate the higher diversity of endophytic fungi in B. polyandra.
Antibacterial activity of the endophytic fungal extracts
The antibacterial activity of the fungal extracts against five test bacteria (Escherichia coli, Pseudomonas lachrymans, Xanthomonas vesicatoria, Ralstonia solanacearum and Bacillus subtilis) are summarized in Table 3. All 22 endophytic fungal extracts showed antibacterial activity against all the test bacteria to different degrees. For instance, the extracts of endophytes Bpf-1, Bpf-3, Bpf-4, Bpf-8, Bpf-9, Bpf-10, Bpf-11, Bpf-12, Bpf-14, and Bpf-22 were more active as antibacterial agents than the extracts of other endophytes. Among them, Bpf-1, Bpf-11, and Bpf-14 showed the highest inhibitory activity against all the test bacteria with inhibition zone diameters exceeding 10 mm. On the other hand, some fungal extracts (i.e., Bpf-7, Bpf-15, and Bpf-19) also exhibited inhibitory activity against all five test bacteria, and the inhibition zone diameters mainly ranged from 5 mm to 10 mm. The extracts of three endophytes (Bpf-5, Bpf-6, and Bpf-13) showed comparatively weak or no inhibition against all test bacteria. Among the test bacteria, E. coli was less susceptible to the endophytic fungal extracts, except for five samples (Bpf-1, Bpf-10, Bpf-11, Bpf-12 and Bpf-14), while R. solanacearum and X. vesicatoria were generally more susceptible.
The magnitude of the Rf value allowed us to determine the polarity of the compounds separated through our elution system. Based on the results of the antibacterial activity results, the Rf values of extracts exhibiting antibacterial activity all ranged from 0.00 to 0.65. Thus, the secondary metabolites of these endophytic fungi were mainly small to moderately polar substances. In this investigation, the gram-negative bacteria were generally more sensitive to the 22 endophytic fungal extracts than the gram-positive bacteria.
Antioxidant activity of the endophytic fungal extracts
The antioxidant activities of 22 endophytic fungal extracts from B. polyandra were evaluated using a DPPH radical scavenging assay. As shown in Fig. 2, all the extracts showed antioxidant activity to varying extents, with IC50 values ranging from 0.45 to 6.9 mg/mL. The extracts of endophytes Bpf-10 (Diaporthe sp.) and Bpf-11 (Botryosphaeria sp.) showed a stronger ability to inhibit DPPH radicals, with IC50 values of 0.46 and 0.45 mg/mL, respectively. However, their activities were lower than that of the standard BHT (0.02 mg/mL). The extracts of endophytes Bpf-12, Bpf-13, and Bpf-14 did not show any significant differences, and their IC50 values were 0.75, 0.73, and 0.71 mg/mL, respectively. The extract of the endophyte Bpf-21 showed the lowest antioxidant activity, with an IC50 value of 6.90 mg/mL.

Antioxidant activities of crude extracts of endophytic fungi isolated from B. polyandra
HPLC analysis of the extracts of endophytes with significant activities
Based on the results of strong antibacterial and antioxidant activities, the extracts of endophyte Bpf-10 (Diaporthe sp.) and Bpf-11 (Botryosphaeria sp.) were selected for further HPLC analysis (Fig. 3). These secondary metabolites mainly had retention times between 2 min and 6 min and between 10 min and 19 min, indicating that endophytes Bpf-10 and Bpf-11 contained major compounds with different polarities. Several fractions suggested that more than one compound produced by two endophytes was responsible for the bioactivity. However, further experiments are required to confirm whether the compounds detected in extracts mediate the antibacterial and antioxidant activities. The obtained HPLC chromatogram provides a theoretical reference for the further isolation, purification and identification of active components from endophytes Bpf-10 and Bpf-11.

HPLC-UV chromatograms of crude extracts of endophytes Bpf-10 and Bpf-11 at 210 nm
Discussion
Endophytic fungi, which are potential producers of medicinal substances, have attracted increasing attention in recent years (Wei et al. 2020). Medicinal plants provide a unique eco-environment for their endophytic fungi. A plethora of previous studies reported that special eco-environmental endophytic fungi might produce special bioactive natural products (Jia et al. 2016). Based on these considerations, we investigated the application of endophytic fungi, especially those isolated from B. polyandra, to evaluate their diversity and biological properties.
Endophytic fungi are detected in different medicinal plants worldwide. Their phylogenetic diversity has been reported in various forms to describe the interaction of fungi with the host plant (Tejesvi et al. 2011; Murdiyah 2017). In the present study, 125 endophytic fungi were isolated from B. polyandra, and 22 isolates were identified successfully based on morphological features and a sequence analysis of the ITS regions. These isolates showed 98.48–100.00% similarity to their assigned taxa. These fungal isolates belonged to one phylum, three classes, six orders and fifteen genera, showing the phylogenetic diversity of the endophytic fungi. Calonectria was the dominant genus and has also been reported in other plants, such as Acacia persea, Sarcococca hookeriana, and Buxus sempervirens (Dann et al. 2012; Wight et al. 2016). Members of a variety of common endophytic genera observed in the present study, such as Trichoderma, Fusarium and Colletotrichum, were typically isolated from different hosts (Hidayat et al. 2016; Ntuba-Jua et al. 2017). In the whole tissue, the large values of H' 2.469 and D' 2.900 revealed that B. polyandra hosted rich and diverse endophytic fungi.
Some research groups have reported the antibacterial activity of endophytic fungi from medicinal plants against various pathogenic microbes (Sathish et al. 2014; Liu et al. 2016; Wahab et al. 2017). Another objective of this study was to assess the antibacterial and antioxidant activities of endophytic fungal extracts. All extracts (Bpf-1 to Bpf-22) exhibited broad-spectrum antibacterial activities against E. coli, P. lachrymans, X. vesicatoria, R. solanacearum, and B. subtilis. Among them, Bpf-10 and Bpf-11 showed the strongest antibacterial activities, suggesting that they could be used as a potential source of antibacterial agents. Our results are supported by previous studies that demonstrated Diaporthe LGMF907 had potent antibacterial activity against E. coli, Saccharomyces cerevisiae, methicillin-sensitive Staphylococcus aureus, and methicillin-resistant S. aureus (de Medeiros et al. 2018). In addition, Botryosphaeria MGN23-3 also displayed strong antibacterial activity against Bacillus cereus and B. subtilis (da Silva et al. 2022). The remaining fungal extracts showed different activities toward the different tested bacteria, which might result from morphological differences in the cell walls of these pathogens (Gunasekaran et al. 2017). R. solanacearum and X. vesicatoria were sensitive to most fungal extracts. This result corroborated the findings reported by Ouyang (Ouyang et al. 2017) that extracts of B. polyandra showed significant antibacterial activity against R. solanacearum.
In the present study, all endophytic fungal extracts showed different levels of antioxidant activity. The extracts of endophytes Bpf-10 (Diaporthe sp.) and Bpf-11 (Botryosphaeria sp.) showed the best antioxidant activities based on the reduction of DPPH. As a proof of their efficiency, Diaporthe sp. MFLUCC16-0682 and Botryosphaeria MGN23-3 were reported to have notable antioxidant activities (Tanapichatsakul et al. 2017; da Silva et al. 2022). In addition, altenusin and djalonensone isolated from Botryosphaeria sp. had antioxidant activities (Xiao et al. 2014). Many previous studies have proven that some polyphenol, flavonoid and tannin compounds seem to play an important role in reducing peroxidation (Mazandarani et al. 2014; Kada et al. 2017). However, polyphenols, flavonoids, and tannins compounds were found to be absent from fungal extracts (Sharma et al. 2022). Hence, further confirmation is needed to determine whether a positive correlation exists between the extracts and these compounds. In the present study, all activities of fungal extracts were lower than that of the standard butylated hydroxytoluene (BHT). However, negative results do not mean an absence of the bioactive constituents in all fungal extracts, and they may contain other active chemical components that produce a definite physiological action. In this study, two endophytic fungi Bpf-10 (Diaporthe sp.) and Bpf-11 (Botryosphaeria sp.) were screened for the first time from medicinal plant B. polyandra, which showed strong antibacterial and antioxidant activities. Furthermore, there are few studies on the biological activities of the secondary metabolites of Gliocladiopsis (Bpf-2), Calonectria (Bpf-3, Bpf-5, Bpf-9, Bpf-15 and Bpf-18), Gliocephalotrichum (Bpf-4), and Cylindrocladium (Bpf-13).
Conclusions
In this study, the antibacterial and antioxidant activities of ethyl acetate extracts of the culturable endophytic fungi from the root of the holoparasitic plant B. polyandra were reported. A total of 125 endophytic fungal isolates were isolated from 60 samples of B. polyandra (30 female samples and 30 male samples). Of them, twenty-two distinct isolates (Bpf-1~Bpf-22) were selected for identification and characterization using molecular and morphological analyses. Fifteen genera were identified, among which Calonectria, Clonostachys, and Botryosphaeria were dominant. The crude extracts of Diaporthe neotheicola Bpf-10 and Botryosphaeria dothidea Bpf-11 showed potent inhibitory activities against DPPH radicals and pathogenic bacteria (E. coli, P. lachrymans, X. vesicatoria, R. solanacearum, and B. subtilis). Moreover, the HPLC chromatogram showed the presence of secondary metabolites with different polarities in the Bpf-10 and Bpf-11 extracts. These findings indicated that endophytic fungi from the holoparasitic plant B. polyandra have great potential to produce antioxidant and antibacterial compounds. Subsequent research will focus on the isolation and identification of the antibacterial and antioxidant compounds from these fungi, as well as on their applications as biocontrol agents.
Materials and methods
Materials
Whole plants of B. polyandra were collected from Chebaling National Nature Reserve, Guangdong Province of China, in September 2015. All the samples were placed in a plastic bag and immediately transported to the laboratory for further study. The plant specimens were authenticated by Dr. Mingxuan Zheng at South China Agricultural University. The plant materials were stored in sealed plastic bags at 4 °C until further use.
Isolation of the endophytic fungi
The endophytic fungi were isolated using the method described by Shan et al. with some modifications (Shan et al. 2019). Plant materials were thoroughly washed for 20 min with running tap water and surface-sterilized with 75% ethanol for 30 s, followed by three rinses with sterilized distilled water. They were then treated with 0.2% mercuric chloride for 20 min and then washed thrice with sterile distilled water. The surface-sterilized tissues were dried on sterile filter papers under aseptic conditions. Finally, each tissue sample was cut into 5 × 5 mm pieces and placed on a potato dextrose agar (PDA) plate containing streptomycin sulfate (500 μg/L). The culture plates were incubated at 28 °C for 1–3 weeks and observed daily. The emerging colonies were subcultured several times on fresh PDA plates to obtain pure isolates. Finally, the pure isolate was transferred into a PDA slant and stored at 4 °C until further use. The colonization frequency (CF %) of each pure isolate was calculated using the following formula:
where “NCOL” represents the number of segments colonized by the emerging fungus, and “Nt” represents the total number of samples segment (Shan et al. 2020).
Morphological characterization of endophytic fungi
For morphological identification, all endophytic fungi that varied in shape, growing area, exudate drop color, growth rates, surface texture, reverse color, radial lines, and concentric were observed and identified according to standard taxonomic manuals and textbooks (Praptiwi et al. 2016).
Molecular characterization of endophytic fungi
DNA was extracted from fresh mycelium using a fungal genomic DNA extraction kit (Shanghai Biological Engineering Co., China) according to the manufacturer’s method. The ITS region of rDNA was amplified by polymerase chain reaction (PCR) and subsequently sequenced with the universal primers ITS4 (5′-TCCTCCGCTTATTGATATGC-3′) and ITS5 (5′-GGAAGTAAAAGTCGTAACAAGG-3′). The PCR system was as follows: 25 μL of PCR Master Mix, 21 μL of double-distilled H2O, 2 μL of template DNA, 1 μL of forward primer ITS4, and 1 μL of reverse primer ITS5. The PCR conditions were set as follows: predenaturation at 95 °C for 2 min; 30 cycles of denaturation at 94 °C for 40 s, annealing at 56 °C for 40 s, extension at 72 °C for 1 min and 20 s; and extension at 72 °C for 10 min. The PCR products were sequenced and purified by Shanghai Biological Engineering Co., China. The obtained DNA sequences were submitted to GenBank and compared with a BLASTn analysis (Shan et al. 2019).
Assessment of endophytic fungi diversity
The diversity of endophytic fungi at each site was estimated according to Margalef’s abundance index (D'), Simpson index (D), the Shannon-Wiener diversity index (H'), and the Pielou species evenness index (J). The following formula was used: D' = (S-1)/ln N, where “S” represents the number of species and “N” is the number of individuals in the sample (Cosoveanu et al. 2018). D = 1 − Σ Pi2, where the ratio “Pi” is the frequency of colonization of the taxon in the sample (Zheng et al. 2013). H' = - Σ Pi ln Pi, where “H'” was used to show the diversity of the endophytic fungal species (Sadeghi et al. 2019). J = H'/ln(S), where “J” denotes the uniformity of the endophytic fungi (Zheng et al. 2013).
Preparation of crude extracts of endophytic fungi
The pure isolates of endophytic fungi were cultivated on PDA plates for 4–7 days. Afterward, 3 to 4 agar plugs with mycelia were inoculated into a 50-mL conical flask containing 20 mL of potato dextrose broth (PDB) (3 flasks for each fungus). All flasks were incubated at 150 rpm on a rotary shaker at 28 °C in the dark for 5–7 days. Each fungal broth and mycelia were inoculated into two flasks containing 20 g of sterile rice for 60 days under aseptic conditions. The fermented product was extracted thrice with ethyl acetate under sonication. Finally, the solvent was evaporated and concentrated using a rotary evaporator to obtain crude extracts. The ethyl acetate crude extracts were stored at 4 °C until use.
Detection of the antibacterial activity of the endophytic fungal extracts
The antibacterial activity of the endophytic fungal extracts was detected using a thin-layer chromatography (TLC)-bioautography assay (Shan et al. 2012). One gram-positive (B. subtilis) and four gram-negative (E. coli, P. lachrymans, X. vesicatoria, and R. solanacearum) bacterial strains were used as test bacteria. All the bacterial cultures were reactivated in Luria-Bertani (LB) broth medium for 12 h at 28 °C, followed by streaking on LB agar plates. The bacterial suspension (108 CFUs/mL) was mixed with molten semisolid LB medium (with 0.5% agar) before use. Five microliter of the ethyl acetate extracts was spotted onto a silica TLC plate, and then 5 μL of a streptomycin sulfate (CK+) solution (0.2 mg/mL) was spotted onto the lower right of the TLC plate. The prepared TLC plates were developed in a glass tank with the petroleum ether to acetone (4:1) (v/v) solvent system. The prepared bacterial suspension was poured uniformly over the TLC plate and incubated at 25 °C for 12 h under humid conditions. Last, the TLC plate was sprayed with a 5 mg/mL solution of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) and then incubated for 2 h. The antibacterial activity of endophytic extracts was assayed by measuring the white inhibition zone diameter on the purple background.
Assessment of the antioxidant activity of the endophytic fungal extracts
The DPPH radical scavenging assay was employed to examine the antioxidant activity of the endophytic fungal extracts using the microtiter plate (96-well) spectrophotometric method (Shan et al. 2019) with some modifications. Briefly, 20 mg of DPPH were dissolved in 100 mL of ethanol to obtain a 0.2 mg/mL DPPH solution (0.004% w/v) for this assay. The stock solutions of test samples were prepared separately by dissolving 0.1 g of ethyl acetate extract in 1 mL of ethanol. A series of stock solutions of different concentrations (20, 10, 5, 2.5, 1.25, 0.625, 0.3125, and 0.15625 mg/mL) were prepared by diluting the solutions with ethanol. Butylated hydroxytoluene (BHT) was used as a positive control and prepared using the same method. The final concentrations of BHT were 0.4, 0.2, 0.1, 0.05, 0.025, 0.0125, 0.00625, and 0.003125 mg/mL. After completing the preparation of all solutions, 80 μL of DPPH were added to each well containing 20 μL of sample solutions or BHT solutions at different concentrations. These reaction mixtures were homogenized well and incubated in the dark for 10 min and then incubated for 30 min in a water bath at 37 °C. The absorbance was measured at 517 nm using a spectrophotometer, and tests were performed in triplicate. Ethanol was used as a reference standard. DPPH inhibition was calculated using the following equation: DPPH inhibition (%) = [(A517 nm of control − A517 nm of sample)/A517 nm of control] × 100, where A is the absorbance obtained for a sample or the control. The median inhibitory concentration (IC50) value was calculated using a linear relationship between the percentage of DPPH inhibition results and their respective results and predictive equations.
HPLC analysis of the extracts of superior endophytes
High-performance liquid chromatography (HPLC) analysis of the fungal extracts was performed using the gradient elution method (Shan et al. 2020) with an XB-C18 reverse-phase (250 mm × 10 mm, 10 μm, Welch, Shanghai, China). Commercial grade water with 0.01% trifluoroacetic acid was used as mobile phase A, and acetonitrile with 0.01% trifluoroacetic acid was used as mobile phase B. The HPLC analysis was performed using a gradient of water to acetonitrile (0–2 min, 20% B, 2–15 min, 20–50% B, 15–16 min 50–100% B, 16–30 min, 100% B) at a flow rate of 1 mL/min, temperature of 40 °C; and UV detection at 210 nm.
Availability of data and materials
The generated nucleotide sequence of the endophytic fungal isolates (isolation number Bpf-1 ~ Bpf-22) can be accessed in GenBank under accession numbers MH378888, MH378889, MH397479 to MH397488 (https://blast.ncbi.nlm.nih.gov/Blast.cgi). The datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Ascêncio PGM, Ascêncio SD, Aguiar AA, Fiorini A, Pimenta RS (2014) Chemical assessment and antimicrobial and antioxidant activities of endophytic fungi extracts isolated from Costus spiralis (Jacq.) Roscoe (Costaceae). Evid Based Complement Alternat Med 1:1–10
Atiphasaworn P, Monggoot S, Gentekaki E, Brooks S, Pripdeevech P (2017) Antibacterial and antioxidant constituents of extracts of endophytic fungi isolated from Ocimum basilicum var. thyrsiflora leaves. Curr Microbiol 74:1185–1193
Bedi A, Adholeya A, Deshmukh SK (2017) Novel anticancer compounds from endophytic fungi. Curr Biotechnol 6:1–17
Cosoveanu A, Sabina SR, Cabrera R (2018) Fungi as endophytes in Artemisia thuscula: juxtaposed elements of diversity and phylogeny. J Fungi 4:2–21
da Silva AA, Polonio JC, Bulla AM, Polli AD, Castro JC, Soares LC et al (2022) Antimicrobial and antioxidant activities of secondary metabolites from endophytic fungus Botryosphaeria fabicerciana (MGN23-3) associated to Morus nigra L. Nat Prod Res. https://doi.org/10.1080/14786419.2021.1947272
Dann EK, Cooke AW, Forsberg LI, Pegg KG, Tan YP, Shivas RG (2012) Pathogenicity studies in avocado with three nectriaceous fungi, Calonectria ilicicola, Gliocladiopsis sp. and Ilyonectria liriodendri. Plant Pathol 61:896–902
Das M, Prakash HS, Nalini MS (2017) Antioxidative and antibacterial potentials of fungal endophytes from Justicia Wynaadensis Heyne: an ethnomedicinal rain forest species of western Ghats. Asian J Pharm Clin Res 10:203–209
de Medeiros AG, Savi DC, Mitra P, Shaaban KA, Jha AK, Thorson JS et al (2018) Bioprospecting of Diaporthe terebinthifolii LGMF907 for antimicrobial compounds. Folia Microbiol 63:499–505
Gunasekaran S, Sathiavelu M, Arunachalam S (2017) In vitro antioxidant and antibacterial activity of endophytic fungi isolated from Mussaenda luteola. J Appl Pharma Sci 7:234–238
Hateet RR (2016) Antibacterial and antioxidant activites of secondary metabolites of endophytic fungus Stemphylium radicinum (Meier, Drechs and Eddy). Ori Pharm Exp Med 57:558–563
Hidayat I, Radiastuti N, Rahayu G, Achmadi SS, Okane I (2016) Endophytic fungal diversity from Cinchona calisaya based on phylogenetic analysis of the ITS ribosomal DNA sequence data. Curr Res Environ Appl Mycol 6:132–142
Jia M, Chen L, Xin HL, Zheng CJ, Rahman K, Han T, Qin LP (2016) A friendly relationship between endophytic fungi and medicinal plants: a systematic review. Front Microbiol 7:1–14
Kada S, Bouriche H, Senator A, Demirtas I, Ozen T, Toptanci BC, Kizil G, Kizil M (2017) Protective activity of Hertia cheirifolia extracts against DNA damage, lipid peroxidation and protein oxidation. Pharm Biol 55:330–337
Liu YH, Hu XP, Li W, Cao XY, Yang HR, Lin ST, Xu CB, Liu SX, Li CF (2016) Antimicrobial and antitumor activity and diversity of endophytic fungi from traditional Chinese medicinal plant Cephalotaxus hainanensis Li. Genet Mol Res 15:1–11
Mao Z, Zhang W, Wu C, Feng H, Peng Y, Shahid H, Cui Z, Ding P, Shan T (2021) Diversity and antibacterial activity of fungal endophytes from Eucalyptus exserta. BMC Microbiol 21:155
Mazandarani M, Ghafourian M, Khormali A (2014) Ethnopharmacology, antibacterial and antioxidant activity of dittrichia graveolens (L.) W. Greuter. which has been used as remedies antirheumatic, anti-inflammation and antiinfection against leishmaniasis in the traditional medicine of Gorgan, Iran. Crescent J Med Biol Sci 1:125–129
Murdiyah S (2017) Endophytic fungi of various medicinal plants collected from evergreen forest Baluran national park and its potential as laboratory manual for mycology course. J Pendidikan Biologi Indonesia 3:64–71
Nafis A, Kasrati A, Azmani A, Ouhdouch Y, Hassani L (2018) Endophytic actinobacteria of medicinal plant Aloe vera: Isolation, antimicrobial, antioxidant, cytotoxicity assays and taxonomic study. Asian Pac J Trop Biomed 8:513–518
Ntuba-Jua GM, Mih AM, Bechem EET (2017) Diversity and distribution of endophytic fungi in different Prunus africana (Hook. f.) Kalkman provenances in Cameroon. Biosci Plant Biol 4:7–23
Ouyang J, Wu C, Wang X, Wang S, Wu H, Wang J, Shan T (2017) The crude extracts of Balaophora polyandra and their antimicrobial activities. Chin J Trop Agric 37:61–65
Patil RH, Patil MP, Maheshwari VL (2016) Bioactive secondary metabolites from endophytic fungi: a review of biotechnological production and their potential applications. Stud Nat Prod Chem 1:189–205
Praptiwi, Palupi KD, Fathoni A, Wulansari D, Ilyas M, Agusta A (2016) Evaluation of antibacterial and antioxidant activity of extracts of endophytic fungi isolated from Indonesian Zingiberaceous plants. Nusantara Biosci 8:306–311
Sadeghi F, Samsampour D, Seyahooei MA, Bagheri A, Soltani J (2019) Diversity and spatiotemporal distribution of fungal endophytes associated with Citrus reticulata cv, Siyahoo. Curr Microbiol 76:279–289
Sathish L, Pavithra N, Ananda K (2014) Evaluation of antimicrobial activity of secondary metabolites and enzyme production from endophytic fungi isolated from Eucalyptus citriodora. J Pharm Res 8:269–276
Shan T, Duan Z, Wu C, Li Z, Wang S, Mao Z (2020) Secondary metabolites of symbiotic fungi isolated from Blaptica dubia and their biological activities. J Environ Entomol 42:170–179
Shan T, Qin K, Xie Y, Zhang W, Mao Z, Wang J (2019) Secondary metabolites of endophytic fungi isolated from Casuarina equisetifolia and their bioactivities. J S China Agr Univ 40:67–74
Shan T, Sun W, Lou J, Gao S, Mou Y, Zhou L (2012) Antibacterial activity of the endophytic fungi from medicinal herb, Macleaya cordata. Afr J Biotechnol 11:4354–4359
Sharma A, Sagar A, Rana J, Rani R (2022) Green synthesis of silver nanoparticles and its antibacterial activity using fungus Talaromyces purpureogenus isolated from Taxus baccata Linn. Micro Nano Syst Lett 10:2–12
Sheik S, Chandrashekar KR (2018) Fungal endophytes of an endemic plant Humboldtia brunonis Wall. of western Ghats (India) and their antimicrobial and DPPH radical scavenging potentiality. Ori Pharm Exp Med. 18:115–125
Tanapichatsakul C, Monggoot S, Gentekaki E, Pripdeevech P (2017) Antibacterial and antioxidant metabolites of Diaporthe spp. isolated from flowers of Melodorum fruticosum. Curr Microbiol 75:476–483
Tao RY, Ye F, He YB, Tian JY, Liu GG, Ji TF, Su YL (2009) Improvement of high-fat-diet-induced metabolic syndrome by a compound from Balanophora polyandra Griff in mice. Eur J Pharmacol 616:314–340
Tejesvi MV, Kajula M, Mattila S, Pirttilä AM (2011) Bioactivity and genetic diversity of endophytic fungi in Rhododendron tomentosum Harmaja. Fungal Divers 47:97–107
Venieraki A, Dimou M, Katinakis P (2017) Endophytic fungi residing in medicinal plants have the ability to produce the same or similar pharmacologically active secondary metabolites as their hosts. Hell Plant Prot J 10:51–66
Wahab MAA, Bahkali AHA, Gorban AME, Hodhod MS (2017) Natural products of Nothophoma multilocularis sp. nov. an endophyte of the medicinal plant Rhazya stricta. Mycosphere. 8:1185–1200
Wang KJ, Zhang YJ, Yang CR (2006) New Phenolic Constituents from Balanophora polyandra with radical-Scavenging activity. Chem Biodivers 3:1317–1324
Wang YG, Yang JB, Wang AG (2013) Hydrolyzable tannins from Balanophora polyandra. Acta Pharm Sin B 3:46–50
Wei J, Chen F, Liu YM, Abudoukerimu A, Zheng Q, Zhang XB, Sun YP, Yimiti D (2020) Comparative metabolomics revealed the potential antitumor characteristics of four endophytic fungi of brassica rapa L. Am Chem Soc 5:5939–5950
Wight MM, Salazar CS, Demers JE, Clement DL, Rane KK, Crouch JA (2016) Sarcococca blight: use of whole genome sequencing for fungal plant disease diagnosis. Plant Dis 100:1093–1100
Xiao J, Zhang Q, Gao YQ, Tang JJ, Zhang AL, Gao JM (2014) Secondary metabolites from the endophytic Botryosphaeria dothidea of Melia azedarach and their antifungal, antibacterial, antioxidant, and cytotoxic activities. J Agric Food Chem 62:3584–3590
Zheng JH, Kang JC, Lei BX, Li QR, Wen TC, Meng ZB (2013) Diversity of endophytic fungi associated with Ginkgo biloba. Mycosystema. 32:671–681
Zhong LY, Zou L, Tang XH, Li WF, Li X, Zhao G, Zhao JL (2017) Community of endophytic fungi from the medicinal and edible plant Fagopyrum tataricum and their antimicrobial activity. Trop J Pharm Res 16:387–396
Acknowledgements
We thank Dr. Mingxuan Zheng from the College of Forestry and Landscape Architecture, South China Agricultural University, for the taxonomic identification of the plant materials.
Funding
This research was co-financed by the National Natural Science Foundation of China (32071766), the Natural Science Foundation of Guangdong Province (2019A1515011554; 2022A1515010944), and the Key Research and Development Projects of Guangdong Province (2020B020214001).
Author information
Authors and Affiliations
Contributions
W.W, S.W. and T.S. collected the plant material. C.W., W.W, X.W., and T.S. performed the isolation and identification of the endophytic fungi. C.W., W.W, X.W., and T.S. evaluated the antimicrobial activity. C.W., H.S., Y.Y, and Y.W. performed the antioxidant activity. C.W., H.S., and Y.Y contributed in the diversity analysis of endophytic fungi. H.S., Y.Y, and Y.W. performed the HPLC analysis of the crude extract. C.W., W.W, H.S., Y.Y, and T.S. prepared the figures and tables. S.W. and T.S. designed the research. All the authors contributed in writing, editing, and revising the manuscript. The author(s) read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
The healthy plants of B. polyandra were collected in September 2015 from Chebaling National Nature Reserve, Guangdong Province, China. The taxonomic identification of the plant materials was performed by Dr. Mingxuan Zheng of College of Forestry and Landscape Architecture (SCAU), where the voucher specimen (SCAULPMH-1509015) of the plant was deposited. All experiments were approved by the College of Forestry and Landscape Architecture (SCAU) and were strictly evaluated in accordance with the IUCN Policy Statement on Research Involving Species at Risk of Extinction and the Convention on the Trade in Endangered Species of Wild Fauna and Flora.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Wu, C., Wang, W., Wang, X. et al. Diversity and communities of culturable endophytic fungi from the root holoparasite Balanophora polyandra Griff. and their antibacterial and antioxidant activities. Ann Microbiol 72, 19 (2022). https://doi.org/10.1186/s13213-022-01676-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13213-022-01676-6