
Abstract
Background
The demonstration that the recently discovered Anopheles symbiont Microsporidia MB blocks malaria transmission in Anopheles arabiensis and undergoes vertical and horizontal transmission suggests that it is a promising candidate for the development of a symbiont-based malaria transmission-blocking strategy. The infection prevalence and characteristics of Microsporidia MB in Anopheles gambiae sensu stricto (s.s.), another primary vector species of malaria in Kenya, were investigated.
Methods
Field-collected females were confirmed to be Microsporidia MB-positive after oviposition. Egg counts of Microsporidia MB-infected and non-infected individuals were used to infer the effects of Microsporidia MB on fecundity. The time to pupation, adult sex ratio and survival were used to determine if Microsporidia MB infection has similar characteristics in the host mosquitoes An. gambiae and An. arabiensis. The intensity of Microsporidia MB infection in tissues of the midgut and gonads, and in carcasses, was determined by quantitative polymerase chain reaction. To investigate horizontal transmission, virgin males and females that were either Microsporidia MB-infected or non-infected were placed in standard cages for 48 h and allowed to mate; transmission was confirmed by quantitative polymerase chain reaction targeting Microsporidia MB genes.
Results
Microsporidia MB was found to naturally occur at a low prevalence in An. gambiae s.s. collected in western Kenya. Microsporidia MB shortened the development time from larva to pupa, but other fitness parameters such as fecundity, sex ratio, and adult survival did not differ between Microsporidia MB-infected and non-infected hosts. Microsporidia MB intensities were high in the male gonadal tissues. Transmission experiments indicated that Microsporidia MB undergoes both maternal and horizontal transmission in An. gambiae s.s.
Conclusions
The findings that Microsporidia MB naturally infects, undergoes maternal and horizontal transmission, and is avirulent in An. gambiae s.s. indicate that many of the characteristics of its infection in An. arabiensis hold true for the former. The results of the present study indicate that Microsporidia MB could be developed as a tool for the transmission-blocking of malaria across different Anopheles species.
Graphical Abstract

Similar content being viewed by others

Background
Anopheles gambiae sensu stricto (s.s.) mosquitoes are efficient vectors of the main malaria parasite Plasmodium falciparum, especially in parts of sub-Saharan Africa, where malaria still remains a huge burden. While frontline insecticide-based vector control strategies have contributed significantly to the decrease in malaria cases [1], there are challenges that need to be overcome, such as the development of insecticide resistance [2, 3]. Alternative mosquito-borne control interventions that utilize symbionts which can be stably maintained in their mosquito hosts, conferring protection against parasites and pathogens, e.g. in Anopheles against P. falciparum [4,5,6] and in Aedes against arboviruses [7,8,9], have been proposed. The discovery that the novel Microsporidia MB symbiont identified in wild Anopheles arabiensis mosquitoes protects these hosts against infection with Plasmodium parasites [10] and is vertically and horizontally transmitted in adult mosquitoes [11], has led to the suggestion that it could be a promising candidate for integrated vector management.
While Microsporidia MB was initially reported in Anopheles arabiensis, which is a member of the Anopheles gambiae sensu lato (s.l.) species complex, there have been reports from Ghana that it infects other members of the An. gambiae species complex, such as Anopheles coluzzii and An. gambiae s.s. [12]. These findings are highly significant since An. gambiae s.l. is the main malaria vector in many parts of sub-Saharan Africa [13,14,15]. As An. arabiensis and An. gambiae s.s. are closely related and often found in sympatry [16], it is possible that Microsporidia MB could have similar characteristics in both of them, including vertical transmission, a lack of sex bias, and avirulence. In the wild, Microsporidia MB is generally found at a prevalence that ranges from 0 to 9% in populations of Anopheles arabiensis, Anopheles funestus, Anopheles gambiae, and Anopheles coluzzii [10,11,12]. Although Microsporidia MB has been reported in diverse Anopheles species, the infection characteristics in species of this genus other than those mentioned above have not yet been investigated. Anopheles mosquitoes collected from western Kenya, where An. gambiae s.s. is the dominant species, were used to determine the prevalence of Microsporidia MB infection in this host. Further, it was investigated if the infection characteristics of Microsporidia MB in An. gambiae s.s. are similar to those previously observed in An. arabiensis.
Methods
Ethical clearance
The Kenya Medical Research Institute (KEMRI) undertook an ethical review and granted ethical approval for this work [KEMRI/Scientific and Ethics Review Unit (SERU)/Centre for Biotechnology Research and Development (CBRD)/230/4341]. Community leaders and village participants enrolled for sample collection were provided with a detailed explanation of the study objectives, activities, procedures, benefits and risks before the commencement of data collection. Through the community leaders, informed consent was obtained from the owners of the households where samples were collected. Participation was on a voluntary basis and participants were free to withdraw from the study at any point.
Description of archived Anopheles samples
Anopheles mosquito DNA samples used for screening Microsporidia MB were retrieved from various projects undertaken at the Centre for Global Health Research in KEMRI between 2019 and 2021. Protein precipitation was used to extract nucleic acids (Puregene; Qiagen, the Netherlands). The DNA samples were from individual adult mosquitoes previously collected from different sites across Busia and Bungoma in western Kenya. The species were confirmed by molecular assay. To assess DNA integrity, a subset was selected at random to determine the quantity and quality of DNA by using a NanoDrop 2000c Spectrophotometer (Thermo Fisher, UK). An average DNA concentration of 61.3 ng/µL (25.73–184.23 ng/µL) and purity of 1.7 (1.42–2.10) were obtained.
Field collection of Anopheles mosquitoes
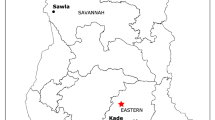
Collections of resting gravid and engorged female mosquitoes were undertaken indoors using manual aspiration. Collection in Busia (− 34.105278W, − 0.463333N) was undertaken between August 2021 and February 2022 from 0700 to 1130 hours using aspirators and electric torches/lights to target endophagic and endophilic gravid individuals that had acquired a blood meal late at night and were likely to be found in crevices or dark corners of the houses [15]. Similarly, collection was by indoor catches, undertaken from 0600 to 0900 hours, in Busia and areas of Bungoma adjacent to where previous studies had found more of the highly anthropophilic An. gambiae s.s. and An. funestus s.l. than An. arabiensis [17, 18]. Sites in Bungoma were excluded from the planned field collection where archived samples showed a low prevalence of Microsporidia MB compared to those from Busia. Collected females were placed in netted paper cups or standard cages and supplied with 10% sucrose before transporting them to KEMRI/Centre for Global Health Research insectary laboratory for processing.
Generating Microsporidia MB-infected semi-field colonies
All wild-caught mosquito females were identified morphologically using the keys of Gillies and Coetzee [19]. In the areas of Busia where mosquitoes were collected, An. gambiae s.l. and An. funestus s.l. are the commonest Anopheles species, with > 93% of the members of the An. gambiae species complex there previously confirmed as An. gambiae s.s. by PCR [18].
The females were placed in individual tubes containing a wet 1-cm × 1-cm Whatman filter paper to induce oviposition, and maintained in an insectary at 28 ± 2.5 °C, relative humidity of 60–80%, under a 12-h day and 12-h night cycle. All of the eggs laid by the Microsporidia MB-infected and non-infected females were counted under a microscope and placed in trays containing water for larval development in a semi-field setup at 30 ± 2.5 °C and relative humidity of 30–40% [11]. Once they had laid eggs, the generation zero (G0) females were set aside for processing to extract DNA, re-confirm their species identity, and screen them for the presence of Microsporidia MB infection, as previously described [10, 11]. The larvae of the Microsporidia MB-infected females were reared in pools (76% of samples, where the offspring of seven females constituted one pool), or as individual broods (24% of samples). The larvae were first fed 0.1 mg TetraMin Baby/larva per day, which was increased to 0.3 mg/larva per day for the late-stage instars before pupation. Adults were maintained in separate cages according to brood or pool and supplied with 10% sucrose. Field-derived G1 non-infected broods or pools were reared under the same conditions but separate from the Microsporidia MB-infected individuals, and were used as control isofemale lineages.
Determination of the effects of Microsporidia MB on An. gambiae s.s. fitness
To determine the effects of Microsporidia MB on host life history traits, 2464 wild-caught gravid female mosquitoes were placed in perforated 1.5-mL microcentrifuge tubes containing a soaked piece of Whatman paper to induce oviposition. The eggs from each female were counted as previously described [10, 11] before placing them in tubes containing water. After egg-laying, 53 G0 females that were confirmed to be An. gambiae s.s. and Microsporidia MB-positive by using qPCR were used to produce Microsporidia MB-positive progeny alongside non-infected controls selected at random (n = 374). The offspring were used to determine (i) the time period of larval development until pupation; (ii) the sex ratio, by counting the number of males and females in the broods of individual or pooled females; (iii) adult survival; and (iv) vertical transmission efficiency, by determining the number of offspring acquiring Microsporidia MB from each wild-caught Microsporidia MB-infected female.
Transmission of Microsporidia MB between live An. gambiae s.s. adults
Microsporidia MB-infected and uninfected An. gambiae s.s. virgin adult offspring were reared in standard 30-cm × 30-cm × 30-cm cages. Virgin male and female pupae were determined by visual examination of the terminalia and hatched in separate cages, as previously described [11]. To increase the chance of transmission, individual Microsporidia MB-infected donor and non-infected male and female recipients of the same age (range, 5–7 days old) were kept together in cages for 48 hr. The number of Microsporidia MB-infected donors of the same age ranged from two to 21 individuals, while those of the virgin uninfected recipients of the opposite sex ranged in number from 12 to 44 individuals (Additional file 4: Table S2). The set up for mates of the same age was in duplicate, and each age group represented an independent experiment. As the horizontal transmission of Microsporidia MB previously observed in An. arabiensis was between individuals of the opposite sex [11], in the present experiment, horizontal transmission was only examined between females and males and vice versa. All of the An. gambiae s.s. mosquitoes were subsequently screened by qPCR to establish Microsporidia MB infection statuses and intensities in both donor and recipient An. gambiae s.s. after the spermathecae had been examined to check for the presence of sperm in the recipient females [11]. A representative number of individuals in a cage (n = 5) was checked prior to screening all of the recipient and donor individuals in the cages for Microsporidia MB infection by qPCR. All of the samples in an experimental cage were used to determine the number of donors infected with Microsporidia MB and the number of recipient individuals acquiring a Microsporidia MB infection.
Microsporidia MB in tissues of An. gambiae s.s.
Microsporidia MB were quantified in dissected tissues of G1 Microsporidia MB-infected An. gambiae s.s. adults, 5–7 days post-emergence. Tissues of the midgut and gonads were separated from the whole mosquito, and the remainder was designated “the carcass,” as previously described [11]. The individual tissues were labeled separately and screened for the presence and intensity of Microsporidia MB by qPCR. Genomic DNA was extracted from the tissues in individual tubes by using protein precipitation [10, 11].
Molecular screening and quantification of Microsporidia MB
The detection and quantification of Microsporidia MB were done with specific primers (MB18SF, CGCCGGCCGTGAAAAATTTA; MB18SR, CCTTGGACGTGGGAGCTATC) previously designed to detect Microsporidia MB in An. arabiensis [10, 11]. Briefly, a 10-µL polymerase chain reaction (PCR) master mix consisting of 2 µL HOT FIREPol Blend Master Mix Ready to Load (Solis BioDyne, Estonia; mix components included HOT FIREPol DNA polymerase, 2 mM of each deoxynucleoside triphosphate and 7.5 mM magnesium chloride), 5 µL of nuclease-free PCR water, 0.5 µL of 5 pmol µL−1 forward and reverse primers, and 1 µL of the sample template, was prepared. The mixture was incubated in a thermocycler set up as follows: initial denaturation at 95 ˚C/15 min, followed by 35 cycles of denaturation at 95˚C/60 s, primer annealing for 90 s at 62 ˚C, extension at 72 ˚C for 60 s, and a final chain elongation step of 72 ˚C for 5 min. A qPCR reaction carried out using the MB18SF/MB18SR primers on a MIC qPCR cycler (Bio Molecular Systems, Australia) was used to determine the intensity of Microsporidia MB infection. These data were normalized by using Anopheles host ribosomal S7 gene primers (S7F, TCCTGGAGCTGGAGATGAAC; S7R, GACGGGTCTGTACCTTCTGG) [21].
Statistical analysis
An unpaired t-test was used to analyze data that were assumed to be normally distributed, while non-normally distributed data were analyzed by either two-tailed unpaired Mann–Whitney U-test or Kruskal–Wallis test following Dunn’s multiple comparisons post hoc test. A Mann–Whitney U-test was used to compare fecundity and development time between Microsporidia MB-infected and non-infected mosquitoes, while the Kruskal–Wallis test was used to estimate the significance of Microsporidia MB infection in tissues. To measure the level of significance of Microsporidia MB positivity rates, a chi-square test was used, while a log-rank test was used to estimate survival. A linear regression was used to establish the correlation coefficient between Go and G1 infection intensities and prevalences. To examine whether Microsporidia MB intensities impacted transmission rate, a mixed-effect model fit by maximum likelihood was used, where the total individuals exposed and the number of donors in the cage were considered random intercepts. All of the analyses were performed using either R (version 4.1.2) or GraphpadPrism software. The data are presented as mean ± SEM, and P < 0.05 was deemed to indicate statistical significance.
Results
Microsporidia MB detected in An. gambiae s.s.
Microsporidia MB was found in 79 out of 5067 DNA samples from mosquitoes collected from the two areas in western Kenya (Fig. 1). The overall Microsporidia MB prevalence was 1.7% (0.92–2.19) and 0.2% (0–0.2) for the total number of samples analyzed from Busia (78/4561) and Bungoma (1/506), respectively (Fig. 2; Additional file 3: Table S1). The average prevalence of Microsporidia MB in An. gambiae s.s. from the two areas was 1.6% (79/5067), and ranged from 0 to 2.2%. These prevalences are lower than those previously observed in sibling species of the An. gambiae species complex, such as An. arabiensis [10], An. gambiae s.s. and An. coluzzii [12]. Microsporidia MB was not detected in An. funestus s.s. from Busia (0/769) or from Bungoma (0/665) (Additional file 3: Table S1). Microsporidia MB was detected in An. arabiensis from Busia at a prevalence of 0.9% (3/333), but none of the An. arabiensis individuals from Bungoma harbored Microsporidia MB. Anopheles gambiae s.s. from Busia accounted for 98.7% (78/79) of individuals of this species harboring Microsporidia MB, while An. gambiae s.s. from Bungoma accounted for 1.3% of these mosquitoes (1/79), with only one individual harboring Microsporidia MB. The 78 Microsporidia MB-infected An. gambiae s.s. from Busia were from 4561 (1.7%) of the mosquitoes collected there; the single infected An. gambiae s.s. from Bungoma was from 506 (0.2%) mosquitoes collected there.
For Busia, 80.5% (4561/5663) of the collected Anopheles mosquitoes were identified as An. gambiae s.s., while 13.6% (769/5663) were identified as An. funestus s.s., and 5.9% (333/5663) were confirmed as An. arabiensis. In Bungoma, An. gambiae s.s. accounted for 43.2% (506/1171) of Anopheles mosquitoes collected there, while An. funestus s.s. was the predominant species at 56.8% (665/1171): An. arabiensis was not detected in this area.
When samples collected in 2022 were included in the analysis, the rate of Microsporidia MB positivity differed significantly between the two areas [χ2(1, n = 5067) = 6.8, P = 0.00917]. There was a lack of statistical significance when the data for the mosquitoes collected in 2020 were analyzed separately [χ2(1, n = 1818) = 2.6968, P = 0.10055] or were combined with those from the 2021 collection where Microsporidia MB was not detected [χ2(1, n = 2603) = 3.8529, P = 0.4966]. The results from the analysis of archived samples collected in 2020 and 2021 informed the subsequent field collection of An. gambiae s.s. from Busia (Microsporidia MB prevalence of 1.1%), where the Microsporidia MB positivity rate was higher than that in Bungoma (Microsporidia MB prevalence of 0.2%). For the 2022 field collections, Microsporidia MB was detected in An. gambiae s.s. (54/2464), and An. arabiensis (3/281) from Busia, but not in An. funestus s.l. (0/181).
Fifty-three of the 54 Microsporidia MB-positive An. gambiae s.s. that laid eggs were used to generate F1 progeny together with 374 non-infected An. gambiae s.s., to explore the life history traits of Microsporidia MB-infected An. gambiae s.s. and the transmission characteristics of Microsporidia MB in them.

Map showing the sampling sites in Busia and Bungoma, western Kenya, where Anopheles gambiae sensu lato (s.l.) infected with Microsporidia MB were collected

Positivity rate of Microsporidia MB (MB) infection in Anopheles gambiae sensu stricto (s.s.) and Anopheles arabiensis collected from the Busia and Bungoma study sites. Bar plots represent the prevalence of Microsporidia MB in An. gambiae s.s. and An. arabiensis from Bungoma and Busia. Error bars represent the SEM (* P < 0.05)
Microsporidia MB is avirulent in An. gambiae s.s.
To determine the characteristics of Microsporidia MB infection in An. gambiae s.s., parameters such as fecundity, larval development time, sex ratio, and adult survival were investigated in infected vs. uninfected isofemale lineages. The number of eggs laid by Microsporidia MB-infected females (n = 53) versus uninfected individuals (n = 374) did not differ significantly (two-tailed Mann–Whitney U-test = 9778, P = 0.874; Fig. 3a), indicating that Microsporidia MB does not have a sterilizing effect on female An. gambiae s.s. A significantly shortened larval development time was observed in An. gambiae s.s. larvae infected with Microsporidia MB (unpaired two-tailed t-test, t = 2.023, df = 106, P < 0.0001; Fig. 3b). No statistically significant difference was observed in the mean pupation rate between Microsporidia MB-infected (76.14 ± 1.88) and uninfected (72.68 ± 3.10) An. gambiae s.s. (unpaired two-tailed t-test, t = 0.955, df = 17, P = 2.11; Fig. 3c). The proportion of male and female offspring from Microsporidia MB-infected and uninfected An. gambiae s.s. did not differ significantly in isofemale lineages (unpaired two-tailed t-test, t = 0.1712, df = 22, P = 0.8657; Fig. 3d) or pooled lineages (Mann–Whitney U-test = 17, P = 0.9026; Additional file 1: Fig. S1A). No statistically significant differences were observed in survival between adult An. gambiae s.s. infected with Microsporidia MB and those that were uninfected (two-sided log-rank Mantel–Cox, χ2 = 0.2406, df = 1, P = 0.624; Fig. 3e). Taken together, these findings suggest that Microsporidia MB harbored in An. gambiae s.s. is avirulent.

Microsporidia MB did not affect certain metrics of Anopheles gambiae s.s. fitness. a Individual values; the line represents the mean number of eggs laid by females infected with Microsporidia MB (MB+) (n = 53) and by uninfected females (M−) (n = 374). Bar plots representing mean larval development time (b) and mean pupation rate (c) of Microsporidia MB-infected generation 1 (G1) and non-Microsporidia MB-infected G1. d Bar plots representing the mean sex ratio of the progenies of Microsporidia MB-infected and non-infected counterparts. e Survival curves representing the longevity of adult An. gambiae s.s. infected with Microsporidia MB and non-infected individuals. The results are expressed as mean ± SEM of independent experiments done in triplicate. ns No significant difference, **** P < 0.0001
Microsporidia MB undergoes vertical and horizontal transmission in An. gambiae s.s.
Microsporidia MB transmission from field-derived females to their offspring was observed at a frequency ranging from 0 to 100% (61.4 ± 8.93%) in 12 isofemale lineages (Fig. 4a) and from 28.6 to 85.3% (59.91 ± 8.46%) in six pooled lineages (Additional file 1: Figure S1B), which led to the hypothesis that Microsporidia MB colonizes and proliferates in the reproductive tissues of An. gambiae s.s.
To gain insights into the distribution of Microsporidia MB infecting An. gambiae s.s. host tissues, such as the midgut and reproductive tissues, and remaining sections (referred to as the carcass here), 7- to 10-day-old G1 adults were dissected and screened for the presence and intensity of Microsporidia MB. The prevalence of Microsporidia MB was high in the gonads of both females (84.21%, n = 36) and males (74.51%, n = 50) compared to female guts (15.79%) and males guts (19.61%) and males carcasses (5.88%) (Fig. 4b). The intensity of Microsporidia MB infection in female reproductive tissue was relatively high, but did not differ significantly from that in the guts (Mann–Whitney U-test = 24, P = 0.494, n = 17; Fig. 4c). The intensity of Microsporidia MB infection was significantly higher in the male gonads than in the male guts [χ2(2) = 135.265, P < 0.05, n = 50]; Fig. 4d) and in male gonads compared to male carcasses [χ2(1, n = 36) = 27.3103, P < 0.05; Fig. 4d]. A weak non-significant positive correlation was observed between the intensities of infection in the G1 and Go [r2 = 0.1276, P = 0.268, number of broods tested = 18; Additional file 2: Figure S2A]. A significant positive correlation was observed between Go infection intensities and the transmission rate of Microsporidia MB to the G1 offspring (r2 = 0.625, P = 0.0056, n = 18; Additional file 2: Fig. S2B), which suggested the likelihood of intensity-dependent maternal transmission.
When virgin Microsporidia MB-positive and non-infected individuals of the opposite sex were kept together in cages, 39 out of 40 Microsporidia MB-positive An. gambiae s.s. males were able to infect at least one female in their cages (range 3.45–12.5%); these results were confirmed by the visual detection of sperm in the female spermathecae before the validation of Microsporidia MB infection by qPCR. For instance, in BUR_KSMS 11 and 12, the spermathecae of three Microsporidia MB-infected individuals that contained sperm were confirmed by qPCR (Additional file 4: Table S2). The number of individuals acquiring Microsporidia MB infection through horizontal transfer determined by a similar molecular methodology ranged from 6.38 to 12.5% (Additional file 4: Table S2). It is also noteworthy that the transfer of Microsporidia MB from infected female(s) to one uninfected male was observed (rate of 6.25%; Additional file 4: Table S2). The lack of a statistically significant difference between infection and successful transmission when the mixed-effect model was applied to the data (P > 0.05) suggests that the transmission of Microsporidia MB from the reproductive tissues of An. gambiae s.s. to their offspring is a complex process.

Transmission of Microsporidia MB from Anopheles gambiae s.s. to their offspring. a Bar plots represent transmission rate of Microsporidia MB from 12 infected field-derived females (Go; n = 12) to their offspring (G1). b Bar plots representing Microsporidia MB prevalence in tissues of male and female An. gambiae s.s. [gonads (gon), guts (gut) and carcass (carc)]. Scatter plots representing mean relative densities of Microsporidia MB transmitted to An. gambiae s.s. tissues (gonads, guts and carcass) of female offspring (c) and gonads and guts of male offspring, but not male carcasses (d). The results are expressed as the mean ± SE from three separate experiments. * P < 0.05, ***P < 0.001
Discussion
The recent discovery of a Microsporidia MB symbiont in An. arabiensis with Plasmodium-transmission blocking capabilities has generated interest in the potential for the development of a symbiont-based alternative malaria control strategy. Other characteristics of Microsporidia MB, such as its transovarial transmission [10], horizontal transmission [11] and avirulence are of high relevance for its potential as a symbiont for the transmission blocking of Plasmodium, since these contribute to its ability to spread through and be maintained in populations of Anopheles mosquitoes. Microsporidia MB has also been found in Anopheles species other than An. arabiensis [11, 12]; however, the characteristics of these infections and their similarity to those of Microsporidia MB in An. arabiensis have not been investigated.
The present study demonstrates that Microsporidia MB can naturally infect An. gambiae s.s. in Kenya, and undergoes transovarial and horizontal transmission without having a deleterious effect on host fitness parameters such as larval development time, pupation rates and adult longevity. These parameters were compared between Microsporidia MB-infected and non-infected Anopheles, but it could not be ascertained if there were relationships between Microsporidia MB infection intensity and these parameters.
The rates of Microsporidia MB infection in the populations of An. gambiae s.s. in Busia (~ 1.4%) and Bungoma (0.3%) were low. Several factors may explain the observed variations in the number of An. gambiae s.s. in the total number of mosquitoes collected from Busia and Bungoma. First, it is likely that seasonality (e.g., the dry and wet seasons in 2020, 2021 and 2022) played a role in the overall abundance of An. gambiae s.s. These highlands areas experience bimodal patterns of rainfall, with peak malaria transmission associated with the long wet season from April to July, while the short wet season is from October to November. Low malaria transmission associated with low vector abundance has been reported for the dry season, which lasts from January to March [22]. Second, as the An. gambiae species complex exhibits great phenotypic plasticity, with a diversity of resting locations and opportunism in blood feeding [15], the collection methods used for the archived samples, such as indoor aspiration and CDC light traps, may have contributed to variation in the determined species composition. Collection methods that solely target mosquitoes that rest indoors can miss individuals that acquire their blood meal indoors but rest outdoors, which may partly explain some of the variation in the total number of individuals of different species captured in this study.
It is notable that the infection rates observed for An. arabiensis were as low as those reported for certain sites in other studies [10]. This suggests that low prevalence could be related to the characteristics of collection sites rather than those of the species of the An. gambiae species complex per se. It is, however, notable that the prevalence rates of Microsporidia MB agree with those recently reported for An. gambiae s.s. and An. coluzzii (~ 1.8%) from Ghana [12]. Overall, these findings indicate that Microsporidia MB does naturally infect An. gambiae s.l. in East Africa. This is significant because An. gambiae s.s. remains an extremely important vector of malaria [13, 15, 20]. Of the three main malaria vectors in western Kenya—An. arabiensis, An. gambiae s.s. and An. funestus s.s.—the latter two predominate in the malaria endemic highlands of western Kenya, and are efficient vectors of malaria due to their endophilic and anthropophilic behaviors [15, 22,23,24,25].
The collection of fewer An. arabiensis in the two areas where sampling was carried out (Additional file 3: Table S1) could be attributed to bias due to our sample collection strategy, which targeted indoor-resting individuals, as this species shows outdoor host-seeking and resting behaviors. In addition, the sites at Busia and Bungoma are in highlands, far from lakes, and in other studies a low number of collected An. arabiensis was attributed to the distance of the sampling site from lakes [14], temporal and spatial variation [16], and/or changes in ecological factors [24]. While sibling species of the genus Anopheles have been found to live in sympatry, An. arabiensis prefers areas with high temperatures and low relative humidity, such as lowlands [12, 16]. The observation that An. gambiae s.l. was the predominant malaria vector in these highland sites of Kenya followed by An. funestus s.l. corroborates previous findings. Although Microsporidia MB was primarily detected in An. gambiae s.s. in the present study, extensive sampling that targets An. funestus in the Kenyan highlands may allow for the detection of Microsporidia MB in these hosts, as this symbiont has been detected in this mosquito species in the lowlands of Ahero [11]. It is possible that scaled-up vector control strategies, such as indoor-based interventions [1, 3], may partly influence mosquito behaviors, including, but not limited to, their preferences for outdoor and/or indoors resting and biting [18]. Thus the use of a single approach for mosquito collection, such as indoor aspiration, may influence the measured species abundance and also the Microsporidia MB positivity rate.
Herren et al. [10] attributed a low Microsporidia MB infection rate in An. arabiensis (0–9%) to several factors. Firstly, the prevalence of Microsporidia MB was found to correlate strongly with season, with infection rates increasing immediately after the rains [10]. While we did not examine seasonal fluctuations, it is possible that changes in the infection rates in An. gambiae s.s. found here may also have been related to the these. Secondly, An. gambiae s.s. from western Kenya have been found to have insecticide-resistance traits and associations with other microbes that may have modulatory effects on their insecticide resistance [26, 27]. Thus, it is possible that the An. gambiae s.s. examined in the present study have multiple insecticide-resistance traits, and the circulation of insecticide-resistant phenotypes of this host may influence Microsporidia MB infection rates in the geographical locations of the present study. Third, changes in the infection rates of maternally inherited symbionts such as Wolbachia have been attributed to changes in temperature, which also affect the densities of specific strains of Wolbachia [28, 29]. Although the effects of temperature on the intensity of Microsporidia MB infection and transmission in mosquito hosts have not been studied, it is possible that chemical interactions similar to those observed in Wolbachia infections [28, 30], or even unique ones, could be associated with the maternal inheritance of symbionts such as Microsporidia MB in mosquitoes. Further investigation of these hypotheses could help to explain why the prevalence of Microsporidia MB in the field remains low despite the observed high transmission rate of Microsporidia MB from field-collected mothers to their offspring.
The results showed that the development of Microsporidia MB in the midgut and reproductive tissues does not impose fitness costs on An. gambiae s.s. Anopheles gambiae s.s. females infected with Microsporidia MB transmitted the infection to their offspring at a rate of approximately 69% (range 0–100%). Low Microsporidia MB intensities were observed in two of the females that did not transmit the infection to their offspring. A high infection intensity of Microsporidia MB was observed in the reproductive tissues of 5- to 7-day-old G1 adults relative to their gut tissues, although in females the intensity of infection in the gut and reproductive tissues did not significantly differ. These results suggest that the gut tissues may be an important reservoir for Microsporidia MB in An. gambiae s.s. A weak positive correlation between Microsporidia MB infection intensities in G0 females and their G1 offspring was observed. It is possible that infection of the reproductive tissues with Microsporidia MB may influence its transmission to offspring, albeit slightly [10], but it is unknown if infection intensities in the midgut are related to the proliferation of Microsporidia MB in reproductive tissues.
Infection of An. gambiae s.s. with Microsporidia MB did not have a negative effect on several of its life history traits. For instance, females infected with Microsporidia MB and non-infected females laid an equivalent number of eggs, and the larvae infected with Microsporidia MB had a marginally significant faster mean development time. Mortality did not differ significantly between Microsporidia MB-infected and non-infected larvae, and infected adults had a similar lifespan to non-infected ones. Whether Microsporidia MB infection results in changes in these parameters when mosquitoes are exposed to nutritional and temperature-related stress is unknown. Previous reports showed that An. arabiensis [10] and An. gambiae s.s. larvae infected with Microsporidia MB had a slightly faster pupal development time than non-infected larvae, but the overall survival rates did not differ between them. Although the larval survival rates of An. gambiae s.s. did not differ between the Microsporidia MB-infected and non-infected individuals in the present study, future studies should investigate if Microsporidia MB infection load influences pupation rate and subsequent survival.
The faster development rate of infected pupae suggests that Microsporidia MB may influence nutrient availability and host metabolic processes [10]. A distortion in the sex ratio favoring females has been previously observed in symbionts that utilize transovarial transmission, such as Dictyocoela microsporidia [31]. The lack of distortion in the sex ratio of An. gambiae s.s. adults infected with Microsporidia MB agrees with previous observations in An. arabiensis [10]. The sex ratio was determined for the emerging adults, but whether there was a similar sex ratio in the eggs laid by females with different infection loads is unknown.
Overall, our findings suggest that Microsporidia MB is avirulent in two species of the An. gambiae species complex, An. arabiensis and An. gambiae s.s., in contrast to other mosquito-associated microsporidians that have been found to be virulent in mosquitoes [32,33,34,35,36,37]. It was shown that Microsporidia MB is transmitted from infected females to their offspring and also horizontally between adults. This is significant, since Microsporidia MB-infected males derived from infected females could be used in a strategy for the dissemination of Microsporidia MB, which would thus avoid the need to release biting females.
Conclusions
The finding that the characteristics of Microsporidia MB infection in An. gambiae s.s. are similar to those reported for An. arabiensis [9, 10] supports the prospect of developing a Microsporidia MB-based malaria transmission-blocking strategy for both of these important vector species. Future studies will need to specifically investigate whether Microsporidia MB protects An. gambiae s.s. against P. falciparum. In addition, the sequencing of Microsporidia MB from both of these species should reveal if there are diverse Microsporidia MB strains that infect different members of the An. gambiae species complex. It would also be of interest to determine if Microsporidia MB can be transmitted between members of the An. gambiae species complex.
Availability of data and materials
All of the datasets supporting the conclusions of this article are included within the article.
Abbreviations
- KEMRI:
-
Kenya Medical Research Institute
- PCR:
-
Polymerase chain reaction
- qPCR:
-
Quantitative polymerase chain reaction
References
Bhatt S, Weiss DJ, Cameron E, Bisanzio D, Mappin B, Dalrymple U, et al. The effect of malaria control on Plasmodium falciparum in Africa between 2000 and 2015. Nature. 2015;526:207–11.
Ochomo E, Bayoh NM, Kamau L, Atieli F, Vulule J, Ouma C, et al. Pyrethroid susceptibility of malaria vectors in four districts of western Kenya. Parasit Vectors. 2014;7:310.
Ranson H, N’Guessan R, Lines J, Moiroux N, Nkuni Z, Corbel V. Pyrethroid resistance in African anopheline mosquitoes: what are the implications for malaria control? Trends Parasitol. 2011;27:91–8.
Dong Y, Manfredini F, Dimopoulos G. Implication of the mosquito midgut microbiota in the defense against malaria parasites. PLoS Pathog. 2009;5:e1000423.
Favia G, Ricci I, Damiani C, Raddadi N, Crotti E, Marzorati M, et al. Bacteria of the genus Asaia stably associate with Anopheles stephensi, an Asian malarial mosquito vector. Proc Natl Acad Sci USA. 2007;104:9047–51.
Bukhari T, Pevsner R, Herren JK. Microsporidia: a promising vector control tool for residual malaria transmission. Front Trop Dis. 2022;3:1–18.
Hoffmann AA, Montgomery BL, Popovici J, Iturbe-Ormaetxe I, Johnson PH, Muzzi F, et al. Successful establishment of Wolbachia in Aedes populations to suppress dengue transmission. Nature. 2011;476:454–9.
Frentiu FD, Zakir T, Walker T, Popovici J, Pyke AT, van den Hurk A, et al. Limited dengue virus replication in field-collected Aedes aegypti mosquitoes infected with Wolbachia. PLoS Negl Trop Dis. 2014;8:1–10.
Ant TH, Herd CS, Geoghegan V, Hoffmann AA, Sinkins SP. The Wolbachia strain wAu provides highly efficient virus transmission blocking in Aedes aegypti. PLoS Pathog. 2018;14:e1006815.
Herren JK, Mbaisi L, Mararo E, Makhulu EE, Mobegi VA, Butungi H, et al. A microsporidian impairs Plasmodium falciparum transmission in Anopheles arabiensis mosquitoes. Nat Commun. 2020;11:2187.
Nattoh G, Maina T, Makhulu EE, Mbaisi L, Mararo E, Otieno FG, et al. Horizontal transmission of the symbiont Microsporidia MB in Anopheles arabiensis. Front Microbiol. 2021;12:647183.
Akorli J, Akorli EA, Naa S, Tetteh A, Atibilla D, Bansa SP, et al. Microsporidia MB is found predominantly associated with Anopheles gambiae s.s. and Anopheles coluzzii in Ghana. Sci Rep. 2021;11:18658.
Hemming-Schroeder E, Zhong D, Machani M, Nguyen H, Thong S, Kahindi S, et al. Ecological drivers of genetic connectivity for African malaria vectors Anopheles gambiae and An. arabiensis. Sci Rep. 2020;10:19946.
Bayoh MN, Mathias DK, Odiere MR, Mutuku FM, Kamau L, Gimnig JE, et al. Anopheles gambiae: historical population decline associated with regional distribution of insecticide-treated bed nets in western Nyanza Province, Kenya. Malar J. 2010;9:1–12.
Sinka ME, Bangs MJ, Manguin S, Coetzee M, Mbogo CM, Hemingway J, et al. The dominant Anopheles vectors of human malaria in Africa, Europe and the Middle East: occurrence data, distribution maps and bionomic précis. Parasit Vectors. 2010;3:117.
Takken W, Smallegange RC, Vigneau AJ, Johnston V, Brown M, Mordue-luntz AJ, et al. Larval nutrition differentially affects adult fitness and Plasmodium development in the malaria vectors Anopheles gambiae and Anopheles stephensi. Parasit Vectors. 2013;6:345.
Githinji EK, Irungu LW, Ndegwa PN, MacHani MG, Amito RO, Kemei BJ, et al. Species composition, phenotypic and genotypic resistance levels in major malaria vectors in Teso North and Teso South sub-counties in Busia County, western Kenya. J Parasitol Res. 2020;2020:1–17.
Machani MG, Ochomo E, Amimo F, Kosgei J, Munga S, Zhou G, et al. Resting behaviour of malaria vectors in highland and lowland sites of western Kenya: implication on malaria vector control measures. PLoS ONE. 2020;15:e0224718.
Gillies M, Coetzee M. A supplement to the Anophelinae of Africa south of the Sahara (Afrotropical region). South African Inst Med Res. 1987;55:1987.
Santolamazza F, Mancini E, Simard F, Qi Y, Tu Z, della Torre A. Insertion polymorphisms of SINE200 retrotransposons within speciation islands of Anopheles gambiae molecular forms. Malar J. 2008;7:163.
Dimopoulos G, Richman A, Müller HM, Kafatos FC. Molecular immune responses of the mosquito Anopheles gambiae to bacteria and malaria parasites. Proc Natl Acad Sci USA. 1997;94:11508–13.
Zhou G, Afrane Y, Vardo-Zalik A, Atiel I, Zhong D, Et A. Changing patterns of malaria epidemiology between 2002 and 2010 in western Kenya: the fall and rise of malaria. PLoS ONE. 2011;6:e20318.
Doumbe-Belisse P, Kopya E, Ngadjeu CS, Sonhafouo-Chiana N, Talipouo A, Djamouko-Djonkam L, et al. Urban malaria in sub-Saharan Africa: dynamic of the vectorial system and the entomological inoculation rate. Malar J. 2021;20:364.
Ndenga B, Githeko A, Omukunda E, Munyekenye G, Atieli H, Wamai P, et al. Population dynamics of malaria vectors in western Kenya highlands. J Med Entomol. 2006;43:200–6.
Mwangangi J, Mbogo C, Orindi B, Muturi E, Midega J, Nzovu J, et al. Shifts in malaria vector species composition and transmission dynamics along the Kenyan coast over the past 20 years. Malar J. 2013;12:13.
Kawada H, Dida GO, Ohashi K, Komagata O, Kasai S, Sonye G, et al. Multimodal pyrethroid resistance in malaria vectors, Anopheles gambiae s.s., Anopheles arabiensis, and Anopheles funestus s.s. in western Kenya. PLoS ONE. 2011;6:e22574.
Omoke D, Kipsum M, Otieno S, Esalimba E, Sheth M, Lenhart A, et al. Western Kenyan Anopheles gambiae showing intense permethrin resistance harbour distinct microbiota. Malar J. 2021;20:77.
Ross PA, Axford KJ, Yang Q, Staunton KM, Ritchie SA, Richardson KM, et al. Heatwaves cause fluctuations in wMel Wolbachia densities and frequencies in Aedes aegypti. PLoS Negl Trop Dis. 2020;12:e0007958.
Mancini MV, Ant TH, Herd CS, Martinez J, Murdochy SM, Gingell DD, et al. High temperature cycles result in maternal transmission and dengue infection differences between Wolbachia strains in Aedes aegypti. MBio. 2021;12:e00250-21.
Carvajal TM, Hashimoto K, Harnandika RK, Amalin DM, Watanabe K. Detection of Wolbachia in field-collected Aedes aegypti mosquitoes in metropolitan Manila, Philippines. Parasit Vectors. 2019;12:361.
Ironside JE, Alexander J. Microsporidian parasites feminise hosts without paramyxean co-infection: support for convergent evolution of parasitic feminisation. Int J Parasitol. 2015;45:427–33.
Andreadis TG, Hall DW. Development, ultrastructure, and mode of transmission of Amblyospora sp. (Microspora) in the mosquito. J Protozool. 1979;26:444–52.
Becnel JJ. Host regulated developmental mechanisms in vector arthropods. Gainesville: University of Florida; 1993. p. 240–5.
Fox RM, Weiser JA. Microsporidian parasite of Anopheles gambiae in Liberia. J Parasitol. 1959;45:21.
Haq N, Reisen WK, Aslamkhan M. The effects of Nosema algerae on the horizontal life table attributes of Anopheles stephensi under laboratory conditions. J Invertebr Pathol. 1981;37:236–42.
Hulls RH. The adverse effects of a microsporidan on sporogony and infectivity of Plasmodium berghei. Trans R Soc Trop Med Hyg. 1971;65:421–2.
Ward RA, Savage KE. Effects of microsporidian parasites on anopheline mosquitoes and malarial infection. Proc Helminthol Soc Wash. 1972;39:434–8.
Acknowledgements
We acknowledge and thank the homeowners in the villages of Kimaeti Bungoma and Akiriamas/Akiriamasit Teso for allowing access to their homes during the mosquito collection, without which this study would not have been possible. We are grateful to Martin Kennedy and Richard Amito for their assistance with planning and the mosquito collections. We are grateful to the director general of KEMRI for permission to publish this work.
Funding
The authors gratefully acknowledge financial support for this research from the following organizations and agencies: Open Philanthropy (SYMBIOVECTOR Track A); the Bill and Melinda Gates Foundation (INV0225840); the Children’s Investment Fund Foundation (SMBV-FFT); the ANTi-VeC network (grant no. AV/AASS/006); the International Foundation for Science, Stockholm, Sweden (through grant no. I-1-F-5852-1 awarded to GN); the Swedish International Development Cooperation Agency; the Swiss Agency for Development and Cooperation; the Australian Centre for International Agricultural Research; the Federal Democratic Republic of Ethiopia; and the government of the Republic of Kenya. The views expressed herein do not necessarily reflect the official opinions of the donors.
Author information
Authors and Affiliations
Contributions
GN and JKH conceived and designed the study. GN and BO conducted the field collection of specimens, assisted by JKH and EO. JKH and EO provided resources for the molecular analysis. GN and BO performed the molecular analyses, and were assisted in this by DO, EMM, LM, and MMG. JKH, EO and LK supervised the work. GN drafted the original manuscript. GN, EO, LK, JKH led the interpretation of the findings. All of the authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
KEMRI undertook the ethical review and granted ethical approval for this work [KEMRI/Scientific and Ethics Review Unit (SERU)/Centre for Biotechnology Research and Development (CBRD)/230/4341]. The community leaders and village participants enrolled in the sample collection were provided with a detailed explanation of the study objectives, activities, procedures, benefits and risks before its commencement. Through the participation of the community leaders we were able to obtain informed consent from the owners of the households where samples were collected. Participation was on a voluntary basis, and participants were free to withdraw from the study whenever they wished.
Consent for publication
Not applicable.
Competing interests
All authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1
: Figure S1. Sex ratio of Microsporidia MB-infected Anopheles gambiae s.s. progeny reared in pools. A Bar plots representing the sex ratio of Microsporidia MB-infected offspring and non-infected counterparts reared in pools (n = 6, where a pool consists of approximately seven individual females). B Bar plots representing the mean prevalence of Microsporidia MB transmitted to An. gambiae s.s. offspring, reared in pools, by females. Error bars represent the SEM percentage (* P < 0.05).
Additional file 2: Figure S2.
Microsporidia MB infection intensities in An. gambiae s.s. influence the rate of transmission to offspring. A Correlation of Go Microsporidia MB intensities with average G1 Microsporidia MB densities [r2 = 0.1276, P = 0.268, number of broods = 18]. B Correlation between G0 Microsporidia MB intensities and Microsporidia MB transmission to offspring (r2 = 0.625, P = 0.0056, n = 18).
Additional file 3
: Table S1. Annual prevalence of Microsporidia MB in Anopheles gambiae s.l. Values indicate yearly (2021–2022) abundance (%) of Microsporidia MB assessed from the DNA of individual female mosquitoes sampled from Bungoma and Busia.
Additional file 4
: Table S2. Horizontal transmission of Microsporidia MB between Anopheles gambiae s.s. adults reared together in cages. Values indicate intensities of Microsporidia MB infection in donor male/female and recipient male/female An. gambiae s.s. individuals reared together in a cage.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Nattoh, G., Onyango, B., Makhulu, E.E. et al. Microsporidia MB in the primary malaria vector Anopheles gambiae sensu stricto is avirulent and undergoes maternal and horizontal transmission. Parasites Vectors 16, 335 (2023). https://doi.org/10.1186/s13071-023-05933-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-023-05933-8