
Abstract
The Philadelphia negative myeloproliferative neoplasms (MPN) compromise a heterogeneous group of clonal myeloid stem cell disorders comprising polycythaemia vera, essential thrombocythaemia and primary myelofibrosis. Despite distinct clinical entities, these disorders are linked by morphological similarities and propensity to thrombotic complications and leukaemic transformation. Current therapeutic options are limited in disease-modifying activity with a focus on the prevention of thrombus formation. Constitutive activation of the JAK/STAT signalling pathway is a hallmark of pathogenesis across the disease spectrum with driving mutations in JAK2, CALR and MPL identified in the majority of patients. Co-occurring somatic mutations in genes associated with epigenetic regulation, transcriptional control and splicing of RNA are variably but recurrently identified across the MPN disease spectrum, whilst epigenetic contributors to disease are increasingly recognised. The prognostic implications of one MPN diagnosis may significantly limit life expectancy, whilst another may have limited impact depending on the disease phenotype, genotype and other external factors. The genetic and clinical similarities and differences in these disorders have provided a unique opportunity to understand the relative contributions to MPN, myeloid and cancer biology generally from specific genetic and epigenetic changes. This review provides a comprehensive overview of the molecular pathophysiology of MPN exploring the role of driver mutations, co-occurring mutations, dysregulation of intrinsic cell signalling, epigenetic regulation and genetic predisposing factors highlighting important areas for future consideration.
Similar content being viewed by others

Introduction
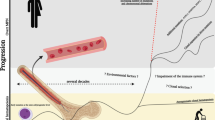
The classical Philadelphia chromosome negative myeloproliferative neoplasms (MPN) are rare clonal neoplastic disorders of the myeloid haematopoietic stem cells (HSC). These disorders are classified into polycythaemia vera (PV) with a predominance of excessive red cell production, essential thrombocythaemia (ET) with a predominance of excessive platelet production and primary myelofibrosis (PMF) with excessive bone marrow scarring and fibrosis. The updated WHO classification also includes pre-fibrotic myelofibrosis (Pre-PMF), distinguishing a group of patients with subtle phenotypic differences from ET and a higher rate of progression to myelofibrosis (MF) [1]. Prognosis is highly variable, but in general, MF significantly limits life expectancy in comparison to PV or ET. A small number of individuals progress to blast phase of disease presenting as acute myeloid leukaemia which is frequently refractory to conventional therapy. Figure 1 characterises the distinguishing clinical features commonly observed in each MPN manifestation.

MPN Heterogeneity. A figure demonstrating the distinct clinical entities observed in MPN patients with a summary of distinguishing clinical features observed in each. *Propensity to enhanced rates of clot formation in ET appear to be variable depending on driver mutation status. **Bleeding can manifest in a minority of ET patients resulting from acquired Von Willebrand disease resulting from very high platelet counts
Despite the obvious differences, similarities in bone marrow morphology, a tendency to arterial and venous thrombus formation and a tendency to secondary myelofibrotic or leukaemic phase transformation links these disorders clinically. These phenotypic similarities had been identified well in advance of the discovery of activating mutations in the JAK2, MPL and CALR genes and the demonstration of activated Janus Kinase (JAK)/signal transducer and activator (STAT) signalling pathway signalling which has helped to further define these disorders [2,3,4,5,6,7,8]. JAK2 V617F mutations are detectable in approximately 95% of PV patients with JAK2 exon 12 mutations present in virtually all remaining PV cases [3,4,5]. The JAK2 V617F mutation is present in approximately 50% of ET and PMF patients with CALR and MPL mutations present in most remaining patients [8, 9]. “Triple negative” patients make up a small percentage of ET and PMF cases. Diagnostic criteria now place a heavy emphasis on demonstrating the presence of these genetic changes to confirm a suspected diagnosis as is demonstrated in Table 1 [10].
With modern diagnostic approaches, it is increasingly clear that the desire for neat classification is often complicated by a spectrum of phenotypic presentation and genetic heterogeneity. A range of co-occurring somatic mutations are frequently detectable at significant variant allele frequencies alongside the JAK2, MPL or CALR mutations [13,14,15]. Complex clonal hierarchies have been observed within MPN patients [16]. These frequently observed co-occurring mutations include genes encoding epigenetic modifiers, transcriptional regulators and mRNA splicing machinery. They are not exclusive to MPN but rather occur across the spectrum of myeloid malignancy [17, 18]. Further complicating the picture, many of these mutations, including JAK2 V617F, are increasingly detected in individuals as we age yet with the majority demonstrating no haematological disease phenotype [19]. This clonal haematopoiesis of indeterminate potential (CHIP) unsurprisingly pre-disposes to the development of myeloid malignancy but appears to also be sufficient to significantly increase cardiovascular risk [19,20,21].
Current therapeutic approaches in MPN aim to limit the risk of thrombosis with antiplatelet agents, anticoagulants, therapeutic venesection and cytoreductive therapies including hydroxycarbamide and interferon-alpha all with established benefits in specific circumstances [22,23,24]. JAK inhibitors including ruxolitinib have provided an additional targeted therapy with clear symptomatic and clinical benefits but limited disease-modifying activity [25,26,27,28]. Haematopoietic stem cell transplantation offers the only opportunity for cure but is rarely suitable due to the significant toxicities and mortality risk associated. It is generally reserved for younger, fitter individuals with higher-risk myelofibrosis or blast phase disease, and outcomes remain poor in these populations [29, 30].
The clinical and genetic similarities and differences in this heterogeneous population offer the opportunity to characterise and elucidate the contributions of various genetic and epigenetic factors to disease pathogenesis. Enhanced availability of such genetic and phenotypic data has meanwhile provided the opportunity to generate individualised prognosis probabilities to MPN patients [15]. This review of current understanding of the molecular pathogenesis of MPN will focus on the role of JAK/STAT and other intracellular signalling pathways, acquired and inherited genetic contributors to disease, epigenetic dysregulation and cellular context and will highlight areas for future research considerations.
JAK/STAT signalling in MPN
The evolutionarily conserved JAK/STAT pathway exists as a critical intracellular mediator of extracellular protein–cell surface receptor interactions. Four genes for JAK proteins exist in the human genome (JAK1, JAK2, JAK3 and TYK2) interacting with seven STAT proteins to mediate differential effects on transcriptional control. JAK proteins associate with numerous cell surface receptors, and thus, JAK/STAT signalling cascades are activated in many metabolic functions, immune cell functions and control of haematopoiesis [31]. Effective control of erythropoiesis, megakaryopoiesis and granulopoiesis is essential to respond to changing physiological demands throughout life and in times of physiological stress or infection. Hormonal signalling with erythropoietin (EPO), thrombopoietin (TPO) and granulocyte-colony stimulating factor (GCSF) drive enhanced production of red cells, platelets and granulocytes through the respective receptors. Activation of the erythropoietin receptor (EPOR), thrombopoietin receptor (MPL) and granulocyte-colony stimulating factor receptor (G-CSFR) then activate JAK/STAT pathways to drive proliferation. JAK/STAT signalling is heavily interconnected with many core cancer signalling pathways and cellular functions including metabolism, cell cycle control, apoptosis, DNA damage response and direct or indirect transcriptional control [32]. Abnormal JAK/STAT signalling has been implicated across a range of myeloid, B and T lymphoid haematological malignancies and solid tumours [33,34,35,36].
Driver mutations activating JAK/STAT signalling
In MPN, constitutive activation of the JAK/STAT signalling pathway is a critical mediator of the pathogenesis. A point mutation in exon 14 of the JAK2 gene results in a single amino acid (valine to phenylalanine) substitution and conformational change in the JH2 pseudo-kinase domain of JAK2. This results in constitutive tyrosine phosphorylation activity by disrupting the normal inhibitory action of the JH2 domain. This JAK2 V617F transcript therefore drives constitutive activation of the JAK/STAT pathway in the absence of EPOR, MPL or G-CSFR ligand binding. This mutation is detected in 95% of PV patients and approximately 50% of ET and PMF patients [5, 37, 38]. The resulting disease phenotype is subject to several additional variables including homo or heterozygosity of the JAK2 V617F, variant allele frequency, additional co-operating mutations and/or external influences including iron deficiency. The remaining 5% of PV patients are almost entirely accounted for by mutations in exon 12 of the JAK2 gene through predominant activation of EPOR signalling pathways driving an erythrocytosis [4]. These JAK2 exon 12 mutations have not been seen in PMF or ET.
The majority of JAK2 V617F negative ET and PMF patients have detectable mutations in MPL or CALR [7, 8, 38]. These mutations drive disease through activation of MPL receptor and subsequent downstream JAK/STAT activation. Generally, the driver mutations occur in a mutually exclusive manner. A number of activating MPL mutations have been identified in the transmembrane domain encoded by exon 10 in both familial and sporadic forms of MPN [8, 39]. These gains of function mutations including W515L and S505N constitutively activate downstream JAK/STAT signalling by removing an inhibitory element and inducing dimerization, respectively [39, 40]. Additional activating or augmenting mutations identified in MPL transmembrane domain by deep mutational scanning screens have also been previously identified in MPN patients demonstrating an inherent susceptibility in the MPL gene [41].
Calreticulin (CALR) is an endoplasmic reticulum chaperone protein which in mutant form will interact directly with the TPO receptor MPL driving TPO independent activation. Numerous mutations in the CALR gene have been described with the majority classed as type 1 resulting from a 52-bp deletion in exon 9 or type 2 with a 5-bp insertion in exon 9. The subsequent activation of MPL and downstream JAK/STAT signalling is dependent on a positively charged C terminus resulting from a frameshift in exon 9 and enabling the CALR lectin binding domain to maintain a stable interaction with MPL [42]. There is a recognised distinct clinical phenotype between patients with detectable type 1 and type 2 CALR mutations in both patients and murine models. Type 1 deletions are significantly over-represented in myelofibrosis and produce a more pronounced MPN phenotype in mice [43].
A small minority of ET and PMF patients fall into the “triple negative” category with no detectable mutation in JAK2, MPL or CALR. The incorporation of additional genetic tests into the work-up of these patients has challenged the notion of true triple negativity in MPN. Retrospective evaluations demonstrate that some of these patients may have other genetic markers of clonality detectable or subsequently test positive for a driver mutation. A few patients remain with characteristic phenotypic and morphological features and no detectable genetic abnormalities [44].
Frequently, these JAK/STAT activating driver mutations are the only detected genetic abnormality in MPN patients with one large study reporting this in 45% of patients using a targeted myeloid next-generation sequencing (NGS) panel [15]. The presence of JAK2, CALR or MPL mutations alone are sufficient to generate an MPN phenotype, albeit polyclonal in nature, in murine models [45]. One study of gene expression profiling by microarray analysis demonstrated features of activated JAK/STAT signalling in MPN patients regardless of clinical phenotype or mutational status [2]. Ruxolitinib, a JAK1/2 inhibitor, is effective across all mutant driver backgrounds [46]. It is therefore clear that constitutively activated JAK/STAT signalling is a key feature of disease pathogenesis.
The presence of the JAK2 V617F mutation is also evidently more than a simple switch for excess proliferation. There is significant heterogeneity in terms of the variant allele frequency (VAF), and therefore, clonal size measured in peripheral blood granulocytes is detectable across the MPN patient population [47]. Patients with homozygosity or high VAF tend towards a PV phenotype rather than ET [48, 49]. And yet, there are many PV patients with a low VAF and similarly ET patients with high VAF. A rare subgroup of patients presenting with splanchnic vein thrombosis (SVT) frequently exhibit normal or near normal blood counts with a small JAK2 V617F clone detectable [50]. The JAK2 V617F mutation is also detectable in individuals with CHIP who exhibit no MPN phenotype. Despite the normal blood counts, these individuals have been observed to have a significantly increased risk of cardiovascular disease [20]. Paradoxically, JAK2 V617F positive ET cases are significantly more likely to have thrombotic complications despite lower platelet counts than their CALR mutated comparators [51]. One suggestion in these cases is that the mutant JAK2 results in qualitative changes enhancing the pro-thrombotic phenotype. There is evidence to suggest enhancing endothelial–erythrocyte interactions via activation of Lu/BCAM or enhanced neutrophil extracellular trap formation [52, 53]. A recent study has demonstrated that in patients with a low VAF (< 20%) and therefore small mutant clones within the peripheral blood granulocytes, there is significant heterogeneity of clone size within the reticulocytes and platelets measured using a quantitative polymerase chain reaction established to measure JAK2 V617F RNA. In many cases, the clonal sizes in the reticulocytes and platelet populations were much higher than the granulocytes perhaps from late expansion of erythroid and platelet precursors [54]. We may not therefore have been accurately assessing clonal size in many of our “low allele” patients, and granulocyte VAF may underestimate the qualitative effect of the mutant JAK2 presence.
STAT proteins in MPN
The complexity of STAT signalling has identified roles for STAT proteins in oncogenesis and tumour suppression, occasionally with conflicting roles in the same tumour type [55]. Investigations of STAT protein recruitment, phosphorylation and ultimately dominance of transcriptional control in MPN have focused on the role of STAT5, STAT1 and STAT3. STAT5 activation was identified early as a key mediator of MPN pathogenesis with experimental work able to demonstrate a dependence on STAT5 to generate a MPN phenotype [56, 57]. STAT5 transcriptional activity is upregulated by the expression of JAK2 V617F in cell lines [37]. In analysis of ex vivo colony forming assays from ET and PV patients, transcriptional analysis demonstrated an enrichment of STAT5A targets with nuclear phosphorylation of STAT5A identified in JAK2 V617F position colonies from both ET and PV patients but not wild type colonies whilst a recent phospho-proteomics approach identified STAT5 and STAT3 as differentially phosphorylated in JAK2 V617F mouse haematopoietic cells [58, 59]. Conditional expression of a null STAT5a/b gene resulted in a failure of a JAK2 V617F mouse model to develop polycythaemia but did not abrogate the risk of myelofibrosis [60]. In an alternative JAK2 V617F mouse model STAT5 deletion resulted in loss of the PV phenotype which could be rescued by STAT5 re-expression [61]. Taken together, STAT5 signalling appears to play a key mediator role in generating the PV phenotype.
Identification of enhanced enrichment of Interferon-gamma target genes in ET in comparison to PV highlighted STAT1 signalling as a potential mediator of differential molecular response between the two disorders. In keeping with this, phosphorylated STAT1 was detectable in ET patients and not PV patients ex vivo [58]. Murine models subsequently demonstrated the loss of STAT1 producing a phenotype favouring erythropoiesis at the expense of megakaryopoiesis and with a reduction in fibrosis [62]. STAT1 phosphorylation at serine 727 may drive proliferation and restrain megakaryocyte differentiation in blast phase MPN. Blocking this serine phosphorylation resulted in different functional outcomes in comparison to blocking tyrosine phosphorylation with ruxolitinib [63]. These results have suggested that an altered balance between STAT1 and STAT5 signalling may be one possible cell intrinsic mechanism of phenotype determination. However, erythroblasts harbouring the JAK2 exon 12 mutations which drive erythrocytosis and are only associated with PV have a transcriptional profile which cannot be distinguished from JAK2 V617F positive ET erythroblasts with no differential in STAT1 activation [64].
Constitutive activation of STAT3 was identified in a number of MPN patients from granulocytes in advance of the discovery of JAK2 V617F, whilst higher levels of STAT3 tyrosine phosphorylation have been identified in JAK2 V617F positive individuals and as a result of JAK2 V617F expression in murine models [65,66,67]. A murine model of STAT3 hyperactivity induced by deletion of suppressor of cytokine signalling (SOCS) 3 spontaneously develops myeloproliferative and lymphoproliferative pathology with serine phosphorylation of STAT3 critical [68]. STAT3 deletion results in an altered MPN phenotype in JAK2 V617F mice with reduced neutrophilia and enhanced thrombocytosis present [69].
The canonical tyrosine phosphorylation, nuclear translocation and transcription factor activity are only one role of the STAT proteins. Unphosphorylated STAT proteins appear to have important roles in the normal maintenance of the epigenome in HSC and progenitor cells [70]. As mentioned, serine phosphorylation of STATs may affect transcriptional control and the methylation of STAT3 by Enhancer of Zeste 2 (EZH2) identified as a mediator of transcriptional control in other solid tumours requires investigation in MPN to fully understand the dynamics at play between STAT proteins and final phenotypes [63, 68, 71]. Therefore, as with all aspects of molecular biology in MPN, differential STAT1/STAT3/STAT5 mobilisation is likely to tell only part of the story.
Non-JAK/STAT signalling in MPN
Outside of JAK/STAT signalling, it is increasingly evident that activation of STAT independent phosphoinositide 3-kinase (PI3K) and mitogen-activated protein kinase (MAPK) signalling pathways is important in the disease pathogenesis of JAK2 V617F positive MPN [37]. CALR mutations have also been observed to activate MAPK signalling pathways albeit with a differential expression profile evident in comparison to JAK2 V617F [72, 73]. There is evidence that either or both signalling of these pathways may remain active in the presence of the JAK inhibitor ruxolitinib. Murine MPN models with JAK2 V617F and MPL W515L drivers have demonstrated persistent activation of the MAPK mediated by platelet-derived growth factor receptor alpha (PDGFRα) in vivo in the setting of ruxolitinib exposure. Combined JAK/MEK inhibition in this model was more efficacious [74]. Persisting phosphorylation of serine residues on STAT5B has been observed in the JAK2 V617F positive SET2 cell line model dependent on PI3K/mechanistic target of rapamycin (mTOR) activation with enhanced efficacy again observed when combining JAK inhibition with either PI3K or mTOR inhibitors [75]. Early phase trials of everolimus, an mTOR inhibitor, in MF patients have previously demonstrated some clinical benefit [76]. Activated MAPK, PI3K/AKT and JAK/STAT signalling are also observed in numerous myeloid malignant phenotypes including acute myeloid leukaemia (AML), chronic myeloid leukaemia (CML), atypical CML, chronic myelomonocytic leukaemia and juvenile myelomonocytic leukaemia [77]. Clearly, improving our understanding of the intricacies of dysregulated signalling cascade activation in MPN patients and the effect of treatment may offer some opportunity to manipulate these processes for more efficacious treatments in select individuals.
Negative regulation of intracellular signalling in MPN
In normal health, intracellular signalling cascades are closely regulated positively by ligand binding to cell surface receptors and negatively by a number of intracellular components acting predominantly as phosphatases or targeting proteins for ubiquitination and subsequent proteasome-mediated degradation. The SOCS proteins are critical negative regulators for JAK/STAT signalling. SOCS3 is a key negative regulator of EPO signalling and therefore erythropoiesis through its interaction with JAK2 and EPOR. There is conflicting evidence on the role of SOCS proteins in the regulation of mutant JAK2. One study has demonstrated that in the presence of the JAK2 V617F mutant, SOCS3 undergoes tyrosine phosphorylation, losing the ability to negatively regulate JAK2 and may in fact potentiate the effect by stabilising the mutant JAK2 [78]. A more recent investigation has demonstrated that mutant JAK2 exhibits comparable inhibition by SOCS3 in in vitro kinase assays [79]. Knockdown of SOCS3 in JAK2 V617F expressing Ba/F3 cells further enhances phosphorylation of STAT3, STAT5 and extracellular signal-regulated kinase 1/2 (ERK1/2) suggesting a tumour suppressor role for SOCS3 in the mutant state [80]. One small study has suggested aberrant regulation of SOCS3 expression in PMF with observation that SOCS3 promoter regions were methylated in 32% of PMF patients but not ET or PV patients [81]. The JAK inhibitor ruxolitinib can clearly reduce the expression of SOCS3 in a number of cellular contexts outside of MPN [82, 83] and we and others have shown that RNA sequencing data confirm reduced expression in MPN cell line models [84, 85]. Histone deacetylase inhibition may upregulate SOCS3 expression in MPN [86,87,88].
CBL encodes the E3 ubiquitin ligase Cbl proto-oncogene (CBL) and is an important regulator of many tyrosine kinases. CBL mutations have been identified in many malignancies and at a low but significant level in MPN [15]. These mutations drive myeloproliferation and result in activated JAK/STAT and PI3K/AKT signalling in murine models [89]. In addition, loss of specific phosphatase activity of phosphatase and tension homolog deleted on chromosome 10 (PTEN), a regulator of PI3K/AKT signalling can drive an MPN phenotype [90]. Conversely, higher expression of dual-specificity phosphatase 1 (DUSP1) may be required in the JAK2 V617F context to protect the cells by moderating JNK/P38 MAPK signalling and protecting against accumulating DNA damage [91].
Figure 2 highlights active intracellular signalling cascades identified in MPN and examples of intracellular negative feedback mechanisms. Understanding the complexities of this intrinsic regulation of intracellular signalling in vivo in the heterogenous cellular contexts of MPN patients may help to improve our understanding of pathogenesis and treatment options.

Activated signalling in MPN. Activated signalling pathways in MPN include JAK/STAT signalling, PI3K/AKT and MAPK (RAS/MEK/ERK) cascades. The balance of predominant STAT1/3/5 signalling may impact on ultimate disease phenotype, whilst persistence of activated MAPK or PI3K signalling or STAT serine phosphorylation may occur despite JAK inhibitor therapy. Examples of intracellular negative regulators of JAK/STAT, PI3K/AKT and MAPK signalling with potential roles in MPN pathophysiology are also shown
Co-occurring mutations and clonal evolution in MPN
With the increasing availability of genetic sequencing in the research and now routine diagnostic setting, the genetic heterogeneity of the MPN group has become increasing clear. A number of pathological mutations are frequently and recurrently identified in MPN patients across a range of genes affecting epigenetic regulation, transcriptional control and splicing machine. These genes commonly include ASXL1, DNMT3A and TET2 at relatively high frequencies in upwards of 5% of patient samples across the MPN spectrum. Others including CBL, SF3B1, EZH2, TP53, SRSF2, USAF1 and IDH1/2 are identified in fewer than 2% of patients in large studies [15]. These mutations are regularly identified in other individuals across the range of myeloid malignancy and in the CHIP population [92,93,94]. These mutations which have been well characterised represent only a small proportion of the overall mutational burden seen as aging progresses in MPN patients when whole genome sequencing approaches are employed [95].
Prognostic implications
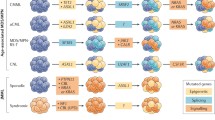
As larger cohorts of individuals continue to be analysed it is clear that the presence of particular mutations is associated both with the disease phenotype and ultimate prognosis. Chronic phase PV and ET patients are significantly more likely to have no additional mutations when compared to PMF patients. The occurrence of particular genes has been significantly associated with disease phenotype. For example, NFE2 mutations correlate with a PV phenotype, spliceosome component mutants and epigenetic regulators EZH2 and ASXL1 are more frequently observed in PMF, whilst other genes including IKZF1 are almost exclusively observed in blast phase disease [15, 96, 97]. Early studies demonstrated clear prognostic implications of particular co-operating mutations. In a cohort of 483 European PMF patients with validation in an American group, ASXL1, EZH2 and SRSF2 mutations independently predicted shortened survival. IDH1/2 or SFSF2 mutations were associated with leukaemic progression in these cohorts with TP53 strongly associated in another cohort [98, 99]. The negative prognostic impact of ASXL1 appears to be evident when present with another high risk mutation but not solely on its own [100]. In a large study of 2035 patients, eight genomic subgroups were identified within the MPN spectrum, each reflecting a different proportion of PV, ET and PMF patients with variable risks of leukaemic or fibrotic transformation and overall survival. Genetic factors including TET2, SRSF2 and ASXL1 mutations contributed to over 50% of the risk factors for fibrotic transformation from PV or ET using a predictive modelling approach. Similarly, over a third of the risk of leukaemic transformation was attributable to genetic factors including TP53 mutation [15]. Figure 3 demonstrates mutated genes identified as contributors to leukaemic and fibrotic transformation from this predictive modelling approach integrated with results from multivariate analysis approaches in other large cohorts [15, 98, 101, 102]. Individualised prognostic evaluations can now be achieved at diagnosis based on the incorporation of genetic and clinical information, and molecular data are now routinely incorporated into prognostic scoring systems including MIPPS70 + and GIPSS [101, 103].

Mutational significance in MPN for overall survival. A summary of determined mutational significance in MPN. The central figure shows genetic contributors to fibrotic (inner yellow ring) and leukaemic (outer red ring) transformation in MPN determined by a predictive modelling approach in 1599 chronic phase and 276 myelofibrosis patients [15]. Underlined genes were contributors to death in myelofibrosis patients in this modelling. Overlaid are a purple ring (A) showing independent risk factors for survival identified in 641 myelofibrosis patients [101], a green ring (B) showing independent risk factors for survival identified in 483 myelofibrosis patients and validated in a further 396 patients [98], and finally a grey ring (C) showing prognostically detrimental genes in 537 myelofibrosis patients and validated in a further 260 patients [102]
Clonal structure and evolution
The variant allele frequency reported by next-generation sequencing (NGS) myeloid panels provides a rough estimate of clonal size and dominance, reflecting both the number of affected cells and whether these cells are hetero or homozygous for the mutation of interest. Variant allele frequencies of driver and co-occurring mutations are seldom identical, reflecting a clonal hierarchy comprising dominant and sub-clones with differing genetic abnormalities and resulting competitive advantages. Application of single-cell technologies in myeloproliferative patients has demonstrated complex clonal hierarchies with dominant and sub-clones demonstrating distinctive transcriptomic signatures within individual patients [16]. The clonal complexity is greater in MPN in comparison to CHIP patients but much less than that observed in AML [104].
This complexity reflects clonal evolution over time, occurring as cells acquire additional genetic or epigenetic changes driving further divergence from the cell of origin. This can significantly alter the clonal structure of the disease with smaller sub-clones developing the competitive advantage to establish dominance. Patients progressing to myelofibrosis and AML often demonstrate greater genetic complexity within clones with increasing numbers of mutations detectable. Ultimately, this results in transcriptional and functional changes which are observed in patients cells underlying the progressive disease phenotype [105]. In chronic phase MPN, the rate of acquisition of new mutations is generally considered to be a slow process. Only two additional mutations were observed over the equivalent of greater than 130 patient years in one study [99]. However, this generalisation, like many, does not reflect the significant heterogeneity across the MPN patient spectrum with both acquisition of new clones resulting from new mutations or minor sub-clones establishing dominance observed in patients transforming from chronic phase to blast phase disease [104, 106]. Meanwhile, approximately one-third of PMF individuals receiving the JAK inhibitor ruxolitinib demonstrated clonal evolution on therapy acquiring new mutations [107]. Perhaps as our repositories of sequential pre- and post-MPN progression sequencing data increases, we may understand the genetic, epigenetic and extrinsic factors explaining the propensity for clonal evolution in a number of individuals.
Recent comprehensive genomic profiling with a whole genome sequencing approach and phylogenetic reconstruction based on the numerous somatic mutations occurring during life have provided evidence that acquisition of both driver and common co-occurring mutations may occur many decades prior to disease presentation. In a number of cases, the acquisition of JAK2 V617F or DNMT3A mutations were predicted to have occurred in utero. This approach reveals a significant variability in the rates of clonal expansion, dependent on the mutational landscape of the clone [95]. A concept of mutational compatibility can be theorised from much of the genetic data available in myeloid malignancy to date. A number of individual mutations are frequently observed together, whilst others appear strictly mutually exclusive of each other [99]. In some cases, this mutational compatibility clearly drives a competitive advantage for the cell such as that demonstrated by FLT3-ITD and NPM1 mutations in AML, rapidly establishing a clonal dominance and driving a clear phenotype [104]. In MPN, this mutational compatibility is more subtle but evident from the high frequencies of many mutations including TET2, ASXL1 and DNMT3A occurring alongside drivers observed across the MPN patient spectrum. It is interesting that many of these co-occurring mutations have been observed to be the initiating mutation prior to the acquisition of one of the classical driver mutations or may occur subsequently to it [108, 109]. This bi-directional co-occurrence suggests more than a simple random coincidence in the acquisition of these mutations.
Mutational order
Acquisition order also appears to be a critical determinant of the resulting disease phenotype. Order of acquisition can be inferred from colony assays or single-cell studies allowing genotyping of the clonal structure. In the case of TET2, it is evident that HSCs with this mutation are transcriptionally altered, driving expansion of the clone within the HSC compartment but with limited excess production of terminally differentiated megakaryocytes or erythrocytes until a second hit with the JAK2 V617F mutation occurs. In contrast, JAK2 V617F first cells behave in precisely the opposite manner [108]. Consistent with these findings, loss of TET2 has been observed to promote clonal expansion and self-renewal of HSCs in murine models [110, 111]. TET2-first individuals tend towards an ET phenotype with higher numbers of single mutant cells in the HSC compartment. In contrast, TET2-second individuals show a predominance of double mutant cells in the HSC compartment with a tendency to PV [108]. Figure 4 demonstrates this pictorially. Similarly, loss of function DNMT3A mutations appears to enhance the self-renewal of HSCs and DNMT3A null HSCs are serially transplantable to a significantly enhanced degree in comparison to normal HSCs [112, 113]. Loss of function of another epigenetic regulator EZH2 has also been observed to enhance self-renewal of the JAK2 V617F positive HSC in a murine model [114]. JAK2 V617F only clones in contrast do not promote an MPN disease phenotype on serial transplantation [111]. Early observations that JAK2 V617F positive individuals could transform to JAK2 V671F negative blast phase disease appear to highlight the ability of pre-JAK2 clone with self-renewing capacity in the stem compartment to later establish dominance over the JAK2 clone [115]. Recent advances have moved away from a traditional model of haematopoiesis in which precursors progress through a series of discrete intermediate stages with decreasing differential potential at each stage to a continuum where the boundaries between stem and progenitor cells are increasingly blurred [116]. The epigenetic regulation of transcriptional control affected by loss of normal TET2 and DNMT3A may allow HSC to access alternative transcriptional programmes, shift along this continuum and promote self-renewal. It is interesting that in CHIP individuals, TET2 and DNMT3A clonal fractions are approximately 25% and 14% smaller than ASXL1 clonal fractions suggesting a differential ability to promote clonal expansion [117]. In any instance, it is clear that the field in which the JAK2 mutation is sown is very different in the context of an earlier mutation. Whether these TET2 or DNMT3A mutant first cells are primed for JAK2 mutagenesis or the subsequent frequency of JAK2 V617F mutations simply reflects random chance and high turnover is not clear at this stage.

Effects of mutation order in MPN. Order of mutation acquisition affects both self-renewal capacity of mutants in the haematopoietic stem cell compartment and proliferative drive ultimately affecting the probability of producing a PV or ET phenotype
Cell of origin
Further cellular context is provided by the cell of origin and ultimate differentiation bias of this cell. Detection of driver mutations in multiple different mature cell compartments including erythroid, megakaryocyte, granulocyte, monocyte and in some cases lymphoid cells provide evidence of an initiating cell with multipotency implicating the HSC population [118]. In keeping with this, JAK2 and CALR mutants are detectable within CD34+, CD38- HSC and CD34+, CD38+ progenitor populations [119]. A murine model incorporating a humanised ossicle niche has demonstrated engraftment of myelofibrosis from the CD34+, CD38− HSC population only, with no engraftment in the CD34+, CD38+ progenitor population, suggesting that the initiating cell resides within this HSC population [120]. A knock-in JAK2 V617F model has also demonstrated the critical role of the HSC population in initiation and maintenance of the polycythaemia phenotype [121]. Therefore, involvement of the multi-potent HSC population appears to be important.
Yet, multipotency of these cells in experimental conditions may not reflect the true balances and equilibriums at play in a complex multi-factorial environment such as the bone marrow. In fact, some of these cells appear to have an in-built differentiation bias from a very early position in the haematopoietic hierarchy. In mouse models, platelet biased HSC populations have been identified at the apex of the hierarchy, with enhanced capability of short- and stable long-term platelet production without loss of self-renewal ability and retain the ability in a proportionally limited manner give rise to lymphoid biased HSCs [122]. It is not clear if HSC exist with other lineage bias [123]. Application of single-cell technologies has demonstrated this megakaryocyte differentiation bias in HSCs in myelofibrosis patients with the majority of megakaryocyte progenitors transcriptionally distinct from normal, with proliferative and fibrosis enhancing gene signatures evident [124]. Similarly, single-cell studies in JAK2 V617F positive ET patients have demonstrated an expanded population of megakaryocyte primed HSCs with increased sensitivity to interferon alpha signalling [125]. These findings point towards an inherent bias in phenotype, at least towards ET or PMF, depending on the bias on the initiating stem cell acquiring the first mutation.
The interaction between the malignant clone and stromal cells by means of pro-inflammatory and cytokine signalling is a key determinant in the formation of a fibrotic phenotype. Myofibroblasts responsible for the deposition of collagen are derived from multipotent mesenchymal progenitor stromal cells [126,127,128]. These myofibroblasts evolve over time, undergoing a maldifferentiation process to lose ability to support haematopoietic tissue and contribute to marrow fibrosis. The presence of the malignant haematopoietic clone and resulting inflammatory milieu provide a continuous drive for myofibroblast differentiation and vicious cycle of fibrosis [128]. Removing Gli1+ mesenchymal stromal cells or interfering with PDGFRA signalling in these cells can ameliorate the fibrotic phenotype [126, 127].
Predisposing factors
A number of risk factors for the development of MPN have been identified. Individuals with CHIP have an enhanced risk of myeloid malignancy [19]. Although these patients are distinguished by the absence of a myeloid neoplasm phenotype, large studies have demonstrated subtle alterations in the full blood counts of these individuals with an increased red cell distribution width and modest increase in white cells and decrease in haemoglobin [117]. More than 75% of mutations in CHIP are accounted for by ASXL1, TET2 and DNMT3A, all evident in greater than 5% of MPN cases, whilst the next five most commonly affected genes JAK2, PPM1D, SRSF2, SF3B1 and TP53 are all identified at frequencies of around 2% in MPN with the exception of JAK2, an MPN driver very commonly identified as discussed [15, 117]. This highlights the similarity between these two groups. Estimates of the prevalence of individuals with detectable JAK2 V617F mutations falling into the CHIP category suggest significantly more than demonstrate an MPN phenotype [129]. Therefore, the risk of transformation to MPN even with the presence of a driver mutation is not absolute. However, as noted, similarly to the overt MPN group, these JAK2 V617F positive CHIP individuals have a significantly increased risk of cardiovascular disease [20]. Evidence to base accurate discrimination between patients with clonal haematopoiesis and a low or high risk of transformation to MPN on the basis of genetic or clinical risk factors is less clear than for those with a high risk of leukaemic transformation, and further investigation is required to understand those at risk.
Familial inheritance and germline predisposition to myeloid malignancy has become an important focus of research in the era of widespread genomic analysis [130]. In fact, the heritable risk of MPN is much more significant than evident in many other cancers and higher than for other myeloid malignancy with a ratio of almost 5 observed to expected cases of MPN in individuals with affected first degree relatives [131]. Another study has suggested prevalence of familial cases at approximately 8% [132]. The commonly observed somatic driver mutations in JAK2 and CALR and MPL W515L are not inherited. Examples of families with a high penetrance germ line mutation driving thrombocytosis including MPL S505N or MPL P106L are very rare, but these should be considered in cases with a strong familial components or paediatric patients [133, 134]. Instead, the familial risk appears to result from the presence of predisposing germline susceptibility factors. The JAK2 46/1 haplotype has been consistently observed to confer a higher risk of acquiring the JAK2 V617F. This is true both for JAK2 V617F positive MPN or CHIP [117, 135]. Similarly, polymorphisms in telomerase reverse transcriptase gene (TERT) have been repeatedly identified as independent risk factors for the development of MPN. TERT functions to ensure telomere stability. Several TERT polymorphisms have been identified as predisposing factors for MPN and are also associated with the development of other solid tumours [136, 137]. In contrast to dyskeratosis congenita, the TERT polymorphisms are associated with telomere lengthening. Increased telomere length has been associated with MPN risk [136]. This extensive genome-wide association study identified a further 15 gene loci in addition to these TERT loci increasing risk of MPN. This included loci within TET2, JAK2, GATA2, ATM, RUNX1 and CHEK2 [136].
Outside of genetic risk factors, the myeloproliferative neoplasms: an in-depth case–control (MOSAICC) study identified a number of potentially modifiable risk factors in the development of MPN. This included childhood household density, low childhood socioeconomic status and a high “pack year” smoking history. Obesity was linked with ET specifically. Alcohol intake was inversely associated with MPN risk [138]. These links may point to a role for inflammation or environmental stressors in modifying the epigenetic risk profile for MPN development. Finally, incidence of PV has traditionally been reported higher in males, whilst the incidence of ET is higher in females. Sex has been observed to be an independent variable for JAK2 V617F allele burden with significantly lower allele burdens reported in women compared to men [139].
Epigenetic dysregulation
Dysregulation of normal epigenetic mechanisms of transcription and translation is increasingly evident in MPN. Activation of the JAK/STAT pathways in response to ligand binding has rapid effect on the chromatin architecture and transcription factor binding profiles [32]. Cytokine-induced changes are different from those induced by chronic constitutive activation of STAT proteins [140]. There is evidence that the histone landscape in MPN is abnormal. These abnormalities can occur even in the absence of any epigenetic modifier mutations. MPL W515L mice demonstrate a differential landscape of H3K27 acetylation [85]. Global levels of H3K9 mono and di-methylation are significantly reduced, mediated by enhanced JMJD1C expression as a result of NFE2 overexpression [141]. The mutant JAK2 V617 protein appears able to directly influence the chromatin landscape in a differential manner and independent of STAT protein interactions. JAK2 V617F can interact and phosphorylate protein arginine methyltransferase 5 (PRMT5), impairing PRMT5 histone methylation. This appears to enhance myeloproliferation [142]. JAK2 can locate to the nucleus and mediate phosphorylation of tyrosine 41 on histone H3 (H3Y41). Enhanced levels of H3Y41 were most abundant in JAK2 V617F containing cell lines. This phosphorylation was reduced by JAK inhibitors and was directly implicated in displacing normal heterochromatin protein 1α [143]. We have recently observed a significant effect of ruxolitinib in the modification of the histone landscape in MPN cell line models and patient samples [84]. In addition, the heterogeneity of histone landscapes will be enhanced by the presence or absence of loss of function mutations in epigenetic modifiers like EZH2 and ASXL1. In JAK2 V617F mice with conditional EZH2 deletion, there is a significant downregulation of the transcriptional repressor H3K27me3 mediated by loss of normal polycomb repressive complex 2 function and upregulation of H3K27 acetylation resulting in the activation of genes associated with PMF pathogenesis [144].
DNA methylation has been observed to be abnormal in MPN. There are also differences between disease phenotypes and during progression to blast phase [145, 146]. Studies of particular genes including SOCS3 and CD18 have suggested differential methylation status in some MPN patients [81, 147]. Commonly mutated regulators of DNA methylation include TET2, DNMT3A and IDH1/2.
As previously described, components of the spliceosome including SF3B1, SRSF2 and USAF1 are observed to be mutated in small numbers of MPN patients, particularly those with myelofibrosis. SF3B1 and JAK2 mutations are commonly observed together in patients with the distinct overlap syndrome and clinical entity MDS/MPN with ringed sideroblasts and thrombocytosis [148]. Mutant JAK2 V617F has been observed to directly phosphorylate a number of components of the splicing machinery differentially from wild type JAK2 in mouse haematopoietic cells. This results in an alteration in JAK2-ERK signalling to maintain the JAK2 V617F clones. JAK2 mutant cells are sensitised to the JAK inhibitor ruxolitinib after inactivation of YBX1, a splicing enzyme [59].
Targeted therapy
As discussed throughout this review, JAK inhibitors, and in particular ruxolitinib, have become a key therapeutic agent in the MPN clinic. They are clearly beneficial in a range of specific MPN patient scenarios with good evidence for spleen volume reduction, haematocrit control and symptom control and some evidence to support a survival benefit in myelofibrosis and with combined hypomethylating agents in blast phase disease [25,26,27, 149]. Initial hopes of a disease modifying effect similar to tyrosine kinase therapy in CML have not materialised with limited reduction in mutant VAF, limited change in marrow fibrosis and often a limited duration of efficacy prior to loss of response. Exploring the nature of this developed resistance to JAK inhibition in detail is a complex and extensive topic beyond the scope of this article. It is interesting to note that further acquired mutations in JAK2 do not appear to be a significant contributor to resistance in patients. There is evidence to support alternative heterodimer formation between JAK2 and JAK1 or TYK1 in MPN cell line models persistently exposed to ruxolitinib and evidence to support recruitment of alternative MAPK signalling bypassing the JAK/STAT pathway as mechanistic explanations of this resistance [150]. The complexity of the genetic changes within the MPN clone may also determine the responsiveness of the cell to ruxolitinib, whilst new somatic mutations driving clonal evolution on therapy and subsequent expansion of new clones has been clearly documented [151, 152].
There is presently significant interest in targeted combination therapies to augment the beneficial effects of JAK inhibition, particularly in myelofibrosis and blast phase disease. Given the complex features of disease pathogenesis, it is not surprising that a range of therapies targeting intracellular signalling cascades, cytokines and epigenetic regulators have been shown to demonstrate some efficacious features in the management of MPN.
Interferon alpha therapy is increasing recognised as a potential disease modifying agent in MPN and therefore should be classed within the targeted therapeutic approaches. There is clear evidence of reductions in the clonal size and longer term “remissions” of disease are possible [153]. Recent single-cell work demonstrated an enhanced sensitivity to interferon treatment in JAK2 V617F positive HSCs from ET patients. Treatment appeared to result in the apoptosis of heterozygous cells whilst establishing quiescence in the homozygous cells [125]. In a murine JAK2 V617F positive model, interferon alpha treatment promoted a shift towards CD41hi expressing HSC population with a megakaryocyte bias and active cell cycling in the JAK2 V617F positive HSCs, ultimately exhausting the mutant clone [154].
The options for novel therapies in MPN have been recently extensively summarised [155]. Table 2 summarises classes of targeted drugs currently under evaluation in MPN divided into categories based on the sections of this review article.
Conclusion
The study of disease provides more than simply a means to alleviating suffering. The insights gained from understanding the molecular pathogenesis of disorders like MPN provide an insight into the phenomenal complexity and simultaneous simplicity with which our cells function. Perhaps most intriguing is the apparent simplicity with which the balance of myeloproliferation is upset and, the relative ease that single mutations in the right cellular context can generate neoplastic clones with an enhanced proliferative drive.
And yet, from the initial discovery of the JAK2 V617F mutation as a key driver in the majority of MPN patients, it is increasing clear that many more subtleties determine the overall disease phenotype, prognosis and whether “disease” develops at all. The presence of additional pathogenic mutations, the order of acquisition, cellular context, germline predisposition factors, the balance of STAT protein signalling alongside PI3K and MAPK signalling, epigenetic dysregulation and extrinsic influences may all affect the ultimate clonal structure, proliferative drive and differentiation capacity of the neoplastic cells. Genetic complexity and heterogeneity across the population and within single individuals provides a significant diagnostic and therapeutic challenge in MPN. As the medical community transitions into an era in which each stage of work-up and treatment of an individual patient can generate large volumes of information on a scale beyond the analytic capacity of single individuals, we will move further away from discrete classification and categorisation of disease towards individualised clinical, genomic and pathological characterisation. The challenge is how we successfully unleash this potential to understand individualised molecular pathogenesis with translation into effective individualised treatments.
Availability of data and materials
Not applicable.
Abbreviations
- MPN:
-
Myeloproliferative neoplasm
- HSC:
-
Haematopoietic stem cell
- PV:
-
Polycythaemia vera
- ET:
-
Essential thrombocythaemia
- PMF:
-
Primary myelofibrosis
- Pre-PMF:
-
Pre-fibrotic primary myelofibrosis
- MF:
-
Myelofibrosis
- JAK:
-
Janus-associated kinase
- STAT:
-
Signal transducer and activator of transcription
- WHO:
-
World Health Organisation
- CHIP:
-
Clonal haematopoiesis of indeterminate potential
- EPO:
-
Erythropoietin
- TPO:
-
Thrombopoietin
- GCSF:
-
Granulocyte colony stimulating factor
- EPOR:
-
Erythropoietin receptor
- MPL:
-
Thrombopoietin receptor
- GCSFR:
-
Granulocyte colony stimulating factor receptor
- CALR:
-
Calreticulin
- VAF:
-
Variant allele frequency
- SOCS:
-
Suppressor of cytokine signalling
- EZH2:
-
Enhancer of Zeste 2
- ERK1/2:
-
Extracellular signal related kinase ½
- PI3K:
-
Phosphoinositide 3-kinase
- MAPK:
-
Mitogen-activated protein kinase
- PDGFRα/PDGRFA:
-
Platelet-derived growth factor receptor alpha
- mTOR:
-
Mechanistic target of rapamycin
- AML:
-
Acute myeloid leukaemia
- CML:
-
Chronic myeloid leukaemia
- CBL:
-
Cbl proto-oncogene
- PTEN:
-
Phosphatase and tension homolog deleted on chromosome 10
- DUSP1:
-
Dual-specificity phosphatase 1
- NGS:
-
Next-generation sequencing
- TERT:
-
Telomerase reverse transcriptase
- PRMT5:
-
Protein arginine methyltransferase 5
- LSD1:
-
Lysine demethylase 1A
References
Arber DA, Orazi A, Hasserjian R, Thiele J, Borowitz MJ, Le Beau MM, et al. The 2016 revision to the World Health Organization classification of myeloid neoplasms and acute leukemia. Blood. 2016;127(20):2391–405.
Rampal R, Al-Shahrour F, Abdel-Wahab O, Patel JP, Brunel JP, Mermel CH, et al. Integrated genomic analysis illustrates the central role of JAK-STAT pathway activation in myeloproliferative neoplasm pathogenesis. Blood. 2014;123(22):e123–33.
Levine RL, Wadleigh M, Cools J, Ebert BL, Wernig G, Huntly BJ, et al. Activating mutation in the tyrosine kinase JAK2 in polycythemia vera, essential thrombocythemia, and myeloid metaplasia with myelofibrosis. Cancer Cell. 2005;7(4):387–97.
Scott LM, Tong W, Levine RL, Scott MA, Beer PA, Stratton MR, et al. JAK2 exon 12 mutations in polycythemia vera and idiopathic erythrocytosis. N Engl J Med. 2007;356(5):459–68.
Baxter EJ, Scott LM, Campbell PJ, East C, Fourouclas N, Swanton S, et al. Acquired mutation of the tyrosine kinase JAK2 in human myeloproliferative disorders. Lancet. 2005;365(9464):1054–61.
Kralovics R, Passamonti F, Buser AS, Teo SS, Tiedt R, Passweg JR, et al. A gain-of-function mutation of JAK2 in myeloproliferative disorders. N Engl J Med. 2005;352(17):1779–90.
Nangalia J, Massie CE, Baxter EJ, Nice FL, Gundem G, Wedge DC, et al. Somatic CALR mutations in myeloproliferative neoplasms with nonmutated JAK2. N Engl J Med. 2013;369(25):2391–405.
Pikman Y, Lee BH, Mercher T, McDowell E, Ebert BL, Gozo M, et al. MPLW515L is a novel somatic activating mutation in myelofibrosis with myeloid metaplasia. PLoS Med. 2006;3(7):e270.
Nangalia J, Green AR. Myeloproliferative neoplasms: from origins to outcomes. Blood. 2017;130(23):2475–83.
Barbui T, Thiele J, Gisslinger H, Kvasnicka HM, Vannucchi AM, Guglielmelli P, et al. The 2016 WHO classification and diagnostic criteria for myeloproliferative neoplasms: document summary and in-depth discussion. Blood Cancer J. 2018;8(2):15.
Mascarenhas J, Heaney ML, Najfeld V, Hexner E, Abdel-Wahab O, Rampal R, et al. Proposed criteria for response assessment in patients treated in clinical trials for myeloproliferative neoplasms in blast phase (MPN-BP): formal recommendations from the post-myeloproliferative neoplasm acute myeloid leukemia consortium. Leuk Res. 2012;36(12):1500–4.
McMullin MF, Harrison CN, Ali S, Cargo C, Chen F, Ewing J, et al. A guideline for the diagnosis and management of polycythaemia vera. A British Society for Haematology Guideline. Br J Haematol. 2019;184(2):176–91.
Tefferi A, Lasho TL, Finke CM, Elala Y, Hanson CA, Ketterling RP, et al. Targeted deep sequencing in primary myelofibrosis. Blood Adv. 2016;1(2):105–11.
Tefferi A, Lasho TL, Guglielmelli P, Finke CM, Rotunno G, Elala Y, et al. Targeted deep sequencing in polycythemia vera and essential thrombocythemia. Blood Adv. 2016;1(1):21–30.
Grinfeld J, Nangalia J, Baxter EJ, Wedge DC, Angelopoulos N, Cantrill R, et al. Classification and personalized prognosis in myeloproliferative neoplasms. N Engl J Med. 2018;379(15):1416–30.
Rodriguez-Meira A, Buck G, Clark SA, Povinelli BJ, Alcolea V, Louka E, et al. Unravelling intratumoral heterogeneity through high-sensitivity single-cell mutational analysis and parallel RNA sequencing. Mol Cell. 2019;73(6):1292-305.e8.
Papaemmanuil E, Gerstung M, Bullinger L, Gaidzik VI, Paschka P, Roberts ND, et al. Genomic classification and prognosis in acute myeloid leukemia. N Engl J Med. 2016;374(23):2209–21.
Zoi K, Cross NC. Molecular pathogenesis of atypical CML, CMML and MDS/MPN-unclassifiable. Int J Hematol. 2015;101(3):229–42.
Jaiswal S, Fontanillas P, Flannick J, Manning A, Grauman PV, Mar BG, et al. Age-related clonal hematopoiesis associated with adverse outcomes. N Engl J Med. 2014;371(26):2488–98.
Jaiswal S, Natarajan P, Silver AJ, Gibson CJ, Bick AG, Shvartz E, et al. Clonal hematopoiesis and risk of atherosclerotic cardiovascular disease. N Engl J Med. 2017;377(2):111–21.
Jaiswal S, Libby P. Clonal haematopoiesis: connecting ageing and inflammation in cardiovascular disease. Nat Rev Cardiol. 2020;17(3):137–44.
Landolfi R, Marchioli R, Kutti J, Gisslinger H, Tognoni G, Patrono C, et al. Efficacy and safety of low-dose aspirin in polycythemia vera. N Engl J Med. 2004;350(2):114–24.
Marchioli R, Finazzi G, Specchia G, Masciulli A, Mennitto MR, Barbui T. The CYTO-PV: a large-scale trial testing the intensity of CYTOreductive therapy to prevent cardiovascular events in patients with polycythemia vera. Thrombosis. 2011;2011:794240.
Gisslinger H, Klade C, Georgiev P, Krochmalczyk D, Gercheva-Kyuchukova L, Egyed M, et al. Ropeginterferon alfa-2b versus standard therapy for polycythaemia vera (PROUD-PV and CONTINUATION-PV): a randomised, non-inferiority, phase 3 trial and its extension study. Lancet Haematol. 2020;7(3):e196–208.
Harrison CN, Vannucchi AM, Kiladjian JJ, Al-Ali HK, Gisslinger H, Knoops L, et al. Long-term findings from COMFORT-II, a phase 3 study of ruxolitinib vs best available therapy for myelofibrosis. Leukemia. 2016;30(8):1701–7.
Griesshammer M, Saydam G, Palandri F, Benevolo G, Egyed M, Callum J, et al. Ruxolitinib for the treatment of inadequately controlled polycythemia vera without splenomegaly: 80-week follow-up from the RESPONSE-2 trial. Ann Hematol. 2018;97:1591–600.
Verstovsek S, Mesa RA, Gotlib J, Levy RS, Gupta V, DiPersio JF, et al. A double-blind, placebo-controlled trial of ruxolitinib for myelofibrosis. N Engl J Med. 2012;366(9):799–807.
Vannucchi AM, Kiladjian JJ, Griesshammer M, Masszi T, Durrant S, Passamonti F, et al. Ruxolitinib versus standard therapy for the treatment of polycythemia vera. N Engl J Med. 2015;372(5):426–35.
Jain T, Mesa RA, Palmer JM. Allogeneic stem cell transplantation in myelofibrosis. Biol Blood Marrow Transplant. 2017;23(9):1429–36.
Gupta V, Kennedy JA, Capo-Chichi JM, Kim S, Hu ZH, Alyea EP, et al. Genetic factors rather than blast reduction determine outcomes of allogeneic HCT in BCR-ABL-negative MPN in blast phase. Blood Adv. 2020;4(21):5562–73.
Bharadwaj U, Kasembeli MM, Robinson P, Tweardy DJ. Targeting Janus kinases and signal transducer and activator of transcription 3 to treat inflammation, fibrosis, and cancer: rationale, progress, and caution. Pharmacol Rev. 2020;72(2):486–526.
Wingelhofer B, Neubauer HA, Valent P, Han X, Constantinescu SN, Gunning PT, et al. Implications of STAT3 and STAT5 signaling on gene regulation and chromatin remodeling in hematopoietic cancer. Leukemia. 2018;32(8):1713–26.
Tiacci E, Ladewig E, Schiavoni G, Penson A, Fortini E, Pettirossi V, et al. Pervasive mutations of JAK-STAT pathway genes in classical Hodgkin lymphoma. Blood. 2018;131(22):2454–65.
Liu Y, Easton J, Shao Y, Maciaszek J, Wang Z, Wilkinson MR, et al. The genomic landscape of pediatric and young adult T-lineage acute lymphoblastic leukemia. Nat Genet. 2017;49(8):1211–8.
Venugopal S, Bar-Natan M, Mascarenhas JO. JAKs to STATs: a tantalizing therapeutic target in acute myeloid leukemia. Blood Rev. 2020;40:100634.
Yu H, Jove R. The STATs of cancer—new molecular targets come of age. Nat Rev Cancer. 2004;4(2):97–105.
James C, Ugo V, Le Couédic JP, Staerk J, Delhommeau F, Lacout C, et al. A unique clonal JAK2 mutation leading to constitutive signalling causes polycythaemia vera. Nature. 2005;434(7037):1144–8.
Mejia-Ochoa M, Acevedo Toro PA, Cardona-Arias JA. Systematization of analytical studies of polycythemia vera, essential thrombocythemia and primary myelofibrosis, and a meta-analysis of the frequency of JAK2, CALR and MPL mutations: 2000–2018. BMC Cancer. 2019;19(1):590.
Ding J, Komatsu H, Iida S, Yano H, Kusumoto S, Inagaki A, et al. The Asn505 mutation of the c-MPL gene, which causes familial essential thrombocythemia, induces autonomous homodimerization of the c-Mpl protein due to strong amino acid polarity. Blood. 2009;114(15):3325–8.
Defour JP, Chachoua I, Pecquet C, Constantinescu SN. Oncogenic activation of MPL/thrombopoietin receptor by 17 mutations at W515: implications for myeloproliferative neoplasms. Leukemia. 2016;30(5):1214–6.
Bridgford JL, Lee SM, Lee CMM, Guglielmelli P, Rumi E, Pietra D, et al. Novel drivers and modifiers of MPL-dependent oncogenic transformation identified by deep mutational scanning. Blood. 2020;135(4):287–92.
How J, Hobbs GS, Mullally A. Mutant calreticulin in myeloproliferative neoplasms. Blood. 2019;134(25):2242–8.
Benlabiod C, Cacemiro MDC, Nedelec A, Edmond V, Muller D, Rameau P, et al. Calreticulin del52 and ins5 knock-in mice recapitulate different myeloproliferative phenotypes observed in patients with MPN. Nat Commun. 2020;11(1):4886.
Michail O, McCallion P, McGimpsey J, Hindley A, Greenfield G, Feerick J, et al. Mutational profiling in suspected triple-negative essential thrombocythaemia using targeted next-generation sequencing in a real-world cohort. J Clin Pathol. 2020;jclinpath-2020-206570.
Mullally A, Lane SW, Brumme K, Ebert BL. Myeloproliferative neoplasm animal models. Hematol Oncol Clin N Am. 2012;26(5):1065–81.
Verstovsek S, Mesa RA, Gotlib J, Levy RS, Gupta V, DiPersio JF, et al. The clinical benefit of ruxolitinib across patient subgroups: analysis of a placebo-controlled, Phase III study in patients with myelofibrosis. Br J Haematol. 2013;161(4):508–16.
Antonioli E, Guglielmelli P, Poli G, Bogani C, Pancrazzi A, Longo G, et al. Influence of JAK2V617F allele burden on phenotype in essential thrombocythemia. Haematologica. 2008;93(1):41–8.
Godfrey AL, Chen E, Pagano F, Silber Y, Campbell PJ, Green AR. Clonal analyses reveal associations of JAK2V617F homozygosity with hematologic features, age and gender in polycythemia vera and essential thrombocythemia. Haematologica. 2013;98(5):718–21.
Passamonti F, Rumi E, Pietra D, Elena C, Boveri E, Arcaini L, et al. A prospective study of 338 patients with polycythemia vera: the impact of JAK2 (V617F) allele burden and leukocytosis on fibrotic or leukemic disease transformation and vascular complications. Leukemia. 2010;24(9):1574–9.
How J, Trinkaus KM, Oh ST. Distinct clinical, laboratory and molecular features of myeloproliferative neoplasm patients with splanchnic vein thrombosis. Br J Haematol. 2018;183(2):310–3.
Rumi E, Pietra D, Ferretti V, Klampfl T, Harutyunyan AS, Milosevic JD, et al. JAK2 or CALR mutation status defines subtypes of essential thrombocythemia with substantially different clinical course and outcomes. Blood. 2014;123(10):1544–51.
De Grandis M, Cambot M, Wautier MP, Cassinat B, Chomienne C, Colin Y, et al. JAK2V617F activates Lu/BCAM-mediated red cell adhesion in polycythemia vera through an EpoR-independent Rap1/Akt pathway. Blood. 2013;121(4):658–65.
Wolach O, Sellar RS, Martinod K, Cherpokova D, McConkey M, Chappell RJ, et al. Increased neutrophil extracellular trap formation promotes thrombosis in myeloproliferative neoplasms. Sci Transl Med. 2018;10(436):eaan8292.
Nienhold R, Ashcroft P, Zmajkovic J, Rai S, Rao TN, Drexler B, et al. MPN patients with low mutant JAK2 allele burden show late expansion restricted to erythroid and megakaryocytic lineages. Blood. 2020;136(22):2591–5.
Zhang HF, Lai R. STAT3 in cancer-friend or foe? Cancers (Basel). 2014;6(3):1408–40.
Funakoshi-Tago M, Tago K, Abe M, Sonoda Y, Kasahara T. STAT5 activation is critical for the transformation mediated by myeloproliferative disorder-associated JAK2 V617F mutant. J Biol Chem. 2010;285(8):5296–307.
Garcon L, Rivat C, James C, Lacout C, Camara-Clayette V, Ugo V, et al. Constitutive activation of STAT5 and Bcl-xL overexpression can induce endogenous erythroid colony formation in human primary cells. Blood. 2006;108(5):1551–4.
Chen E, Beer PA, Godfrey AL, Ortmann CA, Li J, Costa-Pereira AP, et al. Distinct clinical phenotypes associated with JAK2V617F reflect differential STAT1 signaling. Cancer Cell. 2010;18(5):524–35.
Jayavelu AK, Schnoder TM, Perner F, Herzog C, Meiler A, Krishnamoorthy G, et al. Splicing factor YBX1 mediates persistence of JAK2-mutated neoplasms. Nature. 2020;588(7836):157–63.
Walz C, Ahmed W, Lazarides K, Betancur M, Patel N, Hennighausen L, et al. Essential role for Stat5a/b in myeloproliferative neoplasms induced by BCR-ABL1 and JAK2(V617F) in mice. Blood. 2012;119(15):3550–60.
Yan D, Hutchison RE, Mohi G. Critical requirement for Stat5 in a mouse model of polycythemia vera. Blood. 2012;119(15):3539–49.
Duek A, Lundberg P, Shimizu T, Grisouard J, Karow A, Kubovcakova L, et al. Loss of Stat1 decreases megakaryopoiesis and favors erythropoiesis in a JAK2-V617F-driven mouse model of MPNs. Blood. 2014;123(25):3943–50.
Nitulescu II, Meyer SC, Wen QJ, Crispino JD, Lemieux ME, Levine RL, et al. Mediator kinase phosphorylation of STAT1 S727 promotes growth of neoplasms with JAK-STAT activation. EBioMedicine. 2017;26:112–25.
Godfrey AL, Chen E, Massie CE, Silber Y, Pagano F, Bellosillo B, et al. STAT1 activation in association with JAK2 exon 12 mutations. Haematologica. 2016;101(1):e15–9.
Roder S, Steimle C, Meinhardt G, Pahl HL. STAT3 is constitutively active in some patients with Polycythemia rubra vera. Exp Hematol. 2001;29(6):694–702.
Risum M, Madelung A, Bondo H, Bzorek M, Kristensen MH, Stamp IM, et al. The JAK2V617F allele burden and STAT3- and STAT5 phosphorylation in myeloproliferative neoplasms: early prefibrotic myelofibrosis compared with essential thrombocythemia, polycythemia vera and myelofibrosis. APMIS. 2011;119(8):498–504.
Prestipino A, Emhardt AJ, Aumann K, O’Sullivan D, Gorantla SP, Duquesne S, et al. Oncogenic JAK2(V617F) causes PD-L1 expression, mediating immune escape in myeloproliferative neoplasms. Sci Transl Med. 2018;10(429):eaam7729.
Balic JJ, White CL, Dawson R, Gough D, McCormack MP, Jenkins BJ. STAT3-driven hematopoiesis and lymphopoiesis abnormalities are dependent on serine phosphorylation. Cytokine. 2020;130:155059.
Grisouard J, Shimizu T, Duek A, Kubovcakova L, Hao-Shen H, Dirnhofer S, et al. Deletion of Stat3 in hematopoietic cells enhances thrombocytosis and shortens survival in a JAK2-V617F mouse model of MPN. Blood. 2015;125(13):2131–40.
Comoglio F, Park HJ, Schoenfelder S, Barozzi I, Bode D, Fraser P, et al. Thrombopoietin signaling to chromatin elicits rapid and pervasive epigenome remodeling within poised chromatin architectures. Genome Res. 2018;7:227.
Dasgupta M, Dermawan JK, Willard B, Stark GR. STAT3-driven transcription depends upon the dimethylation of K49 by EZH2. Proc Natl Acad Sci USA. 2015;112(13):3985–90.
Kollmann K, Warsch W, Gonzalez-Arias C, Nice FL, Avezov E, Milburn J, et al. A novel signalling screen demonstrates that CALR mutations activate essential MAPK signalling and facilitate megakaryocyte differentiation. Leukemia. 2017;31(4):934–44.
Zini R, Guglielmelli P, Pietra D, Rumi E, Rossi C, Rontauroli S, et al. CALR mutational status identifies different disease subtypes of essential thrombocythemia showing distinct expression profiles. Blood Cancer J. 2017;7(12):638.
Stivala S, Codilupi T, Brkic S, Baerenwaldt A, Ghosh N, Hao-Shen H, et al. Targeting compensatory MEK/ERK activation increases JAK inhibitor efficacy in myeloproliferative neoplasms. J Clin Invest. 2019;129(4):1596–611.
Bartalucci N, Calabresi L, Balliu M, Martinelli S, Rossi MC, Villeval JL, et al. Inhibitors of the PI3K/mTOR pathway prevent STAT5 phosphorylation in. Oncotarget. 2017;8(57):96710–24.
Guglielmelli P, Barosi G, Rambaldi A, Marchioli R, Masciulli A, Tozzi L, et al. Safety and efficacy of everolimus, a mTOR inhibitor, as single agent in a phase 1/2 study in patients with myelofibrosis. Blood. 2011;118(8):2069–76.
Rocca S, Carra G, Poggio P, Morotti A, Brancaccio M. Targeting few to help hundreds: JAK, MAPK and ROCK pathways as druggable targets in atypical chronic myeloid leukemia. Mol Cancer. 2018;17(1):40.
Hookham MB, Elliott J, Suessmuth Y, Staerk J, Ward AC, Vainchenker W, et al. The myeloproliferative disorder-associated JAK2 V617F mutant escapes negative regulation by suppressor of cytokine signaling 3. Blood. 2007;109(11):4924–9.
Varghese LN, Ungureanu D, Liau NP, Young SN, Laktyushin A, Hammaren H, et al. Mechanistic insights into activation and SOCS3-mediated inhibition of myeloproliferative neoplasm-associated JAK2 mutants from biochemical and structural analyses. Biochem J. 2014;458(2):395–405.
Funakoshi-Tago M, Tsuruya R, Ueda F, Ishihara A, Kasahara T, Tamura H, et al. Tyrosine-phosphorylated SOCS3 negatively regulates cellular transformation mediated by the myeloproliferative neoplasm-associated JAK2 V617F mutant. Cytokine. 2019;123:154753.
Fourouclas N, Li J, Gilby DC, Campbell PJ, Beer PA, Boyd EM, et al. Methylation of the suppressor of cytokine signaling 3 gene (SOCS3) in myeloproliferative disorders. Haematologica. 2008;93(11):1635–44.
Stover DG, Gil Del Alcazar CR, Brock J, Guo H, Overmoyer B, Balko J, et al. Phase II study of ruxolitinib, a selective JAK1/2 inhibitor, in patients with metastatic triple-negative breast cancer. NPJ Breast Cancer. 2018;4:10.
Yang X, Jia J, Yu Z, Duanmu Z, He H, Chen S, et al. Inhibition of JAK2/STAT3/SOCS3 signaling attenuates atherosclerosis in rabbit. BMC Cardiovasc Disord. 2020;20(1):133.
Greenfield G, McPherson S, Smith J, Mead A, Harrison C, Mills K, et al. Modification of the histone landscape with JAK inhibition in myeloproliferative neoplasms. Cancers (Basel). 2020;12(9):2669.
Kleppe M, Koche R, Zou L, van Galen P, Hill CE, Dong L, et al. Dual targeting of oncogenic activation and inflammatory signaling increases therapeutic efficacy in myeloproliferative neoplasms. Cancer Cell. 2018;33(1):29-43.e7.
Chen CQ, Yu K, Yan QX, Xing CY, Chen Y, Yan Z, et al. Pure curcumin increases the expression of SOCS1 and SOCS3 in myeloproliferative neoplasms through suppressing class I histone deacetylases. Carcinogenesis. 2013;34(7):1442–9.
Gao SM, Chen CQ, Wang LY, Hong LL, Wu JB, Dong PH, et al. Histone deacetylases inhibitor sodium butyrate inhibits JAK2/STAT signaling through upregulation of SOCS1 and SOCS3 mediated by HDAC8 inhibition in myeloproliferative neoplasms. Exp Hematol. 2013;41(3):261-70 e4.
Akada H, Akada S, Gajra A, Bair A, Graziano S, Hutchison RE, et al. Efficacy of vorinostat in a murine model of polycythemia vera. Blood. 2012;119(16):3779–89.
Nakata Y, Ueda T, Nagamachi A, Yamasaki N, Ikeda KI, Sera Y, et al. Acquired expression of Cbl(Q367P) in mice induces dysplastic myelopoiesis mimicking chronic myelomonocytic leukemia. Blood. 2017;129(15):2148–60.
Zhu HH, Luo X, Zhang K, Cui J, Zhao H, Ji Z, et al. Shp2 and Pten have antagonistic roles in myeloproliferation but cooperate to promote erythropoiesis in mammals. Proc Natl Acad Sci USA. 2015;112(43):13342–7.
Stetka J, Vyhlidalova P, Lanikova L, Koralkova P, Gursky J, Hlusi A, et al. Addiction to DUSP1 protects JAK2V617F-driven polycythemia vera progenitors against inflammatory stress and DNA damage, allowing chronic proliferation. Oncogene. 2019;38(28):5627–42.
Tyner JW, Tognon CE, Bottomly D, Wilmot B, Kurtz SE, Savage SL, et al. Functional genomic landscape of acute myeloid leukaemia. Nature. 2018;562(7728):526–31.
Zink F, Stacey SN, Norddahl GL, Frigge ML, Magnusson OT, Jonsdottir I, et al. Clonal hematopoiesis, with and without candidate driver mutations, is common in the elderly. Blood. 2017;130(6):742–52.
Haferlach T, Nagata Y, Grossmann V, Okuno Y, Bacher U, Nagae G, et al. Landscape of genetic lesions in 944 patients with myelodysplastic syndromes. Leukemia. 2014;28(2):241–7.
Williams N, Lee J, Moore L, Baxter EJ, Hewinson J, Dawson KJ, et al. Phylogenetic reconstruction of myeloproliferative neoplasm reveals very early origins and lifelong evolution. bioRxiv. 2020:2020.11.09.374710.
Tefferi A. Novel mutations and their functional and clinical relevance in myeloproliferative neoplasms: JAK2, MPL, TET2, ASXL1, CBL, IDH and IKZF1. Leukemia. 2010;24(6):1128–38.
Triviai I, Zeschke S, Rentel J, Spanakis M, Scherer T, Gabdoulline R, et al. ASXL1/EZH2 mutations promote clonal expansion of neoplastic HSC and impair erythropoiesis in PMF. Leukemia. 2018;33:99–109.
Vannucchi AM, Lasho TL, Guglielmelli P, Biamonte F, Pardanani A, Pereira A, et al. Mutations and prognosis in primary myelofibrosis. Leukemia. 2013;27(9):1861–9.
Lundberg P, Karow A, Nienhold R, Looser R, Hao-Shen H, Nissen I, et al. Clonal evolution and clinical correlates of somatic mutations in myeloproliferative neoplasms. Blood. 2014;123(14):2220–8.
Luque Paz D, Riou J, Verger E, Cassinat B, Chauveau A, Ianotto JC, et al. Genomic analysis of primary and secondary myelofibrosis redefines the prognostic impact of ASXL1 mutations: a FIM study. Blood Adv. 2021;5(5):1442–51.
Tefferi A, Guglielmelli P, Nicolosi M, Mannelli F, Mudireddy M, Bartalucci N, et al. GIPSS: genetically inspired prognostic scoring system for primary myelofibrosis. Leukemia. 2018;32(7):1631–42.
Guglielmelli P, Lasho TL, Rotunno G, Score J, Mannarelli C, Pancrazzi A, et al. The number of prognostically detrimental mutations and prognosis in primary myelofibrosis: an international study of 797 patients. Leukemia. 2014;28(9):1804–10.
Tefferi A, Guglielmelli P, Lasho TL, Gangat N, Ketterling RP, Pardanani A, et al. MIPSS70+ version 2.0: mutation and karyotype-enhanced international prognostic scoring system for primary myelofibrosis. J Clin Oncol. 2018;36(17):1769–70.
Miles LA, Bowman RL, Merlinsky TR, Csete IS, Ooi AT, Durruthy-Durruthy R, et al. Single-cell mutation analysis of clonal evolution in myeloid malignancies. Nature. 2020;587(7834):477–82.
Geissler K, Gisslinger B, Jager E, Jager R, Schiefer AI, Bogner E, et al. Myelomonocytic skewing in vitro discriminates subgroups of patients with myelofibrosis with a different phenotype, a different mutational profile and different prognosis. Cancers (Basel). 2020;12(8):2291.
Engle EK, Fisher DA, Miller CA, McLellan MD, Fulton RS, Moore DM, et al. Clonal evolution revealed by whole genome sequencing in a case of primary myelofibrosis transformed to secondary acute myeloid leukemia. Leukemia. 2015;29(4):869–76.
Newberry KJ, Patel K, Masarova L, Luthra R, Manshouri T, Jabbour E, et al. Clonal evolution and outcomes in myelofibrosis after ruxolitinib discontinuation. Blood. 2017;130(9):1125–31.
Ortmann CA, Kent DG, Nangalia J, Silber Y, Wedge DC, Grinfeld J, et al. Effect of mutation order on myeloproliferative neoplasms. N Engl J Med. 2015;372(7):601–12.
Nangalia J, Nice FL, Wedge DC, Godfrey AL, Grinfeld J, Thakker C, et al. DNMT3A mutations occur early or late in patients with myeloproliferative neoplasms and mutation order influences phenotype. Haematologica. 2015;100(11):e438–42.
Moran-Crusio K, Reavie L, Shih A, Abdel-Wahab O, Ndiaye-Lobry D, Lobry C, et al. Tet2 loss leads to increased hematopoietic stem cell self-renewal and myeloid transformation. Cancer Cell. 2011;20(1):11–24.
Kameda T, Shide K, Yamaji T, Kamiunten A, Sekine M, Taniguchi Y, et al. Loss of TET2 has dual roles in murine myeloproliferative neoplasms: disease sustainer and disease accelerator. Blood. 2015;125(2):304–15.
Challen GA, Sun D, Jeong M, Luo M, Jelinek J, Berg JS, et al. Dnmt3a is essential for hematopoietic stem cell differentiation. Nat Genet. 2011;44(1):23–31.
Jeong M, Park HJ, Celik H, Ostrander EL, Reyes JM, Guzman A, et al. Loss of Dnmt3a immortalizes hematopoietic stem cells in vivo. Cell Rep. 2018;23(1):1–10.
Shimizu T, Kubovcakova L, Nienhold R, Zmajkovic J, Meyer SC, Hao-Shen H, et al. Loss of Ezh2 synergizes with JAK2-V617F in initiating myeloproliferative neoplasms and promoting myelofibrosis. J Exp Med. 2016;213(8):1479–96.
Theocharides A, Boissinot M, Girodon F, Garand R, Teo SS, Lippert E, et al. Leukemic blasts in transformed JAK2-V617F-positive myeloproliferative disorders are frequently negative for the JAK2-V617F mutation. Blood. 2007;110(1):375–9.
Laurenti E, Gottgens B. From haematopoietic stem cells to complex differentiation landscapes. Nature. 2018;553(7689):418–26.
Bick AG, Weinstock JS, Nandakumar SK, Fulco CP, Bao EL, Zekavat SM, et al. Inherited causes of clonal haematopoiesis in 97,691 whole genomes. Nature. 2020;586(7831):763–8.
Ishii T, Bruno E, Hoffman R, Xu M. Involvement of various hematopoietic-cell lineages by the JAK2V617F mutation in polycythemia vera. Blood. 2006;108(9):3128–34.
Angona A, Alvarez-Larran A, Bellosillo B, Longaron R, Camacho L, Fernandez-Rodriguez MC, et al. Characterization of CD34+ hematopoietic progenitor cells in JAK2V617F and CALR-mutated myeloproliferative neoplasms. Leuk Res. 2016;48:11–5.
Reinisch A, Thomas D, Corces MR, Zhang X, Gratzinger D, Hong WJ, et al. A humanized bone marrow ossicle xenotransplantation model enables improved engraftment of healthy and leukemic human hematopoietic cells. Nat Med. 2016;22(7):812–21.
Mullally A, Poveromo L, Schneider RK, Al-Shahrour F, Lane SW, Ebert BL. Distinct roles for long-term hematopoietic stem cells and erythroid precursor cells in a murine model of Jak2V617F-mediated polycythemia vera. Blood. 2012;120(1):166–72.
Sanjuan-Pla A, Macaulay IC, Jensen CT, Woll PS, Luis TC, Mead A, et al. Platelet-biased stem cells reside at the apex of the haematopoietic stem-cell hierarchy. Nature. 2013;502(7470):232–6.
Carrelha J, Meng Y, Kettyle LM, Luis TC, Norfo R, Alcolea V, et al. Hierarchically related lineage-restricted fates of multipotent haematopoietic stem cells. Nature. 2018;554(7690):106–11.
Psaila B, Wang G, Rodriguez-Meira A, Li R, Heuston EF, Murphy L, et al. Single-cell analyses reveal megakaryocyte-biased hematopoiesis in myelofibrosis and identify mutant clone-specific targets. Mol Cell. 2020;78(3):477-92 e8.
Tong J, Sun T, Ma S, Zhao Y, Ju M, Gao Y, et al. Hematopoietic stem cell heterogeneity is linked to the initiation and therapeutic response of myeloproliferative neoplasms. Cell Stem Cell. 2021;28(3):502-13 e6.
Schneider RK, Mullally A, Dugourd A, Peisker F, Hoogenboezem R, Van Strien PMH, et al. Gli1(+) mesenchymal stromal cells are a key driver of bone marrow fibrosis and an important cellular therapeutic target. Cell Stem Cell. 2017;20(6):785-800 e8.
Decker M, Martinez-Morentin L, Wang G, Lee Y, Liu Q, Leslie J, et al. Leptin-receptor-expressing bone marrow stromal cells are myofibroblasts in primary myelofibrosis. Nat Cell Biol. 2017;19(6):677–88.
Leimkuhler NB, Gleitz HFE, Ronghui L, Snoeren IAM, Fuchs SNR, Nagai JS, et al. Heterogeneous bone-marrow stromal progenitors drive myelofibrosis via a druggable alarmin axis. Cell Stem Cell. 2021;28(4):637-52 e8.
Acuna-Hidalgo R, Sengul H, Steehouwer M, van de Vorst M, Vermeulen SH, Kiemeney LALM, et al. Ultra-sensitive sequencing identifies high prevalence of clonal hematopoiesis-associated mutations throughout adult life. Am J Hum Genet. 2017;101(1):50–64.
Godley LA, Shimamura A. Genetic predisposition to hematologic malignancies: management and surveillance. Blood. 2017;130(4):424–32.
Sud A, Chattopadhyay S, Thomsen H, Sundquist K, Sundquist J, Houlston RS, et al. Familial risks of acute myeloid leukemia, myelodysplastic syndromes, and myeloproliferative neoplasms. Blood. 2018;132(9):973–6.
Rumi E, Passamonti F, Della Porta MG, Elena C, Arcaini L, Vanelli L, et al. Familial chronic myeloproliferative disorders: clinical phenotype and evidence of disease anticipation. J Clin Oncol. 2007;25(35):5630–5.
el El-Harith HA, Roesl C, Ballmaier M, Germeshausen M, Frye-Boukhriss H, von Neuhoff N, et al. Familial thrombocytosis caused by the novel germ-line mutation p.Pro106Leu in the MPL gene. Br J Haematol. 2009;144(2):185–94.
Ding J, Komatsu H, Wakita A, Kato-Uranishi M, Ito M, Satoh A, et al. Familial essential thrombocythemia associated with a dominant-positive activating mutation of the c-MPL gene, which encodes for the receptor for thrombopoietin. Blood. 2004;103(11):4198–200.
Jones AV, Chase A, Silver RT, Oscier D, Zoi K, Wang YL, et al. JAK2 haplotype is a major risk factor for the development of myeloproliferative neoplasms. Nat Genet. 2009;41(4):446–9.
Bao EL, Nandakumar SK, Liao X, Bick AG, Karjalainen J, Tabaka M, et al. Inherited myeloproliferative neoplasm risk affects haematopoietic stem cells. Nature. 2020;586(7831):769–75.
Li H, Xu Y, Mei H, Peng L, Li X, Tang J. The TERT rs2736100 polymorphism increases cancer risk: a meta-analysis. Oncotarget. 2017;8(24):38693–705.
Duncombe AS, Anderson LA, James G, de Vocht F, Fritschi L, Mesa R, et al. Modifiable lifestyle and medical risk factors associated with myeloproliferative neoplasms. Hemasphere. 2020;4(1):e327.
Stein BL, Williams DM, Wang NY, Rogers O, Isaacs MA, Pemmaraju N, et al. Sex differences in the JAK2 V617F allele burden in chronic myeloproliferative disorders. Haematologica. 2010;95(7):1090–7.
Moucadel V, Constantinescu SN. Differential STAT5 signaling by ligand-dependent and constitutively active cytokine receptors. J Biol Chem. 2005;280(14):13364–73.
Peeken JC, Jutzi JS, Wehrle J, Koellerer C, Staehle HF, Becker H, et al. Epigenetic regulation of NFE2 overexpression in myeloproliferative neoplasms. Blood. 2018;131(18):2065–73.
Liu F, Zhao X, Perna F, Wang L, Koppikar P, Abdel-Wahab O, et al. JAK2V617F-mediated phosphorylation of PRMT5 downregulates its methyltransferase activity and promotes myeloproliferation. Cancer Cell. 2011;19(2):283–94.
Dawson MA, Bannister AJ, Göttgens B, Foster SD, Bartke T, Green AR, et al. JAK2 phosphorylates histone H3Y41 and excludes HP1alpha from chromatin. Nature. 2009;461(7265):819–22.
Sashida G, Wang C, Tomioka T, Oshima M, Aoyama K, Kanai A, et al. The loss of Ezh2 drives the pathogenesis of myelofibrosis and sensitizes tumor-initiating cells to bromodomain inhibition. J Exp Med. 2016;213(8):1459–77.
Pérez C, Pascual M, Martín-Subero JI, Bellosillo B, Segura V, Delabesse E, et al. Aberrant DNA methylation profile of chronic and transformed classic Philadelphia-negative myeloproliferative neoplasms. Haematologica. 2013;98(9):1414–20.
McPherson S, Greenfield G, Andersen C, Grinfeld J, Hasselbalch H, Nangalia J, et al. Methylation age as a correlate for allele burden, disease status and clinical response in myeloproliferative neoplasm patients treated with Vorinostat. Exp Hematol. 2019;79:26–34.
Augello C, Cattaneo D, Bucelli C, Terrasi A, Fermo E, Martinelli I, et al. CD18 promoter methylation is associated with a higher risk of thrombotic complications in primary myelofibrosis. Ann Hematol. 2016;95(12):1965–9.
Broseus J, Alpermann T, Wulfert M, Florensa Brichs L, Jeromin S, Lippert E, et al. Age, JAK2(V617F) and SF3B1 mutations are the main predicting factors for survival in refractory anaemia with ring sideroblasts and marked thrombocytosis. Leukemia. 2013;27(9):1826–31.
Mascarenhas JO, Rampal RK, Kosiorek HE, Bhave R, Hexner E, Wang ES, et al. Phase 2 study of ruxolitinib and decitabine in patients with myeloproliferative neoplasm in accelerated and blast phase. Blood Adv. 2020;4(20):5246–56.
Brkic S, Meyer SC. Challenges and perspectives for therapeutic targeting of myeloproliferative neoplasms. Hemasphere. 2021;5(1):e516.
Coltro G, Rotunno G, Mannelli L, Mannarelli C, Fiaccabrino S, Romagnoli S, et al. RAS/CBL mutations predict resistance to JAK inhibitors in myelofibrosis and are associated with poor prognostic features. Blood Adv. 2020;4(15):3677–87.
Mylonas E, Yoshida K, Frick M, Hoyer K, Christen F, Kaeda J, et al. Single-cell analysis based dissection of clonality in myelofibrosis. Nat Commun. 2020;11(1):73.
Masarova L, Patel KP, Newberry KJ, Cortes J, Borthakur G, Konopleva M, et al. Pegylated interferon alfa-2a in patients with essential thrombocythaemia or polycythaemia vera: a post-hoc, median 83 month follow-up of an open-label, phase 2 trial. Lancet Haematol. 2017;4(4):e165–75.
Rao TN, Hansen N, Stetka J, Luque Paz D, Kalmer M, Hilfiker J, et al. JAK2-V617F and interferon-alpha induce megakaryocyte-biased stem cells characterized by decreased long-term functionality. Blood. 2021;137(16):2139–51.
Venugopal S, Mascarenhas J. Novel therapeutics in myeloproliferative neoplasms. J Hematol Oncol. 2020;13(1):162.
Acknowledgements
All figures in the manuscript were created at biorender.com
Funding
This review was undertaken within the Irish Clinical Academic Training (ICAT) Programme, supported by the Wellcome Trust and the Health Research Board (Grant Number 203930/B/16/Z), the Health Service Executive National Doctors Training and the Health and Social Care, Research and Development Division, Northern Ireland.
Author information
Authors and Affiliations
Contributions
GG undertook the literature review and wrote the manuscript. KM and MFM reviewed and amended the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
GG and KM have no competing interests to declare. MFM has undertaken advisory board work and received speaker fees from Novartis and undertaken advisory board work for Celgene, Incyte and AbbVie.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Greenfield, G., McMullin, M.F. & Mills, K. Molecular pathogenesis of the myeloproliferative neoplasms. J Hematol Oncol 14, 103 (2021). https://doi.org/10.1186/s13045-021-01116-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13045-021-01116-z