
Abstract
Background
Bone disease causes short-term or long-term physical pain and disability. It is necessary to explore new drug for bone-related disease. This study aimed to explore the role and mechanism of Salidroside in promoting osteogenic differentiation of adipose-derived stromal cells (ADSCs).
Methods
ADSCs were isolated and treated with different dose of Salidroside. Cell count kit-8 (CCK-8) assay was performed to assess the cell viability of ADSCs. Then, ALP and ARS staining were conducted to assess the early and late osteogenic capacity of ADSCs, respectively.
Then, differentially expressed genes were obtained by R software. Then, Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis of the differentially expressed genes were further analyzed. The expression of OCN, COL1A1, RUNX2, WNT3A, and β-catenin were measured by real-time PCR and Western blot analysis. Last, β-catenin was silenced by small interfering RNA.
Results
Salidroside significantly increased the ADSCs viability at a dose-response manner. Moreover, Salidroside enhanced osteogenic capacity of ADSCs, which are identified by enhanced ALP activity and calcium deposition. A total of 543 differentially expressed genes were identified between normal and Salidroside-treated ADSCs. Among these differentially expressed genes, 345 genes were upregulated and 198 genes were downregulated. Differentially expressed genes enriched in the Wnt/β-catenin signaling pathway. Western blot assay indicated that Salidroside enhanced the WNT3A and β-catenin expression. Silencing β-catenin partially reversed the promotion effects of Salidroside. PCR and Western blot results further confirmed these results.
Conclusion
Salidroside promoted osteogenic differentiation of ADSCs through Wnt/β-catenin signaling pathway.
Similar content being viewed by others
Background
Bone disease causes short-term or long-term physical pain and disability. Osteoblast differentiation is a highly regulated process controlled by a complex signaling pathway [1, 2]. Imbalance between bone formation by osteoblasts and bone resorption by osteoclasts contributes to osteoporosis [3, 4].
At present, drugs for the treatment of osteoporosis can be aimed at promoting osteoblastic bone formation or inhibiting osteoclasts [5]. Among them, parathyroid hormone is the only drug approved by the US Food and Drug Administration to promote bone formation. However, parathyroid hormone can cause osteosarcoma, which limits its large-scale clinical application [6]. Therefore, people’s attention has shifted to natural compounds.
The rhizome and roots of Rhodiola rosea L. have long been used in traditional medicine of China. Salidroside (SR) is a main component of Rhodiola rosea L. and exhibits a variety of pharmacologic properties, including anti-inflammatory, anti-fatigue and anti-oxidant properties, anti-apoptosis, and hypoglycemic effects. Mao et al. [7] found that Salidroside contributes to anti-aging effect in D-galactose induced aging model. Moreover, Salidroside protects cardiomyocyte against hypoxia-induced death through HIF-1α and VEGF-mediated pathway [8]. Zhang et al. [9] revealed that salidroside has protective effects against Abeta (25-35)-induced oxidative stress, which might be a potential therapeutic agent for treating or preventing neurodegenerative diseases. Studies have shown that a variety of natural compounds can affect the differentiation of osteoblasts [10]. However, whether SR could promote bone formation is not yet known.
In the treatment of many different bone diseases, mesenchymal stem cells were widely used in cell therapy [11]. Mesenchymal stem cells have the potential to differentiate into many different cell types such as osteoblasts, chondrocytes, muscle cells, and adipocytes [12,13,14]. Stem cells can be separated from bone marrow, synovium, periosteum, and fat tissue [14,15,16]. Among them, the source of adipose stem cells is abundant and easy to obtain, with little trauma [17]. Moreover, adipose stem cells proliferate faster than bone marrow stem cells. Therefore, adipose stem cells have more advantages than bone marrow stem cells in the cell therapy of bone diseases. Studies have shown that the adipose-derived stromal cells (ADSCs) have active biological properties, and its osteogenic differentiation are critical to the repair of bone loss and bone defect diseases [18]. Adipose-derived mesenchymal stem cells are easy to differentiate into adipocytes rather than osteoblast.
It was also reported that Wnt/β-catenin signaling is anabolic for bone formation [19]. The Wnt signaling pathway influences bone formation during development and bone remodeling during tissue renewal [20, 21]. Wnt inhibitors can promote preadipocyte differentiation by inhibiting Wnt/β-catenin signaling [22]. Therefore, the Wnt/β-catenin signaling pathway serves a notable role in osteogenic differentiation.
In this study, we performed RNA sequencing between normal and Salidroside-treated ADSCs. Moreover, we performed a series of experiments to verify that Salidroside promoted osteogenic differentiation of ADSCs through Wnt/β-catenin signaling pathway.
Material and methods
Isolation and culture of human adipose stem cells
Abdominal subcutaneous adipose tissue was obtained from human adipose tissue through simple liposuction. Then, 200–300 mL sterile PBS was added to every 0.5 g adipose tissue to prevent dehydration. Adipose tissues were cut into small pieces about less than 1 mm3 in size. Sterile saline (37 °C) was added to the homogenized adipose tissue in a ratio of 3:1 (saline to adipose tissue), followed by the addition of stock collagenase solution to a final concentration of 0.5 units/ml. This suspension was placed in a 37 °C shaker at 200 rpm for 60 min. Samples were then passed through a 100-μm filter into a 50-ml conical tube. After washing three times with sterile PBS, cells were then resuspend in high-glucose DMEM containing 20% FBS.
Osteogenic differentiation of ADSCs
The third passage (P3) cells were used for all subsequent experiments. When 80% of the ADSCs were confluent, cells were seeded in 6-well plates and decomposed overnight. Then, ADSCs were changed to induction medium (10% high-glucose DMEM containing 10 mM β-glycerol phosphate, 10 nM dexamethasone, and 60 mM ascorbic acid). ADSCs were treated with osteogenic induction medium for 3 weeks and the medium was changed every 3 days.
ALP staining and ALP activity
The ALP staining was performed using the ALP staining kit according to the instructions. In brief, ADSCs were fixed by 4% paraformaldehyde and then washed with PBS for three times. Staining was performed using BCIP/NBT chromogenic substrate (Vectastain ABC-AmP Kit, Vector Laboratories). An optical microscope (Olympus, Japan) was implemented for the staining result observation and photographing. ALP activity was measured according to the kit instructions. In brief, ADSCs were washed twice with TB buffer and then added 100 μL lysis solution. ADSCs were then separated for 20 min at 12,000 rpm at 4 °C. Then, 45 μL supernatant was added to 100 μL p-nitrophosphatate. After incubating for 30 min, the absorbance at 450 nm was measured with Microplate Reader (BioTek Instruments).
Alizarin Red Staining
The content of calcium was measured by Alizarin Red Staining. In brief, 21 days after induction, ADSCs were fixed by 4% paraformaldehyde for 5 min and then washed by PBS three times. Then, ADSCs were incubated with 0.2% Alizarin Red solution for 30 min. Finally, optical microscope (Olympus, Japan) was implemented for the staining result observation and photographing.
RNA sequencing
ADSCs underwent osteogenic differentiation and then divided into two groups: control and Salidroside-treated groups. ADSCs were treated with Trizol reagent to extract total RNA. Solexa pipeline v1.8 (Off-Line base Caller software, v1.8 Illumina, Foster City, CA, USA) was used to perform image analysis and substrate call. FastQC Software (Shanghai Kangcheng Biotechnology Co., Ltd.) was used to assess the quality and integrity of the RNA samples. HISAT2 software was used to align reads to the reference genome sequence (hg38). Transcripts from all samples were merged using StringTie to create a new reference file, which was then used to estimate transcript abundance in each sample. Differential expression analysis of the genes was performed using the DESeq package in R software (version 3.5.1). The heatmap of differentially expressed genes was drawn using the Complex Heatmap package in R. Gene ontology of the differentially expressed genes mainly including biological process, cellular component, and molecular function. Gene Ontology (GO) was performed using the clusterProfiler package in R software. The Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis of differentially expressed genes was performed using R software (version 3.5.1).
Construction and amplification of recombinant adenoviruses expressing siβ-catenin
For constructing adenoviruses expressing siβ-catenin, siRNAs targeting the coding region of mouse β-catenin were assembled to an adenoviral shuttle vector. The siRNA target sites against human β-catenin-coding region were cloned into the pSES adenoviral shuttle vector to create recombinant adenoviruses. Polybrene (4 μg/mL) was used to increase adenovirus infection efficiency. The effects of the transfection on β-catenin reduction were assayed by Western blot analysis.
Real-time polymerase chain reaction
Total RNA was extracted from ADSCs using the Trizol reagent in accordance with the manufacturer’s protocol. The first strand of cDNA was synthesized using the PrimeScript Reverse Transcriptase (RT) (TaKaRa Code: D2680S) and stored at − 20 °C for later use. The 7500 fast RT-PCR (ABI) was used for real-time quantitative PCR (qPCR). The expression of mRNA was calculated normalized to GAPDH with 2−△△Ct method. The primers used were as follows: OCN, 5′-TCTTAGAACAAATTCTGCCCTTT-3′ (forward) and 5′-TGCTTTGGTCTTGAAATCACA-3′ (reverse); COL1A, 5′-CCTCCTCAGCTCACCTTCTC-3′ (forward) and 5′-GTTGGGAGCCCAAATAGAAA-3′ (reverse); RUNX2, 5′-AAGGTGTACGGCAAGGCTTC-3′(forward); 5′-CGTCAGAGCGAGTGAACCTC-3′ (reverse); GAPDH, 5′-CAATGTGGCCGAGGACTTTG-3′ (forward) and 5′-CATTCTCCTTAGAGAGAAGTGG − 3′ (reverse).
Western blot
After washing human adipose-derived mesenchymal stem cells three times with PBS, total protein was extracted using RIPA protein lysis buffer (Beyotime, Shanghai, China) with freshly added 1% protease inhibitor cocktail and 1 mM phenylmethylsulfonyl fluoride (PMSF). Equal amounts of protein were resolved on a SDS-PAGE gel. After SDS-PAGE, the proteins were transferred to a PVDF membrane by electro-blotting for 1 h at 100 v. Membranes were blocked with 4% (w/v) non-fat milk powder in TBST (0.1% Tween-20 in Tris-buffered saline) at room temperature for 2 h. Then, the membranes were incubated with primary antibodies on a shaker, overnight at 4 °C. After that, the membrane was incubated with HRP-conjugated second antibody at room temperature for 2 h. The determination of the grayscale value was processed using ImageJ software.
Statistical analysis
Statistical analysis was performed using the SPSS 20.0 statistical package (IBM Corp, Armonk, NY, USA). A statistically significant difference between groups was indicated at p < 0.05. Data are expressed as the mean ± standard deviation (SD). Differences were analyzed using one-way analysis of variance (ANOVA) followed by the Tukey’s post hoc test for multiple comparisons. Graphs were made using GraphPad Prism 7 (GraphPad Software).
Results
Salidroside enhanced cell viability of ADSCs
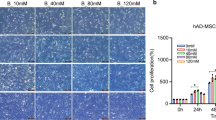
2D structure of Salidroside can be seen in Supplement S1. Salidroside As shown in Fig. 1, Salidroside slightly stimulated the proliferation of ADSCs within a 0.5–50-μM dose range, with 5 μM Salidroside showing maximal effect. However, the cell viability decreased slightly when the Salidroside concentration exceeded 5 μM.

Cell viability of ADSCs in control and different dose of Salidroside was measured by CCK-8 assay
Salidroside enhanced osteogenic differentiation of ADSCs
In order to assess the role of Salidroside in promoting osteogenic differentiation of ADSCs, ALP, and ARS were performed to assess the early and late osteogenic capacity, respectively. As shown in Fig. 2, results indicated that Salidroside showed a dose-dependent osteogenic promotion effect after Salidroside treatment, including early and late osteogenic differentiation effects. Simultaneously, PCR analysis showed that the expression of the osteoblastic differentiation markers OCN (Fig. 3A), COL1A1 (Fig. 3B), and RUNX2 (Fig. 3C) were dose-dependently increased after Salidroside treatment; the difference was statistically significant (P < 0.05).

Salidroside promoted osteogenic differentiation of ADSCs. ALP and ARS staining for ADSCs in control and different dose of Salidroside-treated groups

Real-time PCR analysis of OCN (A), COL1A1 (B), and RUNX2 (C) results of ADSCs in control and different dose of Salidroside-treated groups
Western blot analysis was in agreement with the quantitative real-time PCR (qRT-PCR) results, showing that the protein expression of OCN, COL1A1, and RUNX2 was upregulated in the Salidroside group (P < 0.05, Fig. 4).

Western blot assay to assess the osteogenic markers in control and different dose of Salidroside-treated groups
Bioinformatic analysis of Salidroside induced ADSCs
In order to identify the mechanism of Salidroside for promoting osteogenic differentiation of ADSCs. RNA sequencing and bioinformatic analysis were performed to reveal the mechanism of Salidroside in promoting osteogenic differentiation of ADSCs.
A total of 543 differentially expressed genes were identified between normal and Salidroside-treated ADSCs. Among these differentially expressed genes, 345 genes were upregulated and 198 genes were downregulated. Top fourteen differentially expressed genes can be seen in Fig. 5A.

Identify the differentially expressed genes in control and Salidroside-treated ADSCs. A Heatmap of the top fourteen differentially expressed genes between control and Salidroside-treated ADSCs. B Gene Ontology of the differentially expressed genes. C KEGG pathway of the differentially expressed genes
Gene ontology was divided into three categories: biological process, cellular component, and molecular function. Biological process of the differentially expressed genes mainly including Wnt signaling pathway, canonical Wnt signaling pathway, beta-catenin destruction complex disassembly, regulation of Wnt signaling pathway, regulation of canonical Wnt signaling pathway, cell surface receptor signaling pathway, animal organ development, tissue morphogenesis, and negative regulation of Wnt signaling pathway (Fig. 5B).
Cellular component of the differentially expressed genes mainly including Wnt signalosome, beta-catenin destruction complex, Wnt-Frizzled-LRP5/6 complex, catenin complex, extrinsic component of plasma membrane, plasma membrane protein complex, flotillin complex, beta-catenin-TCF complex, lateral plasma membrane, and plasma membrane (Fig. 5B).
Molecular function of the differentially expressed genes mainly including beta-catenin binding, Wnt-activated receptor activity, Wnt-protein binding, protein binding, gamma-catenin binding, protein domain specific binding, signaling receptor binding, frizzled binding coreceptor activity involved in Wnt signaling pathway, and armadillo repeat domain binding (Fig. 5B).
KEGG pathway mainly including Wnt signaling pathway, gastric cancer, hippo signaling pathway, breast cancer, hepatocellular carcinoma, basal cell carcinoma, pathways in cancer, Cushing’s syndrome, endometrial cancer, and signaling pathways regulating pluripotency of stem cells (Fig. 5C).
Salidroside significantly activated Wnt/β-catenin signaling pathway
In order to assess the mechanism of Salidroside in promoting osteogenic differentiation of ADSCs. We selected Wnt/β-catenin signaling pathway for further analysis. We found that Salidroside significantly increased the Wnt3A and β-catenin expression than control group (Fig. 6, P < 0.05).

Salidroside enhanced osteogenic differentiation of ADSCs through Wnt/β-catenin signaling pathway. Western blot assay that assessed the WNT3A and β-catenin in control and Salidroside-treated ADSCs
Silencing β-catenin partially reversed the osteogenic promotion effects of Salidroside
In order to further confirm the Wnt/β-catenin signaling pathway involved into the Salidroside promoted osteogenic differentiation of ADSCs.
ALP and ARS were then performed to assess the role of Salidroside for promoting osteogenic differentiation of ADSCs. We found that silencing β-catenin partially reversed the ALP activity and calcium deposition that induced by Salidroside (Fig. 7A).

Silencing β-catenin partially reversed the promotion effects of Salidroside. A ALP and ARS staining in control, Salidroside, and Salidroside + si-β-catenin groups. B Western blot assay of the osteogenic markers in control, Salidroside, and Salidroside + si-β-catenin groups
Western blot analysis showing that the protein expression of OCN, COL1A1, and RUNX2 was partially reversed by silencing of β-catenin (P < 0.05, Fig. 7B).
Discussion
In this study, we identified that Salidroside promoted osteogenic differentiation of ADSCs through Wnt/β-catenin signaling pathway. We firstly performed cell count kit-8 (CCK-8) assay to identify the optimal dose range of Salidroside. Then, we performed ALP and ARS staining assays to identify the osteogenic promotion effects of Salidroside. RNA sequencing was then performed to identify the differentially expressed genes between normal and Salidroside-treated ADSCs. Differentially expressed genes mainly enriched in the Wnt/β-catenin signaling pathway. We finally constructed small interfering RNA of β-catenin to verify the mechanism of Salidroside.
Strengths of this study were as follows: (1) we firstly performed RNA sequencing to assess the mechanism of Salidroside in promoting osteogenic differentiation of ADSCs, and (2) results revealed that Salidroside promoted osteogenic differentiation of ADSCs through Wnt/β-catenin signaling pathway.
The process of bone formation includes the initial proliferation of osteoblasts and the development and maturation of the extracellular matrix and the final mineralization. The osteogenic differentiation starts from adhesion and proliferation of several cell types, such as mesenchymal stem cells, pre-osteoblasts, and osteoblasts. Firstly, we performed CCK-8 assay to identify the optimal dose of Salidroside. We selected 5 μM Salidroside for further study. Then, ALP and ARS staining were performed to assess the osteogenic capacity of Salidroside. We found that Salidroside significantly increased osteogenic capacity of ADSCs. This study was in accordance with a previous study, which found that Salidroside protect against bone loss induced by H2O2 [23]. Bai et al. [24] revealed that Salidroside obviously promotes the proliferation of mesenchymal stem cells. These results suggested that Salidroside may be as alternative targets for osteoporosis therapies.
One of the commonly applied methodologies is deep RNA sequencing of proliferating, which indicated changes in the mRNA expression profiles in treatment and control groups. A total of 543 differentially expressed genes were identified between normal and Salidroside-treated ADSCs. Most of the differentially expressed genes mainly enriched in Wnt/β-catenin signaling pathway. Thus, we assessed the Wnt/3A and β-catenin expression in control and Salidroside-treated ADSCs. We found that Salidroside significantly increased Wnt/3A and β-catenin expression. Wnt/β-catenin was involved into multiple cells differentiation including neural differentiation [25], preadipocyte differentiation [26], and enterocyte differentiation [27].
In order to further identify the Wnt/β-catenin involved into the process of osteogenic differentiation of ADSCs. We constructed siRNA of β-catenin to inhibit the Wnt/β-catenin signaling pathway. We found that silencing β-catenin partially blocked the promotion effects of Salidroside on the osteogenic differentiation of ADSCs. Therefore, we concluded that Salidroside promoted osteogenic differentiation of ADSCs through Wnt/β-catenin signaling pathway.
This study has several shortcomings that are worth mentioning. First, the present study did not perform in vivo experiments. Second, receptor of Salidroside did not identify in this study. Third, the optimal dose regimen of Salidroside has not been established.
Conclusion
In conclusion, our data indicate that Salidroside promoted osteogenic differentiation of ADSCs via the canonical Wnt/β-catenin signaling pathway. The findings in this study may provide insights into understanding the mechanism of Salidroside for osteogenesis. Salidroside may be a novel therapy for bone-related diseases.
Availability of data and materials
We state that the data will not be shared since all the raw data are present in the figures included in the article.
Abbreviations
- ADSCs:
-
Adipose-derived stromal cells
- CCK-8:
-
Cell count kit-8
- GO:
-
Gene ontology
- KEGG:
-
Kyoto encyclopedia of genes and genomes
- SR:
-
Salidroside
- RT:
-
Reverse transcriptase
- qPCR:
-
Real-time quantitative PCR
- PMSF:
-
Phenylmethylsulfonyl fluoride
- SD:
-
Standard deviation
- ANOVA:
-
One-way analysis of variance
References
You WL, Xu ZL. Curculigoside promotes osteogenic differentiation of ADSCs to prevent ovariectomized-induced osteoporosis. J Orthop Surg Res. 2021;16(1):279. https://doi.org/10.1186/s13018-021-02389-3.
Wu JQ, Mao LB, Liu LF, Li YM, Wu J, Yao J, et al. Identification of key genes and pathways of BMP-9-induced osteogenic differentiation of mesenchymal stem cells by integrated bioinformatics analysis. J Orthop Surg Res. 2021;16(1):273. https://doi.org/10.1186/s13018-021-02390-w.
Liu M, Ding H, Wang H, Wang M, Wu X, Gan L, et al. Moringa oleifera leaf extracts protect BMSC osteogenic induction following peroxidative damage by activating the PI3K/Akt/Foxo1 pathway. J Orthop Surg Res. 2021;16(1):150. https://doi.org/10.1186/s13018-021-02284-x.
Yang XM, Song YQ, Li L, Liu DM, Chen GD. miR-1249-5p regulates the osteogenic differentiation of ADSCs by targeting PDX1. J Orthop Surg Res. 2021;16(1):10. https://doi.org/10.1186/s13018-020-02147-x.
Wang F, Yang Z, He W, Song Q, Wang K, Zhou Y. Effects of icariin on the proliferation and osteogenic differentiation of human amniotic mesenchymal stem cells. J Orthop Surg Res. 2020;15(1):578. https://doi.org/10.1186/s13018-020-02076-9.
Anastasilakis AD, Makras P, Doulgeraki A, Polyzos SA, Guarnieri V, Papapoulos SE. Denosumab for the treatment of primary pediatric osteoporosis. Osteoporos Int. 2021. https://doi.org/10.1007/s00198-021-06002-5.
Mao GX, Deng HB, Yuan LG, et al. Protective role of salidroside against aging in a mouse model induced by D-galactose. Biomed Environ Sci. 2010;23(2):161–6. https://doi.org/10.1016/s0895-3988(10)60047-5.
Zhang J, Liu A, Hou R, Zhang J, Jia X, Jiang W, et al. Salidroside protects cardiomyocyte against hypoxia-induced death: a HIF-1alpha-activated and VEGF-mediated pathway. Eur J Pharmacol. 2009;607(1-3):6–14. https://doi.org/10.1016/j.ejphar.2009.01.046.
Zhang L, Yu H, Zhao X, Lin X, Tan C, Cao G, et al. Neuroprotective effects of salidroside against beta-amyloid-induced oxidative stress in SH-SY5Y human neuroblastoma cells. Neurochem Int. 2010;57(5):547–55. https://doi.org/10.1016/j.neuint.2010.06.021.
Chen JJ, Zhang NF, Mao GX, He XB, Zhan YC, Deng HB, et al. Salidroside stimulates osteoblast differentiation through BMP signaling pathway. Food Chem Toxicol. 2013;62:499–505. https://doi.org/10.1016/j.fct.2013.09.019.
Liu F, Yuan Y, Bai L, Yuan L, Li L, Liu J, et al. LRRc17 controls BMSC senescence via mitophagy and inhibits the therapeutic effect of BMSCs on ovariectomy-induced bone loss. Redox Biol. 2021;43:101963. https://doi.org/10.1016/j.redox.2021.101963.
Zhang L, Wang Q, Su H, Cheng J. Exosomes from adipose derived mesenchymal stem cells alleviate diabetic osteoporosis in rats through suppressing NLRP3 inflammasome activation in osteoclasts. J Biosci Bioeng. 2021;131(6):671–8. https://doi.org/10.1016/j.jbiosc.2021.02.007.
Lamparelli EP, Lovecchio J. Chondrogenic commitment of human bone marrow mesenchymal stem cells in a perfused collagen hydrogel functionalized with hTGF-β1-releasing PLGA microcarrier. Pharmaceutics. 2021;13(3). https://doi.org/10.3390/pharmaceutics13030399.
Ciardulli MC, Marino L. Dose-response tendon-specific markers induction by growth differentiation factor-5 in human bone marrow and umbilical cord mesenchymal stem cells. Int J Mol Sci. 2020;21(16). https://doi.org/10.3390/ijms21165905.
Lanzillotti C, De Mattei M, Mazziotta C, et al. Long non-coding RNAs and microRNAs interplay in osteogenic differentiation of mesenchymal stem cells. Front Cell Dev Biol. 2021;9:646032. https://doi.org/10.3389/fcell.2021.646032.
Giai Via A, McCarthy MB, de Girolamo L, et al. Making them commit: strategies to influence phenotypic hifferentiation in mesenchymal stem cells. Sports Med Arthrosc Rev. 2018;26(2):64–9. https://doi.org/10.1097/jsa.0000000000000187.
Mazini L, Ezzoubi M, Malka G. Overview of current adipose-derived stem cell (ADSCs) processing involved in therapeutic advancements: flow chart and regulation updates before and after COVID-19. Stem Cell Res Ther. 2021;12(1):1. https://doi.org/10.1186/s13287-020-02006-w.
Zhang M, Matinlinna JP, Tsoi JKH, Liu W, Cui X, Lu WW, et al. Recent developments in biomaterials for long-bone segmental defect reconstruction: a narrative overview. J Orthop Translat. 2020;22:26–33. https://doi.org/10.1016/j.jot.2019.09.005.
Wang K, Zhao Z, Wang X, Zhang Y. BRD4 induces osteogenic differentiation of BMSCs via the Wnt/β-catenin signaling pathway. Tissue Cell. 2021;72:101555. https://doi.org/10.1016/j.tice.2021.101555.
Yang YY, Zhou YM, Xu JZ, Sun LH, Tao B, Wang WQ, et al. Lgr4 promotes aerobic glycolysis and differentiation in osteoblasts via the canonical Wnt/β-catenin pathway. J Bone Miner Res. 2021. https://doi.org/10.1002/jbmr.4321.
Zhu H, Chen D, Xie X, Li Y, Fan T. Melittin inhibits lung metastasis of human osteosarcoma: evidence of Wnt/β-catenin signaling pathway participation. Toxicon. 2021;198:132–42. https://doi.org/10.1016/j.toxicon.2021.04.024.
Yang J, Shi BY. Dickkopf (Dkk)-2 is a beige fat-enriched adipokine to regulate adipogenesis. Biochem Biophys Res Commun. 2021;548:211–6. https://doi.org/10.1016/j.bbrc.2021.02.068.
Mao GX, Wang Y, Qiu Q, Deng HB, Yuan LG, Li RG, et al. Salidroside protects human fibroblast cells from premature senescence induced by H(2)O(2) partly through modulating oxidative status. Mech Ageing Dev. 2010;131(11-12):723–31. https://doi.org/10.1016/j.mad.2010.10.003.
Bai H, Wang CB, Ma XH, Wei YP, Xi R, Zhao Q, et al. Effects of salidroside on proliferation of bone marrow mesenchymal stem cells. Zhongguo Shi Yan Xue Ye Xue Za Zhi. 2014;22(4):1072–7. https://doi.org/10.7534/j.issn.1009-2137.2014.04.035.
Makdissy N, Haddad K, AlBacha JD, et al. Essential role of ATP6AP2 enrichment in caveolae/lipid raft microdomains for the induction of neuronal differentiation of stem cells. Stem Cell Res Ther. 2018;9(1):132. https://doi.org/10.1186/s13287-018-0862-9.
Wang Z, Dai Z, Luo Z, Zuo C. Identification of pyrvinium, an anthelmintic drug, as a novel anti-adipogenic compound based on the gene expression microarray and connectivity map. Molecules. 2019;24(13). https://doi.org/10.3390/molecules24132391.
Kim JT, Li C, Weiss HL, Zhou Y, Liu C, Wang Q, et al. Regulation of ketogenic enzyme HMGCS2 by Wnt/β-catenin/PPARγ pathway in intestinal cells. Cells. 2019;8(9). https://doi.org/10.3390/cells8091106.
Acknowledgements
Not applicable.
Funding
There is no funding for this article.
Author information
Authors and Affiliations
Contributions
XHL, FLC, and HLS design the study and conducted the experiments. FLC and HLS perform the statistically analyses. XHL, FLC, and HLS write the draft. XHL and FLC edited and confirmed the final manuscript. The authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Supplement S1
: 2 D structure of Salidroside.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Li, Xh., Chen, Fl. & Shen, Hl. Salidroside promoted osteogenic differentiation of adipose-derived stromal cells through Wnt/β-catenin signaling pathway. J Orthop Surg Res 16, 456 (2021). https://doi.org/10.1186/s13018-021-02598-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13018-021-02598-w