
Abstract
Stroke is a clinical syndrome characterized by an acute, focal neurological deficit, primarily caused by the occlusion or rupture of cerebral blood vessels. In stroke, neuroinflammation emerges as a pivotal event contributing to neuronal cell death. The occurrence and progression of neuroinflammation entail intricate processes, prominently featuring mitochondrial dysfunction and adaptive responses. Mitochondria, a double membrane-bound organelle are recognized as the “energy workshop” of the body. Brain is particularly vulnerable to mitochondrial disturbances due to its high energy demands from mitochondria-related energy production. The interplay between mitochondria and neuroinflammation plays a significant role in the pathogenesis of stroke. The biological and pathological consequences resulting from mitochondrial stress have substantial implications for cerebral function. Mitochondrial stress serves as an adaptive mechanism aimed at mitigating the stress induced by the import of misfolded proteins, which occurs in response to stroke. This adaptive response involves a reduction in misfolded protein accumulation and overall protein synthesis. The influence of mitochondrial stress on the pathological state of stroke is underscored by its capacity to interact with neuroinflammation. The impact of mitochondrial stress on neuroinflammation varies according to its severity. Moderate mitochondrial stress can bolster cellular adaptive defenses, enabling cells to better withstand detrimental stressors. In contrast, sustained and excessive mitochondrial stress detrimentally affects cellular and tissue integrity. The relationship between neuroinflammation and mitochondrial stress depends on the degree of mitochondrial stress present. Understanding its role in stroke pathogenesis is instrumental in excavating the novel treatment of stroke. This review aims to provide the evaluation of the cross-talk between mitochondrial stress and neuroinflammation within the context of stroke. We aim to reveal how mitochondrial stress affects neuroinflammation environment in stroke.
Similar content being viewed by others


Introduction
Stroke is a clinical syndrome characterized by an acute and focal neurological deficit, caused by the occlusion or rupture of cerebral vessels [1]. Stroke is classified into hemorrhagic stroke caused by blood vessel rupture, and ischemic stroke resulted from blood clot-induced obstruction of cerebral blood flow. Further categorizations within hemorrhagic stroke include intracerebral hemorrhage (ICH) and subarachnoid hemorrhage (SAH) [1, 2]. Notably, over 60% of all incident strokes is ischemia stroke, which constitutes the largest population of stroke [3]. It is noteworthy that stroke ranks as the second leading cause of death in the world. According to a report in 2019 by World Stroke Organization, stroke claimed the lives of over 6.50 million annually, and accounted for at least 1.43 million disability-adjusted life lost [4, 5]. Unfortunately, even a few effective medical or surgical therapies have been demonstrated to enhance the prognosis of stroke patients, survivors of stroke commonly contend with sequelae of neurological impairments and psychiatric disorders, exerting a profound impact on their daily functionality and occupational capacity. Therefore, dedicated research efforts into the pathophysiological mechanisms of stroke are essential for developing new treatments and ultimately improving outcomes for these patients.
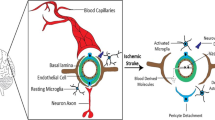
Neuroinflammation is the remarked event of stroke and also contributes to the neural cell death [5,6,7]. In stroke, the pathological condition increases the permeability of blood–brain barrier (BBB) [8], facilitating the infiltration of deleterious substances into neuronal microenvironment. Subsequently, microglia, a specific type of brain resident macrophage, are activated by the damage-associated molecular patterns (DAMPs) released from impaired neurons [9]. And these cells transformed to into the classical M1 phenotype and produce lots of inflammation-related cytokines to recruit other immune cells [10, 11], thereby fostering a neuroinflammation microenvironment in the brain. Similarly, the activation of other glia, such as astrocytes and oligodendrocytes, modulates the production and release of inflammatory cytokines in stroke. Indeed, neuroinflammation plays a pivotal role in causing brain tissue damage after both ischemic and hemorrhagic strokes. Understanding the mechanisms of neuroinflammation is essential for developing effective therapies to improve stroke outcomes.
As cellular powerhouses, mitochondria have garnered significant attention and reevaluation. Recent research has unveiled a profound correlation between mitochondrial dynamics and the pathophysiology of stroke [12, 13]. Mitochondrial dysfunction and adaptive alteration can affect the occurrence and development of neuroinflammation [14]. Dysfunctional mitochondrial dynamics inciting innate immune responses in both resident and infiltrating cells, such as microglia, astrocytes and oligodendrocytes, encompassing calcium-dependent immune activation, phosphorylation of transcription factors, cytokine secretion, organelle translocation, and the release of mitochondrial damage-associated molecular patterns (mDAMPs), even cell death [14, 15], impacting the outcome of stroke. Meanwhile, under neuroinflammation situation, the activation of microglia/astrocytes release various pro-inflammatory factors like IL-6, IL-1β, TNF-α and chemokine [16,17,18], promote the secretion of inflammasome [19]. Inflammasome and pro-inflammation factors associate with mitochondrial dysfunction lead to release reactive oxygen species (ROS) and mDAMPS which, in turn, aggravate inflammation consequently [20]. Eventually, the synergistic effect of neuroinflammation and mitochondrial stress cause brain edema, brain–blood barrier disruption and cell death [21, 22]. Therefore, understanding the correlation between mitochondrial and neuroinflammation in stroke pathogenesis is essential in excavating the novel treatment of stroke. In this review, we provide the evaluation of the cross-talk between mitochondrial stress and neuroinflammation, focusing on stroke. We aim to reveal how mitochondrial stress affects neuroinflammation environment in stroke.
Mitochondria and mitochondrial stress
Mitochondrion, a double membranes-bound organelle, is divided into four suborganellar compartments: outer mitochondrial membrane (OMM), inner mitochondrial membrane (IMM), intermembrane space between OMM and IMM (ISM) and mitochondrial matrix surrounded by IMM [23]. The matrix of human mitochondria contains mitochondrial circular DNA encoding 2 ribosomal RNAs (rRNA) including 12S and 16S rRNA, 13 proteins and 22 transfer RNAs (tRNA) [24]. Although mitochondrial genome is capable of encoding several factors required for the organelle activity, mitochondria still depend on nuclear genome. Most of mitochondrial proteins are encoded by nuclear genome. For example, only 11 proteins of proteins of oxidative phosphorylation (OXPHOS) are encoded by mitochondrial genome whereas 1300 OXPHOS-related proteins are nuclear gene products [25].
These nuclear genome-encoded proteins are transferred to mitochondria based on mitochondrial protein import pathways. Mitochondrial protein import is a conserved cellular process. In this process, most of mitochondrial proteins, which bind to some chaperones such as heat shock protein (HSP) 70 and 90, can be recognized by the translocase of the outer membrane (TOM) complex via their mitochondrial targeting pre-sequences (MTS). TOM complex is a symmetrical dimer that consists of 10 membrane protein subunits including TOM5, TOM6, TOM7, TOM22 and TOM40, which forming a shallow funnel on OMM [26,27,28,29,30]. After imported translocated across OMM, mitochondrial proteins are directed to the presequence translocase of the inner membrane complex (TIM23 complex), followed by the insert into IMM through the translocase of IMM (TIM22) complex, or the import to mitochondrial matrix by the mitochondrial import complex (MIM complex) [31].
Mitochondrial protein import involves the communication to multiple signal pathways in the cytoplasm and organelles [32]. Meanwhile, mitochondria provide the large amounts of energy for organelle activity in cellular processes [32]. Thus, the cross-talks of mitochondria and other organelle cause itself easy to respond to cellular stress. A series of mitochondrial response to stressors is called ‘mitochondrial stress’ (Fig. 1). The moderate mitochondrial stress promotes the cellular adaptive protection to resist harmful stressors. While the continuous mitochondrial stress impairs cells and tissues. Numerous constituents within mitochondria, along with metabolic byproducts, can function as mDAMPs. When released into the cytosol or extracellular environment after mitochondria injured, these mDAMPs trigger pro-inflammatory cytokine production and the recruitment of immune cells [15], which subsequently contributes to oxidative stress, inflammation, energetic impairment cell death and tissue injury [33, 34].

Mitochondrial stress response pathways. Environmental disturbance or impairment result in a decrease in mitochondrial stressor (ROS, mtDNA, ETC inhibition and PAMPs/DAPMs). Mitochondrial stress is activated to respond those stressors. A The mutant form of Ant1/Aac2 impairs the structural stability of TIM22, which in turn inhibits the biogenesis of TIM23 and induces instability in the IMM. This disruption results in proteostasis stress and triggers mitochondrial precursor overaccumulation stress (mPOS). B The unfolded protein response in mitochondria (UPRmt) is regulated by competing organelle targeting sequences within the transcription factor ATFS-1. When ATFS-1 is imported into the mitochondrial matrix via the MTS, it is subjected to degradation. However, in cases of mitochondrial dysfunction where ATFS-1 cannot be imported, it is redirected to the nucleus through the nuclear localization sequence (NLS). There, it activates the transcription of mitochondrial chaperones and proteases. C Under normal conditions, mitochondrial protein import undergoes rigorous monitoring by various quality control mechanisms, including ribosomal quality control (RQC) and mitochondrial translocation-associated degradation (mitoTAD). However, if RQC and mitoTAD are impaired due to mitochondrial dysfunction, a response known as mitochondrial compromised protein import response (mitoCPR) occurs within the cells. D Mitochondria transmit stress signals, activating the integrated stress response (ISR). This classic pathway involves four stress-activated kinases: PKR, PERK, GCN2, and HRI. They phosphorylate eIF2α during cellular stress, inhibiting protein biosynthesis and activating ATP4 transcription, thus promoting adaptive and apoptotic gene expression in response to stress. E Recognition and selective degradation of damaged mitochondria is mediated by mitophagy. The kinase PINK1 is stabilized specifically on damaged mitochondria where it recruits the ubiquitin ligase Parkin, which ubiquitinates multiple mitochondrial outer membrane proteins. Ubiquitinated mitochondria are then engulfed by autophagosomes and trafficked to lysosomes where they are degraded
Mitochondrial stress: stressor, stress response and communication mode
Stressors
Environmental disturbance or impairment result in a decrease in mitochondrial membrane protein (MMP) and misfolded protein accumulation in mitochondria. Mitochondrial stress is activated to respond these alterations. We introduce some of common stressors of mitochondrial stress.
ROS is the biomarker of oxidative stress, which can be produced in various location within the cell from plasma membrane to nucleus [35]. Mitochondrial ROS (mtROS) play a major role in the physiological or pathological condition, although ROS can be generated in many organelles [36]. The results of mtROS accumulation in mitochondria relate to the decrease of MMP, release of cytochrome C and leak of mitochondrial DNA (mtDNA) [37]. The ROS-induced release of cytochrome C and mnntDNA may involve gasdermin D (GSDMD), a pore-forming protein. A current report by Weindel et al. concluded mitochondrial ROS caused GSDMD to direct to mitochondrial membranes. which promoted mitochondrial content outflow to the cytoplasm [38]. Moreover, mitochondrial ROS is involved in the expression of nuclear genes. Peroxisome proliferator-activated receptor γ coactivator 1α (PGC-1α) is associated with the role of ROS in nuclear genome. PGC-1α is a transcript coactivator expressed in the tissues with high energetic demands [39]. PGC-1α is the core factor for oxidative stress and inflammation. On the one hand, PGC-1α interacts nuclear factor kappa B (NF-κB) p65 subunit to block its transcriptional activity on pro-inflammation genes, thereby playing the anti-inflammatory role [40]. As the negative feedback, NF-κB p65 directly inhibits PGC-1α via the binding between them [41]. On other hands, PGC-1α inhibits ROS accumulation in mitochondria via triggering the expression of ROS detoxification proteins such as superoxide dismutase 2 (SOD2) and glutathione peroxidase 1 (GPX1) [42]. The increased ROS activates NF-κB p65 to induce PGC-1α dysfunction, thereby causing the decreased expression of SOD2 and GPX1.
mtDNA is easily affected by environmental disturbance or impairment. mtDNA mutation contributes to mitochondrial stress. Zhang et al. showed mtDNA increase was associated with UPRmt in the C. elegans model [43]. The intriguing finding indicate mtDNA mutation has the potential of inducing stress signals via UPRmt. mtDNA mutation blocks the mitochondria–nuclear communication. Recently, a mouse model with mtDNA mutation develops the specific profile that mtDNA causes the depletion of total cellular nucleotides, suggesting the role of mtDNA mutation in nuclear genome stability [44].
ROS is the common byproduct of OXPHOS. There is a mature regulator mechanism to prevent ROS accumulation based on ROS detoxification proteins. For example, cells performing OXPHOS form the phenotype with the upregulated ERK5 that binds to the promoter of NRF2 as a transcript factor [45]. However, uncoupling of OXPHOS impairs the adjusting in cells and affects mitochondrial homeostasis. In response to the impairment of OXPHOS uncoupling, cells show the adaptive phenotype with the compensatory elevation in PGC-1α, COXIV and ANT3 and the rise in glycolytic metabolism but the reduction in cellular growth [46, 47]. OXPHOS uncoupling inhibits the translocation of mitochondrial protein from nuclear to mitochondria, which further induces mitochondrial unfolded protein response (UPRmt). Thus, OHPHOS uncoupling can cause UPRmt activation.
Electron transport chain (ETC) inhibition impairs OXPHOS and then triggers UPRmt. Mitochondria with ETC inhibition can develop the increased mitochondrial calpain 1 that has the capability of cleaving apoptosis inducing factor, a dual-function protein that can remove ROS in the physiological condition or translocate to nuclear to mediated caspase-dependent and caspase-independent apoptosis when sensing apoptosis stimuli [48]. A dominant mutation occurring in IMS protein is a cause of ETC-related mitochondrial stress. Mitochondrial coiled-coil-helix-coiled-coil-helix domain-containing 10 (CHCHD10) plays the core role of ETC integrity and mitochondrial structure via binding to mitochondrial nuclear retrograde regulator 1 that interacts with complex IV, the last electron receptor of ETC [49]. CHCHD10 mutation contributes to mitochondrial dysfunction with ETC inhibition [50]. Particularly, OMA1 is activated to induce mitochondrial fission, UPRmt and integrated stress response (ISR) when the dominant p.G58R mutation in CHCHD10 occurs [50].
Stress response
Mitochondrial precursor overaccumulation stress (mPOS)
Mitochondrial protein import is exquisitely modulated at multiple aspects, which causes mitochondria easier to develop the complex and multifaceted physiological consequences when exposed to stress. Some of pathogenic stressors that indirectly affect the core protein import inhibit cell adaption to stress through interfering the proteostasis network in the cytoplasm [51], which results in a decrease in protein import that further cause mitochondrial precursor overaccumulation in the cytoplasm. Cells adjust itself survival and death in response to mitochondrial precursor overaccumulation due to severe proteostasis stress, which is called ‘mPOS’ [52]. Ant1, an IMM protein related to ATP/ADP exchange, plays the key role in mPOS. The mutant of Ant1 is capable of inducing mPOS via disturbing the assembly and stability of mitochondrial membranes. The misfolded variants of Aac2, the homologue of Ant1 in yeast, can form aggregates to cause proteostasis stress [53]. Also, it impairs the structural stability of TIM22, which subsequently leads to the defect of TIM23 preprotein translocase [53]. The Aac2-induced instability of IMM and proteostasis stress triggers mPOS. Interestingly, human Ant1 activates a defense mechanism against mPOS in addition to the role in the induction of misfolded protein overaccumulation [54]. The overexpression of mutant Ant1 results in aggregate formation to activate protein degradation, autophagy and cell death, which is assumed as the anti-mPOS mechanism [54].
Mitochondrial unfolded protein response (UPRmt)
Mitochondrial unfold protein response (UPRmt) is a stress response that protects mitochondrial function by stop protein translation and degrade unfolded proteins when mitochondria is impaired by unfolded proteins [55]. UPRmt characterized by the activation of molecular chaperones and proteases is activated to decrease protein toxicity of mitochondrial stress. In UPRmt, activated chaperones are used for protein folding and proteases are for protein degradation [56]. The inefficient import of activating transcription factor associated with stress 1 (ATFS1) is the key event of UPRmt. It has been determined to co-ordinate the protective transcription response in yeast, C. elegans, and mammal during UPRmt. ATFS1 structure contains MTS mediating protein import in the normal condition and nuclear localization signal (NLS) promoting translocation to nuclear under stress [57]. When stressed, ATFS1 import is inhibited and then it, because of NLS, transfers to the nuclear for the transcription activation of targets, including Dnj10, HSP60, skn1, gpd2, TIM23 and TIM17 [57]. UPRmt has the ability to upregulate the efficiency of protein import to prolong mitochondrial lifespan. Intriguingly, ATFS1-dependent activation of UPRmt can promote mitochondrial import, despite its membrane potential is reduced [58], suggesting a positive feedback between UPRmt and relatively healthy mitochondria that these mitochondria transport signals to activate UPRmt which enhance the import of reversive protein for the assembly and biogenesis of mitochondria. In human, UPRmt is regulated by the homologue of ATFS-1 including activating transcription factor 4 (ATF4) and ATF5.
In addition to ATFS1, heat shock transcription factor 1 (HSF1) is also required for UPRmt. In UPRmt, ATFS1 is used to activate the transcription of nuclear genes that contribute to cell fitness involved in metabolism and proteases, while HSF1 plays the role of inducing mitochondrial chaperones. In response to mitochondrial stress, HSF1 is upregulated to bind constitutively to the promoters of genes encoding mitochondrial chaperones, including HSP60, HSP10 and mitochondrial HSP70 [59]. How does mitochondria transmit the stress signal to HSF1 during UPRmt? A recent study showed the potential mechanism involving mtROS and mitochondrial precursor overaccumulation (mtPO). During mitochondrial stress, mtROS released to the cytoplasm oxidates cytosolic HSP40, a co-chaperon of HSP70, which leads to HSP70 recruitment in mtPO that promotes the release of HSF1 to translocate in the nuclear [60]. HSF1-mediated transcription activation appears to stop at the end of the acute stress response, and this protein activated by acute stress cannot induce the transcription of its targets under chronic stress, which suggests HSF1 is required for acute stress rather than chronic or continuous stress [61].
Mitochondrial compromised protein import response (mitoCPR)
In the normal condition, mitochondrial protein import is under rigorous monitor by various mechanism of quality of control, such as ribosomal quality control [62] and mitochondrial translocation-associated degradation [63]. However, the common maintaining mechanisms are not enough to meet the overaccumulation of mitochondrial protein during stress. The Yeast develops compensatory and acute mechanism to mitigate protein import stagnation in the pathological condition, which may be not conversed in higher eukaryotes. In yeast, Pdr3, a transcription factor, induces Cis1 expression that is capable of recruiting Msp1 translocation to TOM complex, which promote the stalled import protein to be degraded in the cytoplasm [64]. The acute mechanism responding to mitochondrial stress is called ‘mitoCPR’. It remains unclear whether mitoCPR occurs in human since there is no obvious ortholog of Pdr3. Moreover, it needs to be uncovered whether mitoCPR function as a pre-order mechanism to respond to the inefficient protein import before mPOS and UPRmt [65].
Integrated stress response (ISR)
Mitochondrial stress is not a single mechanism. It demands the co-ordination of multiple cellular response and signaling pathways. Mitochondria can transmit stress signals to trigger in ISR. The classical ISR pathway consists of four stress-activated kinases, including Protein kinase R, Protein kinase R-like endoplasmic reticulum kinase, general control nonderepressible 2 and heme-regulated inhibitor (HRI) that have the ability to phosphorylate eukaryotic initiation factor 2α (eIF2α) when stressed [66]. Phosphorylated eIF2α inhibits the whole biosynthesis of protein via its activity in transcription inhibition. Meanwhile, it also activates ATP4 transcription to promote the expression of adaptive and apoptotic genes in response to stress [67]. Recent insight into integrated stress response concludes stressed mitochondria leads to the PHB2-STOML2 complex disintegration, thereby activating OMA1 to cleave DELE1 that binds to HRI after relayed to the cytoplasm and induce HRI-mediated eIF2α phosphorylation [67, 68].
Mitophagy
Mitophagy is a cellular process wherein mitochondrial-derived vesicles selectively engulf specific mitochondrial components and transport them to lysosomes or peroxisomes for degradation [69]. This quality control mechanism is pivotal for maintaining mitochondrial content and metabolic homeostasis [69, 70]. In cells, mitochondrial stress and mitophagy are two axes to resist the pathological condition. The former is used to improve the inefficient protein import to mitochondria, and the latter contributes to removing impaired mitochondria [70]. An imbalance between these processes can result in the accumulation of dysfunctional mitochondria, heightened oxygen consumption, and excessive production of ROS [70]. Ultimately, this imbalance can lead to cellular degeneration and activation of cell death pathways [70]. The co-ordination of mitochondrial stress and mitophagy is observed in recent studies. For example, mitophagy can promote global inhibition to protein biogenesis during mPOS via removing the impair mitochondria. In response to mPOS, cells trigger LC3 lipidation to activate mitophagy via NLRX1/RRBP1 complex [71]. There is the co-ordination between mitophagy and UPRmt. Myocardial mitochondria show the co-ordination between mitophagy and UPRmt to protect mitochondria from stress [72]. In C. elegans, FNDC1, an ortholog of FUN14 domain containing 1 (FUNDC1), and ATFS1 [73] mediate the cross-talk between mitophagy and UPRmt. Dysfunction of FNDC1-mediated mitophagy contributed to impaired mitochondria accumulation that cause UPRmt activation [73]. In turn, mitochondrial stress is also able to induce FUNDC1-mediated mitophagy [74].
Communication mode
Nuclear genome encodes most of mitochondrial protein. The forward signaling from nuclear to mitochondria is required for the homeostasis and integrity of mitochondria. Mitochondria also are able to influence the expression of nuclear genes via various communication mediators, which is named ‘mitochondrial retrograde signaling’. Mitochondrial retrograde signaling is responsible for transmitting mitochondrial stress to cells. We conclude common communication molecules in mitochondrial retrograde signaling.
ROS is not just a stressor, but also serves as a communication molecule of mitochondrial stress. ETC inhibition due to OXPHOS uncoupling-related proton leak to promote one electron reduction of oxygen to superoxide, which contributes to the production and release of mitochondrial ROS [75]. Then, ROS, as the signal, activates NRF2-related UCP1 expression (a protein for mitochondrial uncoupling) [76,77,78] and ATF family-mediated UPRmt [79, 80], and recruits Parkin to mitochondria for mitophagy [81, 82].
In multicellular organisms, local stress can be transmitted to the distant cell/tissue to induce systemic regulation for the whole metabolism. The non-autonomous response is mediated by a series of factors named ‘mitokine’. Fibroblast growth factor 21 (FGF21) and growth differentiation factor 15 (GDF15) are two mitokines to be most discussed. FGF21 and GDF15 can be used for the biomarkers in the first-line diagnosis of mitochondrial disease [83, 84]. FGF21 and GDF15 show the different roles in metabolic adaption to mitochondrial stress although both of them are capable of transmitting stress signals.
FGF21 is upregulated in mitochondrial stress. Stressors such as OXPHOS uncoupling increase FGF21 expression to transmit ISR [85]. Mitochondrial dysfunction caused the upregulation of Fgf21 mRNA expression in nucleus [86], upregulation of FGF21 is instrumental in cardio-protection and mitochondrial quantity control. FGF21 is able to activate AMPK/FOXO3/SIRT3 pathway to inhibit the mitochondrial dysfunction in cardiomyocytes [87]. Li et al. showed depletion of FGF21 contributes to the production of mitochondrial ROS [88]. They further determined mitochondrial fusion was activated by FGF21 based on AMPK pathway. FGF21 plays the role in systemic metabolism and mitochondrial stress via interacting with β-klotho, an obligatory co-receptor for FGF21. In cells, the C terminal of FGF21 binds to β-klotho to activate fibroblast growth factor receptor, which promotes cell signaling transduction [89]. Notably, FGF21 shows a characteristic with diurnal oscillations. Circulating FGF21 reaches the peak at the early morning, and then decrease to the minimum at the afternoon [90], suggesting the dynamic role of FGF21 in metabolism regulation.
Similar to FGF21, GDF15 is increased by mitochondrial stressors. In nucleus Crif-1 increase Gdf15 RNA expression via UPRmt/ATF4/CHOP pathway [91]. GDF15 can be induced by ETC inhibition to mediate systemic metabolic flexibility related to glycolipid metabolism [92]. Also, exercise leads to the dramatic upregulation of plasma GDF15 [93]. GDF15 is expressed in multiple tissues, such as lung, heart, brain, kidney, liver, adipose, skeletal muscle and gastrointestinal system. There is a specific receptor of GDF15 expressed, called as ‘GDNF-family receptor α-like (GFRAL)’ in the hindbrain. GDF15 is capable of interacting with GFRAL to orchestrate its role in lipid metabolism. Interestingly, GDF15 also shows the diurnal rhythm in the body. Circulating GDF15 in the day is higher than that in the night in a UCP1-tg mice developing mitochondrial uncoupling [92].
Cross-talk between mitochondrial stress and neuroinflammation in stroke
Neuroinflammation, the inflammatory response following neuronal injury, plays a pivotal role in stroke pathogenesis [9, 94]. Under mitochondrial stress, diverse cell types, such as astrocytes, microglia, endothelial cells, and leukocytes, release pro-inflammatory agents, including chemokines, cytokines, and enzymes. These actions collectively contribute to damage in the cerebral parenchyma [6]. Notably, accumulating evidence highlights the dual nature of these cells in inflammation, with their effects being either advantageous or harmful, depending on the timing of pathway activation or inhibition [95]. Mitochondrial stress after stroke is capable of interacting with neuroinflammation lead to neurovascular unit (NVU) dysfunctions, including neuron death, BBB disruption, and neuroinflammation, and affect the outcome of stroke [13]. The turbulences of mitochondrial function include increased fission, deficient fusion, and impaired or excessive mitophagy [12]. However, the role of mitochondrial stress in neuroinflammation and stroke is still under debate and we will discuss whether mitochondrial stress promotes or inhibits neuroinflammation in stroke in this section (Fig. 2).

Cross-talk between mitochondrial stress and neuroinflammation in stroke. Mitochondrial stress serves as an adaptive mechanism aimed at mitigating the stress induced by the import of misfolded proteins, which occurs in response to a stroke. The significance of mitochondrial stress in the pathogenesis of stroke is highlighted by its ability to interact with neuroinflammation. The impact of mitochondrial stress on neuroinflammation varies depending on its severity. Moderate mitochondrial stress can modulate neuroinflammation, enabling cells to better withstand harmful stressors and protect neurons. In contrast, sustained and excessive mitochondrial stress promotes neuroinflammation, leading to the release of pro-inflammatory cytokines, which negatively impacts cellular and tissue integrity
Mitochondrial stress modulates neuroinflammation to resist stroke
Although much of the research described previously has characterized mitochondrial stress as a deleterious process in stroke-induced brain injury, there is also evidence pointing to its neuroprotective effects.
Mitochondrial stress has been employed as a strategy to mitigate excessive damage resulting from neuroinflammation. A study by Zhu et al. demonstrated the activation of microglia and the UPRmt (reflecting mitochondrial stress) are involved in the pathogenic mechanism underlying hydrocephalus in a kaolin-induced hydrocephalus mouse model in C57BL/6J mice [96]. Subsequent research by the same group suggested a link between pro-inflammatory microglial polarization and UPRmt [97]. ATF5 functions as an upstream regulator that mediates the mitochondrial integrated stress response (ISR). The ATF5–UPRmt axis plays a vital role in facilitating the repair of impaired mitochondria and upholding protein homeostasis within the cellular environment [98]. Specifically, they found that microglia exposed to lipopolysaccharide stimulation exhibited an upregulation of ATF5-dependent UPRmt, which significantly restrained the production of pro-inflammatory cytokines, including TNF-α, IL-6, and IL-1β [97]. This implies that ATF5-dependent UPRmt may serve as a regulatory mechanism to dampen the inflammatory response in microglia and potentially mitigate neuroinflammation-associated damage. Xie et al. found that the activation of UPRmt attenuated neuroinflammation after ischemic stroke [99], and in SAH UPRmt regulated the neuronal mitochondrial homeostasis, which decreased the early brain injury [100].
Moreover, mitochondrial fission is a complex process that entails the division of a single mitochondrion into two separate mitochondria [101]. Prior research has established that the excessive activation of mitochondrial fission is a contributing factor to brain injury following a stroke [102]. However, there is evidence suggesting a neuroprotective effect of mitochondrial fission through the elimination of impaired mitochondrial segments, facilitation of signal transduction, enhancement of ATP production, and stabilization of mitochondrial DNA (mtDNA) [101]. In a study conducted by Busceti et al., wherein selective overexpression of uncoupling protein-2 (UCP2) was carried out in the corpus striatum of stroke-prone spontaneously hypertensive rats, they observed an upregulation of OPA1 and Fis1 [103]. OPA1 is associated with mitochondrial fusion, while Fis1 is related to mitochondrial fission, both processes occurring in the striatum tissue [103]. While the exact causality was not explicitly established, this experiment suggested that UCP2 may exert a neuroprotective effect by modulating mitochondrial fusion and fission.
Furthermore, appropriate mitophagy is instrumental in the improvement of stroke. Mitochondrial stress induces mitophagy to remove impaired mitochondria. In response to the pathological condition, the impaired mitochondrion is divided into two parts, one of which is functional segment, and anther of which undergoes dysfunction. Subsequently, the dysfunctional mitochondrial part is eliminated by mitophagy [104].
In ischemic stroke
The ATP depletion and Ca2+ overloading due to the deletion of oxygen and glucose induce the ISR that the proteins located in mitochondria-associated ER membrane (MAMs) are activated or translocate from ER to MAMs to maintain Ca2+ homeostasis and function as the assembly platform for inflammasomes in the brain when responding to the ischemic condition [105]. Appropriate activation in MAMs contributes to ischemic stroke improvement via regulating neuroinflammation.
FUNDC1 and Voltage Dependent Anion Channel 1 (VDAC1) are two of the MAMs protein. Mitochondrial VDAC1 has been determined to mediate inflammation activation in ischemic stroke. VDAC1 is the key mitochondrial protein that mediates the release of mtDNA in mitochondrial stress. In ischemia, the interaction of VDAC1 with Bax causes Cytochrome C release to the cytoplasm, which subsequently trigger mitochondrial permeabilization to evoke NLRP3 inflammasome assembly and cytokine expressions, including TNF-α, IL-6, IL-12 and interferon α/β [106]. Importantly, the lysine lactylation of VDAC1 is inhibited in neurons during ischemia [107], which contributes to the changes in metabolic phenotype and macrophage polarization. FUNDC1 can sense mitochondrial stress via OPA1 and then activate mitophagy [74]. Mitochondrial stress due to ischemic stroke results in FUNDC1 translocation from ER to mitochondria. In ischemic condition, FUNDC1 holds the stronger binding to LC3B that is required for mitophagy activation [108, 109]. Mitophagy is used to remove impaired mitochondria that promote neuroinflammation [110]. Thus, mitochondrial stress modulates neuroinflammation via the co-ordination with FUNDC1-mediated mitophagy.
In addition to the role of MAMs-related ISR, ATF4 also plays the protective role in stroke-associated neuroinflammation. As mentioned above, mitochondrial stress is capable of resisting neuroinflammation via inducing mitophagy. When UPRmt is activated by ischemic condition, ATF4 can increase Parkin expression to cause mitophagy activation that has the ability to inhibit NLRP3 inflammasomes in the brain [111, 112]. Also, ATF4 is involved in the expressions of Bax, Bcl-2 and ER stress-related genes during ischemic stroke [113]. HSF1, another member of UPRmt, has been found to be related to ischemia disease pathogenesis [114]. After ischemia, HSF1 is highly enriched in the nuclear of P7 neurons and partly in microglia [115]. HSF1 has the capability to modulate the activation in microglia and astrocytes. HSF1 inhibits A1 astrocyte (a phenotype for neurotoxicity and neuroinflammation) through repressing MAPK/NF-κB pathway [116]. In microglia, HSF1 modulates gene expression that affect cell activation and inflammatory cytokines. For example, Liao et al. showed HSF1 could mediate the expression of miR-214-3p and nuclear factor of activated T cells 2 via its activity of transcription factor, thereby inhibiting microglia-mediated neuroinflammation [117]. Furthermore, dietary intake of n-3 polyunsaturated fatty acids attenuated mitochondrial oxidative stress and increased the mitophagy of astrocytes in the condition of hypoxia to limit A1-specific astrocyte polarization, thereby reducing the production of inflammatory cytokines and neurotoxins, subsequently improving the neurological outcomes of mice with ischemic stroke [118]. In addition, Sirtuin-3 (Sirt3), a NAD+-dependent deacetylase, was reported to modulate the UPRmt and attenuate neuroinflammation after cerebral ischemia [99]. Sirt3 is a mitochondrial NAD+-dependent protein deacetylase that regulates the balance of energy metabolism, ROS generation [119]. Sirt3 activates the UPRmt by increasing Foxo3a and Sphkl expression, subsequently Sphkl expression relieves neuroinflammation after ischemic stroke [99]. Thus, activation of UPRmt and mitophagy reveal neuroprotection by mitigate neuroinflammation.
In hemorrhagic stroke
We have mentioned that mild mitochondrial stress activates mitophagy to eliminate neuroinflammation in ischemic stroke. In fact, this important mechanism is triggered as well as in hemorrhagic stroke. A report focusing on SAH in rats suggested mitophagy contributed to the inhibition to oxidative stress and neuron death [110]. Animal experiments showed mitophagy was activated at 24 h after hemorrhagic stroke modeling, with the increases of mitophagy biomarkers including Beclin-2, LC3B and PINK1 [120, 121]. Proteins in mitochondrial stress contributes to the mitophagy activation at this stage. In a study conducted on a SAH rat model, the administration of melatonin was found to reduce ROS levels by upregulating mitophagy [121, 122]. At the 24-h mark after SAH, melatonin treatment led to increased levels of mitophagy-associated proteins, specifically PINK and Parkin. Optineurin (OPTN) a protein can combine with ubiquitinated cargo contributing to mitophagy. OPTN is also associated with PINK1/Parkin-mediated mitophagy [123], which upregulates mitophagy and decreases NLRP3 activation in ICH [124]. This intervention resulted in a decrease in ROS content, mitigated morphological alterations in mitochondria, and ultimately inhibited the activation of the NLRP3 inflammasome [121]. In a rat SAH model, VDAC1 was found to be time-dependently upregulated within 48 h after SAH modeling, which induced mitophagy [125]. VDAC1 promoted mitophagy to reduce DAMPs and inhibit the death in neurons [125]. The role of VDAC1 in mitophagy may involve Ant1, an mPOS-related protein. The complex of VDAC1 and Ant1 drove mitophagy via causing MMP depolarization [126]. Importantly, Ant1 mediates the stability of PINK1 during mitophagy. Zoltan Arany et al. reported Ant1 promoted PINK1 accumulation on OMMs via closing TIM23-mediated PINK1 translocation in the pathological condition [127].
As described above, in mitochondria Sirt3 is a switch of mitochondrial energy regulation [128]. Activation of Sirt3, which promotes UPRmt, attenuates neuroinflammation and mitigates ATP, ROS and mitochondrial complex1 release after ICH [129, 130]. FGF21 functions as the mitokine that transmits stress signals for non-autonomous response. A clinical report determined serum FGF21 is the prognosis biomarker of ICH [131], suggesting its key role in hemorrhagic stroke. FGF21 has found to suppress the production of NF-κB-related cytokines during hemorrhagic disease. Thus, FGF21 has the potential to modulate neuroinflammation in hemorrhagic stroke. GDF15 is also the core mitokine for mitochondrial stress. GDF15 is significantly upregulated in patients with intracerebral and SAH [132]. However, the mechanism of GDF15 in hemorrhagic stroke has been not clearly described. Perhaps, GDF15 modulates cerebral microenvironment via GFRAL–RET heterodimer expressed in the hindbrain [133].
Continuous and severe mitochondrial stress aggravates neuroinflammation during stroke
Stroke pathogenesis involves mitochondrial stress [12]. Neuronal toxicity leads to mitochondrial impairment, including MMP reduction, the increase of protein aggresomes and the dysfunction in UPRmt [13]. Mitochondrial stress is the key event for quantifying control in mitochondria [12]. However, the continuous performance of mitochondrial stress may fail to restore mitochondrial homeostasis and induces the severe mitochondrial injury [134]. Mitochondrial stress responses become the cooperator of inflammation to induce the death in neurons during stroke when they are not enough to resist mitochondrial dysfunction in the pathological condition [13]. Mitochondrial stress affects glia activity. In this part, we review the reports showing the co-operation of severe mitochondrial stress and neuroinflammation in stroke, and describe the intriguing mechanism of severe mitochondrial stress in neuroinflammation based on common glia.
In ischemic stroke
Misfolded proteins are the main cause of secondary brain injury in ischemic stroke [13]. UPRmt is used to decrease misfolded mitochondrial proteins and alleviate the pressure of protein import into mitochondria. However, this response is not enough to inhibit the sustained accumulation of misfolded proteins in ischemic condition. The UPRmt-related protein ATF4 has been determined to be the pro-death transcription factor promoting the progression of ischemic stroke although it plays neuroprotective role in the early stage of ischemic stroke. Overexpression of ATF4 in neurons contributes to the susceptibility to oxidative death. Thus, ATF4 has the harmful effect on neurons undergoing ischemic condition. The mouse model of ischemic stroke with ATF4 deletion shows the resistance to the oxidative stress-induced death in neurons [135]. In cerebral ischemia, the ectopic expression of ATF4 promoted JUNB and ETS1 via the JMJD3-dependent demethylation of H3K27m3, which aggravated neuroinflammation and neuronal death in the brain [136].
Ischemic stroke pathogenesis causes abundant mitochondrial stressors in the central nervous system (CNS). Then, mitochondrial stress leads to the massive release of mitochondrial contents such as mtDNA and mtROS, which promotes pro-inflammatory cytokine production and immunocyte activation during ischemic stroke [12]. Sirt6 is a NAD+-dependent protein deacetylase and ribosylation enzyme, which mediated inflammatory cytokine secretion and ROS production [137]. Overexpression of Sirt6 in endothelial lessen ischemic region by reduce mtROS and oxidized mtDNA, meanwhile decrease TNF-α, IL-18 and IL-1β [22]. A laboratory study based on lipopolysaccharide-stimulated macrophage suggested mtROS activated the deubiquitination of NOD-like receptor thermal protein domain associated protein 3 (NLRP3) to induce NLRP3 inflammasomes activation [138]. NLRP3 inflammasomes commonly expressed in the CNS when ischemic stroke occurs are positively correlated to the cerebral infarct and neurological deficit during stroke [139]. It is instrumental in inhibiting neuron death and neurological deficit during ischemic stroke to remove mtROS by exogenous treatment [140, 141].
Glia positively contribute to neuroinflammation in ischemic stroke. Microglia are the most important immune cell in CNS. Microglia activation following ischemia release inflammatory cytokine which aggravates neuroinflammation. Mitochondrial fission, the most important pathway of mitophagy [142], in microglia was associated with neuroinflammation after cerebral ischemia. Mounting evidence shows that the Janus kinase (JAK) and signal transducers and transcription (STAT) were activated in inflammation [143,144,145]. Dynamin-related protein 1 (Drp1) is a multidomain GTPase provoke mitochondrial fission, which is modulated by JAK2/STAT3 signaling [146]. Thus, activation of JAK2/STAT3 upregulates Drp1 expression then promotes mitochondrial fission in microglia and mediates neuroinflammation. Impaired mitochondria in neurons can evoke astrocyte activation via a crossing cellular pathway [147]. Astrocyte plays dual roles in ischemic stroke. Activated astrocytes support neurological restoration, whereas they also produce pro-inflammatory cytokines to aggravate neuroinflammation in ischemic stroke progression [148]. The sustained signals from impaired mitochondria result in a large amount of cytokine by activated astrocytes, leading to the continuous inflammatory microenvironment in the CNS. Similar to astrocytes, the inflammatory microglia also are maintained by severe mitochondrial stress, which is one of the major causes of inflammation infiltration into the brain during ischemic stroke. Oligodendrocytes are more susceptible to the stress than other types of glia. The activity and morphology of oligodendroglial mitochondria are unique. The differentiation from oligodendrocyte progenitor cells (OPCs) to oligodendrocytes creates the high demand of mitochondria during myelin biosynthesis while OPCs rely less on mitochondria-mediated ATP metabolism after myelination [149]. OPCs positively regulate the sustained inflammation in the brain. Netrin-1 expressed in oligodendrocytes monitors the subcellular location and recruitment of mitochondria [150]. Mitochondria tend to induce oligodendroglial apoptosis in the pathological condition such as ischemia, resulting in myelin loss and neurodegeneration.
In hemorrhagic stroke
Mitochondria are the main source of ROS after hemorrhagic stroke. The animal experiment related to SAH showed ROS burst due to the stress-induced opening of mitochondrial permeability transition pore [151]. As described above, mtROS functions as the communication molecule of mitochondrial stress. Cells start the downstream pathway when sensing mtROS. For example, excessive mitochondrial fission was observed in ICH [11]. Suppression of excessive mitochondrial fission seems to contribute to reduce NLRP3 and pro-inflammatory including IL-1β, IL-18, consequently, attenuate neuronal pyroptosis in ICH [152]. MtROS and mtDNA have been found to play the key role in the activation of NLRP3 inflammasomes. NLRP3 inflammasomes mediate the occurrence and development of pyroptosis that is an inflammatory death in cells and accompanied by the massive release of inflammatory cytokines. NLRP3 inflammasomes were increased in the microglia of mice with hemorrhagic stroke. Impaired mitochondria can activate NLRP3-mediated pyroptosis through mtROS release, which further enhances neuroinflammation and aggravates neuronal injury in hemorrhagic stroke [120, 121, 153].
ATF4, similar to its role in ischemic stroke, becomes a significant target of inhibiting neuronal death in several rodent models of hemorrhagic stroke [154]. Liu et al. identified ATF4 as the differentially expressed gene of ICH and predicted its function was involved in TNF-mediated inflammation [155]. They subsequently determined that ATF4 was upregulated in the rat model of cerebral hemorrhage, and found ATF4 downregulation was instrumental in inhibiting inflammation. Min et al. reported the activated microglia-derived exosomes improved necroptosis in neurons undergoing ICH via reducing ATF4 [156]. In mice with ICH, activated GCNF-eIF2α pathway inhibited ATF4 activity to reduce inflammation infiltration into the CNS [157].
In hemorrhagic stroke-induced neuronal injury, UPRmt dysfunction is associated with the decrease in the complex of nucleotide exchange factor and HSP70. A study focusing UPRmt in hemorrhagic stroke showed a mechanism between GrpE like 1, mitochondrial (GrpEL1), a nucleotide exchange factor, and mitochondrial HSP70 (mtHSP70). Although mtHSP70 upregulation was observed in oxyhemoglobin-induced cell model of hemorrhagic stroke, the constitutive binding of GrpEL1 and mtHSP70 was decreased, which resulted in the UPRmt inhibition that contributed to mitochondrial dysfunction and impairment [100]. Obviously, the sustained pathological condition in neurons contributes to the overaccumulation of mitochondrial stress response. The sustained mitochondrial stress further impairs mitochondrial genome [158], which may serve as the major cause of mitochondrial dysfunction in hemorrhagic stroke.
Conclusion
In summary, disruptions in mitochondrial homeostasis serve as an initial event contributing to NVU dysfunctions and play a pivotal role in the pathophysiology of stroke. The synergy between neuroinflammation and proper mitochondrial stress can effectively eliminate harmful stimuli and facilitate neurological recovery. Conversely, severe mitochondrial stress exacerbates neuroinflammation in the context of stroke. Such proper mitophagy appears to offer neuroprotection by eliminating damaged mitochondria, while excessive mitophagy can detrimentally impact energy production and mitochondria-associated signaling pathways. Consequently, inhibiting mitochondrial stress emerges as a potential and unique therapeutic approach for stroke. Key proteins involved in mitochondrial stress, such as ATF5, ATF4, HSF1, FGF21, and GDF15, hold promise as potential targets for modulating neuroinflammation during stroke.
Despite significant advancements in understanding the mechanisms underlying mitochondrial dynamics and mitophagy following stroke, this field remains complex and necessitates further investigation. The precise effects of mitophagy in the context of stroke demand further exploration, as they hold the potential to yield valuable insights into stroke treatment strategies. Techniques such as genome, transcriptome, proteome, and epigenome sequencing can uncover molecular heterogeneity, offering patient-specific insights into MQC and potentially paving the way for novel therapeutic interventions in stroke.
Availability of data and materials
Not applicable.
Abbreviations
- BBB:
-
Blood–brain barrier
- DAMPs:
-
Damage associated molecular patterns
- OMM:
-
Outer mitochondrial membrane
- IMM:
-
Inner mitochondrial membrane
- OXPHOS:
-
Oxidative phosphorylation
- HSP:
-
Heat shock protein
- TOM:
-
Translocase of the outer membrane
- MTS:
-
Mitochondrial targeting pre-sequences
- MMP:
-
Mitochondrial membrane protein
- GSDMD:
-
Gasdermin D
- PGC-1α:
-
Peroxisome proliferator-activated receptor γ coactivator 1α
- NF-κB:
-
Nuclear factor kappa B
- SOD2:
-
Superoxide dismutase 2
- GPX1:
-
Glutathione peroxidase 1
- UPRmt :
-
Mitochondrial unfolded protein response
- ETC:
-
Electron transport chain
- CHCHD10:
-
Coiled-coil-helix-coiled-coil-helix domain-containing 10
- ISR:
-
Integrated stress response
- mPOS:
-
Mitochondrial precursor overaccumulation stress
- mitoCPR:
-
Mitochondrial compromised protein import response
- ATFS1:
-
Activating transcription factor associated with stress 1
- NLS:
-
Nuclear localization signal
- HSF1:
-
Heat shock transcription factor 1
- mtPO:
-
Mitochondrial precursor overaccumulation
- HRI:
-
Heme-regulated inhibitor
- eIF2α:
-
Eukaryotic initiation factor 2α
- NVU:
-
Neurovascular unit
- CNS:
-
Central nervous system
- OPCs:
-
Oligodendrocyte progenitor cells
- UCP2:
-
Uncoupling protein-2
- MAMs:
-
Mitochondria-associated ER membrane
- VDAC1:
-
Voltage-dependent anion channel 1
References
Murphy SJ, Werring DJ. Stroke: causes and clinical features. Medicine. 2020;48:561–6.
Elijovich L, Patel PV, Hemphill JC 3rd. Intracerebral hemorrhage. Semin Neurol. 2008;28:657–67.
Collaborators GBDS. Global, regional, and national burden of stroke and its risk factors, 1990–2019: a systematic analysis for the Global Burden of Disease Study 2019. Lancet Neurol. 2021;20:795–820.
Feigin VL, Brainin M, Norrving B, Martins S, Sacco RL, Hacke W, Fisher M, Pandian J, Lindsay P. World Stroke Organization (WSO): global stroke fact sheet 2022. Int J Stroke. 2022;17:18–29.
Qin C, Yang S, Chu YH, Zhang H, Pang XW, Chen L, Zhou LQ, Chen M, Tian DS, Wang W. Signaling pathways involved in ischemic stroke: molecular mechanisms and therapeutic interventions. Signal Transduct Target Ther. 2022;7:215.
Jayaraj RL, Azimullah S, Beiram R, Jalal FY, Rosenberg GA. Neuroinflammation: friend and foe for ischemic stroke. J Neuroinflamm. 2019;16:142.
Chen YY, Gong ZC, Zhang MM, Huang ZH. Brain-targeting emodin mitigates ischemic stroke via inhibiting AQP4-mediated swelling and neuroinflammation. Transl Stroke Res. 2023. https://doi.org/10.1007/s12975-023-01170-4.
Gao L, Peng L, Sherchan P, Tang H, Liu Y, Xiao J, Shi H, Luo Y, Tang J, Zhang JH, Xia Y. Inhibition of lysophosphatidic acid receptor 1 relieves PMN recruitment in CNS via LPA1/TSP1/CXCR2 pathway and alleviates disruption on blood–brain barrier following intracerebral haemorrhage in mice. Fluids Barriers CNS. 2023;20:33.
Dabrowska S, Andrzejewska A, Lukomska B, Janowski M. Neuroinflammation as a target for treatment of stroke using mesenchymal stem cells and extracellular vesicles. J Neuroinflamm. 2019;16:178.
Picca A, Guerra F, Calvani R, Romano R, Coelho-Junior HJ, Bucci C, Marzetti E. Mitochondrial dysfunction, protein misfolding and neuroinflammation in Parkinson’s disease: roads to biomarker discovery. Biomolecules. 2021;11:1508.
Wu X, Luo J, Liu H, Cui W, Guo K, Zhao L, Bai H, Guo W, Guo H, Feng D, Qu Y. Recombinant adiponectin peptide ameliorates brain injury following intracerebral hemorrhage by suppressing astrocyte-derived inflammation via the inhibition of Drp1-mediated mitochondrial fission. Transl Stroke Res. 2020;11:924–39.
Yang M, He Y, Deng S, Xiao L, Tian M, Xin Y, Lu C, Zhao F, Gong Y. Mitochondrial quality control: a pathophysiological mechanism and therapeutic target for stroke. Front Mol Neurosci. 2021;14: 786099.
Chen W, Guo C, Feng H, Chen Y. Mitochondria: novel mechanisms and therapeutic targets for secondary brain injury after intracerebral hemorrhage. Front Aging Neurosci. 2020;12: 615451.
de Oliveira LG, Angelo YS, Iglesias AH, Peron JPS. Unraveling the link between mitochondrial dynamics and neuroinflammation. Front Immunol. 2021;12: 624919.
Bader V, Winklhofer KF. Mitochondria at the interface between neurodegeneration and neuroinflammation. Semin Cell Dev Biol. 2020;99:163–71.
Lan X, Han X, Li Q, Yang QW, Wang J. Modulators of microglial activation and polarization after intracerebral haemorrhage. Nat Rev Neurol. 2017;13:420–33.
Nikolopoulos D, Manolakou T, Polissidis A, Filia A, Bertsias G, Koutmani Y, Boumpas DT. Microglia activation in the presence of intact blood–brain barrier and disruption of hippocampal neurogenesis via IL-6 and IL-18 mediate early diffuse neuropsychiatric lupus. Ann Rheum Dis. 2023;82:646–57.
Lawrence JM, Schardien K, Wigdahl B, Nonnemacher MR. Roles of neuropathology-associated reactive astrocytes: a systematic review. Acta Neuropathol Commun. 2023;11:42.
Liu P, Zhou Y, Shi J, Wang F, Yang X, Zheng X, Wang Y, He Y, Xie X, Pang X. Myricetin improves pathological changes in 3×Tg-AD mice by regulating the mitochondria-NLRP3 inflammasome-microglia channel by targeting P38 MAPK signaling pathway. Phytomedicine. 2023;115: 154801.
Yu CH, Davidson S, Harapas CR, Hilton JB, Mlodzianoski MJ, Laohamonthonkul P, Louis C, Low RRJ, Moecking J, De Nardo D, et al. TDP-43 triggers mitochondrial DNA release via mPTP to activate cGAS/STING in ALS. Cell. 2020;183:636-649.e618.
Haileselassie B, Joshi AU, Minhas PS, Mukherjee R, Andreasson KI, Mochly-Rosen D. Mitochondrial dysfunction mediated through dynamin-related protein 1 (Drp1) propagates impairment in blood brain barrier in septic encephalopathy. J Neuroinflamm. 2020;17:36.
Ibrahim AA, Abdel Mageed SS, Safar MM, El-Yamany MF, Oraby MA. MitoQ alleviates hippocampal damage after cerebral ischemia: the potential role of SIRT6 in regulating mitochondrial dysfunction and neuroinflammation. Life Sci. 2023;328: 121895.
Andrieux P, Chevillard C, Cunha-Neto E, Nunes JPS. Mitochondria as a cellular hub in infection and inflammation. Int J Mol Sci. 2021;22:11338.
Ribas de Pouplana L. The mitochondrial tRNA conundrum. Nat Rev Mol Cell Biol. 2020;21:361.
Tang JX, Thompson K, Taylor RW, Olahova M. Mitochondrial OXPHOS biogenesis: co-regulation of protein synthesis, import, and assembly pathways. Int J Mol Sci. 2020;21:3820.
Bausewein T, Mills DJ, Langer JD, Nitschke B, Nussberger S, Kuhlbrandt W. Cryo-EM structure of the TOM core complex from Neurospora crassa. Cell. 2017;170:693-700.e697.
Tucker K, Park E. Cryo-EM structure of the mitochondrial protein-import channel TOM complex at near-atomic resolution. Nat Struct Mol Biol. 2019;26:1158–66.
Araiso Y, Tsutsumi A, Qiu J, Imai K, Shiota T, Song J, Lindau C, Wenz LS, Sakaue H, Yunoki K, et al. Structure of the mitochondrial import gate reveals distinct preprotein paths. Nature. 2019;575:395–401.
Wang W, Chen X, Zhang L, Yi J, Ma Q, Yin J, Zhuo W, Gu J, Yang M. Atomic structure of human TOM core complex. Cell Discov. 2020;6:67.
Guan Z, Yan L, Wang Q, Qi L, Hong S, Gong Z, Yan C, Yin P. Structural insights into assembly of human mitochondrial translocase TOM complex. Cell Discov. 2021;7:22.
Palmer CS, Anderson AJ, Stojanovski D. Mitochondrial protein import dysfunction: mitochondrial disease, neurodegenerative disease and cancer. FEBS Lett. 2021;595:1107–31.
Fox TD. Mitochondrial protein synthesis, import, and assembly. Genetics. 2012;192:1203–34.
Burtscher J, Soltany A, Visavadiya NP, Burtscher M, Millet GP, Khoramipour K, Khamoui AV. Mitochondrial stress and mitokines in aging. Aging Cell. 2023;22: e13770.
Nour Eldine M, Alhousseini M, Nour-Eldine W, Noureldine H, Vakharia KV, Krafft PR, Noureldine MHA. The role of oxidative stress in the progression of secondary brain injury following germinal matrix hemorrhage. Transl Stroke Res. 2023. https://doi.org/10.1007/s12975-023-01147-3.
Thannickal VJ, Fanburg BL. Reactive oxygen species in cell signaling. Am J Physiol Lung Cell Mol Physiol. 2000;279:L1005-1028.
Angelova PR, Abramov AY. Functional role of mitochondrial reactive oxygen species in physiology. Free Radic Biol Med. 2016;100:81–5.
Rizwan H, Pal S, Sabnam S, Pal A. High glucose augments ROS generation regulates mitochondrial dysfunction and apoptosis via stress signalling cascades in keratinocytes. Life Sci. 2020;241: 117148.
Weindel CG, Martinez EL, Zhao X, Mabry CJ, Bell SL, Vail KJ, Coleman AK, VanPortfliet JJ, Zhao B, Wagner AR, et al. Mitochondrial ROS promotes susceptibility to infection via gasdermin D-mediated necroptosis. Cell. 2022;185:3214-3231.e3223.
Rius-Perez S, Torres-Cuevas I, Millan I, Ortega AL, Perez S. PGC-1alpha, inflammation, and oxidative stress: an integrative view in metabolism. Oxid Med Cell Longev. 2020;2020:1452696.
Zhang Y, Chen C, Jiang Y, Wang S, Wu X, Wang K. PPARgamma coactivator-1alpha (PGC-1alpha) protects neuroblastoma cells against amyloid-beta (Abeta) induced cell death and neuroinflammation via NF-kappaB pathway. BMC Neurosci. 2017;18:69.
Alvarez-Guardia D, Palomer X, Coll T, Davidson MM, Chan TO, Feldman AM, Laguna JC, Vazquez-Carrera M. The p65 subunit of NF-kappaB binds to PGC-1alpha, linking inflammation and metabolic disturbances in cardiac cells. Cardiovasc Res. 2010;87:449–58.
St-Pierre J, Drori S, Uldry M, Silvaggi JM, Rhee J, Jager S, Handschin C, Zheng K, Lin J, Yang W, et al. Suppression of reactive oxygen species and neurodegeneration by the PGC-1 transcriptional coactivators. Cell. 2006;127:397–408.
Zhang Q, Wang Z, Zhang W, Wen Q, Li X, Zhou J, Wu X, Guo Y, Liu Y, Wei C, et al. The memory of neuronal mitochondrial stress is inherited transgenerationally via elevated mitochondrial DNA levels. Nat Cell Biol. 2021;23:870–80.
Hamalainen RH, Landoni JC, Ahlqvist KJ, Goffart S, Ryytty S, Rahman MO, Brilhante V, Icay K, Hautaniemi S, Wang L, et al. Defects in mtDNA replication challenge nuclear genome stability through nucleotide depletion and provide a unifying mechanism for mouse progerias. Nat Metab. 2019;1:958–65.
Khan AUH, Allende-Vega N, Gitenay D, Garaude J, Vo DN, Belkhala S, Gerbal-Chaloin S, Gondeau C, Daujat-Chavanieu M, Delettre C, et al. Mitochondrial complex I activity signals antioxidant response through ERK5. Sci Rep. 2018;8:7420.
Desquiret V, Loiseau D, Jacques C, Douay O, Malthiery Y, Ritz P, Roussel D. Dinitrophenol-induced mitochondrial uncoupling in vivo triggers respiratory adaptation in HepG2 cells. Biochim Biophys Acta. 2006;1757:21–30.
Vaughan RA, Garcia-Smith R, Bisoffi M, Trujillo KA, Conn CA. Effects of caffeine on metabolism and mitochondria biogenesis in rhabdomyosarcoma cells compared with 2,4-dinitrophenol. Nutr Metab Insights. 2012;5:59–70.
Chen Q, Thompson J, Hu Y, Lesnefsky EJ. The mitochondrial electron transport chain contributes to calpain 1 activation during ischemia–reperfusion. Biochem Biophys Res Commun. 2022;613:127–32.
Purandare N, Somayajulu M, Huttemann M, Grossman LI, Aras S. The cellular stress proteins CHCHD10 and MNRR1 (CHCHD2): partners in mitochondrial and nuclear function and dysfunction. J Biol Chem. 2018;293:6517–29.
Liu YT, Huang X, Nguyen D, Shammas MK, Wu BP, Dombi E, Springer DA, Poulton J, Sekine S, Narendra DP. Loss of CHCHD2 and CHCHD10 activates OMA1 peptidase to disrupt mitochondrial cristae phenocopying patient mutations. Hum Mol Genet. 2020;29:1547–67.
Coyne LP, Chen XJ. mPOS is a novel mitochondrial trigger of cell death—implications for neurodegeneration. FEBS Lett. 2018;592:759–75.
Wang X, Chen XJ. A cytosolic network suppressing mitochondria-mediated proteostatic stress and cell death. Nature. 2015;524:481–4.
Liu Y, Wang X, Chen XJ. Misfolding of mutant adenine nucleotide translocase in yeast supports a novel mechanism of Ant1-induced muscle diseases. Mol Biol Cell. 2015;26:1985–94.
Liu Y, Wang X, Coyne LP, Yang Y, Qi Y, Middleton FA, Chen XJ. Mitochondrial carrier protein overloading and misfolding induce aggresomes and proteostatic adaptations in the cytosol. Mol Biol Cell. 2019;30:1272–84.
Anderson NS, Haynes CM. Folding the mitochondrial UPR into the integrated stress response. Trends Cell Biol. 2020;30:428–39.
Inigo JR, Chandra D. The mitochondrial unfolded protein response (UPRmt): shielding against toxicity to mitochondria in cancer. J Hematol Oncol. 2022;15:98.
Nargund AM, Pellegrino MW, Fiorese CJ, Baker BM, Haynes CM. Mitochondrial import efficiency of ATFS-1 regulates mitochondrial UPR activation. Science. 2012;337:587–90.
Xin N, Durieux J, Yang C, Wolff S, Kim HE, Dillin A. The UPRmt preserves mitochondrial import to extend lifespan. J Cell Biol. 2022;221: e202201071.
Katiyar A, Fujimoto M, Tan K, Kurashima A, Srivastava P, Okada M, Takii R, Nakai A. HSF1 is required for induction of mitochondrial chaperones during the mitochondrial unfolded protein response. FEBS Open Bio. 2020;10:1135–48.
Sutandy FXR, Gossner I, Tascher G, Munch C. A cytosolic surveillance mechanism activates the mitochondrial UPR. Nature. 2023;618(7966):849–54.
Zhang H, Shao S, Zeng Y, Wang X, Qin Y, Ren Q, Xiang S, Wang Y, Xiao J, Sun Y. Reversible phase separation of HSF1 is required for an acute transcriptional response during heat shock. Nat Cell Biol. 2022;24:340–52.
Joazeiro CAP. Mechanisms and functions of ribosome-associated protein quality control. Nat Rev Mol Cell Biol. 2019;20:368–83.
Martensson CU, Priesnitz C, Song J, Ellenrieder L, Doan KN, Boos F, Floerchinger A, Zufall N, Oeljeklaus S, Warscheid B, Becker T. Mitochondrial protein translocation-associated degradation. Nature. 2019;569:679–83.
Weidberg H, Amon A. MitoCPR—a surveillance pathway that protects mitochondria in response to protein import stress. Science. 2018;360: eaan4146.
Eckl EM, Ziegemann O, Krumwiede L, Fessler E, Jae LT. Sensing, signaling and surviving mitochondrial stress. Cell Mol Life Sci. 2021;78:5925–51.
Nwosu GO, Powell JA, Pitson SM. Targeting the integrated stress response in hematologic malignancies. Exp Hematol Oncol. 2022;11:94.
Cheng M, Yu H, Kong Q, Wang B, Shen L, Dong D, Sun L. The mitochondrial PHB2/OMA1/DELE1 pathway cooperates with endoplasmic reticulum stress to facilitate the response to chemotherapeutics in ovarian cancer. Int J Mol Sci. 2022;23:1320.
Guo X, Aviles G, Liu Y, Tian R, Unger BA, Lin YT, Wiita AP, Xu K, Correia MA, Kampmann M. Mitochondrial stress is relayed to the cytosol by an OMA1-DELE1-HRI pathway. Nature. 2020;579:427–32.
Guan R, Zou W, Dai X, Yu X, Liu H, Chen Q, Teng W. Mitophagy, a potential therapeutic target for stroke. J Biomed Sci. 2018;25:87.
Palikaras K, Tavernarakis N. Mitochondrial homeostasis: the interplay between mitophagy and mitochondrial biogenesis. Exp Gerontol. 2014;56:182–8.
Killackey SA, Bi Y, Soares F, Hammi I, Winsor NJ, Abdul-Sater AA, Philpott DJ, Arnoult D, Girardin SE. Mitochondrial protein import stress regulates the LC3 lipidation step of mitophagy through NLRX1 and RRBP1. Mol Cell. 2022;82:2815-2831.e2815.
Wang Y, Jasper H, Toan S, Muid D, Chang X, Zhou H. Mitophagy coordinates the mitochondrial unfolded protein response to attenuate inflammation-mediated myocardial injury. Redox Biol. 2021;45: 102049.
Lim Y, Berry B, Viteri S, McCall M, Park EC, Rongo C, Brookes PS, Nehrke K. FNDC-1-mediated mitophagy and ATFS-1 coordinate to protect against hypoxia-reoxygenation. Autophagy. 2021;17:3389–401.
Chen M, Chen Z, Wang Y, Tan Z, Zhu C, Li Y, Han Z, Chen L, Gao R, Liu L, Chen Q. Mitophagy receptor FUNDC1 regulates mitochondrial dynamics and mitophagy. Autophagy. 2016;12:689–702.
Berry BJ, Trewin AJ, Amitrano AM, Kim M, Wojtovich AP. Use the protonmotive force: mitochondrial uncoupling and reactive oxygen species. J Mol Biol. 2018;430:3873–91.
Chang SH, Jang J, Oh S, Yoon JH, Jo DG, Yun UJ, Park KW. Nrf2 induces Ucp1 expression in adipocytes in response to beta3-AR stimulation and enhances oxygen consumption in high-fat diet-fed obese mice. BMB Rep. 2021;54:419–24.
Oo SM, Oo HK, Takayama H, Ishii KA, Takeshita Y, Goto H, Nakano Y, Kohno S, Takahashi C, Nakamura H, et al. Selenoprotein P-mediated reductive stress impairs cold-induced thermogenesis in brown fat. Cell Rep. 2022;38: 110566.
Wang P, Geng J, Gao J, Zhao H, Li J, Shi Y, Yang B, Xiao C, Linghu Y, Sun X, et al. Macrophage achieves self-protection against oxidative stress-induced ageing through the Mst-Nrf2 axis. Nat Commun. 2019;10:755.
Liu C, Zhang A. ROS-mediated PERK-eIF2alpha-ATF4 pathway plays an important role in arsenite-induced L-02 cells apoptosis via regulating CHOP-DR5 signaling. Environ Toxicol. 2020;35:1100–13.
Hu D, Liu Z, Qi X. UPR(mt) activation protects against MPP(+)-induced toxicity in a cell culture model of Parkinson’s disease. Biochem Biophys Res Commun. 2021;569:17–22.
Zhang Y, Xi X, Mei Y, Zhao X, Zhou L, Ma M, Liu S, Zha X, Yang Y. High-glucose induces retinal pigment epithelium mitochondrial pathways of apoptosis and inhibits mitophagy by regulating ROS/PINK1/Parkin signal pathway. Biomed Pharmacother. 2019;111:1315–25.
Xiao B, Goh JY, Xiao L, Xian H, Lim KL, Liou YC. Reactive oxygen species trigger Parkin/PINK1 pathway-dependent mitophagy by inducing mitochondrial recruitment of Parkin. J Biol Chem. 2017;292:16697–708.
Lehtonen JM, Auranen M, Darin N, Sofou K, Bindoff L, Hikmat O, Uusimaa J, Vieira P, Tulinius M, Lonnqvist T, et al. Diagnostic value of serum biomarkers FGF21 and GDF15 compared to muscle sample in mitochondrial disease. J Inherit Metab Dis. 2021;44:469–80.
Li Y, Li S, Qiu Y, Zhou M, Chen M, Hu Y, Hong S, Jiang L, Guo Y. Circulating FGF21 and GDF15 as biomarkers for screening, diagnosis, and severity assessment of primary mitochondrial disorders in children. Front Pediatr. 2022;10: 851534.
Keipert S, Ost M, Johann K, Imber F, Jastroch M, van Schothorst EM, Keijer J, Klaus S. Skeletal muscle mitochondrial uncoupling drives endocrine cross-talk through the induction of FGF21 as a myokine. Am J Physiol Endocrinol Metab. 2014;306:E469-482.
Fu T, Xu Z, Liu L, Guo Q, Wu H, Liang X, Zhou D, Xiao L, Liu L, Liu Y, et al. Mitophagy directs muscle-adipose crosstalk to alleviate dietary obesity. Cell Rep. 2018;23:1357–72.
Jin L, Geng L, Ying L, Shu L, Ye K, Yang R, Liu Y, Wang Y, Cai Y, Jiang X, et al. FGF21-sirtuin 3 axis confers the protective effects of exercise against diabetic cardiomyopathy by governing mitochondrial integrity. Circulation. 2022;146:1537–57.
Li X, Hong Y, He H, Jiang G, You W, Liang X, Fu Q, Han S, Lian Q, Zhang Y. FGF21 mediates mesenchymal stem cell senescence via regulation of mitochondrial dynamics. Oxid Med Cell Longev. 2019;2019:4915149.
Lee S, Choi J, Mohanty J, Sousa LP, Tome F, Pardon E, Steyaert J, Lemmon MA, Lax I, Schlessinger J. Structures of beta-klotho reveal a ‘zip code’-like mechanism for endocrine FGF signalling. Nature. 2018;553:501–5.
Yu H, Xia F, Lam KS, Wang Y, Bao Y, Zhang J, Gu Y, Zhou P, Lu J, Jia W, Xu A. Circadian rhythm of circulating fibroblast growth factor 21 is related to diurnal changes in fatty acids in humans. Clin Chem. 2011;57:691–700.
Chung HK, Kim JT, Kim HW, Kwon M, Kim SY, Shong M, Kim KS, Yi HS. GDF15 deficiency exacerbates chronic alcohol- and carbon tetrachloride-induced liver injury. Sci Rep. 2017;7:17238.
Ost M, Igual Gil C, Coleman V, Keipert S, Efstathiou S, Vidic V, Weyers M, Klaus S. Muscle-derived GDF15 drives diurnal anorexia and systemic metabolic remodeling during mitochondrial stress. EMBO Rep. 2020;21: e48804.
Conte M, Martucci M, Mosconi G, Chiariello A, Cappuccilli M, Totti V, Santoro A, Franceschi C, Salvioli S. GDF15 plasma level is inversely associated with level of physical activity and correlates with markers of inflammation and muscle weakness. Front Immunol. 2020;11:915.
Gao L, Shi H, Sherchan P, Tang H, Peng L, Xie S, Liu R, Hu X, Tang J, Xia Y, Zhang JH. Inhibition of lysophosphatidic acid receptor 1 attenuates neuroinflammation via PGE2/EP2/NOX2 signalling and improves the outcome of intracerebral haemorrhage in mice. Brain Behav Immun. 2021;91:615–26.
Maida CD, Norrito RL, Daidone M, Tuttolomondo A, Pinto A. Neuroinflammatory mechanisms in ischemic stroke: focus on cardioembolic stroke, background, and therapeutic approaches. Int J Mol Sci. 2020;21:6454.
Zhu J, Lee MJ, Chang HJ, Ju X, Cui J, Lee YL, Go D, Chung W, Oh E, Heo JY. Reactive microglia and mitochondrial unfolded protein response following ventriculomegaly and behavior defects in kaolin-induced hydrocephalus. BMB Rep. 2022;55:181–6.
Zhu J, Lee MJ, An JH, Oh E, Chung W, Heo JY. ATF5 attenuates the secretion of pro-inflammatory cytokines in activated microglia. Int J Mol Sci. 2023;24:3322.
Sorrentino V, Menzies KJ, Auwerx J. Repairing mitochondrial dysfunction in disease. Annu Rev Pharmacol Toxicol. 2018;58:353–89.
Xiaowei X, Qian X, Dingzhou Z. Sirtuin-3 activates the mitochondrial unfolded protein response and reduces cerebral ischemia/reperfusion injury. Int J Biol Sci. 2023;19:4327–39.
Ma C, Gao B, Wang Z, You W, Yu Z, Shen H, Li X, Li H, Zhang X, Wang Z, Chen G. GrpEL1 regulates mitochondrial unfolded protein response after experimental subarachnoid hemorrhage in vivo and in vitro. Brain Res Bull. 2022;181:97–108.
An H, Zhou B, Ji X. Mitochondrial quality control in acute ischemic stroke. J Cereb Blood Flow Metab. 2021;41:3157–70.
He M, Kittur FS, Hung CY, Zhang J, Jing L, Sane DC, Li PA, Xie J. A novel plant-produced Asialo-rhuEPO protects brain from ischemic damage without erythropoietic action. Transl Stroke Res. 2022;13:338–54.
Busceti CL, Cotugno M, Bianchi F, Forte M, Stanzione R, Marchitti S, Battaglia G, Nicoletti F, Fornai F, Rubattu S. Brain overexpression of uncoupling protein-2 (UCP2) delays renal damage and stroke occurrence in stroke-prone spontaneously hypertensive rats. Int J Mol Sci. 2020;21:4289.
Zorov DB, Vorobjev IA, Popkov VA, Babenko VA, Zorova LD, Pevzner IB, Silachev DN, Zorov SD, Andrianova NV, Plotnikov EY. Lessons from the discovery of mitochondrial fragmentation (fission): a review and update. Cells. 2019;8:175.
Jiang RQ, Li QQ, Sheng R. Mitochondria associated ER membranes and cerebral ischemia: molecular mechanisms and therapeutic strategies. Pharmacol Res. 2023;191: 106761.
Hu H, Guo L, Overholser J, Wang X. Mitochondrial VDAC1: a potential therapeutic target of inflammation-related diseases and clinical opportunities. Cells. 2022;11:3174.
Yao Y, Bade R, Li G, Zhang A, Zhao H, Fan L, Zhu R, Yuan J. Global-scale profiling of differential expressed lysine-lactylated proteins in the cerebral endothelium of cerebral ischemia–reperfusion injury rats. Cell Mol Neurobiol. 2023;43:1989–2004.
Liu R, Xu C, Zhang W, Cao Y, Ye J, Li B, Jia S, Weng L, Liu Y, Liu L, Zheng M. FUNDC1-mediated mitophagy and HIF1alpha activation drives pulmonary hypertension during hypoxia. Cell Death Dis. 2022;13:634.
Cai Y, Yang E, Yao X, Zhang X, Wang Q, Wang Y, Liu J, Fan W, Yi K, Kang C, Wu J. FUNDC1-dependent mitophagy induced by tPA protects neurons against cerebral ischemia–reperfusion injury. Redox Biol. 2021;38: 101792.
Zhang T, Wu P, Budbazar E, Zhu Q, Sun C, Mo J, Peng J, Gospodarev V, Tang J, Shi H, Zhang JH. Mitophagy reduces oxidative stress via Keap1 (Kelch-like epichlorohydrin-associated protein 1)/Nrf2 (nuclear factor-E2-related factor 2)/PHB2 (prohibitin 2) pathway after subarachnoid hemorrhage in rats. Stroke. 2019;50:978–88.
He Q, Li Z, Meng C, Wu J, Zhao Y, Zhao J. Parkin-dependent mitophagy is required for the inhibition of ATF4 on NLRP3 inflammasome activation in cerebral ischemia–reperfusion injury in rats. Cells. 2019;8:897.
Zhang J, Chen H, Huang W, Zhou C, Li J, Xing S, Chen L, Li C, Dang C, Liu G, et al. Unfolded protein response is activated in the ipsilateral thalamus following focal cerebral infarction in hypertensive rats. Clin Exp Pharmacol Physiol. 2016;43:1216–24.
Gu Y, Ren K, Wang L, Jiang C, Yao Q. Rg1 in combination with mannitol protects neurons against glutamate-induced ER stress via the PERK-eIF2 alpha-ATF4 signaling pathway. Life Sci. 2020;263: 118559.
Ma D, Qiao J, Qu Q, He F, Chen W, Yu B. Weighted gene co-expression network analysis to investigate the key genes implicated in global brain ischemia/reperfusion injury in rats. Adv Clin Exp Med. 2020;29:649–59.
Sun X, Crawford R, Liu C, Luo T, Hu B. Development-dependent regulation of molecular chaperones after hypoxia-ischemia. Neurobiol Dis. 2015;82:123–31.
Li L, Li Y, He B, Li H, Ji H, Wang Y, Zhu Z, Hu Y, Zhou Y, Yang T, et al. HSF1 is involved in suppressing A1 phenotype conversion of astrocytes following spinal cord injury in rats. J Neuroinflamm. 2021;18:205.
Liao Y, Gu Y, Wang J, Tian Y, Ni X, Zhou L, Ye Y, Xia G. HSF1 inhibits microglia activation to attenuate neuroinflammation via regulating miR-214-3p and NFATc2 in Parkinson’s disease. Folia Neuropathol. 2023;61:53–67.
Cao J, Dong L, Luo J, Zeng F, Hong Z, Liu Y, Zhao Y, Xia Z, Zuo D, Xu L, Tao T. Supplemental N-3 polyunsaturated fatty acids limit A1-specific astrocyte polarization via attenuating mitochondrial dysfunction in ischemic stroke in mice. Oxid Med Cell Longev. 2021;2021:5524705.
Shen Y, Wu Q, Shi J, Zhou S. Regulation of SIRT3 on mitochondrial functions and oxidative stress in Parkinson’s disease. Biomed Pharmacother. 2020;132: 110928.
Sun B, Yang S, Li S, Hang C. Melatonin upregulates nuclear factor erythroid-2 related factor 2 (Nrf2) and mediates mitophagy to protect against early brain injury after subarachnoid hemorrhage. Med Sci Monit. 2018;24:6422–30.
Cao S, Shrestha S, Li J, Yu X, Chen J, Yan F, Ying G, Gu C, Wang L, Chen G. Melatonin-mediated mitophagy protects against early brain injury after subarachnoid hemorrhage through inhibition of NLRP3 inflammasome activation. Sci Rep. 2017;7:2417.
Peng J, Zuo Y, Huang L, Okada T, Liu S, Zuo G, Zhang G, Tang J, Xia Y, Zhang JH. Activation of GPR30 with G1 attenuates neuronal apoptosis via src/EGFR/stat3 signaling pathway after subarachnoid hemorrhage in male rats. Exp Neurol. 2019;320: 113008.
Nguyen TN, Sawa-Makarska J, Khuu G, Lam WK, Adriaenssens E, Fracchiolla D, Shoebridge S, Bernklau D, Padman BS, Skulsuppaisarn M, et al. Unconventional initiation of PINK1/Parkin mitophagy by optineurin. Mol Cell. 2023;83:1693-1709.e1699.
Cheng Y, Liu M, Tang H, Chen B, Yang G, Zhao W, Cai Y, Shang H. iTRAQ-based quantitative proteomics indicated Nrf2/OPTN-mediated mitophagy inhibits NLRP3 inflammasome activation after intracerebral hemorrhage. Oxid Med Cell Longev. 2021;2021:6630281.
Li J, Lu J, Mi Y, Shi Z, Chen C, Riley J, Zhou C. Voltage-dependent anion channels (VDACs) promote mitophagy to protect neuron from death in an early brain injury following a subarachnoid hemorrhage in rats. Brain Res. 2014;1573:74–83.
Wang WJ, Wang Y, Chen HZ, Xing YZ, Li FW, Zhang Q, Zhou B, Zhang HK, Zhang J, Bian XL, et al. Orphan nuclear receptor TR3 acts in autophagic cell death via mitochondrial signaling pathway. Nat Chem Biol. 2014;10:133–40.
Hoshino A, Wang WJ, Wada S, McDermott-Roe C, Evans CS, Gosis B, Morley MP, Rathi KS, Li J, Li K, et al. The ADP/ATP translocase drives mitophagy independent of nucleotide exchange. Nature. 2019;575:375–9.
Mishra Y, Kaundal RK. Role of SIRT3 in mitochondrial biology and its therapeutic implications in neurodegenerative disorders. Drug Discov Today. 2023;28: 103583.
Zhou Z, Fan Y, Zong R, Tan K. The mitochondrial unfolded protein response: a multitasking giant in the fight against human diseases. Ageing Res Rev. 2022;81: 101702.
Wu Y, Hu Q, Wu X, Cai YN, Zhang YZ, Wu YX, Zhu G, Luo JN, Cheng HB, Yu JG, et al. P7C3-A20 attenuates microglial inflammation and brain injury after ICH through activating the NAD(+)/Sirt3 pathway. Oxid Med Cell Longev. 2023;2023:7857760.
Chen K, Huang W, Wang J, Xu H, Ruan L, Li Y, Wang Z, Wang X, Lin L, Li X. Increased serum fibroblast growth factor 21 levels are associated with adverse clinical outcomes after intracerebral hemorrhage. Front Neurosci. 2023;17:1117057.
Song L, Soderholm M, Svensson EH, Borne Y, Engstrom G. Circulating growth differentiation factor 15 levels are associated with risk of both intracerebral and subarachnoid hemorrhage. Front Neurol. 2021;12: 664010.
Lockhart SM, Saudek V, O’Rahilly S. GDF15: a hormone conveying somatic distress to the brain. Endocr Rev. 2020;41: bnaa007.
Chen T, Wang Y, Wang YH, Hang CH. The Mfn1-βIIPKC interaction regulates mitochondrial dysfunction via Sirt3 following experimental subarachnoid hemorrhage. Transl Stroke Res. 2022;13:845–57.
Lange PS, Chavez JC, Pinto JT, Coppola G, Sun CW, Townes TM, Geschwind DH, Ratan RR. ATF4 is an oxidative stress-inducible, prodeath transcription factor in neurons in vitro and in vivo. J Exp Med. 2008;205:1227–42.
Wu G, Zhang X, Li S, Wang L, Bai J, Wang H, Shu Q. Silencing ATF4 inhibits JMJD3-dependent JUNB/ETS1 axis and mitigates cerebral ischemic injury. J Biochem Mol Toxicol. 2022;36: e23070.
He Y, Xiao Y, Yang X, Li Y, Wang B, Yao F, Shang C, Jin Z, Wang W, Lin R. SIRT6 inhibits TNF-α-induced inflammation of vascular adventitial fibroblasts through ROS and Akt signaling pathway. Exp Cell Res. 2017;357:88–97.
Ren JD, Wu XB, Jiang R, Hao DP, Liu Y. Molecular hydrogen inhibits lipopolysaccharide-triggered NLRP3 inflammasome activation in macrophages by targeting the mitochondrial reactive oxygen species. Biochim Biophys Acta. 2016;1863:50–5.
Shi M, Chen J, Liu T, Dai W, Zhou Z, Chen L, Xie Y. Protective effects of remimazolam on cerebral ischemia/reperfusion injury in rats by inhibiting of NLRP3 inflammasome-dependent pyroptosis. Drug Des Devel Ther. 2022;16:413–23.
Shi L, Liang F, Zheng J, Zhou K, Chen S, Yu J, Zhang J. Melatonin regulates apoptosis and autophagy via ROS-MST1 pathway in subarachnoid hemorrhage. Front Mol Neurosci. 2018;11:93.
Hu Q, Liang X, Chen D, Chen Y, Doycheva D, Tang J, Tang J, Zhang JH. Delayed hyperbaric oxygen therapy promotes neurogenesis through reactive oxygen species/hypoxia-inducible factor-1α/β-catenin pathway in middle cerebral artery occlusion rats. Stroke. 2014;45:1807–14.
Wang J, Chen A, Xue Z, Liu J, He Y, Liu G, Zhao Z, Li W, Zhang Q, Chen A, et al. BCL2L13 promotes mitophagy through DNM1L-mediated mitochondrial fission in glioblastoma. Cell Death Dis. 2023;14:585.
Banerjee S, Biehl A, Gadina M, Hasni S, Schwartz DM. JAK-STAT signaling as a target for inflammatory and autoimmune diseases: current and future prospects. Drugs. 2017;77:521–46.
Hu X, Li J, Fu M, Zhao X, Wang W. The JAK/STAT signaling pathway: from bench to clinic. Signal Transduct Target Ther. 2021;6:402.
Philips RL, Wang Y, Cheon H, Kanno Y, Gadina M, Sartorelli V, Horvath CM, Darnell JE Jr, Stark GR, O’Shea JJ. The JAK-STAT pathway at 30: much learned, much more to do. Cell. 2022;185:3857–76.
Zhou K, Chen J, Wu J, Wu Q, Jia C, Xu YXZ, Chen L, Tu W, Yang G, Kong J, et al. Atractylenolide III ameliorates cerebral ischemic injury and neuroinflammation associated with inhibiting JAK2/STAT3/Drp1-dependent mitochondrial fission in microglia. Phytomedicine. 2019;59: 152922.
Gao L, Liu F, Hou PP, Manaenko A, Xiao ZP, Wang F, Xu TL, Hu Q. Neurons release injured mitochondria as “help-me” signaling after ischemic stroke. Front Aging Neurosci. 2022;14: 785761.
Patabendige A, Singh A, Jenkins S, Sen J, Chen R. Astrocyte activation in neurovascular damage and repair following ischaemic stroke. Int J Mol Sci. 2021;22:4280.
Meyer N, Rinholm JE. Mitochondria in myelinating oligodendrocytes: slow and out of breath? Metabolites. 2021;11:359.
Nakamura DS, Lin YH, Khan D, Gothie JM, de Faria O Jr, Dixon JA, McBride HM, Antel JP, Kennedy TE. Mitochondrial dynamics and bioenergetics regulated by netrin-1 in oligodendrocytes. Glia. 2021;69:392–412.
Yan H, Zhang D, Hao S, Li K, Hang CH. Role of mitochondrial calcium uniporter in early brain injury after experimental subarachnoid hemorrhage. Mol Neurobiol. 2015;52:1637–47.
Yang M, Deng S, Jiang J, Tian M, Xiao L, Gong Y. Oxytocin improves intracerebral hemorrhage outcomes by suppressing neuronal pyroptosis and mitochondrial fission. Stroke. 2023;54:1888–900.
Chen S, Zuo Y, Huang L, Sherchan P, Zhang J, Yu Z, Peng J, Zhang J, Zhao L, Doycheva D, et al. The MC(4) receptor agonist RO27-3225 inhibits NLRP1-dependent neuronal pyroptosis via the ASK1/JNK/p38 MAPK pathway in a mouse model of intracerebral haemorrhage. Br J Pharmacol. 2019;176:1341–56.
Karuppagounder SS, Alim I, Khim SJ, Bourassa MW, Sleiman SF, John R, Thinnes CC, Yeh TL, Demetriades M, Neitemeier S, et al. Therapeutic targeting of oxygen-sensing prolyl hydroxylases abrogates ATF4-dependent neuronal death and improves outcomes after brain hemorrhage in several rodent models. Sci Transl Med. 2016;8:328ra329.
Liu T, Li X, Cui Y, Meng P, Zeng G, Wang Y, Wang Q. Bioinformatics analysis identifies potential ferroptosis key genes in the pathogenesis of intracerebral hemorrhage. Front Neurosci. 2021;15: 661663.
Wei M, Li C, Yan Z, Hu Z, Dong L, Zhang J, Wang X, Li Y, Zhang H. Activated microglia exosomes mediated miR-383-3p promotes neuronal necroptosis through inhibiting ATF4 expression in intracerebral hemorrhage. Neurochem Res. 2021;46:1337–49.
Lu Z, Wang Z, Yu L, Ding Y, Xu Y, Xu N, Li R, Tang J, Chen G, Zhang JH. GCN2 reduces inflammation by p-eIF2alpha/ATF4 pathway after intracerebral hemorrhage in mice. Exp Neurol. 2019;313:16–25.
Lin YF, Schulz AM, Pellegrino MW, Lu Y, Shaham S, Haynes CM. Maintenance and propagation of a deleterious mitochondrial genome by the mitochondrial unfolded protein response. Nature. 2016;533:416–9.
Funding
This work was supported by a grant from Hainan Provincial Natural Science Foundation of China (No. 822MS205), to Dr. Ling Gao, a grant from Hainan Provincial Natural Science Foundation of China (No. 821QN1005) to Dr. Li Peng, a grant from Hainan Province Clinical Medical research Center (No. LCYX202107) to Dr. Ying Xia, a grant from Key research and development program of Hainan province (No. ZDYF2021GXJS017) and Hainan Province Clinical Medical Center.
Author information
Authors and Affiliations
Contributions
LG and LP drafted the manuscript and drew the figures. JW worked on manuscript editing. YX and JHZ worked on manuscript conception and revision. All authors contributed to the manuscript in conceptualization and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable as no patients/participants were involved in this review.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Gao, L., Peng, L., Wang, J. et al. Mitochondrial stress: a key role of neuroinflammation in stroke. J Neuroinflammation 21, 44 (2024). https://doi.org/10.1186/s12974-024-03033-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12974-024-03033-7