
Abstract
Background
Telocytes play key roles in maintenance of organ/tissue function and prevention of organ injury. However, there are great challenges to investigate telocytes functions using primary telocytes, due to the difficulties of isolation, identification, and stability. The present study aims at constructing continuous cell strain of mouse lung telocyte cell line with stable characters by gene modification and investigating biological behaviors and responses of gene-modified telocytes to inflammation.
Methods
Mouse primary lung telocytes were isolated and identified using immune-labeling markers and immunoelectron microscopy. Primary telocytes were transformed with Simian vacuolating virus 40 small and large T antigen (SV40). Biological characters, behaviors morphology, and proliferation of those gene-modified telocytes were defined and monitored dynamically for 50 generations, as compared with primary lung telocytes. Cell cycle of mouse primary lung telocytes or gene-modified telocytes was detected by flow cytometry.
Results
Gene modified telocytes of generations 5, 10, 30 and 50 were observed with telopodes and also showed CD34 and ckit positive. Multiple cellular morphology were also observed on telocyte cell-line under monitor of celliq and enhanced cell proliferation were showed. SV40 transduction was also reduced apoptosis and increased the ratio of S and G2 phases in telocyte cell-line.
Conclusion
We successfully constructed mouse lung telocyte cell-line which maintained the biological properties and behaviors as primary telocytes and could responses to inflammation induced by LPS. Thus, gene-modified lung telocytes, Telocyte Line, would provide a cell tool for researchers exploring the roles and applications of telocytes involved in physiological and pathological states in future.
Similar content being viewed by others


Background
Telocytes (TCs) have been found widely spreaded in a large number of organs and tissues of mammals, including atrial and ventricular myocardium, bladder, lung skeletal muscle, gastrointestinal tract, eyes, and others. TCs communicate with neighboring cells by homo- and heterocellular contacts and transfer genetic information and signaling molecules to influence other cells [1, 2]. Shoshkes-Carme et al. [3] recently demonstrated that TCs can be the major source of Wnt signaling, and play dependent roles in proliferation of intestinal stem cells and epithelial renewal. This particular study provides solid evidence that forkhead box L1-positive TCs contribute to the formation of the subepithelial plexus of the intestine, support the entire epithelium, and provide niche signals to intestinal stem cells, by producing Wnt ligands and maintaining Wnt signal pathway. Although TCs are often allocated within the stem cell niche, pulmonary TCs have different genetic profiles and biological functions from stem cells, fibroblasts, alveolar type II cells, airway basal cells, proximal airway cells, and T cells from bronchial lymph nodes or from lungs, respectively [4,5,6,7]. TCs functionally and constructively support and connect tissue cells, function as signal massage carriers, and benefit gene and cell therapies [8,9,10,11,12].
Gene modification is considered as a potential therapy to genetically prevent and treat diseases, even though a large number of factors still need to furthermore clarified, e.g. safety, efficacy, and complexity. With rapid development of gene editing technology, the precision and efficacy of gene modification are improved significantly [13, 14]. Safety profiles of gene modification as part of limitations require more preclinical and clinical evidence. The present study investigates potential effects of gene modification on morphological phenomes, biological behaviors and functions, as well as responses to inflammation in lung TCs. An anti-aging gene from Simian vacuolating virus 40 was transferred as the target gene into primary TCs isolated from mouse lungs to improve the longevity of cells, since primary TCs hardly survive in the in vitro system for a few weeks or months [5,6,7].
Materials and methods
TCs isolation and primary cell culture
Mice were provided by Animal Facility in Biomedical Research Center of Zhongshan Hospital, Fudan University. This study was approved by the Fudan University Ethical Committee for animal experiments.
Mouse lung TCs were isolated and prepared as escribed previously [4]. In brief, female BABL/c mice aged 6–8 weeks were used and anaesthetized. The trachea and lung tissues were isolated and collected into sterile tubes, containing Dulbecco’s Modified Eagle’s Medium plus F12 (DMEM/F12) integration (Gibco, NY, USA), supplemented with 100 UI/ml penicillin, 0.1 mg/ml streptomycin (Sigma-Aldrich Shanghai Trading Co Ltd., Shanghai, China). Samples were placed on ice and transported to the cell culture laboratory within 30 min. The tissues were cut into about 1 × mm3 in sterile DMEM/F12 and incubated in 10 mg/ml collagenase type II (Sigma-Aldrich, St. Louis, MO, USA) and 2000 U/ml deoxyribonuclease I (Sigma-Aldrich) on an orbital shaker for 4 h, at 37 °C. Cell suspension was separated by 40-m-diameter cell strainer (BD Falcon, NJ, USA), and dispersed cells were collected by centrifugation. Cells were resuspended in DMEM/F12, supplemented with 10% fetal calf serum, 100 UI/ml penicillin, 0.1 mg/ml streptomycin (Sigma-Aldrich), and cultured in a incubator, with 5% CO2 in air, at 37 °C, for 30 min, to wipe off most fibroblasts. Cell suspension was transferred to other flakes and cultured for another 12 h before changing culture medium. Culture medium was changed every 48 h. Cells were examined by phase contrast microscope, under an inverted Olympus phase contrast microscope (1 × 51; Tokyo, Japan).
Lentivirus construction and infection
Lentivirus particles containing the small and large T antigen of anti-aging gene from Simian vacuolating virus 40 (SV40) gene were constructed (HanyinCo., Shanghai, China) according to the former articles [15]. The oligo (listed 5′–3′) SV40-F: CCG GAATTCATGGATAAAGTTTTAAACAGAGAGGAATC and SV40-R: GCTCTAGATTACAAGTCCTCTTCAGAAATGAG. TCs were infected on 6-well plates for 12 h at 37 °C, replaced fresh medium, and harvested for qPCR analysis to determine the SV40 mRNA expression level after being cultured for 96 h at 37 °C.
RNA extraction and PCR
Primary TCs and TCs transformed with SV40 (TCsSV40) were seeded in 24-well plates with a density of 104 cells/well and cultured for 24 h at 37 °C. Cells were washed thrice with PBS, and total RNA was isolated and transcribed into single-stranded cDNA using the 1st Strand cDNA Synthesis Kit for RT-PCR (AMV, Roche) following the recommendations of the manufacturer.
cDNA was synthesized from 1 µg of total RNA using PrimeScript® RT reagent Kit (Takara Bio Inc., Shiga, Japan). PCR was performed with 1 µl of cDNA using GoTaq polymerase (Promega) for 25 cycles with specific primers for SV40. PCR reaction products were resolved and stained with Gelred (Biotium Inc., Newark, USA).
Detection of cell bio-behaviors
The bio-behaviors of TCs and TCsSV40 were recorded and analyzed using a Cell-IQ cell culturing platform (Chip-Man Technologies, Tampere, Finland), equipped with a phase-contrast microscope (Nikon CFI Achromat phase contrast objective with 10 magnification) and a camera20. The bio-behaviors, including cell proliferation, division, death, cell morphology, and cell movement, can be monitored and recorded as time-lapse data by this Cell-IQ system uses machine vision technology. Images were captured at about 30 min intervals for 48 h. Analysis was carried out with a freely distributed Image software (McMaster Biophotonics Facility, Hamilton, ON), using the Manual Tracking plugin created by Fabrice Cordelie´res (Institute Curie, Orsay, France).
RNA microarrays and long non-coding RNA (lncRNA) classification pipeline
RNA microarrays and lncRNA classification pipeline were tested in primary TCs or TCsSV40. Briefly, total RNA was collected using NucleoSpin® RNA Plus according the manufacturer’s protocol (Macherey–Nagel, Inc., Düren, Germany). Microarray and quality controls of gene expression profiling were performed after RNA and cDNA amplifications, using the GeneChip® Human Transcriptome Array 2.0 gene chip (Affymetrix, Inc., UK) with 67,528 genes. Gene expression data from each group were analyzed using Expression Console and Transcriptome Analysis Console 3.0.0.466 (Affymetrix). The differentially expressed mRNA and lncRNAs were used for a hierarchical clustering analysis (HCA) in Cluster and TreeView (https://sourceforge.net/projects/jtreeview/files/).
Immunofluorescent staining
Double immunofluorescent staining for CD34 and vimentin was performed as previously reported [21]. In brief, primary TCs or TCsSV40 in 1, 5, 10, 30, or 50 generations were load and cultured on glass bottom cell culture dishes with 20 mm diameter glass (NEST, Nanjing, China) and were fixed in 4% paraformaldehyde containing 0.05% Triton-X-100 for 20 min. The cells were washed thrice with PBS and blocked in 5% bovine serum albumin (BSA) for 1 h and incubated overnight at 4 °C with mouse anti-CD34 antibody and goat anti-vimentin antibody or rabbit anti-ckit antibody (1:200 dilution; Abcam, Cambridge, UK) diluted in 1% BSA in PBS. Cells were washed in PBS thrice and incubated with PE conjugated anti-goat secondary antibodies and FITC conjugated anti-rabbit secondary antibodies and/or FITC conjugated anti-mouse secondary antibodies (1:200 dilution; Jackson ImmunoResearch, USA). The nuclear were marked by DAPI staining, according to the manufacture’s instruction (KeyGEN BioTECH, Nanjing, China).
Transmission electron microscopy
The ultrastructure of cells were observed under transmission electron microscopy (TEM) as previously reported (14). In brief, primary TCs or TCsSV40 in 1, 5, 10, 30, and 50 generations were cultured, collected, and fixed in 4% glutaraldehyde (pH 7.3, 4 °C) for 4 h. Cells were then washed with 0.1 M cacodylate buffer and post-fixed with 1% osmium tetroxide in 0.1 M cacodylate buffer (pH 7.3, 4 °C). After fixing, cells were dehydrated in a graded series of ethanol, impregnated in propylene oxide (immersed overnight in a mixture of propylene oxide and Epon 812 resin), and embedded in Epon 812. Ultrathin sections at 70 nm were cut on a Leica LKB-II (Nußloch, Germany), collected on Formvar-coated copper grids, stained with uranyl acetate and lead citrate, and observed at an acceleration voltage of 80 kV electron microscope (JEOL JEM-1230, Tokyo, Japan).
Immunoelectron microscopy
Ultrathin sections were prepared and collected on nickel grids. Immunolabeling staining for CD34/Vimentin and ckit/platelet-derived growth factor receptor α (PDGFR-α) was used as previously reported [22]. In brief, sections were incubated in 50 mM Glycine for 30 min and washed in Ultra-pure Water thrice for 5 min. Sections were etched in 1% sodium periodate for 10 min following washing in Ultra-pure water. Sections were incubated in the blocking buffer for 20 min and labeled with rabbit anti-ckit antibody, mouse anti-CD34 antibody, goat anti-vimentin antibody and/or rat anti PDGFR-α antibody (1:200 dilution; Abcam) at 4 °C for 24 h. The nickel grids were washed in PBS for 5 min 12 times, blocked within 1% BSA for 20 min, and incubated with 10 nm gold conjugated anti-goat secondary antibodies, 18 nm gold conjugated anti-mouse secondary antibodies, 25 nm gold conjugated anti-rat secondary antibodies, and/or 40 nm gold conjugated anti-rabbit secondary antibodies (1:200 dilution; Abcam) for 2 h. Nickel grids were dried on filter paper and observed with transmission electronic microscopy (TEM). The staining controls included cells stained only with the second antibodies with gold labelling or the first antibodies.
Cell cycle assay
Propidium iodide (PI) staining was used for cell cycle analysis of primary TCs and TCsSV40 as described in manufacturer. In brief, cells were collected and fixed in 75% ethanol at 4 °C for overnight. After centrifuging and washing, staining buffer (BD Pharmingen, NJ, USA) with 0.5 ml PI/RNase was added to each tube for 15 min at room temperature. Samples were examined with a fluorescence-activated cell sorting flow cytometer (FACS Aria II, Becton, Dickinson and Company, NJ, USA) and DNA histograms were analyzed with Flowjo 7.6.1 software. Each test was repeated thrice.
Statistics
Data were expressed as mean ± SEM analyzed using SPSS Statistics 20 (IBM, Chicago, USA). Statistical differences between two groups were compared by t-test. Statistical differences among more than two groups were determined using ANOVA. p value less than 0.05 was considered significant.
Results
Telopodes (Tps) as one of characteristic structures of TCs were demonstrated in Fig. 1a, b. The c-kit/CD117, CD34 and vimentin in primary lung TCs were detected and shown in Fig. 1c, d, f–h. TEM tomography also showed that TCs have narrow and flat cellular prolongations surrounding other TCs in Fig. 1i, j. Mitochondria and endoplasmic reticulums in cytoplasm and nuclear of TCs were shown (Fig. 1k). Cytomembrane were also shown clearly under TEM (Fig. 1l).

Characteristics of primary lung telocytes (TCs). a, b Telocytes with telopodes (black arrow heads), c positive staining of c-kit (green), and telopodes (white arrow heads), d positive staining CD34 (green), and telopodes (white arrow heads), e nuclear staining with DAPI, f positive staining of vimentin (red), g positive staining of CD34 (green), h merge of DAPI, vimentin, and CD34, i telopode ultrastructure (Tp), j organelle and cell membrane, k mitochondrial (black arrows), mitochondrial vacuoles (white arrow head) and endoplasmic reticulum (black arrow head) in the cytoplasm, and l microvilli under TEM
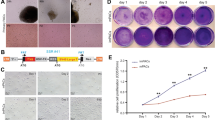
The quality of SV40 gene insert in TCsSV40 were defined with SV40 mRNA expression and shown in Fig. 2a. Characteristics of telocytes in TCsSV40 were identified and telopodes of TCsSV40 were observed and recorded in Fig. 2b. The positive staining of vimentin and CD34 was detected in primary lung TCs and TCsSV40, as presented in Fig. 2c, d.

TCs characteristics of lung TCs transferred with SV40 (TCsSV40). a SV40 mRNA expression of TCsSV40 cells, as compared with primary lung TCs or SV40-negative cells, b telopodes of TCsSV40 at 2 (b1), 5 (b2), 10 (b3), or 20 generations (b4), c positive staining of c-kit (green) in TCsSV40 at 2 (c1), 5 (c2), 10 (c3), or 20 generations (c4), d positive staining of CD34 (green) in TCsSV40 at 2 (d1), 5 (d2), 10 (d3), or 20 generations (d4), and e positive staining of CD34 (green) and vimentin (red) in TCsSV40 at 2 (e1), 5 (e2), 10 (e3), 20 (e4), 30 (e5), 40 (e6), 50 (e7), or 60 generations (e8), where cell nuclei were stained with DAPI
In order to demonstrated the immortalization and stability of TCsSV40, telopodes were observed in TCsSV40 cultured for 1, 5, 10 and 20 generations SV40 mRNA positively expressed in TCsSV40 cells through generations, as compared with primary lung TCs or SV40-negative cells (Fig. 2a). Characteristics of TCsSV40 were furthermore evaluated at 2, 5, 10, or 20 generations, respectively, including telopodes (Fig. 2b1–b4), positive staining of c-kit (Fig. 2c1–c4), positive staining of CD34 (Fig. 2d1–d4), or positive staining of both CD34 and vimentin at 2 (e1), 5 (e2), 10 (e3), 20 (e4), 30 (e5), 40 (e6), 50 (e7), or 60 generations (e8), where cell nuclei were stained with DAPI. Dynamic alterations of TCsSV40 at 2, 5, 10, 20, 30, 50, or 60 generations were recorded automatically each 30 min for 48 h and cell morphological phenomes were presented each 12 h, respectively, in Additional file 1: Figure S1. Figure 3 demonstrated that immuno-positive staining of the vimentin labeled with particle diameter at 10 nm, CD34 at18 nm, platelet-derived growth factor receptor α (PDGFRα) at 25 nm, or ckit at 40 nm in lung TCsSV40 under transmission electronic microscopy. Tomography of TCs was taken immediately after transfer with SV40 tomography, to show mitochondria-rich cytoplasm and surrounded nucleus as well as telopodes with mitochondria (Fig. 3a). PDGFRα, ckit, or vimentin and CD34 were easily detected in TCsSV40 at 2 (b, c, d), 5 (e, f), 10 (g, h), 30 (i, j), or 50 generations (k, l), respectively. Those findings of transmission electronic microscopy tomography expression demonstrated that the characteristic structures and expressions of specific markers could be stable and consistent until generation 60.

Immuno-staining transmission electronic microscopy tomography of vimentin, CD34, platelet-derived growth factor receptor α (PDGF), or ckit of lung TCs transferred with SV40 (TCsSV40). a TCsSV40 tomography, and positive staining of vimentin (
 , 10 nm), CD34 (
, 10 nm), CD34 (
 , 18 nm), PDGF (
, 18 nm), PDGF (
 , 25 nm), or ckit (
, 25 nm), or ckit (
 , 40 nm) in TCsSV40 at 2 (b–d), 5 (e, f), 10 (g, h), 30 (i, j), or 50 generations (k, l)
, 40 nm) in TCsSV40 at 2 (b–d), 5 (e, f), 10 (g, h), 30 (i, j), or 50 generations (k, l)
The profiles of transcriptional factor and lncRNA genes between primary lung TCs and TCsSV40 were compared and listed in Tables 1 and 2, and the hotmap was shown in Fig. 4. As compared with purified primary lung TCs, 367 or 621 genes were up- or down-regulated in purified TCsSV40, 668 or 890 genes in non-purified lung TCsSV40, or 36 genes up-regulated in non-purified primary lung TCs. As compared with non-purified TCsSV40, 71 or 116 genes were up- or down-regulated in purified TCsSV40. Details of transcriptional factor and lncRNA gene profiles were listed in Additional file 2: Table S1. COL3A1 and SFRP2, which code alpha 1 type III collagen and secreted frizzled-related protein 2, respectively, significantly up-regulated in purified TCsSV40, as compared with purified primary lung TCs. The patterns of transcriptional factor and lncRNA gene profiles were shown in Fig. 4a, b, respectively). The top 5 transcriptional factor genes are COL3A1, SLIT3, FST, NNAT and PCDH17, respectively. The top 5 lncRNA genes are TC17000728.hg.1, TC06001978.hg.1, TC08000302.hg.1, TC07001784.hg.1 and TC03003114.hg.1, respectively.

Hotmap of differential Genes of primary TCs, SV40-transformed TCs, primary lung cells, and SV40-transformed primary lung cells. a Hotmap of differential mRNAs. b Hotmap of LncRNAs of primary TCs, SV40-transformed TCs, primary lung cells, and SV40-transformed primary lung cells
We noticed the proliferation rate of lung TCsSV40 at all generations that we detected was significantly higher than that of primary lung TCsSV40 (Fig. 5a), of which the highest proliferation rate was observed in lung TCsSV40 at generation 2, while the lowest in at generation 50. Lung TCsSV40 were mainly located in cell cycle phases of S and G2, while primary lung TCs in sub-G1 phase (Fig. 5b). Furthermore, Fig. 5c demonstrates the alterations of cell cycle phases of Lung TCsSV40 compared with primary lung TCs. We found that the number of proliferation (Fig. 6b1–c1) or differentiated TCsSV40 (Fig. 6b3) significantly decreased in LPS administration at 1 or 0.1 μg/ml, respectively. LPS at 1 μg/ml caused a significant cell death (Fig. 6b2). Administration with SB216763 significantly inhibited the cell death (Fig. 6b2) or differentiated number (Fig. 6b3) of TCsSV40 treated with LPS at doses of 1.0 or 0.1 μg/ml, respectively, while could prevented LPS high dose-decreased proliferating as well as induced cell death (Fig. 6c1, c2). Administration with LY294002 significantly inhibited the cell death (Fig. 6b2) or differentiated number (Fig. 6b3) of TCsSV40 treated with LPS at doses of 0.1 μg/ml, respectively, while could prevented LPS high dose-decreased proliferating as well as induced cell death (Fig. 6c1, c2). Figure 7 demonstrates that the ratio of TCsSV40 proliferation, death, and dividing after cells were challenged with vehicle or TNFα at different concentrations of 0.2, 20, or 200 μg/ml and treated with vehicle, SB216763 or LY294002. The number of proliferating cells significantly reduced 48 h after administration with TNFα at doses of 200 μg/ml (Fig. 7b1, c1). Cell proliferation, cell death and differentiated number analysis of LY294002 or SB216763 stimulated TCsSV40 by celliq were shown in Additional file 3: Figure S2. The representative photos of cell bio-behaviors of TCsSV40 stimulated by LPS, LY294003 and SB216763 recorded by celliq were shown in Additional file 4: Figure S3. Additional file 5: Figure S4 showed the representative photos of cell bio-behaviors of TCsSV40 stimulated by TNF-α, LY294003 and SB216763 recorded by celliq.

Cell bio-behaviors record and analysis by celliq. a The analysis forproliferation rate of primary TCs and TCsSV40 within generation 2, 5, 10, 30 and 50. b The comparison of sub-G1 phase, G1phase, S phase and G2 phase in primary TCs and TCsSV40. c Representative pictures of cell cycle phases for primary TCs and TCsSV40. *p < 0.05

Cell proliferation, cell death and differentiated number analysis of LPS, LY294002 and/or SB216763 stimulated TCsSV40 by celliq. a1–c1 Cell proliferation analysis of LPS, LY294002 and/or SB216763 stimulated TCsSV40. a2–c2 Cell death analysis of LPS, LY294002 and/or SB216763 stimulated TCsSV40. a3–c3 Differentiated number analysis of LPS, LY294002 and/or SB216763 stimulated TCsSV40

Cell proliferation, cell death and differentiated number analysis of TNF-α, LY294002 and/or SB216763 stimulated TCsSV40 by celliq. a1–c1 Cell proliferation analysis of TNF-α, LY294002 and/or SB216763 stimulated TCsSV40. a2–c2 Cell death analysis of TNF-α, LY294002 and/or SB216763 stimulated TCsSV40. a3–c3 Differentiated number analysis of TNF-α, LY294002 and/or SB216763 stimulated TCsSV40
Discussion
TCs play an important role in the occurrence and progression of acute and chronic lung injury, asthma, and lung cancer [1, 10, 12], responsible for the highest mortality and morbidity of patients. TCs can directly communicate with a large number of cells within organ and contribute to tissue repair and regeneration, as potential alternative of cell therapies [10]. We initially isolated and purified TCs from human lung and airway tissues [16], and defined the special identity and genomic phenomes of lung TCs, different from lung stem cells, fibroblasts, alveolar type II cells, airway basal cells, proximal airway cells, CD8+ T cells from bronchial lymph nodes, and CD8+ T cells from lungs [4, 5, 17]. Our previous studies have proved the independence and specificity of human lung TCs at genomic levels and proposed TCs as a major source to connect cells, e.g. between TCs per se, or between TCs with other cells. The immunocytochemical markers of TCs include CD34, vimentin, c-kit, CD34/c-kit, CD34/vimentin, or PDGFRα [3, 10, 16, 17]. The specificity of TCs and telocytes-specific biomarkers for the identification are still to be furthermore defined, since there are a large number of telocyte heterogeneities on source, preparation, pathway, duration, and measurable variables. There are obvious differences of TCs among tissues and organs, dependent upon biological functions and characters of TC-connected tissues and organs. TCs in lung have the specificity of connection with air–liquid epithelial cells, tolerance to movement and pressure, and flexibility among barriers. In order to overcome those limits and difficulties, the present study develops a mouse lung telocyte cell-line by gene editing with lentivirus particles containing the anti-aging gene from Simian vacuolating virus 40 (SV40) gene. In the present study, we found morphology, immune biomarkers, and ultrastructure of SV40-positive TCs are similar and coincident with those of primary TCs directly isolated from mouse lungs or cultured for days. Dynamic observations of bio-behaviors demonstrated that TCsSV40 proliferation obviously increased, rather than cell movement. The capability of TCs SV40 proliferation declined by increased consecutive passages and became more stable at the 50th passage, which was still significantly higher than the primary TCs. TCsSV40 provides a repeatable and stable cell tool for deep investigation of biological roles.
SV40-infection or transformation of lung TCs alters gene expression profiles of cells at certain degrees. SV40 as a polyomavirus with icosahedral capsids of 45 nm and a 5.25 kb-long circular double-stranded DNA can replicate in macaques as its natural host, leading to chronic asymptomatic infections. SV40 small and large T antigen was transduced using lentivirus to immortalize primary cells [18]. SV40 has been strongly considered as a clinical candidate of gene delivery, replacement, or therapy, due to the lack of immunogenicity in humans and capacity to induce immune tolerance to transgene proteins. Toscano et al. [19] used the current SV40 vector genome to generate Vero-based packaging cell vector particles and expresses a large amount of the SV40 large T antigen. It indicates that SV40 delivery can be an approach or alternative of clinical gene therapy. In the present study, mouse lung TCs were transduced and immortalized with a sequence of SV40 small and large T antigen in lentivirus. Excepted for increased capability of cell proliferation, we found that morphology, measured biomarkers, movement, and ultrastructure of TCsSV40 are the same as primary TCs directly harvested from lungs. However, we did find a number of gene expression in TCsSV40 were different from primary TCs, although the meaning and values of altered gene expression profiles in TCsSV40 remain unclear.
There are numbers of questions and considerations on biological behaviors of cells with gene editing, especially about the long-term side-effects and pathophysiological responses of gene-edited cells to challenges. Although the aim of the present study to establish a mouse lung telocyte cell-line for deeply understanding molecular mechanisms of TCs, it should be aware the regulation and translational ethics of preclinical activities for the genome editing [20]. Gene-edited TCs in the present study do not need to perform the large-scale cross-platform comparisons of safety and specificity of those edited TCs as discussed for the potential of clinical applications [21], while we noticed a clear off-target effects of mouse lung TCs on the survival time and passage of TCs. The second passage of TCsSV40 had the strongest capacity of cell proliferation, while such capacity declined with the increase of telocyte passage. We found the characterizations of phenomes and functions of TCsSV40 were more stable between consecutive passages 5 and 30. Our finding is similar to the report from Taciak et al. [22] that the phenotype and functional stability of the RAW 264.7 cell line were evaluated from the 5th to 50th passage and suggested the RAW 264.7 cell line could remain stable between 10th and 30th passage. Both studies strongly suggest that it should be extremely careful to apply more than the 30th passage of the telocyte cell-line and RAW 264.7 cell line for preclinical research, in order to avoid the questions of data reliability.
TCsSV40 showed a dose-dependent response to challenges and therapies which are one of important functional properties of cells. We selected LPS as a stimulus of infection and inflammation and TNFα as an inflammatory mediator at different concentrations treated with or without signal pathway inhibitors and found the proliferative capacity of TCsSV40 declined with an increased concentration of LPS or TNFα, similar to responses of primary TCs [23]. Phosphoinositide 3-kinase (PI3K) is a family of related intracellular signal transducer enzymes to phosphorylate the 3-position hydroxyl group of the inositol ring of phosphatidylinositol, perform cellular functions (e.g. cell growth, proliferation, differentiation, and survival), and regulate cell responses to drug therapy [24]. Glycogen synthase kinase 3 (GSK3) is a serine/threonine protein kinase to regulate the addition of phosphate molecules onto serine and threonine amino acid residues, carry out biological function (e.g. cellular proliferation, migration, glucose regulation, and apoptosis), and help cellular reprogramming for clinical cartilage repair [25]. TCsSV40 were sensitive to therapeutic effects of both PI3K and GSK3 inhibitors on prevention of LPS or TNFα-reduced proliferation. Of those pathways, GSK3 seems more dominate in telocyte response to challenges. However, more basic and pre-clinical studies are still needed to ensure the safety and efficacy of gene-edited cells before clinical application. Standard and strict application for clinical research ethics must be carried out before the implementation of clinical research. We suggest that lung TCsSV40 can be applied for further studies to understand molecular mechanisms by which TCs communicate with other cells.
Conclusion
Lung TCs play important roles in maintenance of cell–cell communication within the lung and pulmonary function. The present study initially established a mouse lung telocyte cell-line and evaluated the characteristics of TCs phenomes and functions by screening gene expression profiles, ultrastructure, cell biomarkers, and responses to challenges and therapies. We defined the comparability, characteristics and stability in various passages and optimized phenotypic and functional stability of telocyte lines for application. Thus, our data suggest that the newly established lung telocyte cell-line will provide an important tool to understand roles of TCs in maintenance of lung anatomy and functions and develop a new alterative of therapies for lung diseases.
Abbreviations
- Tps:
-
telocytes
- Tps:
-
telopodes
- LPS:
-
lipopolysaccharides
- SV40:
-
simian vacuolating virus 40 small and large antigen
- TEM:
-
transmission electron microscopy
- TNF:
-
tumor necrosis factor
- SPSS:
-
Statistical Package for the Social Sciences
- ANOVA:
-
one-way analysis of variance
- PDGFRα:
-
platelet-derived growth factor receptor α
References
Song D, Cretoiu D, Cretoiu SM, et al. Telocytes and lung disease. Histol Histopathol. 2016;31(12):1303–14.
Wang J, Jin M, Ma WH, et al. The history of telocyte discovery and understanding. Adv Exp Med Biol. 2016;913:1–21.
Shoshkes-Carmel M, Wang YJ, Wangensteen KJ, et al. Subepithelial telocytes are an important source of Wnts that supports intestinal crypts. Nature. 2018;557(7704):242–6.
Song D, Cretoiu D, Zheng M, et al. Comparison of chromosome 4 gene expression profile between lung telocytes and other local cell types. J Cell Mol Med. 2016;20(1):71–80.
Wang J, Ye L, Jin M, et al. Global analyses of chromosome 17 and 18 genes of lung telocytes compared with mesenchymal stem cells, fibroblasts, alveolar type II cells, airway epithelial cells, and lymphocytes. Biol Direct. 2015;10:9.
Sun X, Zheng M, Zhang M, et al. Differences in the expression of chromosome 1 genes between lung telocytes and other cells: mesenchymal stem cells, fibroblasts, alveolar type II cells, airway epithelial cells and lymphocytes. J Cell Mol Med. 2014;18(5):801–10.
Zheng M, Sun X, Zhang M, et al. Variations of chromosomes 2 and 3 gene expression profiles among pulmonary telocytes, pneumocytes, airway cells, mesenchymal stem cells and lymphocytes. J Cell Mol Med. 2014;18(10):2044–60.
Rusu MC, Cretoiu D, Vrapciu AD, et al. Telocytes of the human adult trigeminal ganglion. Cell Biol Toxicol. 2016;32(3):199–207.
Song D, Cretoiu D, Wang X. Mitochondrial DNA in telocytes. Adv Exp Med Biol. 2017;1038:55–70.
Ye L, Song D, Jin M, et al. Therapeutic roles of telocytes in OVA-induced acute asthma in mice. J Cell Mol Med. 2017;21(11):2863–71.
Zheng Y, Wang X. Roles of telocytes in the development of angiogenesis. Adv Exp Med Biol. 2016;913:253–61.
Shi L, Dong N, Chen C, et al. Potential roles of telocytes in lung diseases. Semin Cell Dev Biol. 2016;55:31–9.
Gammage PA, Viscomi C, Simard ML, et al. Genome editing in mitochondria corrects a pathogenic mtDNA mutation in vivo. Nat Med. 2018;24(11):1691–5.
Wang W, Wang X. Single-cell CRISPR screening in drug resistance. Cell Biol Toxicol. 2017;33(3):207–10.
Blum W, Pecze L, Felley-Bosco E, et al. Establishment of immortalized murine mesothelial cells and a novel mesothelioma cell line. Vitro Cell Dev Biol Anim. 2015;51(7):714–21.
Zheng Y, Li H, Manole CG, et al. Telocytes in trachea and lungs. J Cell Mol Med. 2011;15(10):2262–8.
Zhu Y, Zheng M, Song D, et al. Global comparison of chromosome X genes of pulmonary telocytes with mesenchymal stem cells, fibroblasts, alveolar type II cells, airway epithelial cells, and lymphocytes. J Transl Med. 2015;13:318.
Blum W, Schwaller B. Calretinin is essential for mesothelioma cell growth/survival in vitro: a potential new target for malignant mesothelioma therapy? Int J Cancer. 2013;133(9):2077–88.
Toscano MG, van der Velden J, van der Werf S, et al. Generation of a vero-based packaging cell line to produce SV40 gene delivery vectors for use in clinical gene therapy studies. Mol Ther Methods Clin Dev. 2017;6:124–34.
Ando D, Meyer K. Gene editing: regulatory and translation to clinic. Hematol Oncol Clin North Am. 2017;31(5):797–808.
Lux CT, Scharenberg AM. Therapeutic gene editing safety and specificity. Hematol Oncol Clin North Am. 2017;31(5):787–95.
Taciak B, Bialasek M, Braniewska A, et al. Evaluation of phenotypic and functional stability of RAW 264.7 cell line through serial passages. PLoS ONE. 2018;13(6):e0198943.
Chi C, Jiang XJ, Su L, et al. In vitro morphology, viability and cytokine secretion of uterine telocyte-activated mouse peritoneal macrophages. J Cell Mol Med. 2015;19(12):2741–50.
Lv J, Wang L, Shen H, et al. Regulatory roles of OASL in lung cancer cell sensitivity to Actinidia chinensis Planch root extract (acRoots). Cell Biol Toxicol. 2018;34(3):207–18.
Driessen BJH, Logie C, Vonk LA. Cellular reprogramming for clinical cartilage repair. Cell Biol Toxicol. 2017;33(4):329–49.
Authors’ contributions
DS, MX and XW designed the study and patient specimen collection. DS, MX, RM and YZ completed the experimental process, data collection, literature search and generation of figures. DS, MX and XW wrote and edited the manuscript. RQ, DW and HF provided the experimental technical and laboratory facilities support. All authors reviewed the manuscript. All authors read and approved the final manuscript.
Acknowledgements
We wish to thank Mr. Hongjian Gao for his assistance in preparing cell samples for electron microscope and immuno-electron microscope detection.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
Not applicable.
Consent for publication
Not applicable.
Ethics approval and consent to participate
This study was approved by the Fudan University Ethical Committee for animal experiments.
Funding
This work was supported by Grants from the National Natural Science Foundation of China (81700008).
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author information
Authors and Affiliations
Corresponding authors
Additional files
Additional file 1: Figure S1.
Representative photos of cell bio-behaviors of primary TCs and TCsSV40 at 2, 5, 10, 20, 30, or 50 generations recorded for 0 h, 12, 24, 36 h and 48 h captured by celliq.
Additional file 3: Figure S2.
Cell proliferation, cell death and differentiated number analysis of LY294002 or SB216763 stimulated TCsSV40 by celliq.
Additional file 4: Figure S3.
Cell bio-behaviors of TCsSV40 stimulated by LPS, LY294003 and SB216763 recorded by celliq.
Additional file 5: Figure S4.
Cell bio-behaviors of TCsSV40 stimulated byTNF-α, LY294003 and SB216763 recorded by celliq.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Cite this article
Song, D., Xu, M., Qi, R. et al. Influence of gene modification in biological behaviors and responses of mouse lung telocytes to inflammation. J Transl Med 17, 158 (2019). https://doi.org/10.1186/s12967-019-1870-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12967-019-1870-y