
Abstract
Background: In livestock breeding, oocyte cryopreservation is crucial for preserving and transferring superior genetic traits. This study was conducted to examine the additional effect of melatonin to maturation and vitrification media on the in vitro developmental capacity, mitochondrial distribution, and intensity of buffalo oocytes. The study involved obtaining ovaries from a slaughterhouse and conducting two phases. In the first phase, high-quality oocytes were incubated in a maturation medium with or without 10−9M melatonin for 22 h (at 38.5°C in 5% CO2). Matured oocytes were fertilized in vitro and cultured in SOF media for seven days. In the second phase, vitrified in vitro matured oocytes were stored in vitrified media (basic media (BM) containing a combination of cryoprotectants (20% Ethyl Glycol and 20% Dimethyl sulfoxide), with or without melatonin, and then stored in liquid nitrogen. Normal vitrified/thawed oocytes were fertilized in vitro and cultured as described. Finally, the matured oocytes from the fresh and vitrified/thawed groups, both with and without melatonin, were stained using DAPI and Mitotracker red to detect their viability (nuclear maturation), mitochondrial intensity, and distribution using a confocal microscope. The study found that adding 10−9M melatonin to the maturation media significantly increased maturation (85.47%), fertilization rate (84.21%)cleavage (89.58%), and transferable embryo (48.83%) rates compared to the group without melatonin (69.85%,79.88%, 75.55%, and 37.25% respectively). Besides that, the addition of melatonin to the vitrification media improved the recovery rate of normal oocytes (83.75%), as well as the cleavage (61.80%) and transferable embryo (27.00%) rates when compared to the vitrified TCM group (67.46%, 51.40%, and 17.00%, respectively). The diffuse mitochondrial distribution was higher in fresh with melatonin (TCM + Mel) (80%) and vitrified with melatonin (VS2 + Mel groups) (76.70%), Furthermore, within the same group, while the mitochondrial intensity was higher in the TCM + Mel group (1698.60) than other group. In conclusion, Melatonin supplementation improves the developmental competence and mitochondrial distribution in buffalo oocytes in both cases(in vitro maturation and vitrification).
Similar content being viewed by others

Introduction
Over the past few years, ART (Assisted Reproductive Techniques) such as embryo transfer, artificial insemination, IVEP (in vitro embryo production), and cryopreservation have gained popularity. These techniques potentially improve reproductive efficiency, increase the production of superior animals, increasing the offspring from selected females, and also help in reduction of generation intervals in buffaloes [1]. Producing high-quality embryos from cows through in vitro production is a widely used and effective technique [2]. Maturation of the oocyte is still one of the most critical parts of the in vitro embryo production process. However, buffalo oocytes matured in vivo have higher quality and development than those matured in vitro [3]. Excessive reactive oxygen species (ROS) lead to harmful effects on cell components like fragmentation in DNA, oxidation in protein, peroxidation in lipid, and mitochondrial damage [4]. Antioxidants are included in in-vitro maturation (IVM) media to enhance oocyte maturation and embryo development [5]. Melatonin, also known as Mel, is recognized for its powerful antioxidant and anti-apoptosis properties. It can protect DNA and other components from damage and reduce oxidative stress caused by ROS in all cells [6]. Melatonin can improve the development of in vitro embryos in bovine [7], mice [8], pigs [9], dromedary camel [10], and humans [11]. Also, Mel can protect cumulus cells from nuclear fragmentation while lowering ROSlevels in bovine oocytes [12]. Melatonin can also improve the expansion of cumulus cell, the maturation rate in nucleus, cleavage and blastocyst rates in bufflo oocytes [13].
Oocyte cryopreservation can be achieved through slow freezing or vitrification, but the conventional slow-freezing method for bovine oocytes and embryos frequently causes cellular injury due to osmotic stress and ice crystallization [14]. Cryopreservation through vitrification is a highly efficient method that can minimize damage caused by low temperatures and lessen the impact of osmotic shock on embryos. The success of the cryopreservation process depends on a fine balance between the freezing rate and the use of cryoprotective agents (CPAs) to help in prevent the dehydration of cells and the formation of ice within them [15]. In the vitrification of buffalo oocytes, the most common cryoprotectants used are ethylene glycol and Dimethyl sulfoxide [16]. Addition of melatonin to the vitrification media of mice oocytes improves the oocyte quality after warmed by reducing oxidative stress which maintains oocyte mitochondrial function [17]. The addition of 10-9M Mel improved the developmental competence of parthenogenetic vitrified/ warmed in oocyte mice [18].
Mitochondria have critical roles in cellular energy metabolism for the majority of oocyte processes [19]. Oocyte quality is influenced by mitochondria; the location of mitochondria and the amount of mitochondrial transcripts are two key indications of oocyte cytoplasmic development [20]. The mitochondrial system is highly sensitive to temperature changes [21].Vitrification can affect mitochondrial function and increase ROS levels, leading to a loss of subsequent embryo development ability [22], as well as in people [23], mice [24], porcine [25], and bovine [26]. Melatonin can recover the balance of mitochondrial function and improve the meiotic maturation in vitrified mice oocytes [27]. Also could improve mitochondrial function and quality of in vitro matured aged mice oocytes [28].
Our research has shown that adding melatonin to the vitrification media can enhance the developmental competence and mitochondrial intensity of camel oocytes after being thawed [10]. This study is the first one to investigate the effects of melatonin on both maturation and vitrification media. Our goal was to examine how melatonin affects the morphology, developmental competence, mitochondrial intensity, and distribution of fresh in-vitro matured and/or vitrified/thawed in-vitro matured buffalo oocytes.
Materials and methods
Chemicals and media used in this study were purchased from Sigma-Aldrich (Sant. Louis, MO, USA) unless otherwise mentioned.
Collection of oocytes and IVM
The laboratory received buffalo ovaries from the slaughterhouse in a thermos filled with warm normal saline solution at 37 °C (0.9% NaCl + 100 IU penicillin and 100μg/ml streptomycin). The ovaries were cleaned several times in warmed saline solution, then placed in a water bath at 37 °C till the oocytes were ready to be aspirated. An 18-gauge needle connected to a sterile syringe containing aspiration and washing medium consists of PBS, 4 mg/ml BSA and 50 μg/ml gentamicin to aspirate COCs (cumulus oocyte complexes) from follicles 2–8 mm in diameter. Thereafter, follicular content was transferred to Falcon tube and left for 10–15 min to settle at 37 °C. The COCs were examined under a stereo microscope and rinsed for 3-times in oocyte maturation medium. According to Kandil et al. [29], buffalo oocyte quality was assessed. Depending on the cumulus investment and equally granulated ooplasm, there were four groups of COCs under a stereomicroscope (90 x) as follows:
-
a-Excellent: Oocytes with at least five layers of fully developed cumulus cells (CC) and evenly granulated dark ooplasm.
-
b- Good: One to four layers of cumulus cells and evenly granulated dark cytoplasm.
-
c- Fair: Oocytes are incompletely encircled by cumulus cells, and the ooplasm has little granulation.
-
d-Denuded: Oocytes were covered by zona pellucida and had no cumulus cells.
Excellent and good oocytes transferred to in vitro maturation medium (TCM-199 + 10% FCS + 10 μg/ml FSH + 50 μg/ml gentamicin) with or without 10 −9 M Mel. (M5250, Sigma). The COCs were in vitro matured in an incubator at 38.5°C in a humidified atmosphere with 5% CO2 for 22 h. After 22 h from incubation, the cytoplasmic maturation of oocytes assessed according to the degree of cumulus expansion, according to the extent of cumulus-cell development, the cytoplasmic maturation of buffalo oocytes was evaluated and divided into 4-grades according to Kandil et al. [29]:
-
G0: Without expansion.
-
GI: With slight expansion.
-
GII: Moderate expansions.
-
GIII: Full expansionII.
Maturation of oocytes’ nuclear determined by the presence of the 1st polar body (Pb) in the perivitelline space. The 1st Pb was detected using an inverted microscope at 20X magnification. The expansion rate was obtained by dividing the number of oocytes per grade by the total number of oocytes and then the results were multiplied by 100. The nuclear maturation rate (M II) was calculated by dividing the number of mature oocytes with 1st Pb on the total number of oocytes, and the results were multiplied by 100.
In vitro embryo production
To prepare for fertilization, matured oocytes with full cumulus expansion was washed in a fertilization medium, specifically Fert-TALP [30] supplemented with 6 mg/ml BSA. Meanwhile, the frozen semen straw was thawed in a 37°C water bath for 30 s. The spermatozoa were then suspended in 5 ml of sperm-TALP medium, containing 10μg/ml heparin and 4 mg/ml BSA, before undergoing centrifugation for 10 min at 1800 rpm (344 g) in 15 ml conical tubes. After removing the supernatant, the sperm pellet was resuspended in Fert-TALP media for 3 ml and centrifuged for 5 min at 1800 rpm. Once centrifugation was complete, the supernatant was removed, and the sperm pellet was suspended in Fert-TALP supplemented with 20 μM penicillamine, 10μM/ml hypotaurine, and 1µM/ml epinephrine (PHE) + 6mg/ml BSA.. Next, the sperm suspension was placed into a 4-well culture plate with the oocytes and covered with warm 200 ul mineral oil. The concentration of sperm was adjusted to 1 × 10 6 sperm/ml. The sperm and oocytes were co-incubated for 18 h at 38.5°C and 5% CO2 in humid air.
The zygotes were decomulated using multiple pipetting and then washed three times and placed in a (modified synthetic oviduct fluid, mSOF) solution with 5 mg/ml of BSA and 50 μg/ml of gentamycin. Then, they incubated at 38.5°C with 5% CO2 in a humidified environment. The embryos were observed using an inverted microscope to determine their cleavage rate and development to the morula and blastocyst stages on Days 5 and 7. We change the mSOF medium every 2 days. Also, we checked the rate of cleavage and embryo development on days 2, 5, and 7. Then the detection of the cleavage rate and transferable embryos (morula and blastocyst) is calculated as follows:
-
a-Cleavage rate was obtained by dividing the number of cleaved zygotes by the total number of fertilized oocytes with the second polar body multiplied by 100.
-
b-The transferable embryo (morula and blastocyst) was obtained by dividing the number of morula and blastocyst by the total number of cleaved zygotes multiplied by 100 [16].
Oocytes vitrification
Matured oocytes without cumulus cells with 1st polar body were placed in basic media (BM). This medium consists of 9.5 ml TCM-199, 0.5 ml FCS, and 50 μg/ml gentamicin. Afterward, oocytes were placed in vitrification solution one (VS,1 consists of BM, 10% Ethyl Glycol (EG), and 10% Dimethyl sulfoxide (DMSO)) for 1 min. Then oocytes are transferred to vitrification solution two (VS2, contains BM, 20% EG, and 20% DMSO) for 30 s. Finally, it loaded in a VS2 medium [16].
Oocytes loading
Oocytes were loaded into 0.25ml of French straw using a micro-classic pipette (Karl Hecht No. 558). First loading, about 20 ul hold medium consisting of BM + 0.5 M sucrose was drawn and separated from the oocytes in the vitrification solution (25 ul) using an air bubble chamber from two sides then drawn hold medium to the end of the straw. About 25–30 oocytes and polyvinyl powder were loaded into the straw to seal the loaded straw. The straw was exposed to liquid nitrogen (LN2) vapor for 10 s. then plunged into LN2 and stored for 7 days [16].
Oocytes warming
After 7 days of being vitrified, straws containing vitrified oocytes were warmed in a 37°C water bath for 10–15 s. The straws were shaken to mix the vitrification solution and holding medium with oocytes, then the warmed oocytes were transferred to a new thawing medium that contained sucrose. The sucrose was gradually diluted in a three-step process, starting with a concentration of 0.5 M, followed by 0.3 M, and finally 0.17 M, with 1 min of equilibration time in each solution. The oocytes were washed thrice with fresh BM [16].
Evaluation of morphological changes
Recovered vitrified oocytes were evaluated under an inverted microscope and the recovered vitrified oocyte rate equaled the number of recovered oocytes divided by the number of vitrified oocytes multiplied by 100.
Normal oocytes were spherical and symmetrical, showing no evidence of damage to the membrane, swelling, degeneration, or fragmented or shrinking cytoplasm. Conversely, abnormal oocytes exhibited a ruptured or cracked zona pellucida (ZP), fragmented or shrunk cytoplasm, and signs of degeneration or leakage of cellular contents. \(\mathrm{Morphologically}\;\mathrm{normal}\;\mathrm{oocyte}\;\mathrm{ rate} =\frac{\text{No}.\;\mathrm{of}\;\mathrm{normal}\;\mathrm{vitrified}\;-\;\mathrm{thawed}\;\mathrm{oocytes}\;\mathrm{with}\;1\mathrm{st}\;\mathrm{polar}\;\mathrm {body}\;x\;100}{\mathrm{Total}\;\mathrm{no}.\;\mathrm{of}\;\mathrm{recovered}\;\mathrm{invitro}\;\mathrm{matured}\;\mathrm{vitrified}\;/\;\mathrm{thawed}\;\mathrm{oocytes}}\) [16].
Detection of mitochondrial distribution and intensity:
In-vitro matured buffalo oocytes either fresh or vitrified/warmed were stained using Mitochondrion specific fluorescent probe, Mito Tracker Red FM (thermo fisher). Confocal microscopy (Zeiss LSM 710) was used to determine mitochondrial distribution in the oocytes according to manufacturer instructions. In-vitro matured buffalo oocytes fresh or vitrified/ warmed were incubated with a final concentration of 500 nM Mito Tracker Red FM in PBS for 30 min at 37 ºC in the incubator. Oocytes were washed twice in PBS and co-incubated with PBS containing five µg/mL DAPI to counterstain the nucleus and detect the 1st polar body in matured oocytes (MII). DAPI detection in excitation 358 mm and emission in 4601mm wavelength using a confocal microscope in 20X magnification. Oocytes were then washed and mounted in PBS and were visualized in a glass bottom culture plate 12 mm diameter (thermo fisher) using a confocal microscope (Zeiss 710). The Mito Tracker red fluorescence was observed using an argon laser in a 581nm excitation line and 644 nm emission filters. One optical section was examined for each oocyte, in the plane where the nucleus was visible. The determination of the mitochondrial distribution was as follows: peripheral mitochondrial distributions were in the oocytes in which no mitochondria were found at the center of the oocyte; semi-peripheral distributions were in those with an inhomogeneous distribution of mitochondria in the inner region of the oocyte; and diffuse distributions were those with a uniform distribution of mitochondria across the entire inner region of the oocyte cytoplasm. The mitochondrial intensity is automatically detected through the software of a confocal microscope [16]. The experiment was replicated three times, with a group of 10 oocytes in each replicate.
Experimental design
Experiment 1: effect of melatonin addition to maturation media on maturation rate and developmental competence of buffalo oocytes
The total number of buffalo ovaries used in this experiment was 290 and excellent (n = 283) and good (n = 240) oocytes were used for IVM.
Two groups of in vitro matured oocytes The control group (TCM n = 289) underwent maturation without any melatonin treatment, while the treated group (TCM + Mel n = 234) was exposed to 10 −9 M melatonin. After maturation, the oocytes were fertilized in-vitro and cultured using the abovementioned method. This was done to determine the rate of maturation, cleavage, and transferable embryo.
Experiment 2: effect of melatonin on morphological changes of vitrified/warmed in-vitro matured buffalo oocytes
The total number of buffalo ovaries used in this experiment was 322 and excellent (n = 345) and good (n = 360) oocytes were used for IVM.
Excellent and good oocytes were in-vitro matured as experiment 1. In-vitro matured oocytes with 1st PB (526) were used for vitrification in three groups: vitrified matured oocytes without Mel as control (VTCM n = 176), vitrified matured oocytes with Mel in maturation medium (VTCM + Mel n = 182) and vitrified matured oocytes with Mel in vitrification medium (VS2 + Mel n = 168) all groups were vitrified using 20%Ethelyn Glycol + 20% Dimethyl sulfoxide as described previously The vitrified in vitro matured buffalo oocytes were thawed after one week of vitrification for morphological evaluation.
Experiment 3: effect of melatonin on developmental competence of vitrified/thawed in vitro matured buffalo oocytes
Normal oocytes used for this experiment were 374. VTCM (n = 112), VTCM + Mel (n = 128) and VS2 + Mel (n = 134).
Morphologically normal vitrified/thawed oocytes were subjected to in vitro fertilization as described above. At the end of the fertilization period, the presumptive zygotes were cultured in mSOF medium for 7 days. The cleavage and transferable embryo rate were evaluated
Experiment 4: effect of melatonin on mitochondrial distribution and intensity of fresh and vitrified/warmed in-vitro matured buffalo oocytes
A total number of 150 fresh in-vitro matured and in-vitro matured vitrified / thawed warmed buffalo oocytes were stained using a Mitochondrion-specific fluorescent probe, Mito Tracker Red FM (Thermo Fisher). Confocal microscopy (Zeiss LSM 710) was used to determine mitochondrial distribution and intensity in the oocytes as described previously. DAPI stain was used to counterstain the nucleus and enable the determination of nuclear maturation.
Statistical analysis
The IBM SPSS software program version 25.0 was used to analyze the data. The data was described using numbers and percentages. The significance of differences was determined using an independent t-test, analysis of variance (ANOVA), followed by a post hoc test (P ≤ 0.01). The chi-square test was used for categorical variables.
Results:
Experiment 1: effect of melatonin addition in in-vitro maturation media on maturation rate and developmental competence of buffalo oocytes
Effect of melatonin addition in in-vitro maturation media on cytoplasmic maturation (cumulus expansion rate) of matured buffalo oocytes
Results in Table 1 demonstrate that the addition of 10-9M melatonin to maturation media resulted in a significant (P<0.05) increase in the percentage of oocytes with GIII cumulus expansion by (61.11%) when compared with oocytes cultured without melatonin media (39.44%). The TCM group showed a significant increase in cumulus expansion rate GII, GI, and G0 (23.18%, 20.76%, and 16.60 %, respectively) when compared with TCM +Mel (18.80%, 12.82 %, and 7.26%, respectively).
Effect of melatonin addition in in-vitro maturation media on nuclear maturation rate (1st Pb) of matured buffalo oocytes
The maturation rate of buffalo oocytes was significantly (P < 0.05) higher in the TCM + Mel group (85.47%) when compared with the TCM group (68.85%). The buffalo oocytes without polar bodies matured in the TCM group showed a significant increase (P < 0.05) (14.18%) when compared with oocytes cultured in the TCM + Mel group (6.83%). The degenerated oocytes showed a significant increase in the TCM group (16.95%) than TCM + Mel (7.69%) (Table 2).
Effect of melatonin on developmental competence of in vitro matured buffalo oocytes
The fertilization rate, cleavage rate, and transferable embryo rate of in-vitro matured oocytes were significantly higher (P < 0.05) in TCM + Mel 84.21%, 89.58%, 48.83% respectively) when compared with TCM (79.88%,75.55%, and 37.25% respectively), (Table 3).
Experiment 2: effect of melatonin on morphological changes of vitrified/ warmed in-vitro matured buffalo oocytes
Table 4 showed no significant differences between the percentages of recovered in-vitro matured buffalo oocytes after thawing in vitrified groups.
There were significant differences (P ≤ 0.01) between the percentages of normal in-vitro matured vitrified buffalo oocytes after warming in VTCM, VTCM + Mel, and VS2 + Mel groups (67.46%, 72.72%, and 83.75%, respectively). There were significant differences between the percentages of morphologically abnormal in in-vitro matured vitrified buffalo oocytes after warming in VTCM, VTCM + Mel, and VS2 + Mel groups (32.53%, 27.27%, 16.25%, respectively) (Table 4).
The morphological abnormality of vitrified oocytes with leakage of cellular content and fragmented cytoplasm was significantly higher in the VTCM + Mel group (31.25% and 25%, respectively) than in the VTCM group (22.22% and 9.26%, respectively) and VS2 + Mel group (23.1% and 11.53%). The percentage of vitrified oocytes with cracking of zona pellucida and zona rapture was significantly higher in the VTCM group (25.93% and 14.81%, respectively) than inVS2 + Mel (19.23% and 7.69%, respectively) and VTCM + Mel group (10.42% and 8.33%., respectively). The Percentage of oocytes with shrinkage of cytoplasm was significantly higher in theVS2 + Mel group (38.45%) than VTCM and VTCM + Mel groups (27.78% and 25%, respectively) (Table 5). The abnormality of matured oocytes was significantly (P < 0.01) higher in shrinking in the cytoplasm, cracking of zona pellucida, and leakage of cellular content when compared with fragment cytoplasm and zona rapture in the VTCM group. The leakage, shrinking, and fragment of the cytoplasm were significantly higher than the cracking and rupture of the zona pellucida of oocytes in the VTCM + Mel group. Moreover, the shrinking of cytoplasm, cracking of zona pellucida, and leakage of cellular content are significantly higher than fragmented cytoplasm and zona rapture in the VS2 + Mel group.
Experiment 3: effect of melatonin on developmental competence of in-vitro matured vitrified/warming buffalo oocytes
The fertilization rate showed no significant differences between the VTCM group (82.14%), VTCM + Mel group (89.06%), and VS + Mel group (85.07%). The cleavage rate and transferable embryos rate of vitrified /warmed in-vitro matured buffalo oocytes was significantly (P < 0.01) increased in VTCM + Mel (63.20%, 28.20% respectively) and VS2 + Mel groups (61.80%, 27% respectively), when compared with VTCM group (51.40%, 17%, respectively). In contrast, there was no significance between VTCM + Mel and VS2 + Mel in both cleavage and transferable embryo rates (Table 6, Fig. 1).

Buffalo oocyte quality and oocyte developmental competence using inverted microscope Zeiss using magnification 20X, A showed oocyte quality G = good, Exc = Excellent, B cytoplasmic maturation GIII = full expansion, GII = Modred expansion, GI = low expansion, C nuclear matured oocytes, 1st PB = 1st polar body, D Fertilized oocytes 2nd PB = 2nd polar body, E transferable embryo, mor = morula, EBL = early blastocyst
Experiment 4: effect of melatonin on mitochondrial distribution and intensity of fresh in-vitro matured and vitrified/warming in-vitro matured buffalo oocytes
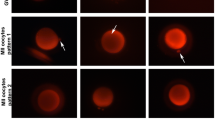
More mitochondria diffusely and centrally distributed were significantly increased in the TCM + Mel group (80%, 20% respectively); semi-diffused distribution was significantly higher in the TCM group (20%); peripheral distribution was significantly higher in the VTCM + Mel group (60%); semi- peripheral distribution was significantly higher in VTCM group (70%) (Table 7, Fig. 2). The defused mitochondrial distribution was significantly higher (P < 0.01) in the TCM, TCM + mel., VTCM + Mel, and VS2 + Mel than other distributions, while in VTCM the simi-peripheral distribution was significantly higher than other distributions.

Effect of melatonin addition on in vitro maturation and /or vitrification media on viability and mitochondrial distribution and intensity on in vitro matured fresh and vitrified /thawed buffalo oocytes using confocal microscope zeiss 710 in magnification 200X
The mean number of mitochondrial intensity of fresh mature oocytes in the TCM + Mel group (1698.60) was significantly higher (P < 0.01) than the fresh TCM group (456.48) and vitrified/ warmed groups. Among vitrified groups, there was no significant difference in mitochondrial intensity between the TCM + Mel group (327.67) and the VS2 + Mel group (298.47) (Table 8).
Discussion
Melatonin is a hormone that regulates various reproductive functions, including the competence of oocyte development. Melatonin is unique in its hydrophilic and lipophilic properties, allowing it to move quickly throughout all cellular compartments [31]. This means it can be found in the cytosol, nuclei, and mitochondria [32]. Unlike other antioxidants, melatonin has the most desirable characteristics. Its metabolites work together to detoxify radicals [33] and it also stimulates the action of important antioxidant enzymes such as glutathione peroxidase (GSH) and superoxide dismutase (SOD) [34].
The antioxidant melatonin shows promise in boosting the development of oocytes. Our study has revealed that melatonin can improve buffalo oocytes' maturation rate and developmental potential. We observed a significant difference in cumulus expansion and nuclear maturation between the oocytes cultured in the TCM group and those treated with TCM + Mel. Such findings are consistent with previous studies that have demonstrated the ability of melatonin to protect oocytes from oxidative stress and enhance their maturation process. Moreover, melatonin has been reported to promote the maturation of porcine oocytes [35], buffalo [13, 36], mice [28], bovine [37], ovine [38], and humans [39]. Melatonin has been found to improve cytoplasmic maturation in bovine oocytes by returning organelles to their proper locations, increasing the levels of intracellular GSH and ATP, activating genes responsible for oocyte maturation [37, 40], and enhancing antioxidant capacity in mouse eggs [41] Additionally, melatonin can regulate gene expression in both oocytes and cumulus cells [35], studies have shown the positive effects of melatonin on the development of blastocysts in various animals, including bovine [37], sheep [42] (porcine [35], mice [28], buffalo [13, 36], goat [43]), and even human [44]. However, some studies have suggested that melatonin may not be as effective in bovine, as adding it to an IVM medium did not result in higher cleavage or blastocyst rates [45]. These differing results may be due to factors such as the dosage, supplier, or batch of melatonin used for IVM. Melatonin has been found to improve the developmental ability of low-quality oocytes. The goats treated with melatonin showed a higher growth rate to the blastocyst stage than untreated oocytes [46]. Additionally, melatonin enhances preantral follicle pig oocytes' developmental competence [47] and cleavage and blastocyst rate of inferior quality bovine oocytes [48]. Melatonin also helps to lower the levels of ROS in oocytes [49, 50], safeguarding them from the negative effects of oxidative stress and preventing meiotic spindle changes [51], DNA damage [52], apoptosis [53], endoplasmic reticulum stress [54], and aging [55]. Furthermore, it increases the antioxidant capacity in mouse oocytes [35].
This study is the first to explore the impact of melatonin on the physical changes of vitrified/thawed in vitro matured buffalo oocytes. The findings show that vitrification with 10-9M melatonin resulted in significant differences (p ≤ 0.01) between the VTCM + Mel, VS2 + Mel, and VTCM groups. The VTCM + Mel group had a high percentage of vitrified oocytes with leaked cellular content and fragmented cytoplasm (31.25% and 25%, respectively). The VTCM group had a high percentage of vitrified oocytes with cracking of zona pellucida and zona rapture (25.93% and 14.81%, respectively). The VS2 + Mel group had a high percentage of oocytes with cytoplasmic shrinkage (38.45%). Ismail's [16] previous research indicated that vitrification of buffalo oocytes significantly affected their morphology. Melatonin enhanced the normal physical changes of vitrified buffalo oocytes, in agreement with previous reports on mouse oocytes. It is important to note that mammalian oocytes are very sensitive to temperature and osmotic pressure changes due to their complex subcellular structure [56]. Cryopreservation can cause significant damage to both oocytes and embryos, with the oocyte membrane being the primary site of damage [57]. Vitrification can lead to hardening of the ZP of oocytes, which can significantly impact their ability to develop [57]. However, studies have shown that melatonin, a potent antioxidant and free radical scavenger, can help reduce oocyte ROS levels. This has been observed in pigs [58], buffalo [13], bovine [59], and camel [10] species. Additionally, melatonin helps maintain adequate levels of endogenous antioxidant enzymes [60] and boosts the level of the antioxidant glutathione (GSH) [61].
After undergoing the process of vitrification and thawing, the oocytes experienced a decrease in GSH synthesis [62] and an increase in ROS production, leading to a reduction in their developmental competence [62].
However, the addition of melatonin has been found to enhance the development of vitrified/thawed oocytes during in vitro development [56]. Recent studies have shown that melatonin supplementation can improve the developmental competence of oocytes in buffalo, and this positive impact is attributed to its ability to reduce oxidative stress [63]. Further research has demonstrated that ROS detoxification is crucial in restoring embryo metabolic functions after thawing [64]. Adding melatonin to maturation, culture, and vitrification media has been found to significantly influence the developmental competence of bovine oocyte and embryo vitrification procedure [65]. Adding 10–7 mol/L melatonin to the vitrification-warming and in vitro maturation media showed a significant increase in the maturation rate for mouse GV oocytes [66]. Adding melatonin to in vitro maturation media also improved the meiotic maturation of mouse vitrified oocytes [27]. Moreover, melatonin improved the in vitro survival rate of mouse preantral follicles after vitrification-thawing [67]. Additionally, it enhanced the developmental potential of parthenogenetically activated embryos produced from vitrified mature oocytes in mice [68]. The developmental competence of vitrified-warmed mouse oocytes can be improved by adding melatonin (10–9 mol/L) during warming, recovery, parthenogenetic activation, and in vitro culture of embryos. This improvement is mainly due to the modulation of oxidative stress, improvement of mitochondrial function, regulation of spindle assembly and chromosome arrangement, and inhibition of apoptosis [18]. However, melatonin may not always be effective in improving the development of cryopreserved metaphase II mouse oocytes, as some studies have shown. This may be due to varying concentrations and culture conditions of melatonin [69]. This study found that a higher percentage of oocytes in the TCM + Mel group (80%) had a diffuse distribution of mitochondria throughout the cytoplasm compared to other groups. This result is consistent with Ismail's findings [16]. The peripheral distribution of mitochondria was significantly higher in the vitrified VTCM + Mel group (60%), while the semi-peripheral distribution was significantly higher in the vitrified VTCM group (70%). The fresh TCM + Mel group showed significant central distribution (20%). The distribution of mitochondria in mature oocytes is usually homogeneous throughout the cytoplasm, providing equal distribution of mitochondria between zygote cells after fertilization [70]. It has also been shown that a higher density and distribution of active mitochondria is correlated with faster nuclear maturation [71]. Antioxidants added to the maturation media can enhance oocyte mitochondrial activity. Previous studies have demonstrated that melatonin acts as a protective agent that can promote oocyte development [72]. Melatonin enhances oocyte mitochondrial function, increases ATP production [73], and reduces mitochondrial DNA damage [11]. In addition, studies show that melatonin can have positive effects on goats [74] and bovine oocytes. It lowers intracytoplasmic ROS levels, increases mitochondrial activity, and boosts ATP content [37]. Melatonin increases mitochondrial function after an in vitro culture of sheep's secondary follicles. To be more precise, bovine oocytes treated with melatonin had higher normal distribution rates of mitochondria than untreated oocytes [49]. Melatonin addition also increases the rates of oocytes with dispersed mitochondria in the cytoplasm. Furthermore, it reduces mitochondrial oxidative stress in mouse embryos [75] and can potentially improve the quality of human oocytes [11]. Melatonin has been found to be beneficial in enhancing the developmental competence of cryopreserved oocytes. This is achieved through the reduction of oxidative stress and protection of mitochondrial function. Melatonin also indirectly promotes the meiotic competence of oocytes while decreasing the production of ROS during oocyte maturation [76]. Melatonin is a crucial substance that helps improve the quality of oocyte vitrification and reduces oxidative stress imbalance in PCOS patients, thus maintaining oocyte mitochondrial function [17]. Studies have shown that the process of vitrification and warming can affect the intracellular oxidative status, leading to an increase in ROS activity that may harm the structure, distribution, and function of mitochondria in oocytes [77]. In buffalo, mitochondria are arranged in the cortical region [78], which is the first site to be damaged during freezing [79]. Moreover, damage to oocyte mitochondria during vitrification procedures may occur with or without visible morphological changes [80]. The distribution and intensity of mitochondria can impact the cytoplasmic and nuclear maturation of oocytes [81]. Melatonin is a potent antioxidant that specifically targets mitochondria [65]. In a recent study, the group treated with melatonin showed significantly higher mitochondria intensity than the other groups. These results are consistent with Kandil's [10] findings that mitochondrial intensity was significantly higher in melatonin-treated oocytes of camels.
In conclusion
Adding 10 −9M melatonin to the maturation or vitrification medium can enhance the in vitro developmental capacity of buffalo oocytes, and increase the viability, mitochondrial distribution, and intensity of fresh and vitrified thawed buffalo oocytes.
Availability of data and materials
Not applicable.
References
Baruselli PS, de Carvalho JGS, Elliff FM, da Silva JCB, Chello D, de Carvalho NAT. Embryo transfer in buffalo (Bubalus bubalis). Theriogenology. 2020;150:221–8.
Currin L, Baldassarre H, de Macedo MP, Glanzner WG, Gutierrez K, Lazaris K, et al. Factors affecting the efficiency of in vitro embryo production in prepubertal mediterranean water buffalo. Animals. 2022;12(24):3549.
Lonergan P, Fair T. Maturation of Oocytes in Vitro. Annu Rev Anim Biosci. 2016;4(1):255–68. Available from: https://www.annualreviews.org/doi/10.1146/annurev-animal-022114-110822.
Lopes AS, Lane MTJ. Oxygen consumption and ROS production are increased at the time of fertilization and cell cleavage in bovine zygotes. Hum Reprod. 2010;25:2762–73.
Mukherjee A, Malik H, Saha AP, Dubey A, Singhal DK, Boateng S, et al. Resveratrol treatment during goat oocytes maturation enhances developmental competence of parthenogenetic and hand-made cloned blastocysts by modulating intracellular glutathione level and embryonic gene expression. J Assist Reprod Genet. 2014;31(2):229–39. Available from: http://link.springer.com/10.1007/s10815-013-0116-9.
Cruz MHC, Leal CLV, Cruz JF, Tan DX, Reiter RJ. Essential actions of melatonin in protecting the ovary from oxidative damage. Theriogenology. 2014;82(7):925–32. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0093691X14003525.
An Q, Peng W, Cheng Y, Lu Z, Zhou C, Zhang Y, et al. Melatonin supplementation during in vitro maturation of oocyte enhances subsequent development of bovine cloned embryos. J Cell Physiol. 2019;234(10):17370–81. Available from: https://onlinelibrary.wiley.com/doi/10.1002/jcp.28357.
Lan K-C, Lin Y-C, Chang Y-C, Lin H-J, Tsai Y-R, Kang H-Y. Limited relationships between reactive oxygen species levels in culture media and zygote and embryo development. J Assist Reprod Genet. 2019Feb 10;36(2):325–34. Available from: http://link.springer.com/10.1007/s10815-018-1363-6.
Lin T, Lee JE, Kang JW, Oqani RK, Cho ES, Kim SB, et al. Melatonin supplementation during prolonged in vitro maturation improves the quality and development of poor-quality porcine oocytes via anti-oxidative and anti-apoptotic effects. Mol Reprod Dev. 2018;85(8–9):665–81. Available from: https://onlinelibrary.wiley.com/doi/10.1002/mrd.23052.
Kandil OM, Takla MS, Noseir WMB, Hattab SA. Effects of vitamin C and melatonin on the viability and mitochondrial distribution of in vitro matured dromedary camel oocytes. Egypt J Chem. 2022;65(3):395–405.
Liu Y-J, Ji D, Liu ZB, Wang TJ, Xie FF, Zhang ZG, et al. Melatonin maintains mitochondrial membrane potential and decreases excessive intracellular Ca2+ levels in immature human oocytes. Life Sci. 2019;235:116810. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0024320519307374.
Rodrigues-Cunha MC, Mesquita LG, Bressan F, del Collado M, Balieiro JCC, Schwarz KRL, et al. Effects of melatonin during IVM in defined medium on oocyte meiosis, oxidative stress, and subsequent embryo development. Theriogenology. 2016;86(7):1685–94. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0093691X16302278.
El-Sanea AM, Abdoon ASS, Kandil OM, El-Toukhy NE, El-maaty AMA, Ahmed HH. Effect of oxygen tension and antioxidants on the developmental competence of buffalo oocytes cultured in vitro. Int J One Heal. 2021;14(1):78–84.
Martínez-Burgos M, Herrero L, Megías D, Salvanes R, Montoya MC, Cobo AC, et al. Vitrification versus slow freezing of oocytes: effects on morphologic appearance, meiotic spindle configuration, and DNA damage. Fertil Steril. 2011;95(1):374–7. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0015028210022156.
Ribeiro JC, Carrageta DF, Oliveira PF, Bernardino RL, Alves MG. Aquaporins and animal gamete cryopreservation: advances and future challenges. Animals. 2022;12(3):359.
Ismail EA, Elsayed MAI, Hemeida NA, Abdoon ASS, Kandil OM. Effect of different cryoprotectant agents on mitochondrial distribution and developmental competence in buffalo oocyte (Bubalus Bubalis). Egypt J Chem. 2022;65(1):539–53.
Jiang Y, Shi H, Liu Y, Zhao S, Zhao H. Applications of Melatonin in Female Reproduction in the Context of Oxidative Stress. Oxid Med Cell Longev. 2021;2021:6668365.
Pan B, Qazi IH, Guo S, Yang J, Qin J, Lv T, et al. Melatonin improves the first cleavage of parthenogenetic embryos from vitrified–warmed mouse oocytes potentially by promoting cell cycle progression. J Anim Sci Biotechnol. 2021;12(1):1–17.
Brevini TAL, Vassena R, Francisci C, Gandolfi F. Role of adenosine triphosphate, active mitochondria, and microtubules in the acquisition of developmental competence of parthenogenetically activated pig oocytes1. Biol Reprod. 2005;72(5):1218–23. Available from: https://academic.oup.com/biolreprod/article-lookup/doi/10.1095/biolreprod.104.038141.
Babayev E, Wang T, Szigeti-Buck K, Lowther K, Taylor HS, Horvath T, et al. Reproductive aging is associated with changes in oocyte mitochondrial dynamics, function, and mtDNA quantity. Maturitas. 2016;93:121–30. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0378512216301451.
Gualtieri R, Kalthur G, Barbato V, Di Nardo M, Adiga SK, Talevi R. Mitochondrial dysfunction and oxidative stress caused by cryopreservation in reproductive cells. Antioxidants. 2021;10(3):1–23.
Gao Z, Yao G, Zhang H, Liu H, Yang Z, Liu C, et al. Resveratrol protects the mitochondria from vitrification injury in mouse 2-cell embryos. Cryobiology. 2020;95:123–9. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0011224020300857.
Lei T, Guo N, Liu JQ, Tan MH, Li YF. Vitrification of in vitro matured oocytes: effects on meiotic spindle configuration and mitochondrial function. Int J Clin Exp Pathol. 2014;7(3):1159–65. Available from: http://www.ncbi.nlm.nih.gov/pubmed/24696732.
Lei T, Guo N, Tan M, Li Y. Effect of mouse oocyte vitrification on mitochondrial membrane potential and distribution. J Huazhong Univ Sci Technol [Medical Sci]. 2014;34(1):99–102. Available from: http://link.springer.com/10.1007/s11596-014-1238-8.
Dai J, Wu C, Muneri CW, Niu Y, Zhang S, Rui R, et al. Changes in mitochondrial function in porcine vitrified MII-stage oocytes and their impacts on apoptosis and developmental ability. Cryobiology. 2015;71(2):291–8. Available from: https://linkinghub.elsevier.com/retrieve/pii/S001122401500214X.
Zhao XM, Du WH, Wang D, Hao HS, Liu Y, Qin T, et al. Effect of cyclosporine pretreatment on mitochondrial function in vitrified bovine mature oocytes. Fertil Steril. 2011;95(8):2786–8. https://doi.org/10.1016/j.fertnstert.2011.04.089.
Gao L, Du M, Zhuan Q, Luo Y, Li J, Hou Y, et al. Melatonin rescues the aneuploidy in mice vitrified oocytes by regulating mitochondrial heat product. Cryobiology. 2019;89(May):68–75. https://doi.org/10.1016/j.cryobiol.2019.05.005.
Almohammed ZNH, Moghani-Ghoroghi F, Ragerdi-Kashani I, Fathi R, Tahaei LS, Naji M, et al. The effect of melatonin on mitochondrial function and autophagy in in vitro matured oocytes of aged mice. Cell J. 2020;22(1):9–16.
Kandil OM, Abdoon ASS, Murakami M, Otoi T, Suzuki T. New technique, using a portable co2 incubator, for the production of in vitro fertilized egyptian buffalo embryos. J Reprod Dev. 1999;45(5):315–20.
Parrish JJ. Bovine In vitro fertilization: In vitro oocyte maturation and sperm capacitation with heparin. Theriogenology. 2013;81(1). https://doi.org/10.1016/j.theriogenology.2013.08.005.
Acuna-Castroviejo DN-NM, Reiter RJ, Escames G. Melatonin actions in the heart; more than a hormone. Melatonin Res. 2018;1:21–6.
Venegas C, García JA, Escames G, Ortiz F, López A, Doerrier C, et al. Extrapineal melatonin: analysis of its subcellular distribution and daily fluctuations. J Pineal Res. 2012Mar;52(2):217–27. Available from: https://onlinelibrary.wiley.com/doi/10.1111/j.1600-079X.2011.00931.x.
Manchester LC, Coto-Montes A, Boga JA, Andersen LP, Zhou Z, Galano A, et al. Melatonin: an ancient molecule that makes oxygen metabolically tolerable. J Pineal Res. 2015;59:403–19.
Fischer TW, Kleszczyński K, Hardkop LH, Kruse N, Zillikens D. Melatonin enhances antioxidative enzyme gene expression (CAT, GPx, SOD), prevents their UVR-induced depletion, and protects against the formation of DNA damage (8-hydroxy-2’-deoxyguanosine) in ex vivo human skin. J Pineal Res. 2013;54(3):303–12. Available from: https://onlinelibrary.wiley.com/doi/10.1111/jpi.12018.
Yang L, Wang Q, Cui M, Li Q, Mu S, Zhao Z. Effect of Melatonin on the In Vitro Maturation of Porcine Oocytes, Development of Parthenogenetically Activated Embryos, and Expression of Genes Related to the Oocyte Developmental Capability. Animals. 2020;10(2):209. Available from: https://www.mdpi.com/2076-2615/10/2/209.
Nagina G, Asima A, Nemat U, Shamim A. Effect of melatonin on maturation capacity and fertilization of Nili-Ravi buffalo (Bubalus bubalis) oocytes. Open Vet J. 2016;6(2):128–34.
Zhao XM, Wang N, Hao HS, Li CY, Zhao YH, Yan CL, et al. Melatonin improves the fertilization capacity and developmental ability of bovine oocytes by regulating cytoplasmic maturation events. J Pineal Res. 2018;64(1):e12445. Available from: https://onlinelibrary.wiley.com/doi/10.1111/jpi.12445.
Barros VRP, Monte APO, Santos JMS, Lins TLBG, Cavalcante AYP, Gouveia BB, et al. Effects of melatonin on the in vitro growth of early antral follicles and maturation of ovine oocytes. Domest Anim Endocrinol. 2020;71:106386. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0739724019300645.
Nishihara T, Hashimoto S, Ito K, Nakaoka Y, Matsumoto K, Hosoi Y, et al. Oral melatonin supplementation improves oocyte and embryo quality in women undergoing in vitro fertilization-embryo transfer. Gynecol Endocrinol. 2014;30(5):359–62. Available from: http://www.tandfonline.com/doi/full/10.3109/09513590.2013.879856.
Pang Y, Zhao S, Sun Y, Jiang X, Hao H, Du W, et al. Protective effects of melatonin on the in vitro developmental competence of bovine oocytes. Anim Sci J. 2018;89(4):648–60.
Wu, Pan, Qazi, Yang, Guo, Yang, et al. Melatonin improves in vitro development of vitrified-warmed mouse germinal vesicle oocytes potentially via modulation of spindle assembly checkpoint-related genes. cells. 2019;8(9):1009. Available from: https://www.mdpi.com/2073-4409/8/9/1009.
Tian X, Wang F, Zhang L, He C, Ji P, Wang J, et al. Beneficial effects of melatonin on the in vitro maturation of sheep oocytes and its relation to melatonin receptors. Int J Mol Sci. 2017;18(4):834.
Saeedabadi S, Abazari-Kia AH, Rajabi H, Parivar K, Salehi M. Melatonin improves the developmental competence of goat Oocytes. Int J Fertil Steril. 2018;12(2):157–63.
Nishihara T, Hashimoto S, Ito K, Nakaoka Y, Matsumoto K, Hosoi Y, et al. Oral melatonin supplementation improves oocyte and embryo quality in women undergoing in vitro fertilization-embryo transfer. Gynecol Endocrinol. 2014;30(5):359–62.
Tsantarliotou MP, Attanasio L, De Rosa A, Boccia L, Pellerano G, Gasparrini B. The effect of melatonin on bovine in vitro embryo development. Ital J Anim Sci. 2007;6(SUPPL. 1):488–9.
Soto-Heras S, Roura M, Catalá MG, Menéndez-Blanco I, Izquierdo D, Fouladi-Nashta AA, et al. Beneficial effects of melatonin on in vitro embryo production from juvenile goat oocytes. Reprod Fertil Dev. 2018;30(2):253. Available from: http://www.publish.csiro.au/?paper=RD17170.
Cao Z, Gao D, Tong X, Xu T, Zhang D, Wang Y, et al. Melatonin improves developmental competence of oocyte-granulosa cell complexes from porcine preantral follicles. Theriogenology. 2019;133:149–58. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0093691X19301220.
Yang M, Tao J, Chai M, Wu H, Wang J, Li G, et al. Melatonin improves the quality of inferior bovine oocytes and promoted their subsequent IVF embryo development: Mechanisms and results. Molecules. 2017;22(12):2059.
El-Raey M, Geshi M, Somfai T, Kaneda M, Hirako M, Abdel-Ghaffar AE, et al. Evidence of melatonin synthesis in the cumulus oocyte complexes and its role in enhancing oocyte maturation in vitro in cattle. Mol Reprod Dev. 2011;78(4):250–62.
Keshavarzi S, Salehi M, Farifteh-Nobijari F, Hosseini T, Hosseini S, Ghazifard A, et al. Melatonin modifies histone acetylation during in vitro maturation of mouse oocytes. Cell J. 2018;20(2):244–9.
Banerjee J, Maitra D, Diamond MP, Abu-Soud HM. Melatonin prevents hypochlorous acid-induced alterations in microtubule and chromosomal structure in metaphase-II mouse oocytes. J Pineal Res. 2012;53(2):122–8. Available from: https://onlinelibrary.wiley.com/doi/10.1111/j.1600-079X.2012.00977.x.
Leem J, Bai G, Kim J, Oh JS. Melatonin protects mouse oocytes from DNA damage by enhancing nonhomologous end‐joining repair. J Pineal Res. 2019;67(4). Available from:https://onlinelibrary.wiley.com/doi/10.1111/jpi.12603.
Tripathi A, PremKumar KV, Pandey AN, Khatun S, Mishra SK, Shrivastav TG, et al. Melatonin protects against clomiphene citrate-induced generation of hydrogen peroxide and morphological apoptotic changes in rat eggs. Eur J Pharmacol. 2011Sep;667(1–3):419–24. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0014299911006881.
Park HJ, Park JY, Kim JW, Yang SG, Jung JM, Kim MJ, et al. Melatonin improves the meiotic maturation of porcine oocytes by reducing endoplasmic reticulum stress during in vitro maturation. J Pineal Res. 2018;64(2):e12458. Available from: https://onlinelibrary.wiley.com/doi/10.1111/jpi.12458.
Lord T, Nixon B, Jones KT, Aitken RJ. Melatonin Prevents postovulatory oocyte aging in the mouse and extends the window for optimal fertilization in vitro1. Biol Reprod. 2013;88(3). Available from: https://academic.oup.com/biolreprod/article-lookup/doi/10.1095/biolreprod.112.106450.
Li W, Cheng K, Zhang Y, Meng Q, Zhu S, Zhou G. No effect of exogenous melatonin on development of cryopreserved metaphase II oocytes in mouse. J Anim Sci Biotechnol. 2015;6(1):1–7. https://doi.org/10.1186/s40104-015-0041-0.
Kandil OM, Aboelwafa FB, Ismail EA, Kandeel SM, Ghanem N, Gamal El-Din AEK. Effects of Ascorbic Acid on Maturation Rate, Morphology, and Gene Expression of Vitrified In Vitro Matured Dromedary Camel Oocytes. World’s Vet J. 2022;12(4):418–29.
Kang J-T, Koo O-J, Kwon D-K, Park H-J, Jang G, Kang S-K, et al. Effects of melatonin on in vitro maturation of porcine oocyte and expression of melatonin receptor RNA in cumulus and granulosa cells. J Pineal Res. 2009;46(1):22–8. Available from: https://onlinelibrary.wiley.com/doi/10.1111/j.1600-079X.2008.00602.x.
Mogas T. Update on the vitrification of bovine oocytes and in vitro-produced embryos. Reprod Fertil Dev. 2019;31(1):105–17.
Fathi M, Salama A, El-Shahat KH, EL-Sherbiny HR, Abdelnaby EA. Effect of melatonin supplementation during IVM of dromedary camel oocytes (Camelus dromedarius) on their maturation, fertilization, and developmental rates in vitro. Theriogenology. 2021;172:187–92. https://doi.org/10.1016/j.theriogenology.2021.05.021.
Gao C, Han HB, Tian XZ, Tan DX, Wang L, Zhou GB, et al. Melatonin promotes embryonic development and reduces reactive oxygen species in vitrified mouse 2-cell embryos. J Pineal Res. 2012;52(3):305–11. Available from: https://onlinelibrary.wiley.com/doi/10.1111/j.1600-079X.2011.00944.x.
Jiao GZ, Cao XY, Cui W, Lian HY, Miao YL, Wu XF, et al. Developmental Potential of Prepubertal Mouse Oocytes Is Compromised Due Mainly to Their Impaired Synthesis of Glutathione. Sun QY, editor. PLoS One. 2013;8(3):e58018. Available from: https://dx.plos.org/10.1371/journal.pone.0058018.
Succu S, Pasciu V, Manca ME, Chelucci S, Torres-Rovira L, Leoni GG, et al. Dose-dependent effect of melatonin on postwarming development of vitrified ovine embryos. Theriogenology. 2014;81(8):1058–66. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0093691X1400065X.
Donnay I, Auquier P, Kaidi S, Carolan C, Lonergan P, Mermillod P, et al. Vitrification of in vitro produced bovine blastocysts: methodological studies and developmental capacity. Anim Reprod Sci. 1998;52(2):93–104. Available from: https://linkinghub.elsevier.com/retrieve/pii/S0378432098000980.
Parnpai R, Liang Y, Ketudat-Cairns M, Somfai T, Nagai T. Vitrification of buffalo oocytes and embryos. Theriogenology. 2016;86(1):214–20.
Wu Z, Pan B, Qazi IH, Yang H, Guo S, Yang J, et al. Melatonin improves in vitro development of vitrified-warmed mouse germinal vesicle oocytes potentially via modulation of spindle assembly checkpoint-related genes. Cells. 2019;8(9):1009.
Ganji R, Nabiuni M, Faraji R. Development of mouse preantral follicle after in vitro culture in a medium containing melatonin. Cell J. 2015;16(4):546–53.
Pan B, Yang H, Wu Z, Qazi I, Liu G, Han H, et al. Melatonin improves parthenogenetic development of vitrified-warmed mouse oocytes potentially by promoting G1/S cell cycle progression. Int J Mol Sci. 2018;19(12):4029. Available from: http://www.mdpi.com/1422-0067/19/12/4029.
Kim MK, Park EA, Kim HJ, Choi WY, Cho JH, Lee WS, et al. Does supplementation of in-vitro culture medium with melatonin improve IVF outcome in PCOS? Reprod Biomed Online. 2013;26(1):22–9. Available from: https://linkinghub.elsevier.com/retrieve/pii/S1472648312006037.
Stojkovic M, Machado SA, Stojkovic P, Zakhartchenko V, Hutzler P, Gonçalves PB, et al. Mitochondrial Distribution and Adenosine Triphosphate Content of Bovine Oocytes Before and After In Vitro Maturation: Correlation with Morphological Criteria and Developmental Capacity After In Vitro Fertilization and Culture1. Biol Reprod. 2001;64(3):904–9. Available from: https://academic.oup.com/biolreprod/article-lookup/doi/10.1095/biolreprod64.3.904.
Yu Y, Dumollard R, Rossbach A, Lai FA, Swann K. Redistribution of mitochondria leads to bursts of ATP production during spontaneous mouse oocyte maturation. J Cell Physiol. 2010;224(3):672–80.
Hu KL, Ye X, Wang S, Zhang D. Melatonin application in assisted reproductive technology: a systematic review and meta-analysis of randomized trials. Front Endocrinol (Lausanne). 2020;11. Available from: https://www.frontiersin.org/article/10.3389/fendo.2020.00160/full.
He C, Wang J, Zhang Z, Yang M, Li Y, Tian X, et al. Mitochondria synthesize melatonin to ameliorate its function and improve mice oocyte’s quality under in vitro conditions. Int J Mol Sci. 2016;17(6):939. Available from: http://www.mdpi.com/1422-0067/17/6/939.
Soto-Heras S, Catalá M-G, Roura M, Menéndez-Blanco I, Piras A-R, Izquierdo D, et al. Effects of melatonin on oocyte developmental competence and the role of melatonin receptor 1 in juvenile goats. Reprod Domest Anim. 2019;54(2):381–90. Available from: https://onlinelibrary.wiley.com/doi/10.1111/rda.13378.
Zhang Y, Li W, Ma Y, Wang D, Zhao X, Zeng C, et al. Improved development by melatonin treatment after vitrification of mouse metaphase II oocytes. Cryobiology. 2016;73(3):335–42. https://doi.org/10.1016/j.cryobiol.2016.09.171.
He M, Zhang T, Yang Y, Wang C. Mechanisms of oocyte maturation and related epigenetic regulation. Front Cell Dev Biol. 2021;9(March):1–18.
Barros VRP, Monte APO, Santos JMS, Lins TLBG, Cavalcante AYP, Gouveia BB, et al. Melatonin improves development, mitochondrial function and promotes the meiotic resumption of sheep oocytes from in vitro grown secondary follicles. Theriogenology. 2020;144:67–73. https://doi.org/10.1016/j.theriogenology.2019.12.006.
Clérico G, Taminelli G, Veronesi JC, Polola J, Pagura N, Pinto C, et al. lMitochondrial function, bastocyst development and live foals born after ICSI of immature vitrified/warmed equine oocytes matured with or without melatonin. Theriogenology. 2021;160:40–9.
Mondadori RG, Santin TR, Fidelis AAG, Name KPO, da Silva JS, Rumpf R, et al. Ultrastructure of in vitro oocyte maturation in buffalo ( Bubalus bubalis ). Zygote. 2010;18(4):309–14. Available from: https://www.cambridge.org/core/product/identifier/S0967199409990335/type/journal_article.
Ogawa B, Ueno S, Nakayama N, Matsunari H, Nakano K, Fujiwara T, et al. Developmental Ability of Porcine In Vitro Matured Oocytes at the Meiosis II Stage After Vitrification. J Reprod Dev. 2010;56(3):356–61. Available from: http://www.jstage.jst.go.jp/article/jrd/56/3/56_10-005H/_article.
Cao X, Li J, Xue H, Wang S, Zhao W, Du Z, et al. Effect of vitrification on meiotic maturation, mitochondrial distribution and glutathione synthesis in immature silver fox cumulus oocyte complexes. Theriogenology. 2017;91:104–11. Available from: https://doi.org/10.1016/j.theriogenology.2016.12.037.
Acknowledgements
The authors acknowledge the financial support through the National Research Centre project ID: 13050417 titles (Overcome effect of harsh environmental condition on early embryonic developmental competence in buffalo) and this work done in the Embryo and Genetic Resources Conservation Bank in the National Research Centre.
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB). This work is financially supported by National Research Centre project ID: 13050417 titles (Overcome effect of harsh environmental condition on early embryonic developmental competence in buffalo).
Author information
Authors and Affiliations
Contributions
O.K. designed the experiment and supported all the equipment, chemicals, and facilities, S.A. brought the samples and did lab work, E.I. helped in the lab work, O.K., S.A., R.A., N.I. help in writing the manuscript and analysis of data.
Corresponding author
Ethics declarations
Ethics approval and consent to particpate
This investigation was conducted by standard protocols, with no pain or injury to the buffalo. Additionally, the procedures of experiments were approved by the Ethics Committee of the National Research Centre, Cairo, Egypt.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Kandil, O.M., Rahman, S.M.A.E., Ali, R.S. et al. Effect of melatonin on developmental competence, mitochondrial distribution, and intensity of fresh and vitrified/thawed in vitro matured buffalo oocytes. Reprod Biol Endocrinol 22, 39 (2024). https://doi.org/10.1186/s12958-024-01209-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12958-024-01209-7