
Abstract
Background
The suprachiasmatic nucleus (SCN) and the cholinergic system of various regions of the hypothalamus participate in the regulation of gonadotropin-releasing hormone (GnRH) and gonadotropin secretion, which are necessary for the occurrence of ovulation. In the present study, our goal was to analyse the effects of unilaterally blocking the muscarinic receptors in the SCN on ovulation and steroid secretion.
Methods
Cyclic rats were randomly allotted to one of the experimental groups. Groups of 8–14 rats were anaesthetized and microinjected with 0.3 μl of saline or a solution of atropine (62.5 ng in 0.3 μl of saline) into the left or right SCN at 09.00 or 19.00 h during diestrus-1 or on the proestrus day. The rats were euthanized on the predicted day of oestrus, and evaluated ovulation and levels of progesterone and oestradiol. Other groups of 10 rats were microinjected with atropine into the left or right SCNs at 09.00 h on the proestrus day, were euthanized eight h later, and luteinizing hormone (LH) was measured.
Results
At 09.00 or 19.00 h during diestrus-1, atropine microinjections into the SCNs on either side did not modify ovulation. The atropine microinjections performed at 09.00 h of proestrus into either side of the SCN blocked ovulation (right SCN: 1/9 ovulated vs. 9/10 in the saline group; left SCN: 8/14 ovulated vs. 10/10 in the saline group). The LH levels at 17.00 h in the rats that were microinjected with atropine at 09.00 h of proestrus were lower than those of the controls. In the non-ovulating atropine-treated rats, the injection of synthetic LH-releasing hormone (LHRH) restored ovulation. Atropine treatment at 19.00 h of proestrus on either side of the SCN did not modify ovulation, while the progesterone and oestradiol levels were lower.
Conclusion
Based on the present results, we suggest that the cholinergic neural information arriving on either side of the SCN is necessary for the pre-ovulatory secretion of LH to induce ovulation. Additionally, the regulation of progesterone and oestradiol secretion by the cholinergic innervation of the SCN varies with the time of day, the day of the cycle, and the affected SCN.
Similar content being viewed by others


Background
The suprachiasmatic nucleus (SCN) of the hypothalamus contains a “central pacemaker” that orchestrates circadian rhythms. The circadian system is important for successful reproduction because it ensures that the period of maximal fertility aligns with peak of sexual motivation [1]. The SCN regulates gonadotropin-releasing hormone (GnRH) secretion, the pre-ovulatory surge of luteinizing hormone (LH) and ovulation [2–5].
The pulsatile frequency and secretion amplitude of GnRH regulate gonadotropin and sex steroid hormone secretion, follicular maturation and ovulation [6–11]. According to Robertson et al. [12], there are two possible routes by which the SCN could regulate GnRH neurons and hence the LH surge. One pathway involves the direct innervation of the GnRH neurons by the SCN. Direct synapses exist between the neurons in the SCN and GnRH neurons in the preoptic anterior area (POA), and vasoactive intestinal peptide (VIP) and vasopressin (AVP) act as neurotransmitters in these synapses [13, 14]. The second pathway involves an indirect circuit linking the SCN with the GnRH neurons through a relay station in the anteroventral periventricular nucleus (AVPV), which is a hypothalamic region that is critical for the GnRH/LH surge in rodents. The SCN sends AVPergic fibres directly to the AVPV [15] where kisspeptidergic neurons have been identified [9]. These neurons are probably responsible for generating the oestradiol-mediated GnRH and LH surges that are required for ovulation [16].
The central cholinergic system regulates the circadian system and modulates the activities of neurons in the SCN. In rats, the SCN receives cholinergic projections from the basal forebrain, including the nucleus basalis magnocellularis (NBM), the brainstem (from the pedunculopontine tegmental nucleus (PPTg)), and the latero-dorsal tegmental nucleus (LDTg) [17]. According to Yang et al. [18], multiple cholinergic-modulatory actions occur via multiple receptors and ion channels that may allow acetylcholine to exert delicate control of the excitabilities of SCN neurons in different physiological settings. Damaging the NBM with quinolic acid reduces SCN cholinergic input and decreases the expressions of VIP and AVP by SCN neurons. The extents of the declines in the expressions of VIP and AVP correlate with the numbers of destroyed cholinergic afferents, which supports the notion that acetylcholine plays a direct role in regulating the metabolic activities of SCN neurons [19].
The SCN sends a cholinergic efferent to the diagonal band of Broca and the medial septum [17, 20], and GnRH neurons are present in these two nuclei [21], which suggests the possibility that the cholinergic system of the SCN regulates GnRH neurons via a direct cholinergic pathway. According to Turi et al. [22], the GnRH neurons are regulated by cholinergic neurons located in different areas of the brain in apposition to GnRH neurons. Everett et al. [23] demonstrated that at proestrus, the cholinergic system participates in the regulation of ovarian hormone secretion and ovulation. Unilaterally implanting atropine crystals into either side of the preoptic-anterior hypothalamic area (POA-AHA) blocks ovulation in a manner that depends on the side into which the POA-AHA atropine is implanted and the day of the oestrous cycle on which the treatment is performed [24]. Moreover, GnRH injection at the predicted time of proestrus restores ovulation [25]. The participation of the muscarinic system in the regulation of progesterone and oestradiol serum levels varies with the hours and days of the oestrous cycle [26], [27].
Asymmetry is a natural trait that is displayed across the range of basic particles to complex organ functions [28]. The SCN consists of two bilaterally paired nuclei [29], each of which is capable of generating a behavioural activity rhythm [30] and may act as an independent oscillator [31]. To our knowledge, there is no information on the effects on spontaneous ovulation that are elicited by the unilateral block of the cholinergic receptors in the SCN. Since in rats on diestrus-1 the unilateral implant of atropine on either side of the POA-AHA blocked ovulation, while the same treatment with rats on proestrus day did not [24] and the subcutaneously injecting atropine sulphate to rats on each day of the oestrous cycle blocked ovulation, depending on day and hour of treatment [32], the aims of the present study were to analyse the effects of unilaterally blocking the SCN’s muscarinic system in rats during diestrus-1 or proestrus on ovulation. According to Zhao and Kriegsfeld [33] immortalized GnRH cells exhibit intrinsic daily changes in their sensitivity to neurochemicals stimulating their activity, which did not depend on clock genes. To test the possibility that blocking SCN cholinergic inputs has an effect on ovulation regulation, the effects of injecting atropine into either side of the SCN were evaluated at two different times of day, i.e., during the light period at 09.00 h and at the beginning of the dark period at 19.00 h.
Methods
Animals
This study was performed with adult, virgin, 3-month-old, female, CIIZ-V strain rats weighing 230–260 g from our own breeding stock. The animals were maintained under controlled light (on from 05:00 to 19:00 h) and temperature (22 ± 2 °C) conditions with free access to food (Purina S.A., México) and tap water. The animals’ oestrous cycles were monitored via cytological examinations of daily vaginal smears. Only rats that exhibited at least two consecutive 4-day oestrus cycles were used in the experiment.
Unilateral microinjection of atropine into the left or right SCN
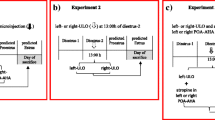
The experimental protocol is illustrated in Fig. 1a and b. At 09.00 or 19.00 h, groups of 4-day cyclic rats in diestrus-1 or proestrus were anaesthetized with sodium pentobarbital (30 mg/kg I.P.; Pisabental, PISA Agropecuaria S.A. de C.V. México). To assess the effectiveness of the anaesthesia, we used the tail pinch method [34]. After verifying that the rats were anaesthetized, they were placed in a stereotaxic apparatus (David Kopf Instruments, Tujunga, CA). The skin of the skull was sectioned, and the left or right side of the skull was drilled with a 1-mm bit. Subsequently, a 29-gauge stainless steel microinjection needle was lowered into the left or right side of the SCN. The SCN was located according to the coordinates of the Paxinos and Watson [35] atlas (left SCN: −0.3 mm AP, +0.2 mm ML, and −9.1 mm DV; right SCN: −0.3 mm AP, −0.4 mm ML, and −9.1 mm DV). The microinjection needle was connected to a 25-μl Hamilton syringe placed on a microinjection pump (CMA/100; BAS, Stockholm, Sweden) with a Teflon tube (0.65 mm OD 9, 0.12 mm OI; Bioanalytical Systems, Inc., West Lafayette, IN). Next, 0.3 μl of saline or 62.5 ng of atropine (based on reference [24, 25, 32]) in 0.3 μl of saline (atropine is a non-specific muscarinic blocker with a half-life of 2 h; Sigma Chemical Co., St. Louis, MO) was microinjected into the left (saline n = 36; atropine n = 42 rats) or right SCN (saline n = 36; atropine n = 35 rats). After surgery, each animal was placed in an individual cage with warm sawdust and heated with an incandescent lamp until the animal awoke.

Schematic representation of the treatment design. The groups of rats were microinjected with atropine in the left or right SCN at 09.00 or 19.00 h during diestrus-1 (a), and the other groups were microinjected on the proestrus day (b). In panel c, note that the group of rats were microinjected with atropine in the left or right SCN at 09.00 h and at 14.00 h these rats were subcutaneously injected with 3.7 μg/kg synthetic LHRH. All animals were sacrificed on the predicted vaginal oestrus day
To examine the potential diffusion of the liquid from one SCN into the other, ten rats were microinjected with 0.3 μl of methylene blue dye in saline solution (10 mg/ml) in the left or right SCN [36]. No diffusion of methylene blue dye into the contralateral SCN, the ipsilateral nuclei (including the POA), or the third ventricle was observed.
Effects of the unilateral microinjection of atropine on the pre-ovulatory LH peak
To assess whether the atropine treatment modified the pre-ovulatory LH surge that occurs at 17.00 h on the proestrus day, groups of rats were injected at 09.00 h on the proestrus day with either vehicle or atropine in the left SCN (n = 10 per group) or right SCN (n = 10 per group) following the procedures described above. The rats were sacrificed at 17.00 h on the same day.
Effects of injecting synthetic LH-releasing hormone on the ovulation responses of non-ovulating rats microinjected with atropine in the SCN
Following the procedures described above, groups of rats on the proestrus day were treated at 09.00 h with atropine in the left SCN (n = 9 per group) or right SCN (n = 9 per group) and treated again at 14.00 h (5 h later). The rats were subcutaneously injected with 3.7 μg/kg of synthetic LH-releasing hormone (LHRH)-Gly-OH (Sigma Chemical Co., St. Louis, MO) diluted in saline following the protocol of Humphrey and colleagues [37]. The rats were sacrificed the next day (predicted oestrus) between 09.00 and 10.00 h (Fig. 1c).
Effects of the microinjection of atropine into the SCN at 09.00 h on the proestrus day on ovarian morphology
To assess the effects the microinjection of the atropine into the SCN on ovarian morphology, the left and right ovaries from the experimental rats were removed, cleaned of adherent fat tissue, fixed in Bouin’s solution for 24 h, dehydrated and embedded in paraffin. The ovaries (left and right) of three randomly selected rats from each group [i.e., the atropine-microinjected and vehicle-injected groups] were serially sectioned at 10 μm and stained with haematoxylin-eosin. All ovary sections were analysed for the presence of antral follicles and new and old corpora lutea with the aid of a Nikon binocular microscope.
Autopsy procedures
On the predicted vaginal oestrus day (72 h or 24 h after treatment), the rats were sacrificed by decapitation [38, 39]. The blood of the trunk from each animal was collected, allowed to clot and centrifuged at 3,000 rpm for 15 min. The serum was stored at −20 °C until the progesterone, oestradiol and LH concentrations were measured via specific radioimmunoassays. Upon autopsy, the oviducts were dissected, and the numbers of ova shed were counted with the aid of a stereoscopic microscope (Nikon, Model C-PS). The brain was subsequently dissected and quickly placed on a plate cooled with dry ice to verify the accuracy of the microinjection site.
Brain histologic processing
The brains of all rats treated with vehicle or atropine were frozen, and 100-μm sections were obtained on a cryostat (MICROM HM 505 N, Walldorf, Germany). The section were mounted on slides and stained with 1 % cresyl violet. All results of the present study are based on rats with verified microinjections into the SCN (Fig. 2a, b).

Diagrammatic representation of the locations of the microinjection sites. a The black circles represent the microinjection sites. Schematic illustration of a coronal section taken from the rat brain atlas of Paxinos and Watson [35]. b Nissl-stained coronal sections illustrating the trajectories of the microneedles into the left SCN. 3 V: third ventricle; OC: optic chiasm; SCN: suprachiasmatic nucleus. The arrowheads indicate the trajectory of the microinjections into the left side of the SCN
Hormone measurements
The serum concentrations of progesterone (ng/ml) and oestradiol (pg/ml) were measured using radioimmunoassay (RIA) with kits purchased from Diagnostic Products (Los Angeles, CA, USA). The intra-assay coefficients of variation were 8.35 and 8.12 % for the progesterone and oestradiol assays, respectively, and the inter-assay coefficients of variation were 9.45 and 9.28 % for the progesterone and oestradiol assays, respectively.
The LH levels in the sera (ng/ml) were measured using the double antibody RIA technique with reagents and protocols kindly supplied by the NIADDK National Pituitary Program (Bethesda, MD, USA). The intra- and inter-assay variations were approximately 5.1 and 6.5 %, respectively, for LH. The results are expressed in terms of the NIADDK standards RP-2 for FSH and LH.
Statistical analysis
The statistical analyses were performed using GraphPad Instant 3. The ovulation rates (i.e., the numbers of ovulating animals/the numbers of treated animals) were analysed using Fisher’s exact probability or Chi-square tests. Data regarding the numbers of ova shed were analysed using Kruskal–Wallis tests followed by a Mann–Whitney U tests. The hormonal serum level results were analysed using analysis of variance (ANOVA) followed by Tukey’s tests. When two means were compared, we used Student’s t-tests. P-values below 0.05 were considered statistically significant.
Results
Ovulation and the numbers of ova shed
To investigate whether the cholinergic system of the left and/or right SCN regulated spontaneous ovulation in the rat, the ovulation rates and the numbers of the ova shed were determined. The ovulation rates and the numbers of ova shed were not modified by the microinjection of atropine into either side of the SCN at 09.00 or 19.00 h during diestrus-1 (Table 1).
The microinjection of atropine into the left SCN at 09:00 h during proestrus resulted in a lower ovulation rate (57 % of the atropine-treated rats ovulated vs. 100 % of the saline-injected group; p < 0.001, chi-square test). Only 11 % of the rats that were microinjected with atropine into the right SCN ovulated, whereas 90 % of those microinjected with saline ovulated. No differences in the numbers of ova shed by the ovulating rats that were microinjected with vehicle or atropine were observed (Table 1).
At 19.00 h during proestrus, the atropine treatments on either side of the SCN did not modify the ovulation rate or the number of ova shed (Table 1).
Progesterone and oestradiol serum levels in the rats microinjected with atropine during diestrus-1
Compared with the saline-microinjected animals, atropine microinjection at 09.00 h in either SCN did not modify the serum progesterone or oestradiol levels (Fig. 3).

Steroid hormone serum levels in the rats treated at 09.00 and 19.00 h during diestrus-1. Means ± SEMs of the progesterone (ng/ml) and oestradiol serum levels (pg/ml) in the rats subjected to microinjections of vehicle into the left (Vh L-SCN) or right (Vh R-SCN) SCN or atropine microinjection into the left (ATR L-SCN) or right (ATR R-SCN) SCN at 09.00 or 19.00 h during diestrus-1. *p < 0.05 vs. the respective groups microinjected with Vh (ANOVA followed by Tukey’s test). ♦p < 0.001 vs. ATR-L-SCN (ANOVA followed by Tukey’s test)
In the rats that were microinjected with atropine in the right SCN at 19.00 h, the progesterone levels were higher, whereas atropine treatment in the left SCN resulted in lower oestradiol levels compared to the rats that were microinjected with the vehicle (Fig. 3). Atropine microinjection in the right SCN yielded higher progesterone levels compared with those of the rats that were treated in the left SCN (Fig. 3).
Serum progesterone and oestradiol levels in the rats that were microinjected with atropine during proestrus
The rats in proestrus that were treated with atropine into either side of the SCN at 09.00 h exhibited significantly lower ovulation rates; consequently, the progesterone and oestradiol levels in the ovulating and non-ovulating animals were analysed separately.
Compared with the group that was microinjected with vehicle, the non-ovulating rats that were microinjected with atropine into the left SCN (n = 6) exhibited lower progesterone levels. Atropine microinjection into the right SCN (n = 8) yielded lower oestradiol levels compared with those of the rats that were treated in the left SCN (n = 8; Fig. 4). In the ovulating rats that were microinjected with atropine in the left SCN (n = 8), the serum progesterone and oestradiol levels were not different from those of the saline-microinjected rats. A statistical analysis of the difference between the ovulating rats that were treated with atropine in the right SCN and the corresponding vehicle-treated group was not possible because only one rat in the right SCN atropine treatment group ovulated (Fig. 4).

Steroid hormone serum levels of the rats treated at 09.00 h on the proestrus day. The means ± the SEMs for the progesterone (ng/ml) and oestradiol serum levels (pg/ml) of the non-ovulating and ovulating rats that received microinjections of the vehicle in the left (Vh L-SCN) or right (Vh R-SCN) SCN or atropine microinjection in the left (ATR L-SCN) or right (ATR R-SCN) SCN at 09.00 h during proestrus. (1) Represents the level of one animal’s ovulation. * p < 0.01 vs. the respective groups microinjected with Vh (ANOVA followed by Tukey’s test). ♦p < 0.05 vs. ATR-L-SCN (ANOVA followed by Tukey’s test)
In the proestrus rats that were treated at 19.00 h with atropine into the right SCN, the progesterone levels were lower than those in the corresponding vehicle-treated group. The atropine treatment in the left SCN resulted in lower oestradiol levels compared with those of the corresponding vehicle-treated group of rats (Fig. 5).

Steroid hormone serum levels of the rats treated at 19.00 h on the proestrus day. The means ± the SEMs of the progesterone (ng/ml) and oestradiol serum levels (pg/ml) in the rats subjected to microinjections of vehicle in the left (Vh L-SCN) or right (Vh R-SCN) SCN or atropine microinjections in the left (ATR L-SCN) or right (ATR R-SCN) SCN at 19.00 h during proestrus. * p < 0.05 vs. their respective groups that were microinjected with Vh (Student’s t -test)
LH serum levels at 17.00 h during proestrus
The LH levels in the rats that were microinjected with atropine at 09.00 h on either side of the SCN and sacrificed at 17.00 h were lower than the LH levels the corresponding vehicle-microinjected group (Fig. 6). Atropine treatment in the right SCN yielded lower LH levels compared with those of the rats with left SCN treatments (ATR R-SCN 1.4 ± 0.2 vs. ATR L-SCN 5.1 ± 1.3; p < 0.023, Student’s t -test).

LH serum levels evaluated at 17.00 h on the proestrus day. The means ± the SEMs for the LH serum levels (ng/ml) of the rats subjected to microinjection of the vehicle in the left (Vh L-SCN) or right (Vh R-SCN) SCN or atropine microinjection into the left (ATR L-SCN) or right (ATR R-SCN) SCN at 09.00 h during proestrus and sacrificed at 17.00 h on the same day. * p < 0.01 vs. the respective groups that were microinjected with Vh (Student’s t -test)
Influence of LHRH on ovulation in the rats that were microinjected with atropine in the SCN
Because the microinjection of atropine into the SCN at 09.00 h during proestrus blocked ovulation, and the pre-ovulatory release of GnRH is required for ovulation, we examined whether the microinjection of atropine into the SCN altered the pre-ovulatory release of GnRH. The results obtained from the rats that were microinjected with atropine in either SCN were combined. Among the rats that were microinjected with atropine in the SCN, ovulation was induced in 77 % of the rats that were treated following a subcutaneous injection of synthetic LHRH (Fig. 7).

Influence of LHRH on ovulation in the rats that were microinjected with atropine. Ovulation rates of the rats subjected to microinjections of atropine in the SCN (ATR-SCN) at 09.00 h during proestrus and rats microinjected with atropine followed by injection with 3.7 μg/kg per body weight of synthetic LHRH at 14.00 h on the same day (ATR-SCN + LHRH). The animals were sacrificed 18–20 h after treatment on the predicted oestrus day. *p < 0.025 vs. the atropine group (Fisher’s exact probability test)
Ovarian morphology
Independently of the day and time, vehicle microinjection into either side of the SCN did not modify the ovarian morphology (Fig. 8a). A histological analysis of the ovaries of the non-ovulating rats that were treated with atropine in either side of the SCN at 09.00 h during proestrus revealed the presence of a few antral follicles and old corpora lutea (Fig. 8b).

Ovarian histologies of the rats with atropine into the SCN at 09.00 h during proestrus. The micrographs correspond to the greatest section of the ovary at 10-μm thick that was stained with haematoxylin-eosin. The rats were sacrificed on the predicted vaginal oestrus day. a the ovaries of rats following the microinjection of the vehicle into either SCN (Vh-SCN) in which several fresh corpora lutea can be observed and some antral follicles can be observed. b Ovaries of rats microinjected with atropine in either SCN (ATR-SCN); F, antral follicles; CL, corpora lutea. 4X microscopic lens, Scale Bar = 200 μm
Discussion
The present results suggest that the cholinergic neural information arriving to either side of the SCN at 09.00 h is necessary for the pre-ovulatory secretions of GnRH and LH that result in spontaneous ovulation in proestrus rats. This neural information does not appear to participate in the regulation of GnRH or LH secretion on the proestrus day at 19.00 h or during diestrus-1 at 09.00 or 19.00 h.
Funabashi et al. [40] demonstrated that the injection of pentobarbital on the morning of proestrus (8.00–09.00 h) does not interfere with the pulsatile secretion of LH. Domínguez and Smith [41] demonstrated that in 4- and 5-day cyclic rats on the proestrus day, the injection of pentobarbital at 09.00 h does not modify ovulation. A similar effect was observed in the present study. In rats on the proestrus day, the injection of barbiturates or atropine at 13.00 h blocks ovulation, and the injection GnRH, LH or human chorionic gonadotrophin (hCG) at 17.00 h (4 h later) restored ovulation, which suggests that the blocking agents (barbiturates or atropine) affect the spontaneous release of LH [41–43].
We have previously demonstrated that systemically blocking the cholinergic system via the subcutaneous injection of atropine sulphate blocks spontaneous ovulation on the predicted oestrus day in a manner that depends on the day and time of the oestrous cycle, which suggests that gonadotropin secretion throughout the oestrous cycle is controlled by a circadian rhythm and that this circadian rhythm is closely related to the cholinergic system [32].
We have previously demonstrated that the blockade of the cholinergic system via the implantation of atropine crystals at 13.00 h into either side of the POA-AHA during diestrus-1, into the right POA-AHA during oestrus, and into the left POA-AHA during diestrus-2 results in the lack of ovulation on the predicted day of oestrus. However, implantation on either side of the hypothalamus during pro-oestrus does not modify the rate of ovulation [24]. In the present study, the unilateral block at 09.00 h of the muscarinic receptors in the SCN of rats on the proestrus day partially blocked ovulation, whereas the same treatment had no effect in rats during diestrus-1. These results are the opposite of those observed when we blocked the cholinergic system in the POA-AHA region (i.e., complete blockade in diestrus-1 rats and a lack of effect in proestrus rats). Taken together, the present and previous results suggest that, during the first three days of the cycle, the POA-AHA regions participate in the regulation of GnRH secretion and ovulation, whereas the participation of the SCN’s cholinergic system occurs only during the beginning of the proestrus day.
In the adult rat, the SCN’s cholinergic afferents that originate in the NBM play an important role in maintaining the anatomy and chemo-architecture of the SCN [19, 44]. On the afternoon of proestrus, the AVP innervation arising from the SCN stimulates the pre-ovulatory LH surge [45]. Loh et al. [46] demonstrated that, in female VIP-knockout mice, the ovulatory response is reduced. In the present study, the ovulation blockade that resulted from the atropine treatment of the rats in proestrus at 09.00 h might have resulted from a decrease in the neural signals from the AVPergic and/or VIPergic neurons that regulate the GnRH neurons located in the POA. Moreover, this decrease would prevent the pre-ovulatory LH surge required for ovulation. The loss of VIP signalling can result in a decrease in reproduction [46].
According to Cruz et al. [47], blocking the muscarinic receptors in the POA-AHA during diestrus-2 or oestrus appears to be linked to the inhibition of ovarian follicular population growth in the ovary ipsilateral to the treated hypothalamic area. In another experiment, the blockade of M1R muscarinic receptors in the left ovary was observed to block ovulation, whereas this block had no effect when performed in the right ovary. Based in these results, the authors suggested that the M1R receptors in the left ovary might regulate ovulation asymmetrically through a stimulatory neural signal that is relayed to the hypothalamus via the vagus nerve and induces GnRH secretion, which then triggers ovulation [48].
Ovulation is a complex process that leads to the release of the mature oocyte from the ovarian follicle, induced by the LH surge [49]. Part of the events for LH-induced ovulation include the weakening of the follicle wall by proteolytic digestion, apoptotic follicle cell death, and follicle contraction. Overall, in mammals these coordinated events are crucial for follicle rupture and subsequent expulsion of the oocyte [50]. In proestrus day, acetylcholine binding to M1R in the follicular cells of the left ovary generates a neural signal involved in the regulation of GnRH secretion, which in turn, stimulates LH secretion, which is required for ovulation [48].
Although present results cannot rule out the possibility that the pre-ovulatory LH surge underwent a phase change (advance or delay), the presence of a few antral follicles and old corpora lutea in the ovaries of non-ovulating rats suggest that the low levels of LH measured at 17.00 h during proestrus were not sufficient to stimulate the final follicle maturation phase. Our interpretation is supported by the observed ovulation induced by injecting LHRH at 14.00 h to atropine treated rats during proestrus and by Cruz et al. [47] results showing that in rats on oestrus day, blocking the muscarinic receptors in the right POA-AHA resulted in a greater number of pre-ovulatory atretic follicles in the ipsilateral ovary, suggest that the lack of ovulation in rats with an atropine implant on the right POA-AHA was the result of an insufficient LH surge on proestrus. Based on these results we suggest that the cholinergic signal originating at the SCN regulates the follicle’s final maturation phase, perhaps through a cholinergic neuronal pathway.
Because the injection of synthetic LHRH into the rats that were treated with atropine in the right SCN induced ovulation, we propose that the cholinergic signal originating in the SCN is necessary for the regulation of GnRH and LH secretion. In proestrus rats, the injection of atropine into the left or right SCN at 09.00 h resulted in lower LH levels, which suggests that on the morning of proestrus, the cholinergic system innervating the SCN regulates gonadotropin secretion in a stimulatory manner. Moreover, the present results suggest that a pre-ovulatory LH surge shift was not produced. In vitro studies have demonstrated that carbachol (a nonspecific acetylcholine agonist) acts directly on the SCN to alter the phase of the rhythm of neuronal activity and that atropine and pirenzepine (a M1-selective antagonist) block carbachol’s phase-shifting effects [51]. Cruz et al. [52] reported that atropine-sulphate treated animals exhibit a 24-h delay in the LH preovulatory surge.
There is evidence that kisspeptin regulates GnRH secretion at two different levels, i.e., the kisspeptinergic signal originating from the AVPV that acts on the somata of GnRH neurons [53] and the arcuate kisspeptinergic neurons at the GnRH axons [54]. Because the SCN sends AVPergic information to both the AVPV and arcuate kisspeptinergic neurons, the present results suggest that the somata of the SCN cells that release AVP are stimulated by acetylcholine ligand binding on their muscarinic receptors. The differences in LH levels observed in the proestrus rats that were microinjected at 17.00 h with atropine into either SCN suggest that the cholinergic signals originating from the left and right SCNs modulate GnRH secretion from two different zones of the hypothalamus.
There is evidence that the regulation exerted by the hypothalamus on ovarian function exhibits presents asymmetry [55] and that this asymmetry varies during the oestrous cycle [24, 25]. Gerendai and Halász [55] suggested that the right side of the hypothalamus plays a dominant role in the control of reproductive functions. We have previously demonstrated that the right side of POA-AHA exhibits greater choline acetyltransferase activity during the day of oestrous, whereas this activity is greater in the left POA-AHAs of rats studied during diestrus-2 [56]. In the SCN, there is evidence of asymmetries, i.e., the asymmetrical patterns of activity from each side of the SCN support the hypothesis that independent oscillators exist on each side. This asymmetry could reflect a more general feature of brain organization, namely, functional lateralization [31]. According to Yang and Okamura [57] and Yan and Silver [58], the expressions of Per1 in the middle and rostral SCN regions are asymmetric. In adult male Syrian hamsters, exposure to constant light induces a splitting in their locomotor activity that results in asymmetry in the Per1 expression in the SCN [59]. In the present study, the differences in the capacities of atropine microinjections into the right and left SCNs at 09.00 h of proestrus to affect spontaneous ovulation (i.e., the ovulation rate) suggest that there are differences in the sensitivities of the muscarinic systems of each SCN. These results suggest that the right SCN seems to be more committed to the regulation of ovarian function. Although additional studies are required to clarify this idea, we cannot rule out the notion that the differences could be related to the numbers of muscarinic receptors in each SCN.
The SCN regulates the activity of the adrenal gland by acting on neurons that release corticotrophin-releasing hormone (CRH) and AVP [60, 61]. We have previously demonstrated that the adrenal glands are the main source of progesterone during the oestrous cycle in rats [26]. According to Kornya et al. [62], the cholinergic action on steroid production in human granulosa cells is mediated through the muscarinic route, and cholinergic neurotransmission may play a physiological role in intraovarian regulatory pathways. According to Cruz et al. [27], the participation of the cholinergic system in the regulation of ovarian oestradiol secretion varies throughout the oestrous cycle, and in unilaterally ovariectomized rats, this participation also depends on the in situ ovary.
In the present study, the changes in the progesterone and oestradiol levels in the rats that underwent unilateral atropine microinjection depended on the hormone measured, the time of day of the treatment, and the side of the SCN treated, which suggests that the neuroendocrine mechanisms that regulate hormone secretion exhibit asymmetry. Additionally, the regulations of progesterone and oestradiol secretions are modulated by SCN signals after pre-ovulatory LH release occurs.
Conclusions
Based on the present results, we propose that, for ovulation to occur, the system regulating GnRH and LH secretion requires a cholinergic signal from the left or right SCN on the morning (09.00 h) of proestrus. Together, the present results suggest that the cholinergic innervation of the SCN participates in the regulation of progesterone and oestradiol secretions and that this participation varies according to the hour and day of the cycle studied and exhibits functional asymmetry.
Abbreviations
ARC, arcuate nucleus; AVP, vasopressin; AVPV, anteroventral periventricular nucleus; GnRH, gonadotropin-releasing hormone; LDTg, latero-dorsal tegmental nucleus; LH, luteinizing hormone; NBM, nucleus basalis magnocellularis; POA, preoptical area; PPTg, pedunculopontine tegmental nucleus; RP3V, rostral periventricular area of the third ventricle; SCN, suprachiasmatic nucleus; VIP, vasoactive intestinal peptide
References
Urlep Z, Rozman D. The interplay between circadian system, cholesterol synthesis, and steroidogenesis affects various aspects of female reproduction. Front Endocrino (Lausanne). 2013;4:11.
Brown-Grant K, Raisman G. Abnormalities in reproductive function associated with the destruction of the suprachiasmatic nuclei in female rats. Proc R Soc Lond B Biol Sci. 1977;198(1132):279–96.
Samson WK, McCann SM. Effects of suprachiasmatic nucleus lesions on the hypothalamic LH-releasing hormone (LHRH) content and gonadotropin secretion in the ovariectomized (OVX) female rat. Brain Res Bull. 1979;4(6):783–8.
Wiegand SJ, Terasawa E. Discrete lesions reveal functional heterogeneity of suprachiasmatic structures in regulation of gonadotropin secretion in the female rat. Neuroendocrinology. 1982;34(6):395–404.
Ma YJ, Kelly MJ, Rönnekleiv OK. Pro-gonadotropin-releasing hormone (ProGnRH) and GnRH content in the preoptic area and the basal hypothalamus of anterior medial preoptic nucleus/suprachiasmatic nucleus-lesioned persistent estrous rats. Endocrinology. 1990;127(6):2654–64.
Ojeda SR, Lomniczi A, Mastronardi C, Heger S, Roth C, Paret AS, Matagne V, Mungenast AE. Minireview: the neuroendocrine regulation of puberty: is time ripe for a systems biology approach? Endocrinology. 2006;147(3):1166–74.
Dungan HM, Gottsch ML, Zeng H, Gragerov A, Bergmann JE, Vassilatis DK, Clifton DK, Steiner RA. The role of kisspeptin-GPR54 signaling in the tonic regulation and surge release of gonadotropin-releasing hormone/luteinizing hormone. J Neurosci. 2007;27(44):12088–95.
Clarkson J, d’Anglemont de Tassigny X, Moreno AS, Colledge WH, Herbison AE. Kisspeptin-GPR54 signaling is essential for preovulatory gonadotropin-releasing hormone neuron activation and the luteinizing hormone surge. J Neurosci. 2008;28(35):8691–7.
Clarkson J, Herbison AE. Oestrogen, kisspeptin, GPR54 and the pre-ovulatory luteinising hormone surge. J Neuroendocrinol. 2009;21(4):305–11.
Pinilla L, Aguilar E, Dieguez C, Millar RP, Tena-Sempere M. Kisspeptins and reproduction: physiological roles and regulatory mechanisms. Physiol Rev. 2012;92(3):1235–316.
Plant TM. 60 years of neuroendocrinology: The hypothalamo-pituitary–gonadal axis. J Endocrinol. 2015;226(2):T41–54.
Robertson JL, Clifton DK, de la Iglesia HO, Steiner RA, Kauffman AS. Circadian regulation of Kiss1 neurons: implications for timing the preovulatory gonadotropin-releasing hormone/luteinizing hormone surge. Endocrinology. 2009;150(8):3664–71.
Van der Beek EM, Wiegant VM, van der Donk HA, van den Hurk R, Buijs RM. Lesions of the suprachiasmatic nucleus indicate the presence of a direct vasoactive intestinal polypeptide-containing projection to gonadotrophin-releasing hormone neurons in the female rat. J Neuroendocrinol. 1993;5(2):137–44.
Tonsfeldt KJ, Chappell PE. Clocks on top: the role of the circadian clock in the hypothalamic and pituitary regulation of endocrine physiology. Mol Cell Endocrinol. 2012;349(1):3–12.
Vida B, Deli L, Hrabovszky E, Kalamatianos T, Caraty A, Coen CW, Liposits Z, Kalló I. Evidence for suprachiasmatic vasopressin neurones innervating kisspeptin neurones in the rostral periventricular area of the mouse brain: regulation by oestrogen. J Neuroendocrinol. 2010;22(9):1032–9.
Colledge WH. Kisspeptins and GnRH neuronal signalling. Trends Endocrinol Metab. 2009;20(3):115–21.
Bina KG, Rusak B, Semba K. Localization of cholinergic neurons in the forebrain and brainstem that project to the suprachiasmatic nucleus of the hypothalamus in rat. J Comp Neurol. 1993;335(2):295–307.
Yang JJ, Wang YT, Cheng PC, Kuo YJ, Huang RC. Cholinergic modulation of neuronal excitability in the rat suprachiasmatic nucleus. J Neurophysiol. 2010;103(3):1397–409.
Madeira MD, Pereira PA, Silva SM, Cadete-Leite A, Paula-Barbosa MM. Basal forebrain neurons modulate the synthesis and expression of neuropeptides in the rat suprachiasmatic nucleus. Neuroscience. 2004;125(4):889–901.
Hut RA, Van der Zee EA. The cholinergic system, circadian rhythmicity, and time memory. Behav Brain Res. 2011;221(2):466–80.
Silverman AJ, Livne I, Witkin JW. The gonadotrophin-releasing hormone (GnRH), neuronal systems: Immunocytochemistry and in situ hybridization. In: Neill JD, editor. Knobil and Neill’s physiology of reproduction. New York: Raven; 1994. p. 1683–706.
Turi GF, Liposits Z, Hrabovszky E. Cholinergic afferents to gonadotropin-releasing hormone neurons of the rat. Neurochem Int. 2008;52(4–5):723–8.
Everett JW, Sawyer CH, Markee JE. A neurogenic timing factor in control of the ovulatory discharge of luteinizing hormone in the cyclic rat. Endocrinology. 1949;44(3):234–50.
Cruz ME, Jaramillo LP, Domínguez R. Asymmetric ovulatory response induced by a unilateral implant of atropine in the anterior hypothalamus in the cyclic rat. J Endocrinol. 1989;123(3):437–9.
Cruz ME, Castro J, Domínguez R. A comparative analysis of the neuroendocrine mechanisms regulating ovulation, affected by a unilateral implant of atropine in the preoptic-anterior hypothalamic area, in intact and hemiovariectomized adult rats. J Endocrinol. 1992;133(2):205–10.
Flores A, Meléndez G, Palafox MT, Rodríguez JO, Barco AI, Chavira R, Domínguez R, Cruz ME. The participation of the cholinergic system in regulating progesterone secretion through the ovarian-adrenal crosstalk varies along the estrous cycle. Endocrine. 2005;28(2):145–51.
Cruz ME, Flores A, Palafox MT, Meléndez G, Rodríguez OJ, Chavira R, Domínguez R. The role of the muscarinic system in regulating estradiol secretion varies during the estrous cycle: the hemiovariectomized rat model. Reprod Biol Endocrinol. 2006;4:43.
Bock GR, Marsh J. Biological asymmetry and handedness. In: “CIBA Foundation symposium 162”, Toronto. Chichester/New York/Brisbane/Singapore: Wiley; 1991.
Michel S, Marek R, vanderleest HT, vansteensel MJ, Schwartz WJ, Colwell CS, Meijer JH. Mechanism of bilateral communication in the suprachiasmatic nucleus. Eur J Neurosci. 2013;37(6):964–71.
Pickard GE, Turek FW. Splitting of the circadian rhythm of activity is abolished by unilateral lesions of the suprachiasmatic nuclei. Science. 1982;215(4536):1119–21.
Zhang L, Aguilar-Roblero R. Asymmetrical electrical activity between the suprachiasmatic nuclei in vitro. Neuroreport. 1995;6(3):537–40.
Domínguez R, Riboni L, Zipitría D, Revilla R. Is there a cholinergic circadian rhythm throughout the oestrous cycle related to ovulation in the rat? J Endocrinol. 1982;95(2):175–80.
Zhao S, Kriegsfeld J. Daily changes in GT1–7 cell sensitivity to GnRH secretagogues that trigger ovulation. Neuroendocrinology. 2009;89:448–57.
Goodchild CS, Serrao JM, Kolosov A, Boyd BJ. Alphaxalone reformulated: a water-soluble intravenous anesthetic preparation in sulfobutyl-ether-β cyclodextrin. Anesth Analg. 2015;120(5):1025–31.
Paxinos G, Watson C. The rat brain in stereotaxic coordinates. 5th ed. London: Elsevier Academic; 2004.
Lohman RJ, Liu L, Morris M, O’Brien TJ. Validation of a method for localised microinjection of drugs into thalamic subregions in rats for epilepsy pharmacological studies. J Neurosci Methods. 2005;146(2):191–7.
Humphrey RR, Dermody WC, Brink HO, Bousley FG, Schottin NH, Sakowski R, Vaitkus JW, Veloso HT, Reel JR. Induction of luteinizing hormone (LH) release and ovulation in rats, hamsters, and rabbits by synthetic luteinizing hormone-releasing factor (LRF). Endocrinology. 1973;92(5):1515–26.
Derr RF. Pain perception in decapitated rat brain. Life Sci. 1991;49(19):1399–402.
Dhibi S, Ettaya A, Elfeki A, Hfaiedh N. Protective effects of Artemisia arborescens essential oil on oestroprogestative treatment induced hepatotoxicity. Nutr Res Pract. 2015;9(5):466–71.
Funabashi T, Jinnai K, Sano A, Shinohara K, Kimura F. Pentobarbital stimulates the activity of the GnRH pulse generator interacting with opioid neurons in rats in proestrus. Psychoneuroendocrinology. 2000;25:277–87.
Domínguez R, Smith ER. Barbiturate blockade of ovulation on days other than proestrus in the rat. Neuroendocrinology. 1974;14(3):212–23.
Weick RF, Smith ER, Domínguez R, Dhariwal AP, Davidson JM. Mechanism of stimulatory feedback effect of estradiol benzoate on the pituitary. Endocrinology. 1971;88(2):293–301.
Everett JW. “Pituitary and hypothalamus: perspectives and overview”. In: Neill JD, editor. Knobil and Neill’s physiology of reproduction. St. Louis: Elsevier Press; 2006. p. 1289–308.
Abbott SM, Arnold JM, Chang Q, Miao H, Ota N, Cecala C, Gold PE, Sweedler JV, Gillette MU. Signals from the brainstem sleep/wake centers regulate behavioral timing via the circadian clock. PLoS One. 2013;8(8):e70481.
Palm IF, Van Der Beek EM, Wiegant VM, Buijs RM, Kalsbeek A. Vasopressin induces a luteinizing hormone surge in ovariectomized, estradiol-treated rats with lesions of the suprachiasmatic nucleus. Neuroscience. 1999;93(2):659–66.
Loh DH, Kuljis DA, Azuma L, Wu Y, Truong D, Wang HB, Colwell CS. Disrupted reproduction, estrous cycle, and circadian rhythms in female mice deficient in vasoactive intestinal peptide. J Biol Rhythms. 2014;29(5):355–69.
Cruz ME, Flores A, Domínguez R. The cholinergic system of the preoptic-anterior hypothalamic areas regulates the ovarian follicular population in an asymmetric way. Endocrine. 2014;47(3):913–22.
Cruz ME, Flores A, Alvarado BE, Hernández CG, Zárate A, Chavira R, Cárdenas M, Arrieta-Cruz I, Gutiérrez-Juárez R. Ovulation requires the activation on proestrus of M1 muscarinic receptors in the left ovary. Endocrine. 2015;49(3):809–19.
Espey LL, Richards JS. Ovulation. In: Neil JD, editors. Physiology of Reproduction. Amsterdam: Academic Press; 2006. p. 425–74.
Crespo D, Goetz FW, Planas JV. Luteinizing hormone induces ovulation via tumor necrosis factor α-dependent increases in prostaglandin F2α in a nonmammalian vertebrate. Sci Rep. 2015;5:14210.
Liu C, Gillette MU. Cholinergic regulation of the suprachiasmatic nucleus circadian rhythm via a muscarinic mechanism at night. J Neurosci. 1996;16(2):744–51.
Cruz ME, Villegas G, Domínguez-González A, Chavira R, Domínguez R. Ovulation delay induced by blockade of the cholinergic system on dioestrus-1, is related to changes in dopaminergic activity of the preoptic anterior-hypothalamic area of the rat. Brain Res Bull. 2001;54(4):339–44.
Smarr BL, Morris E, de la Iglesia HO. The dorsomedial suprachiasmatic nucleus times circadian expression of Kiss1 and the luteinizing hormone surge. Endocrinology. 2012;153(6):2839–50.
d’Anglemont de Tassigny X, Fagg LA, Carlton MB, Colledge WH. Kisspeptin can stimulate gonadotropin-releasing hormone (GnRH) release by a direct action at GnRH nerve terminals. Endocrinology. 2008;149(8):3926–32.
Gerendai I, Halász B. Asymmetry of the neuroendocrine system. News Physiol Sci. 2001;16:92–5.
Sánchez MA, López-García JC, Cruz ME, Tapi R, Domínguez R. Asymmetrical changes in the choline acetyltransferase activity in the preoptic-anterior hypothalamic area during the oestrous cycle of the rat. Neuroreport. 1994;5(4):433–4.
Yan L, Okamura H. Gradients in the circadian expression of Per1 and Per2 genes in the rat suprachiasmatic nucleus. Eur J Neurosci. 2002;15(7):1153–62.
Yan L, Silver R. Day-length encoding through tonic photic effects in the retinorecipient SCN region. Eur J Neurosci. 2008;28(10):2108–15.
Mahoney CE, Brewer JM, Bittman EL. Central control of circadian phase in arousal-promoting neurons. PLoS One. 2013;8(6):e67173.
Buijs RM, Wortel J, Van Heerikhuize JJ, Feenstra MG, Ter Horst GJ, Romijn HJ, Kalsbeek A. Anatomical and functional demonstration of a multisynaptic suprachiasmatic nucleus adrenal (cortex) pathway. Eur J Neurosci. 1999;11(5):1535–44.
Chung S, Son GH, Kim K. Circadian rhythm of adrenal glucocorticoid: its regulation and clinical implications. Biochim Biophys Acta. 2011;1812(5):581–91.
Kornya L, Bódis J, Koppán M, Tinneberg HR, Török A. Modulatory effect of acetylcholine on gonadotropin-stimulated human granulosa cell steroid secretion. Gynecol Obstet Invest. 2001;52(2):104–7.
Acknowledgements
We thank the “Posgrado en Ciencias Biológicas, UNAM” and CONACyT for the support provided for this study. This work is a requirement for obtaining the degree of Doctor of Biological Sciences. We also thank M Sc A. Domínguez-González for the revision of the English manuscript.
Funding
The experimental work was supported by UNAM-DGAPA-PAPIIT No. IN217016; CONACyT Grant 236908.
Availability of data and materials
All datasets on which the conclusions of the paper rely are available to readers.
Authors’ contributions
EV, LML and RD planned the experiments. EV, DAR and NL performed the experiments. EV, LML, PDM, AT and RD participated in the analysis and discussion of the results. MC and RCH participated in the performance of the RIAs to measure the different hormones levels. All authors have read and approved the final version of the manuscript.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Ethics approval and consent to participate
All experiments were performed in strict accordance with the Mexican Law of Animal Treatment and Protection Guidelines and the specifications of the Mexican Official Standard NOM-062-ZOO-1999. The Institutional Committee of the Facultad de Estudios Superiores Zaragoza, Universidad Nacional Autónoma de México approved the experimental protocols (FES/DEPUCI/236/14). All efforts were made to minimize the number of animals used and their suffering.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Vieyra, E., Ramírez, D.A., Lagunas, N. et al. Unilaterally blocking the muscarinic receptors in the suprachiasmatic nucleus in proestrus rats prevents pre-ovulatory LH secretion and ovulation. Reprod Biol Endocrinol 14, 34 (2016). https://doi.org/10.1186/s12958-016-0168-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12958-016-0168-7