
Abstract
Background
Inflammatory breast cancer (IBC) represents a deadly aggressive phenotype of breast cancer (BC) with a unique clinicopathological presentation and low survival rate. In fact, obesity represents an important risk factor for BC. Although several studies have identified different cellular-derived and molecular factors involved in IBC progression, the role of adipocytes remains unclear. Cancer-associated adipose tissue (CAAT) expresses a variety of adipokines, which contribute to tumorigenesis and the regulation of cancer stem cell (CSC). This research investigated the potential effect of the secretome of CAAT explants from patients with BC on the progression and metastasis of the disease.
Methods
This study established an ex-vivo culture of CAAT excised from IBC (n = 13) vs. non-IBC (n = 31) patients with obesity and profiled their secretome using a cytokine antibody array. Furthermore, the quantitative PCR (qPCR) methodology was used to validate the levels of predominant cytokines at the transcript level after culture in a medium conditioned by CAAT. Moreover, the impact of the CAAT secretome on the expression of epithelial-mesenchymal transition (EMT) and cells with stem cell (CSC) markers was studied in the non-IBC MDA-MB-231 and the IBC SUM-149 cell lines. The statistical differences between variables were evaluated using the chi-squared test and unpaired a Student’s t-test.
Results
The results of cytokine array profiling revealed an overall significantly higher level of a panel of 28 cytokines secreted by the CAAT ex-vivo culture from IBC patients with obesity compared to those with non-IBC. Of note, interleukin-6 (IL-6), interleukin-8 (IL-8), and monocyte chemo-attractant protein 1 (MCP-1) were the major adipokines secreted by the CAAT IBC patients with obesity. Moreover, the qPCR results indicated a significant upregulation of the IL-6, IL-8, and MCP-1 mRNAs in CAAT ex-vivo culture of patients with IBC vs. those with non-IBC. Intriguingly, a qPCR data analysis showed that the CAAT secretome secretions from patients with non-IBC downregulated the mRNA levels of the CD24 CSC marker and of the epithelial marker E-cadherin in the non-IBC cell line. By contrast, E-cadherin was upregulated in the SUM-149 cell.
Conclusions
This study identified the overexpression of IL-6, IL-8, and MCP-1 as prognostic markers of CAAT from patients with IBC but not from those with non-IBC ; moreover, their upregulation might be associated with IBC aggressiveness via the regulation of CSC and EMT markers. This study proposed that targeting IL-6, IL-8, and MCP-1 may represent a therapeutic option that should be considered in the treatment of patients with IBC.
Similar content being viewed by others

Background
Breast cancer (BC) is the second leading cause of death among women globally. An estimated 1.7 million women have been diagnosed with BC, accounting for over 521,900 deaths worldwide [1, 2]. The number of BC-related death is expected to increase by 20% in the coming years, with over 95% of new cases being diagnosed over the age of 40 [3]. In Egypt, patients with BC present at a young age and have more aggressive properties than do patients in Western countries [4, 5].
Inflammatory breast cancer (IBC) is the most lethal subtype of BC [6,7,8]. IBC is diagnosed at a younger age compared with other types of BC [9]. Clinically, IBC is defined by distinct and painful clinical symptoms. For example, Patients with IBC have inflamed red or swollen breasts and a “peau d’orange” appearance [10,11,12]. However, IBC is still misdiagnosed as an infection [13].
Obesity is one of the strongest determinants of BC risk and survival rate [14]. Obesity is a condition in which the body’s fat levels are abnormally high and is defined as a body mass index (BMI) ≥ 30 kg/m2 [15, 16]. Obese patients with BC have a bigger tumor mass and a higher risk of lymphatic invasion as a consequence of obesity [17]; moreover, they have a poor prognosis and decreased overall survival compared with their lean counterparts [18,19,20]. Obesity promotes resistance to anticancer therapy in BC [21] by preventing the exposure of cancer cells to drugs, as reported previously [22, 23]. Moreover, the poor recurrence of BC may be adversely affected by obesity [24,25,26]. Because BC grows in the vicinity of adipose tissue (AT), AT dysfunction resulting from obesity is considered a critical determinant of cancer progression [27]. AT is a highly complex tissue that accounts for approximately 10% of the breast size [28, 29].
AT contains a heterogeneous population of cells, such as adipocytes, peri-adipocytes, adipose stromal/stem cells, fibroblasts, and endothelial cells [30,31,32]. As a result of excess calory intake, adipocytes begin to expand (hypertrophy), which in turn can induce an increase in adipose cell number (hyperplasia), to accommodate the excess energy [31], thus leading to obesity.
Typically, the principal function of AT is the storage of excess energy in the form of triglycerides [33, 34]. Moreover, it is a critical endocrine organ that expresses a multiplicity of determined biologically active molecules that control many systemic physiological processes [35, 36]. Collectively, adipokines or adipocytokines are the secretions of AT [36,37,38].
In BC, the alteration of the adipokine profile, including molecules such as IL-6, IL-8, and MCP-1, has a significant impact on cancer progression [2] as it promotes cancer proliferation and invasion, controls epithelial derived-proteins and the production of growth factors, and induces other cells of the tumor microenvironment (TME), to trigger invasion [35] via different mechanisms such as the induction of cancer stem cell-like traits and the modulation of the influencing epithelial–mesenchymal transition (EMT) [39]. The EMT, in which cancer cells lose their epithelial characteristics and acquire mesenchymal-like characteristics, is considered one to be of the essential steps in BC metastasis because it induces the acquisition of cells with stem cell (CSC) characteristics, which that can initiate new tumors [40].
In the present study, an ex-vivo culture of cancer-associated adipose tissue (CAAT) obtained from human patients with non-IBC and IBC during curative surgery was established, to mimic the in-vivo conditions of the TME. Moreover, to determine the predominant adipokines in this context, the mediators/adipokines secreted by the CAAT ex-vivo culture were profiled. Furthermore, their effect on the expression of CSC- and EMT-related genes in BC cell lines was studied.
Methods
Patient samples
For the purpose of patient enrolment in this study, Institutional Review Board (IRB#00006379) approval was obtained before surgical treatment. The experimental protocol was approved by the research ethics committee of the General Surgery Department, Faculty of Medicine, Ain Shams University, Cairo, Egypt. Patients who were preoperatively diagnosed as BC cases in the breast clinic of Ain Shams University Hospitals by clinical examination, mammography, ultrasound, and Tru-cut biopsy and declared their informed consent in written form prior to participation, including agreements for enrolment in this study and publication of data were enrolled in the study. Mammary AT was collected from 44 Egyptian BC patients with BMI ≥ 30 who underwent modified radical mastectomy or conservative surgery. The patients enrolled in this study were divided into two groups; the non-IBC (n = 31) and IBC (n = 13) groups who were selected for MRM surgical treatment. The clinical and pathological in the non-IBC and IBC groups who participated in the present study are listed in Table 1.
Sample collection and handling
AT was obtained from the safety margin, i.e., 3–5 cm away from the resected mass, and no cancer cells were detected by the pathologist. All tissue specimens were collected in a medium and immediately transported in an ice box from the operation room to the laboratory within 1 h, for further experiments.
Culture of cancer-associated adipose tissue (CAAT) and preparation of conditioned medium (CM)
The procedures for handling the tissue specimens were performed under aseptic conditions in the biological safety cabinet level II. CAAT (approximately 300 mg) specimens isolated from patients with non-IBC and IBC were washed with phosphate-buffered saline (PBS) twice and transferred to a 35 mm tissue culture dish containing a warm DMEM/F-12 medium supplemented with 10% fetal bovine serum (FBS) (Gibco, Waltham, Massachusetts, USA). Subsequently, the CAAT specimens were carefully minced with a sterile sharp scalpel into tiny pieces (approximately 0.5 mm in diameter) in a sterile 60 mm petri dish. The CAAT fragments were distributed equally, and a pre-warmed DMEM-F12 medium supplemented with 10% FBS was added and the dish was incubated for 24 h in a humidified atmosphere of 5% CO2 at 37°C. The following day, the medium was discarded, followed by starvation by adding 1 mL of serum-free culture medium and incubation for 24 h. Next, the CAAT-CM was collected and centrifuged at 800 xg at 4°C for 5 min, to separate it into the cell pellet and supernatant. The supernatant was collected and centrifuged at 2000 xg at 4°C for 10 min to eliminate cell debris, aliquoted, and stored at -80°C for cytokine profiling and conditioning breast cancer cell line culture medium. CAAT ex-vivo cultures were homogenized in TRI Reagent® (Sigma life science, Missouri, USA) to prepare the total RNA for real-time PCR experiments. Unless otherwise stated, all tissue culture medium were from Lonza (Basel, Switzerland), and supplies were from Greiner bio-one (Frickenhausen, Germany).
Profiling of non-IBC and IBC CAAT secretomes using a human cytokine array
The Ray-BioTM cytokine antibody array-C3 (RayBiotech, Inc. GA, USA) was used to detect 42 different cytokines and adipokines in the CAAT-CM of patients in non-IBC and IBC groups. Briefly, the secretome of CAAT ex-vivo culture was concentrated 10-fold using Amicon Ultracell 10K filters (Millipore, Billerica, MA). The kit components were allowed to equilibrate at room temperature (RT). As described in the kit manual, the cytokine array membrane was incubated with 2 mL of blocking buffer for 1 h at RT. Subsequently, the blocking buffer was discarded, and the membrane was incubated with 1 mL of CAAT-CM overnight at 4°C. After several washing steps, the membrane was incubated with a biotinylated antibody cocktail overnight at 4°C. Following the incubation with the primary antibody and washing steps, horseradish peroxidase-conjugated streptavidin was added overnight at 4°C. Finally, signals were developed using chemiluminescence reagents, and the cytokine spot signal intensity was evaluated by subtraction from the background and normalization to positive controls on the same membrane using the ImageJ software (NIH Bethesda, Maryland, USA).
Cell culture
The IBC cell line SUM-149, was a gift from Prof. Dr. Bonnie F. Sloane (Department of pharmacology, School of Medicine, Wayne State University, Detroit, 48201 MI, USA, and the non-IBC MDA-MB-231 cell line, was a gift from Prof. Dr. Martin Götte, a Münster University Hospital, Germany. SUM-149 cells were maintained in HAM’s-F12 medium containing 5% FBS, 1% penicillin/streptomycin, insulin, and hydrocortisone in a humidified atmosphere of 5% CO2 at 37°C. MDA-MB-231 cells were grown in a DMEM medium containing 10% FBS, 1% glutamine, and 1% penicillin/streptomycin in a humidified atmosphere of 5% CO2 at 37°C.
Cell viability assay
Using 96-well plates, MDA-MB-231 (3.0 X 103/well) and SUM-149 (5.0 X 103/well) cells were incubated with complete DMEM and HAM’s-F12 medium for 48 h. Cells were starved in a culture medium without FBS for 24 h. Then, MDA-MB-231 and SUM-149 cells were treated with DMEM and HAM’s-F12 medium supplemented with 1% FBS in the absence (control-CM) or presence of 25%, 50%, and 75% of CAAT-CM for an additional 24 h. Subsequently, MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium Bromide) was added, and the absorbance of treated cells compared with the control was measured at 570 nm using an Infinite®200 PRO NanoQuant instrument (Tecan, Zürich, Switzerland).
Stimulation of the breast cancer cell lines MDA-MB-231 and SUM-149 with CAAT-CM
The cells were plated and at 80% confluence, were washed with warm PBS. They were then starved for 24 h in a serum-free growth medium. Subsequently, the cells were seeded in medium containing 1% FBS (Gibco, Waltham, Massachusetts, USA) and 25% CAAT-CM from patients with non-IBC and IBC for 24 h, respectively. In parallel, control cells were seeded in DMEM and HAM’s-F-12 medium containing 1% FBS. Finally, cells were lysed to isolate total RNA. All tissue culture medium were from Lonza (Basel, Switzerland), and supplies were from Greiner Bio-One (Frickenhausen, Germany), unless otherwise stated.
RNA isolation and reverse transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA was isolated from cultured cells (MDA-MB-231 and SUM-149) and fresh CAAT ex-vivo cultures using the TRIzol reagent (Ambion, Life Technologies) according to the manufacturer’s guidlines. About 1 μg RNA was reverse-transcribed into cDNA using the high capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA, USA) according to the manufacturer’s instructions after measurement of the concentration using Infinite®200 PRO NanoQuant (Tecan, Zürich, Switzerland). Next, using SYBR Green universal master mix Kit (Applied Biosystems, Foster City, CA, USA), CD24, CD44, E-cadherin, vimentin, IL-6, Il-8, and MCP-1 gene expression levels were measured. The qPCR was performed in step-one plus amplification system (Applied Biosystems, San Francisco, CA, USA). Samples were incubated for initial denaturation at 95 °C for 10 min and then subjected to 40 PCR cycles as follows: 95°C for 15 sec, 55°-60°C for 1 min, followed by an amplification plot and melting curve to confirm specific product amplification. The ΔΔCt method was used to quantify gene fold expression and data normalized to 18S rRNA for cell lines [41,42,43] and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) for CAAT samples [44]. The selected reference genes are stable under our experimental conditions, as published before by the authors for both cell lines [45, 46].
Wound healing assay
The MDA-MB-231 and SUM-149 cells were cultivated in a 6-well tissue culture plate until their growth rate reached a confluence of ~ 80%. After that, the cells were washed with PBS, and the monolayer was scratched gently with a 200 μL pipette tip across the center of the well. After creating the scratched area, wells were washed twice with PBS to remove the detached cells and incubated in 25% CAAT-CM for 24 h. Cell migration to the scratch area was monitored under the Zeiss Axiophot bright-field microscope at 0 and 24 h.
Statistical analysis
All data are presented as the mean ± SEM except for the data in Additional file 1, which are presented as the mean ± SD as indicated. Data were analyzed using IBM SPSS version 15.0 for Windows (SPSS, Chicago, IL, USA). Differences among variables were evaluated using the chi-squared test and unpaired Student’s t-test. Pearson’s rank correlation test was used to analyze the correlations. The level of significance was set at P < 0.05.
Results
Establishment of a CAAT ex-vivo culture from patients with non-IBC and IBC
CAAT that was cultured as an ex-vivo culture to enable cells to be organized into structures mimicking their in-vivo conditions (Fig. 1A) showed increased secretion of lipid droplets (LDs) (Fig. 1B). A lipid-specific dye (Oil red O) is used to stain lipid droplets, and the Images are provided Supplemental Figure 1.

Photomicrographs representing ex-vivo culture CAAT immediately after surgery. A Representative microscopic images showing an ex-vivo culture of CAAT cultured in DMEM/F-12 medium supplemented with 10% FBS for 24 h in a humidified atmosphere of 5% CO2 at 37 °C. B Representative microscopic images focusing on the lipid droplets (LDs) secreted from CAAT after ex-vivo culture, scale bar 200 μm
IL-6, IL-8, and MCP-1 are elevated in the secretome of IBC vs. non-IBC-derived CAAT
To identify the predominant cytokines and chemokines secreted by AT, the secretome of CAAT was profiled using the Ray-BioTM cytokine antibody array-C3 (RayBiotech, Inc. GA, USA). As depicted in Supplemental Table 1, there was a significant increase in 28 secreted cytokines in the secretome of CAAT from IBC compared with non-IBC samples. Interestingly, relative to the non-IBC group, IL-6, IL-8, and MCP-1 were the predominant cytokines and chemokines in the secretome of CAAT from patients with IBC (P < 0.05 for all, Fig. 2).

profiling of cytokines/chemokines/growth factors secreted from CAAT of non-IBC patients vs. IBC patients. A Representative RayBioTM cytokine antibody array 3 of secreted CAAT-CM from IBC compared to non-IBC patients after 24 h. Boxes indicate the localization of cytokines that show a statistically significant difference between non-IBC patients in the left panel and IBC patients in the right panel. B Bars represent the mean ± SD of the signal intensity value of each cytokine secreted by an ex-vivo culture of CAAT of non-IBC (n = 10) and IBC (n = 10). The cytokine array image was quantified using ImageJ software. The Student’s t-test was used to determine the significant differences in secretion levels of adipokines /growth factors between IBC versus non-IBC patients. *Represents P < 0.05, **represents P ≤ 0.01, ***represents P ≤ 0.001, and # represents P ≤ 0.0001 as determined by Student’s t-test
The IL-6, IL-8, and MCP-1 mRNAs are upregulated in ex-vivo culture of CAAT isolated from non-IBC vs. IBC samples.
Using qRT-PCR, the expression of IL-6, IL-8, and MCP-1 was further verified at the transcript level in explants of CAAT from both the non-IBC and IBC groups. The qRT-PCR analysis showed a significant upregulation of IL-6 (4.17-fold, P = 0.004; Fig. 3A), IL-8 (3.46-fold, P = 0.0001; Fig. 3B), and MCP-1 (2.75-fold, P = 0.039; Fig. 3C) mRNAs in IBC vs. non-IBC samples.

qRT-PCR comparing the expression level of the predominant cytokines secreted by CAAT isolated from non-IBC versus IBC patients. A A significantly higher mRNA expression level of IL-6, (B) IL-8, and (C) MCP-1 of CAAT ex-vivo culture from patients with IBC vs. non-IBC by 4.17, 3.46, and 2.75 folds, respectively. ** P < 0.01, # P < 0.0001.* P < 0.05 as determined by unpaired Student t-test. Quantification of target genes mRNA was expressed relative to the housekeeping gene (GAPDH) mRNA. All bar graphs represent fold change+/−SEM (n of non-IBC = 21 and n of IBC = 11). D A significant up-regulation in IL-6 mRNA expression of obese IBC patients that had lymph node (LN) metastasis (n = 10, P = 0.006) in the right panel and no change in IL-6 mRNA expression of obese non-IBC patients that have LN metastasis (n = 17, P = 0.324) in the left panel was observed. E The transcript level of MCP-1 is significantly up-regulated in obese IBC patients that have LN metastasis (n = 10, P = 0.042) in the right panel, but not in obese non-IBC patients that have LN metastasis (n = 17, P = 0.146) in the left panel. F A significant down-regulation in IL-8 mRNA expression of obese non-IBC patients that express PR receptor (n = 21, P = 0.012) in the left panel but not in obese IBC patients that express PR receptor (n = 10, P = 0.816) in the right panel
Moreover, significant upregulation of the in IL-6 mRNA was detected (P = 0.006) in IBC patients with obesity who had lymph node (LN) metastasis (n = 10; Fig. 3D, the left panel) when compared with the non-IBC group (n = 17) for which its expression was not changed (P = 0.324; Fig. 3D, right panel). Similarly, significant upregulation of the MCP-1 mRNA (n = 10, P = 0.042) was observed in patients with obesity and IBC who had LN metastasis (Fig. 3E, left panel). Conversely, there was no difference in MCP-1 mRNA expression in patients with obesity and non-IBC who have LN metastasis (n = 17, P = 0.146; Fig. 3E, right panel). Furthermore, significant (P = 0.012) downregulation of the IL-8 mRNA was detected in patients with obesity and non-IBC who expressed the progesterone (PR) receptor (n = 21; Fig. 3F, left panel). However, there was no change in IL-8 mRNA in patients with obesity and IBC who expressed the PR receptor (n = 10, P = 0.816; Fig. 3F, right panel).
CAAT-CM effect on the viability of the MDA-231 and SUM-149 cell line
To determine the effect of medium conditioned by an ex-vivo culture of CAAT from patients with obesity in the non-IBC and IBC groups on the proliferation of MDA-231 and SUM-149 cells, respectively, cells were seeded in the absence (control-CM) or presence of 25%, 50%, and 75% CAAT-CM diluted with DMEM-F12 medium for 24 h. There was no change in the proliferation of MDA-231 and SUM-149 cells treated with 25% CAAT-CM compared with the control-CM (P = 0.555 and P = 0.98), respectively. Additionally, a significant decrease in the proliferation of SUM-149 cells was observed after incubation with 50% CAAT-CM (P = 0.039), whereas proliferation was not changed in MDA-231 cells treated with 50% CAAT-CM compared with control-CM. A significant decrease in the proliferation of MDA-231 cells was detected in the presence of 75% CAAT-CM (P = 0.003), whereas the proliferation of SUM-149 cells was not affected by the presence of 75% CAAT-CM as shown in Fig. 4.

CAAT-CM effect on MDA-231 and SUM-149 cells proliferation. MDA-MB-231 and SUM-149 cells were exposed to different concentrations (25%, 50%, and 75%) of CAAT-CM from obese non-IBC and IBC patients for 24 h vs. control cells. (A) No effect was observed in MDA-MB-231 cells growth when seeded in 25%, 50% CAAT-CM, and 75% CAAT-CM downregulated cell proliferation significantly. (B) The proliferation of SUM-149 cells was downregulated significantly by the presence of 50% CAAT-CM, but it was not affected by 25% or 50% CAAT-CM. MTT was used to assess the proliferation of cells. Bar graphs represent the old change ± SEM. **Represents P ≤ 0.01 as determined by the Student’s t-test
CAAT-CM of the non-IBC and IBC groups altered the expression of CSC and EMT markers in MDA-MB-231 and SUM-149 cells
Notably, cancer-associated adipocytes in the breast contribute to cancer growth by communicating with cancer cells by releasing adipokines [47]. MDA-MB-231 cells treated with non-IBC-CAAT-CM exhibited an elongated spindle-shaped morphology compared with untreated cells, whereas SUM-149 cells treated with IBC-CAAT-CM clustered into aggregates as observed in Fig. 5A and B, respectively.

CAAT-CM effect on BC cell morphological features, CSC, and EMT markers gene expression. Photomicrographs represent the morphological change of (A) MDA-MB-231 and (B) SUM-149 BC cells after treatment with 25 % CAAT-CM for 24 h. MDA-MB-231 cells showed elongated spindle-shaped morphology compared to untreated cells. In contrast, SUM-149 cell morphology clustered as aggregates. Cell morphology was observed by phase-contrast microscopy. Scale bar, 200 μm. (C) The mRNA expression of CD24 was significantly downregulated in the treated MDA231 with CAAT-CM compared with the control. D no change in the mRNA expression of CD44 was observed, E the mRNA expression of vimentin tended to be significantly upregulated in the treated MDA231 cells with CAAT-CM compared with the control by (3.08 fold), and F the mRNA expression of E-Cadherin was significantly downregulated in the treated MDA231 cells with CAAT-CM compared with the control. G The mRNA expression of CD24 tended to be significantly downregulated in the treated SUM149 cells, H No change in the mRNA expression of CD44 and I vimentin was observed, and J the mRNA expression of E-Cadherin was significantly upregulated in the treated SUM149 cells with CAAT-CM compared with the control by (2.38 fold). *P < 0.05 as determined by Student’s t-test. MDA-231 and SUM-149 cells were seeded in the absence (control) and presence of 25% CAAT-CM for 24 h (treated). Relative quantification of mRNA expression was analyzed using qRT-PCR (n of non-IBC = 18 and n of IBC = 9). Quantification of target genes mRNA was expressed relative to the housekeeping gene (18S) mRNA. The data shown are the mean ± SEM
The effect of the secretome of CAAT isolated from patients with obesity and BC was investigated on the mRNA expression levels of CSC markers, such as CD24 and CD44 as well as EMT markers, such as vimentin and E-cadherin, was investigated in MDA-231 and SUM-149 cells using qRT-PCR. The results showed that the CAAT-CM significantly downregulated the mRNAs of the surface marker CD24 (0.37-fold, P = 0.024; Fig. 5C) and epithelial E-cadherin marker (0.50-fold, P = 0.003; Fig. 5F) of MDA-231 cells upon stimulation with CAAT-CM from non-IBC groups. However, no change was observed in surface marker CD44 (1.45-fold, P = 0.371; Fig. 5D). Moreover, the vimentin mesenchymal marker tended to be significantly upregulated in MDA-231 upon stimulation (3.08-fold, P = 0.074; Fig. 5E). Moreover, the expression level of the mRNA for the surface CD24 marker tended to be significantly downregulated (0.76-fold, P = 0.098; Fig. 5G) upregulated in SUM-149 upon stimulation with CAAT-CM from IBC groups. In contrast, no changes were observed in the level of expression of the mRNAs for the CD44 (3.40-fold, P = 0.478; Fig. 5H) and vimentin (2.04-fold, P = 0.463; Fig. 5I) markers. Besides, the results showed that the epithelial E-cadherin marker was significantly upregulated (2.38-fold, P = 0.036; Fig. 5J).
CAAT-CM effect on the motility of the MDA-231 and SUM-149 cell lines
MDA-MB-231 cells showed a significant acceleration of scratch wound closure by the presence of 25% CAAT-CM compared with control (P = 0.004) as shown in Fig. 6C. Besides, SUM-149 cells showed a trend significance acceleration of scratch wound closure in response to 25% CAAT-CM compared with control (P = 0.087) as clear in Fig. 6D.

CAAT-CM effect on MDA-231 and SUM-149 cells motility. Representative photographs of (A) MDA-MB-231 and (B) SUM-149 BC cells in the wound healing assay at 0 hr and at 24 h. Scale bar, 200 μm. (C) The percentage of wound closure assay reveals a higher migratory potential of MDA-MB-231 cells compared with the control upon stimulation with 25% CAAT-CM. (D) The percentage of wound closure assay tended to show significance in SUM-149 cell migration compared with the control upon stimulation with 25% CAAT-CM. The area of wound closure was imaged at the indicated times and measured using the ImageJ software. Bar graphs represent a fold change ± SEM. **Represents P ≤ 0.01 as determined by the Student’s t-test
Discussion
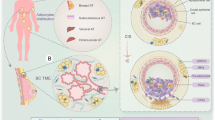
CAAT is an influential player in BC progression, as this type of cancer grows in the AT vicinity [27]. As a consequence, numerous studies have begun to provide insights into the role of AT in BC patients with obesity and its impact on BC migration and development [27, 35, 48, 49].
In fact, dysfunctional AT is associated with an increased risk for various cancers, including the esophageal adenocarcinoma, pancreas, colon, rectum, prostate, and ovarian cancer, as well as BC, in patients with obesity via the expression of a variety of pro-inflammatory adipocytokines and other factors that play a pivotal role in cancer metastasis [50]. Among these adipokines, IL-6, IL-8, and MCP-1 are released by adipocytes [50, 51]. In the present study, adipose tissues were isolated from patients with non-IBC and IBC and cultured ex-vivo; this experiment showed that adipocytokines are a potential candidate linking obesity with BC progression.
The present study aimed to profile the secretome of CAAT obtained during MRM as an ex-vivo patient tissue culture. The ex-vivo patient tissue culture provides mammary epithelial cells with a basement membrane-like gel, which enables cells to be organized into structures that mimick the in-vivo conditions and maintain their growth [52]. Moreover, cell–matrix interactions that modulate the cytoskeleton are enabled when cells grow as an ex-vivo culture [53]. Lastly, a growing body of evidence has suggested that the ex-vivo culture 3D model can preserve cell physiological functions to a greater extent than do those grown in 2D culture [54].
The aims of the present study also included the measurement of the major cytokines and chemokines in the culture medium of CAAT collected from obese non-IBC patients vs. obese IBC patients. The cytokine array showed a significantly higher level of a panel of 28 cytokines secreted by CAAT of obese IBC patients vs. non-IBC patients, with a more than three-fold change observed for the predominant cytokines IL-6, IL-8, and MCP-1. Furthermore, the levels of expression of the IL-6, IL-8, and MCP-1 mRNAs were upregulated in IBC- and non-IBC-derived CAAT. To our knowledge, the present study demonstrated for the first time that the CAAT of obese IBC groups had significantly higher levels of the IL-6, IL-8, and MCP-1 mRNAs and protein levels in CAAT culture medium compared with that of non-IBC obese groups. This result agrees with previous studies that demonstrated that the upregulation of IL-6, IL-8, and MCP-1 is correlated with a poor prognosis of BC in patients with obesity [55]. Regarding the patients, they had a poorer compared with those the non-IBC [9].
It is widely acknowledged that adipocytes play an influential role in the promotion of cancer growth and progression [56]. Additionally, it has been shown that the release of the IL-6 and IL-8 cytokines in metabolic disorders, such as obesity, is increased to mediate the crosstalk between adipocytes and BC cells within the tumor microenvironment [37]. Adipocytes become fertile soil for tumors in patients with, thus allowing them to spread distantly for distant and protecting them from therapeutic treatment [56].
IL-6 has been identified as a marker linked to increased lymph node in patients with BC [57]. Interestingly, a higher expression of IL-6 positively correlates with lymph node in obese IBC groups but not in non-IBC groups enrolled in the present study. These findings are in accordance with the study that highlighted that the IL-6 level in CAAT increases with increasing lymph node involvement [50]. In this regard, IL6 may contribute to the increased IBC aggressiveness and progression via stimulating aromatase expression in AT, thus stimulating estrogen synthesis [58].
Besides, the results analyses provide a significant positive correlation between the lymph node metastasis and MCP-1 transcription level in the obese IBC patients but not in non-IBC patients. This is in accordance with the study mentioned MCP-1 is associated with lymph node metastasis and tumor malignancy of the breast [59].
To extend these findings to in-vitro cultures and to understand better the effect of the secretome of CAAT on the expression of CSC markers such as CD24 and CD44 as well as EMT markers such as vimentin and E-cadherin, in both non-IBC (MDA-231) and IBC (SUM-149) cell lines, first, the best concentration of CAAT-CM was determined and shown not to be toxic to the cells using an MTT assay. The data revealed no changes in MDA-MB-231 cell growth was observed when seeded in 25% and 50% CAAT-CM, whereas 75% CAAT-CM was toxic to the BC cell line and decreased its viability. Additionally, there was no effect on SUM-149 cell proliferation in the presence of 25% and 75% CAAT-CM, whereas 50% CAAT-CM decreased cell proliferation. The present results disagree with other studies showing an absence of changes in the proliferation of MCF-10A, HBL100, MCF-7, and IBH-7 cells when incubated with 50 % and 75% ATCM compared with the control [2]. The differences between the present study and the previous study may be attributed to the different types of cell lines used here or the biological properties of Egyptian patients with BC compared with other patients populations. In this regard, we suggest that 25% CAAT-CM is the best concentration based on the data evaluation and that it is preferable to use the lowest amount of CAAT-CM to preserve the CAAT-CM of patients with BC for further experiments.
The data revealed that CAAT-CM significantly downregulated the mRNA for the CD24 surface marker in MDA-231 cells and tended to significantly downregulates it in SUM-149 cells. A pioneering study has indicated that CD24 is a significant marker linked to tumor metastasis, increased cell proliferation, motility, and invasiveness of abnormal fat accumulation in the body, and is associated with poor clinical outcomes in breast carcinomas [60].
Moreover, vimentin, which is a mesenchymal marker, tended to be significantly upregulated in MDA-231 cells upon stimulation, whereas no change was observed in SUM-149 cells. Such findings are consistent with other reports demonstrating that the co-culture of MDA-231 cells with tumor cells induces the EMT phenotype, which is characterized by the upregulation of the vimentin mesenchymal marker [61, 62]. Furthermore, the data agree with another study that indicated that vimentin expression is necessary for the migration and invasion of BC and that its blockage, in turn, triggers a decrease in BC cell migration and invasiveness [63]. These activities were mediated by PCKα/PP2A/C-SRC and JAK-STAT signaling pathway activation, resulting in STAT3 phosphorylation [64].
Furthermore, the present results revealed that the secretome of CAAT upregulated the E-cadherin epithelial marker in SUM-149 cells and downregulated it in MDA231 cells, which is the hallmark of tumor emboli formation in patients with IBC. This data agreed with another study carried out by our lab, which showed that E-cadherin was overexpressed in the IBC compared with the non-IBC group [13]. Additionally, such findings are consistent with the opposite role played by E-cadherin in patients with non-IBC and IBC. The increased tumor proliferation and metastasis observed in patients with non-IBC are attributed to the loss of E-cadherin expression [65], whereas the disease aggressiveness detected in patients with IBC is attributed to upregulation of the E-cadherin epithelial marker [66]. This may be ascribed to the role of the E-cadherin molecule in allowing carcinoma cells to migrate as a clump of cells that block blood vessels in different organs, causing organ failure and death, as mentioned in a previous study [67]. In this regard, the data indicate that CAAT-CM may play a role in the stem cell properties of cancer cells, thus potentially triggering cancer recurrence.
Study Strengths and Limitations
This study had the following strengths: (1) it was the first to culture CAATs obtained during MRM or curative surgery as an ex-vivo patient tissue culture, and (2) it allowed identification of the major cytokines in CAAT culture medium of obese IBC patients compared with those with non-IBC.
However, this study also had several limitations: (1) this intriguing observation requires further study to determine whether the prominent IL-6, IL-8, and MCP-1 adipokines affect CSCs by knockdown them, and future studies using larger patient cohorts will help define their total prognostic and predictive value in IBC, and (2) the interpretation of the results obtained for the IL-6, IL-8, and MCP-1 cytokines in correlation with BMI was limited because the patients were nearly obese due to the increased obesity rate in Egypt [68], which may contribute to BC. Therefore, only obese patients with BC were enrolled in the present study exclusively.
Conclusions
The study findings suggest that secretions from CAAT collected from the tumor margin of patients with obesity increased IBC aggressiveness by regulating the CSC and EMT phenotypes and are considered an important prognostic factor for BC recurrence. Hence, targeting IL-6, IL-8, and MCP-1 may represent a therapeutic option for obese IBC patients.
The study validates the usage of CAAT as an ex-vivo model that mimics in vivo patients’ tumor microenvironment to assess the key inflammatory meditators and study their effect on modulating gene expression of stem cell markers associated with cancer poor prognosis and relapse.
Herein, CAAT of tumor margins of obese IBC patients is characterized by the secretion of inflammatory mediator IL-6, IL-8, and MCP-1 that plays a significant role in proliferation, motility, invasion, induction of proteolytic activity, and drug resistance in the tumor microenvironment as described before by the authors (Mohamed et al., 2014 and 2021).
The ex-vivo model of CAAT can be used in preclinical studies for screening CAAT derived mediators (adipokines) in different types of cancer, not only breast cancer, and identify how CAAT plays a crucial in cancer relapse and metastasis in obese patients
Availability of data and materials
The data related to this study can be available from the corresponding author upon reasonable request.
Abbreviations
- IBC:
-
Inflammatory breast cancer
- BC:
-
Breast cancer
- CAAT:
-
Cancer-associated adipose tissue
- AT:
-
Adipose tissue
- CSCS :
-
cancer stem cells
- EMT:
-
Epithelial-mesenchymal transition
- BMI:
-
Body Mass Index
- ASCs:
-
Adipose stromal/stem cells
- TME:
-
Tumours Micronvironmental
- CMS :
-
Conditioned medium
- IL-6 :
-
Interleukin (IL)-6
- IL-8:
-
Interleukin (IL)-8
- MCP-1/ CCL2 :
-
monocyte chemoattractant protein 1
- GAPDH:
-
glyceraldehyde-3-phosphate dehydrogenase
- LD:
-
lipid droplet
- MTT:
-
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
- Vim:
-
vimentin
- MRM:
-
modified radical mastectomy
- PBS:
-
phosphate-buffered saline
References
De Silva S, Tennekoon KH, Karunanayake EH. Overview of the genetic basis toward early detection of breast cancer. Breast Cancer Targets Therapy. 2019;11:71–80.
Fletcher SJ, Sacca PA, Pistone-creydt M, Coló FA, Serra MF, Santino FE, et al. Human breast adipose tissue : characterization of factors that change during tumor progression in human breast cancer. J Exp Clin Cancer Res. 2017:1–13.
Saeg F, Anbalagan M. Breast cancer stem cells and the challenges of eradication: a review of novel therapies. 2018;5:1–9.
Harwansh RK, Deshmukh R. Breast cancer: An insight into its inflammatory, molecular, pathological and targeted facets with update on investigational drugs. Crit Rev Oncol Hematol [Internet]. 2020;154:103070. Available from: https://doi.org/https://doi.org/10.1016/j.critrevonc.2020.103070
Schlichting JA, Soliman AS, Schairer C, Harford JB, Hablas A, Ramadan M, et al. Breast Cancer by Age at Diagnosis in the Gharbiah, Egypt, Population-Based Registry Compared to the United States Surveillance, Epidemiology, and End Results Program, 2004-2008. Biomed Res Int. 2015;2015.
Nouh MA, Mohamed MM, El-Shinawi M, Shaalan MA, Cavallo-Medved D, Khaled HM, et al. Cathepsin b: A potential prognostic marker for inflammatory breast cancer. J Transl Med. 2011;9:1–8.
Mohamed HT, Untereiner V, Proult I, Ibrahim SA, Götte M, El-Shinawi M, et al. Characterization of inflammatory breast cancer: a vibrational microspectroscopy and imaging approach at the cellular and tissue level. Analyst. 2018;143:6103–12.
Fares M, Taha Mohamed H, Ibrahim SA, Hosney M, Rady MI, El-Shinawi M, et al. Incidence of Human Cytomegalovirus in Breast Carcinoma Tissues is Associated with A Higher Expression of Growth Factor Receptor-Bound Protein 2. Egypt J Hospital Med [Internet]. 2019;75:2358–65 Available from: https://ejhm.journals.ekb.eg/article_30754_b8d84e5d384b134ccd109242869610c9.pdf.
Barkataki S, Joglekar-Javadekar M, Bradfield P, Murphy T, Dickson-Witmer D, van Golen KL, et al. Inflammatory Breast Cancer: A Panoramic Overview. J Rare Dis ResTreatment. 2018;3:37–43.
Mohamed HT, Boffa MB, Mohamed MM, Cavallo-Medved D, El-Ghonaimy EA, Bazzi ZA, et al. IL-10 correlates with the expression of carboxypeptidase B2 and lymphovascular invasion in inflammatory breast cancer: The potential role of tumor infiltrated macrophages. Curr Probl Cancer. 2018;42:215–30.
Nouh MA, Mohamed MM, El-Shinawi M, Shaalan MA, Cavallo-Medved D, Khaled HM, et al. Cathepsin b: A potential prognostic marker for inflammatory breast cancer. J Transl Med [Internet]; 2011;9:1. Available from: http://www.translational-medicine.com/content/9/1/1
Huang A, Cao S, Tang L. The tumor microenvironment and inflammatory breast cancer. J Cancer. 2017;8:1884–91.
Mohamed MM. Inflammatory breast cancer : New factors contribute to disease etiology: A review. J Adv Res [Internet]. 2014;5:525–536. Available from: https://doi.org/10.1016/j.jare.2013.06.004
Laudisio D, Muscogiuri G, Barrea L, Savastano S, Colao A. Obesity and breast cancer in premenopausal women: Current evidence and future perspectives. Eur J Obstetr Gynecol Reprod Biol [Internet]. 2018;230:217–221. Available from: https://doi.org/10.1016/j.ejogrb.2018.03.050
Liu X, Sun Q, Hou H, Zhu K, Wang Q, Liu H, et al. The association between BMI and kidney cancer risk. 2018;
Jiang SHUZ, Lu WEN, Zong XUEF, Ruan HYUN, Liu YI. Obesity Hypertension (Review). 2016:2395–9.
Lee K, Kruper L, Dieli-Conwright CM, Mortimer JE. The Impact of Obesity on Breast Cancer Diagnosis and Treatment. Curr Oncol Rep. 2019;21:19–24.
Renehan AG, Soerjomataram I, Tyson M, Egger M, Zwahlen M, Coebergh JW, et al. Incident cancer burden attributable to excess body mass index in 30 European countries. Int J Cancer. 2010;702:692–702.
Protani M, Coory M, Martin JH. Effect of obesity on survival of women with breast cancer : systematic review and meta-analysis. Breast Cancer Res Treat. 2010:627–35.
Fuentes-Mattei E, Velazquez-Torres G, Phan L, Zhang F, Chou PC, Shin JH, et al. Effects of obesity on transcriptomic changes and cancer hallmarks in estrogen receptor-positive breast cancer. J Natl Cancer Inst. 2014;106.
Lengyel E, Makowski L, DiGiovanni J, Kolonin MG. Cancer as a Matter of Fat: The Crosstalk between Adipose Tissue and Tumors. Trend Cancer [Internet]. 2018;4:374–384. Available from: https://doi.org/10.1016/j.trecan.2018.03.004
Vaysse C, Muller C, Fallone F. Obesity: An heavyweight player in breast cancer’s chemoresistance. Oncotarget. 2019;10:3207–8.
Ewertz M, Jensen MB, Gunnarsdóttir KÁ, Højris I, Jakobsen EH, Nielsen D, et al. effect of obesity on prognosis after early-stage breast cancer. J Clin Oncol. 2011;29:25–31.
Emaus A, Bragelien M, Steinar V. Metabolic profile, physical activity, and mortality in breast cancer patients. Breast Cancer Res Treat. 2010;121:651–60.
Vaysse C, Lomo J, Garred O, Fjeldheim F, Lofteroed T, Schlichting E, et al. Inflammation of mammary adipose tissue occurs in overweight and obese patients exhibiting early-stage breast cancer. NPJ Breast Cancer [Internet]. 2017;3:1–9. Available from: https://doi.org/10.1038/s41523-017-0015-9
DSM C, Vieira AR, Aune D, Bandera EV, Greenwood DC, Mctiernan A, et al. Body mass index and survival in women with breast cancer-systematic literature review and meta-analysis of 82 follow-up studies. Ann Oncol. 2014;25:1901–14.
Wu Q, Li B, Li Z, Li J, Sun S, Sun S. Cancer-associated adipocytes: Key players in breast cancer progression. J Hematol Oncol. 2019;12:1–15.
Lorincz AM, Sukumar S. Molecular links between obesity and breast cancer. Endocr Relat Cancer. 2006;13:279–92.
Zhu W, Nelson CM. Adipose and mammary epithelial tissue engineering. Biomatter. 2013;3:1–6.
Kocan B, Maziarz A, Tabarkiewicz J, Ochiya T, Banaś-Ząbczyk A. Trophic Activity and Phenotype of Adipose Tissue-Derived Mesenchymal Stem Cells as a Background of Their Regenerative Potential. Stem Cells Int. 2017;2017:1653254.
Sethi JK, Vidal-Puig AJ. Thematic review series: Adipocyte Biology. Adipose tissue function and plasticity orchestrate nutritional adaptation. J Lipid Res. 2007;48:1253–62.
Schäffler A, Büchler C. Concise Review: Adipose Tissue-Derived Stromal Cells-Basic and Clinical Implications for Novel Cell-Based Therapies. Stem Cells. 2007;25:818–27.
Zoico E, Rubele S, De Caro A, Nori N, Mazzali G, Fantin F, et al. Brown and beige adipose tissue and aging. Front Endocrinol. 2019;10:1–10.
Fenzl A, Kiefer FW. Brown adipose tissue and thermogenesis. Horm Mol Biol Clin Invest. 2014;19:25–37.
Jardé T, Perrier S, Vasson MP, Caldefie-Chézet F. Molecular mechanisms of leptin and adiponectin in breast cancer. Eur J Cancer. 2011;47:33–43.
Kajimura S, Saito M. NIH Public Access. 2014;225–49.
Fain JN, Madan AK, Hiler ML, Cheema P, Bahouth SW. Comparison of the release of adipokines by adipose tissue, adipose tissue matrix, and adipocytes from visceral and subcutaneous abdominal adipose tissues of obese humans. Endocrinology. 2004;145:2273–82.
Berg AH, Scherer PE. Adipose tissue, inflammation, and cardiovascular disease. Circ Res. 2005;96:939–49.
Liu C, Zhao Q, Yu X. Bone Marrow Adipocytes, Adipocytokines, and Breast Cancer Cells: Novel Implications in Bone Metastasis of Breast Cancer. Front Oncol. 2020;10:1–15.
Avtanski D, Garcia A, Caraballo B, Thangeswaran P, Marin S, Bianco J, et al. Cytokine Resistin induces breast cancer cells epithelial to mesenchymal transition ( EMT ) and stemness through both adenylyl cyclase-associated protein 1 ( CAP1 ) -dependent and CAP1-independent mechanisms. Cytokine [Internet]. 2019;120:155–164. Available from: https://doi.org/10.1016/j.cyto.2019.04.016
Ogino T, Matsunaga N, Tanaka T, Tanihara T, Terajima H, Yoshitane H, et al. Post-transcriptional repression of circadian component clock regulates cancer-stemness in murine breast cancer cells. Elife. 2021;10:1–26.
Tarek A, El-Sayed SK, Woodward WA, El-Shinawi M, Hirshon JM, Mohamed MM. Inflammatory Breast Cancer: The Cytokinome of Post-Mastectomy Wound Fluid Augments Proliferation, Invasion, and Stem Cell Markers. Curr Issues Mol Biol. 2022;44:2730–44.
Wanandi SI, Syahrani RA, Arumsari S, Wideani G, Hardiany NS. Profiling of gene expression associated with stemness and aggressiveness of ALDH1A1-expressing human breast cancer cells. Malaysian J Med Sci. 2019;26:38–52
Piruzyan M, Shitanda I, Shimauchi Y, Okita G, Tsurekawa Y, Moriuchi M, et al. A novel condition of mild electrical stimulation exerts immunosuppression via hydrogen peroxide production that controls multiple signaling pathway. Public Library of Science; PLoS One. 2020;15.
Ibrahim SA, Gadalla R, El-Ghonaimy EA, Samir O, Mohamed HT, Hassan H, et al. Syndecan-1 is a novel molecular marker for triple negative inflammatory breast cancer and modulates the cancer stem cell phenotype via the IL-6/STAT3, Notch and EGFR signaling pathways. Mol Cancer. 2017;16:57.
Fares M, Mohamed HT, Ibrahim SA, Hosney M, Rady M, El-Shinawi M, et al. Incidence of Human Cytomegalovirus in Breast Carcinoma Tissues is Associated with A Higher Expression of Growth Factor Receptor-Bound Protein 2. Egypt J Hosp Med. 2019;75:2358–65.
D’Esposito V, Ambrosio MR, Giuliano M, Cabaro S, Miele C, Beguinot F, et al. Mammary Adipose Tissue Control of Breast Cancer Progression: Impact of Obesity and Diabetes. Front Oncol. 2020;10:1–8.
Gyamfi J, Lee YH, Eom M, Choi J. Interleukin-6/STAT3 signalling regulates adipocyte induced epithelial-mesenchymal transition in breast cancer cells. Scientific Reports [Internet]. 2018;8:1–13. Available from: https://doi.org/10.1038/s41598-018-27184-9
Cozzo AJ, Fuller AM, Makowski L. Contribution of adipose tissue to development of cancer. Compr Physiol. 2018;8:237–82.
Picon-ruiz M, Morata-tarifa C, Valle-goffin JJ. Take free quizzes online at acsjournals . com / ce Obesity and Adverse Breast Cancer Risk and Outcome : Mechanistic Insights and Strategies for Intervention. 2017;67.
Fouad TM, Kogawa T, Liu DD, Shen Y, Masuda H, El-Zein R, et al. HHS Public Access. 2016;152:407–16.
Padmanaban V, Grasset EM, Neumann NM, Fraser AK, Henriet E, Matsui W, et al. Organotypic culture assays for murine and human primary and metastatic-site tumors. Nature Protocols [Internet]. 2020;15:2413–2442. Available from: https://doi.org/10.1038/s41596-020-0335-3
Mohamed MM. Monocytes conditioned media stimulate fibronectin expression and spreading of inflammatory breast cancer cells in three- dimensional culture: A mechanism mediated by IL-8 signaling pathway. Cell Commun Signal. 2012;10:1–13.
Saji Joseph J, Tebogo Malindisa S, Ntwasa M. Two-Dimensional (2D) and Three-Dimensional (3D) Cell Culturing in Drug Discovery. Cell Cult. 2019;21-42.
Cho YA, Sung MK, Yeon JY, Ro J, Kim J. Prognostic role of interleukin-6, interleukin-8, and leptin levels according to breast cancer subtype. Cancer Res Treat. 2013;45:210–9.
Bandini E, Rossi T, Gallerani G, Fabbri F. Adipocytes and microRNAs Crosstalk: A Key Tile in the Mosaic of. Breast Cancer Microenviron. 2019.
Ibrahim SA, El-ghonaimy EA, Hassan H, Mahana N, Abdelbaky M, El-mamlouk T, et al. Hormonal-receptor positive breast cancer: IL-6 augments invasion and lymph node metastasis via stimulating cathepsin B expression. J Adv Res [Internet]. 2016;7:661–670. Available from: https://doi.org/10.1016/j.jare.2016.06.007.
Ibrahim SA, El-Ghonaimy EA, Hassan H, Mahana N, Mahmoud MA, El-Mamlouk T, et al. Hormonal-receptor positive breast cancer: IL-6 augments invasion and lymph node metastasis via stimulating cathepsin B expression. J Adv Res [Internet]. 2016;7:661–670. Available from: https://doi.org/10.1016/j.jare.2016.06.007.
Wang J, Zhuang ZG, Xu SF, He Q, Shao YG, Ji M, et al. Expression of CCL2 is significantly different in five breast cancer genotypes and predicts patient outcome. Int J Clin Exp Med. 2015;8:15684–91.
Chen X, Jung S, University G, Korea Qi Zhao S, Zheng P, Liu Y, et al. Amplification of the CD24 Gene Is an Independent Predictor for Poor Prognosis of Breast Cancer. 2019; Available from: https://portal.gdc.cancer.gov/,
Wei X, Li S, He J, Du H, Liu Y, Yu W, et al. Tumor-secreted PAI-1 promotes breast cancer metastasis via the induction of adipocyte-derived collagen remodeling. Cell Communication and Signaling. Cell Commun Signal. 2019;17:1–18.
Bochet L, Dabek M, Majed B, Wang YY, Meulle A, Salles B, et al. Cancer-Associated Adipocytes Exhibit an Activated Phenotype and Contribute to Breast Cancer Invasion. Cancer Res. 2011:2455–66.
Tanaka K, Tokunaga E, Inoue Y, Yamashita N, Saeki H, Okano S, et al. Impact of Expression of Vimentin and Axl in Breast Cancer. Clin Breast Cancer [Internet]. 2016;16:520-526.e2. Available from: https://doi.org/10.1016/j.clbc.2016.06.015
Li J, Han X. Adipocytokines and breast cancer. Curr Prob Cancer [Internet]. 2018;42:208–214. Available from: https://doi.org/10.1016/j.currproblcancer.2018.01.004
Kowalski PJ, Rubin MA, Kleer CG. E-cadherin expression in primary carcinomas of the breast and its distant metastases. Breast Cancer Res. 2003;5:217–22.
Kleer CG, Van GKL, Ph D, Braun T, Ph D, Merajver SD, et al. Persistent E-Cadherin Expression in Inflammatory. Breast Cancer. 2001.
Elisha Y, Kalchenko V, Kuznetsov Y, Geiger B. Dual role of E-cadherin in the regulation of invasive collective migration of mammary carcinoma cells. Scientific Reports [Internet]. 2018;8:1–15. Available from: https://doi.org/10.1038/s41598-018-22940-3.
Zaki ME, Basha W, Yousef RN, Awad M. Serum Vascular Endothelial Growth Factor in Egyptian Obese Women with Insulin Resistance. Open Access Maced J Med Sci. 2019;7:1330–4.
Acknowledgements
This work was conducted at Cancer Biology Research Laboratory (CBRL), Department of Zoology, Faculty of Science, Cairo University, Giza, Egypt and Galala University Labs, Suez, Egypt. We thank Dr. Hossam Taha Mohamed, Research Visitor, CBRL and Assistant professor, Faculty of Biotechnology, October University for Modern Sciences and Arts (MSA). The authors are supported by the Academy of Scientific Research and Technology (ASRT), Egypt, APPLE Call Grant Number 10142021205540 (PI: MMM, CO-PI: MES).
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB). This paper received no funding that supports the publication fee.
Author information
Authors and Affiliations
Contributions
MMM formulated the research idea and designed the research strategy and experimental protocols with the support of MES and SS. MMM, SAI, and SS supervised all study experiments. MMM critically reviewed the manuscript and response to reviewer comments with the support of SS. Surgeon MES was responsible for enrolling patients. AS collected patients’ clinical and conducted all practical experiments of the study. AS analysed the data using Statistical Package of the Social Sciences and biomedical informatics software. AS, MMM, SS, SAI. MES. Drafted and wrote the manuscript with the input of all co-authors. The author(s) read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethical approval and consent to participate
Institutional Review Board (IRB#00006379) approval was obtained before surgical treatment, and the protocol was approved by the research ethics committee of the General Surgery Department, Faculty of Medicine, Ain Shams University, Cairo, Egypt
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1:
Supplemental Figure 1. Photomicrographs representing ex-vivo cultured CAAT and secreted lipid droplets stained with oil red O.
Additional file 2:
Supplemental Table S1. A statistical difference in CAAT secretome levels of IBC vs. non-IBC patients.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Ibrahim, A.S., El-Shinawi, M., Sabet, S. et al. Role of adipose tissue-derived cytokines in the progression of inflammatory breast cancer in patients with obesity. Lipids Health Dis 21, 67 (2022). https://doi.org/10.1186/s12944-022-01678-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12944-022-01678-y