
Abstract
Cancer progression is continuously controlled by the immune system which can identify and destroy nascent tumor cells or inhibit metastatic spreading. However, the immune system and its deregulated activity in the tumor microenvironment can also promote tumor progression favoring the outgrowth of cancers capable of escaping immune control, in a process termed cancer immunoediting. This process, which has been classified into three phases, i.e. “elimination”, “equilibrium” and “escape”, is influenced by several cancer- and microenvironment-dependent factors. Senescence is a cellular program primed by cells in response to different pathophysiological stimuli, which is based on long-lasting cell cycle arrest and the secretion of numerous bioactive and inflammatory molecules. Because of this, cellular senescence is a potent immunomodulatory factor promptly recruiting immune cells and actively promoting tissue remodeling. In the context of cancer, these functions can lead to both cancer immunosurveillance and immunosuppression. In this review, the authors will discuss the role of senescence in cancer immunoediting, highlighting its context- and timing-dependent effects on the different three phases, describing how senescent cells promote immune cell recruitment for cancer cell elimination or sustain tumor microenvironment inflammation for immune escape. A potential contribution of senescent cells in cancer dormancy, as a mechanism of therapy resistance and cancer relapse, will be discussed with the final objective to unravel the immunotherapeutic implications of senescence modulation in cancer.
Similar content being viewed by others

Background
There are several factors influencing tumor progression, including genetic alterations, epigenetic modifications, oncogenic signals, and inflammatory cytokines. Among these, the contribution of the immune response on cancer cell fate is crucial, playing a dual role potentially limiting or, paradoxically, promoting tumor growth [1, 2].
In 1909, Paul Ehrlich postulated the concept that the immune system can recognize and eliminate cancer cells (Ned Tijdschr Geneeskd. 1909;5:273–290) and in the late 1950s Burnet and Thomas further expanded this concept with the immunosurveillance theory of cancer, hypothesizing that lymphocytes act as sentinels capable of recognizing and eliminating nascent tumor cells before they manifested disease [3] (L Thomas, H Lawrence - New York: Hoeber-Harper, 1959 - IET). However, the concept of cancer immunosurveillance as currently understood, was developed by Robert Schreiber who formulated the theory of cancer immunoediting [1]. This not only incorporates the original notion of cancer immunosurveillance but also recognizes that even after immunosurveillance escape, a tumor phenotype is continuously sculpted by the immune system. According to this view, cancer immunoediting is a multi-step process consisting in three phases: “Elimination”, “Equilibrium” and “Escape”, also referred to as the three “Es”.
Considering the numerous biological and genetic mechanisms that regulate aging, herein we focus on one of the key processes of aging — cellular senescence — which links aging and cancer together. Senescent cells (SnCs) accumulate during aging, contributing to tissue dysfunction and different pathological conditions, as well as playing a role in tumorigenesis and cancer progression. It has been described that the elimination of SnCs can potentially reduce age-related pathologies, including cancer [4]. In healthy individuals, activation of the immune system can rapidly clear SnC accumulation following induction, strongly decreasing their detrimental activity in tissues [4, 5]. However, during cancer progression or in older individuals, this control is less efficient or partially inhibited, leading to the accumulation of SnCs [6]. The impact of senescence in the regulation of anti-cancer immune responses is only beginning to be elucidated. In this regard, induction of senescence has been correlated with tumor suppression by inhibiting tumorigenesis of pre-neoplastic cells. On the other hand, SnCs have also been implicated as an active driving force to cancer progression, by sustaining immune-related hallmarks of cancer, such as inflammation and immune evasion. In this review, we will discuss how senescence, in response to various intrinsic and extrinsic stimuli, can regulate cancer development through a journey in the different steps of cancer immunoediting.
Senescence at a glance: definition, markers and SASP
The word senescence derives from the Latin word “senex” meaning “growing old”. Indeed, senescence was originally described in 1961 as the cellular program based on cell cycle arrest distinguishing human diploid fibroblasts after long-term in vitro culture [7]. Today, senescence is seen as a hallmark of aging and cancer [8, 9]. Cells undergoing senescence display distinctive morphological and functional alterations, but without unique molecular markers. They are characterized by a deep proliferative arrest (upregulation of cyclin-dependent kinase inhibitors p16 and/or p21), changes in cell morphology (size enlargement and perinuclear lamin-B1 downmodulation), epigenetic modifications (senescence-associated heterochromatin foci), and activation of specific signaling pathways (DNA damage response (DDR) and PI3K/FOXO/mTOR). The expansion of the lysosomal compartment accounts for the senescence-associated beta-galactosidase (SA-β-Gal) activity and represents the most reliable cellular marker of senescence along with lipofuscin accumulation [10]. Throughout life, several intrinsic and extrinsic injuries, via different pathways, trigger senescence (Fig. 1A).
Oncogene-induced senescence (OIS) following aberrant hyperproliferation, and chemotherapy- and radiotherapy- induced senescence (TIS), have long been described as key components in tumor biology [11]. However, how and to what extent cellular senescence affects cancer is still unclear. SnCs exhibit robust metabolic reprogramming, leading to a peculiar senescence-associated secretory phenotype (SASP), which consists of the release of bioactive molecules and inflammatory factors into the surrounding microenvironment resulting in immune cell recruitment and orchestration of tissue homeostasis [12] (Fig. 1B). It is reasonable to assume that senescence developed as a cellular stress response to disarm and shut down cell division, in due course removing damaged cells by alerting the immune system [13].
SASP composition is heterogeneous and highly dynamic. It is cell-type specific, trigger-dependent and changes over time, with successive stages of factor secretion. Nevertheless, a core group of molecules can be identified by an overlapping presence of Angiogenin, CCL2, GM-CSF, GRO-α, -β, -γ, ICAM-1, IGFBPs, IL-1, IL-6, CXCL8, MMP-1, MMP-3, MMP-10, Osteoprotegerin [14]. Recently the extracellular vesicles have been included as key mediators within SASP [15] and contribute to modulate the phenotype of recipient cells resulting in inflammation, cancer and immune cell modulation, and paracrine senescence induction [16,17,18,19,20,21,22].
Collectively, these factors characterize a troop of SnCs aimed at tissue remodeling, bolstered through the immune system, with inevitable repercussions on tumor progression. An additional layer of complexity is also related to the induction of a stromal SASP that can interact with tumor cells and may be involved in tumor initiation and progression, although how it potentially affects immune cells is still unclear [23,24,25].
The NF-kB and C/EBPβ transcription factors are the main drivers for the expression of SASP-related genes, with the transcription factor GATA4 also implicated [26]. In addition, the discovery that SnCs lose nuclear envelope integrity, leading to the production of cytoplasmic chromatin fragments (CCFs), links SASP induction to the cGAS-STING pathway as it will be extensively discussed in a separate section below.

Senescence inducers and paracrine effects of SnCs
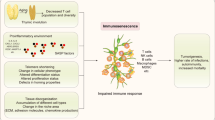
Overview of senescence inducers and paracrine effects of SnCs on the surrounding microenvironment. The senescence program can be activated by different stress stimuli (shown on the left) such as cytotoxic chemotherapeutic drugs, replicative and oxidative stress. On the other hand, SASP factors produce paracrine effects impinging on cancer onset and progression (shown on the right). SASP can induce senescence of surrounding cells (paracrine senescence), facilitate tissue remodeling and recruitment of immune cells
Created with BioRender.com with granted permission and license
Senescence-driven immune surveillance: the “elimination” phase
The elimination phase is classically regarded as a bidirectional crosstalk between pre-malignant/malignant cells and cells of the immune system in which the pressure of the immune system is successful at halting cancer cells (Fig. 2A). This phase is considered largely dependent on the mutational burden of tumor cells, with the expression of tumor neoantigens (TNAs). Much relevance is given to the immunogenicity of TNAs and its associated immune surveillance (be it natural or therapy-elicited) [27]. This straightforward view is now moving towards a more complex picture, in which different cancer cell phenotypes, such as stem- and dormant-like phenotypes, along with several elements of the tumor microenvironment (TME) heterogeneously affect antitumor innate and adaptive immunity. In this scenario, cellular senescence has recently been proposed as an emerging hallmark of cancer [9] and it acts as a double-edged sword showing opposing effects on tumor progression [28]. Regarding its contribution to the elimination of cancer cells, it should be noted that the senescent phenotype can occur at different times and in different cell types of tumor stages. OIS characterizes premalignant cells and is a major barrier to tumorigenesis. It restrains tumor growth by arresting the proliferation of cells at risk of malignant transformation and, at the same time, promotes the immune-mediated clearance of dangerous cells. TIS is an alternative cellular fate to other forms of regulated cell death triggered by conventional chemotherapy, radiation, and molecularly targeted agents. Thus, TIS commonly comes when cancer is clinically evident, a stage of tumor usually ascribed to the equilibrium or escape phases. Moreover, TIS can affect cancer cells as well as cells of the TME, such as stromal and endothelial cells, with different biological outcomes. The impact of these senescence-related temporal and spatial differences is not fully understood. Data from literature support the notion that cellular senescence can promote the elimination phase by at least three mechanisms: (i) immune cell recruitment; (ii) direct killing enhancement; (iii) antigenicity/adjuvanticity properties.
i) immune cell recruitment. The first evidence of a role of senescence to cancer immunosurveillance through immune cell recruitment came from groundbreaking work by Scott Lowe and colleagues. By treating lymphoma with cyclophosphamide, a drug usually reported to induce apoptosis, they showed that cellular senescence developed in vivo following chemotherapy [28]. Moving on, investigating a genetically modified mouse model, they provided direct evidence that cellular senescence triggered by p53 restoration in liver carcinoma induced by the oncogenic H-RAS expression, resulted in tumor regression due to an innate immune response against senescent tumor cells. This response was guided by macrophages and Natural Killer (NK) cells migrated into the tumor bed following the release of CCL2 (previously called MCP1), CXCL1, and IL-15 [29]. Similar findings were reported by others in sarcomas [30]. Thus, p53 controls molecular programs central for senescence, whilst complementary pathways mainly regulated by NF-κB contributes to the SASP, which provides a plethora of inflammatory factors e.g., IL-6, CXCL8, CXCL1, and ICAM1 necessary for the recruitment and activation of immune cells [31]. Importantly, wild-type p53 has been described to interact with NF-κB both in a cooperative and also antagonistic manner [32]. The final outcome can be context-dependent and modulated by factors such as IKKα activity [33] or the common TP53 codon 72 polymorphism [34]. In this scenario, it is not surprising that the alterations of p53 activity caused by different TP53 mutations can result in a dysregulated senescence and tumor secretome, affecting the complex crosstalk between tumor cells and a plethora of cell types in the surrounding stromal microenvironment. Indeed, mutant p53 activities are often dependent on other transcription factors expressed (or activated) in a stimulus and/or cell-specific manner (reviewed in [35]). In this regard, a shift of the secretome from a tumor suppressive (driven by wt-p53) to a cancer-promoting one has been described in models of p53 loss of function (LOF), dominant-negative (DN) inhibition by mutant p53 or gain of neomorphic functions (GOF) [36,37,38].
In a different model, the SASP of N-RAS-dependent pre-malignant senescent hepatocytes, which supplies CCL2 and IL-1α, is sufficient to drive an immune response that limits liver cancer development. In particular, CCR2+ myeloid cells are recruited into the tumor bed and a CD4+ T cell-mediated response stimulates macrophages to clear pre-malignant SnCs [39, 40]. In this way, OIS effectively contributes to the elimination phase by alerting both the innate and adaptive arms of the immune system. Moreover, CCL2 plays a key role in the elimination of senescent cancer cells in liver carcinoma mostly by driving NK cell recruitment to the tumor [41]. Accordingly, the abrogation of SASP gene expression by bromo and extra terminal domain (BET) protein inhibitors, leads to the loss of immune-mediated cancer surveillance [42]. In senescent nevi prone to melanoma transformation, senescence induction by epigenetic drugs is associated with a considerable increase of melanoma infiltrating CD11b+ innate immune cells [43]. Collectively, these studies have changed the understanding of senescence, from a cell-autonomous (intrinsic) defense mechanism restraining cell proliferation, towards an extrinsic tumor barrier involving the action of the immune system.
The ability of senescence in recruiting immune cells to tumors underpins the clinical success of different cytostatic agents adopted in cancer therapy. A combined treatment with a MEK inhibitor (trametinib) and a CDK4/6 inhibitor (palbociclib) in K-RAS-mutant lung and pancreatic cancer cells was able to induce cellular senescence with cytotoxic effects in vivo due to the recruitment (via CCL2, CCL4, CCL5, CXCL10, CX3CL1) and activation (via IL-15, IL-18, TNF-α) of immune cells, namely NK cells [44]. TIS can also sustain SASP-mediated vascular remodeling in pancreatic ductal adenocarcinoma with the result of improving both drug delivery and lymphocyte infiltration to the tumor core, leading to windows of opportunities for enhancing susceptibility to otherwise ineffective chemo- and immuno-therapies [45]. Hence, TIS can revert the equilibrium or escape phases by restoring the elimination phase.
ii) direct killing enhancement. The immune clearance of SnCs, a process called “senescence surveillance”, is of great relevance to tumor progression as the life span of SnCs in tissues deeply affects the inflammatory processes. Different cellular types are implicated in the immunosurveillance of SnCs, with macrophages and NK cells the most involved (Fig. 2). In a model of liver carcinogenesis by CCl4 fibrogenic treatment with coadministration of the carcinogen diethylnitrosamine, p53-dependent senescence skewed macrophage polarization towards an “M1-like” phenotype with SnC-targeting capacity [36]. In a different approach designed to identify unique biomarkers of senescence, senescent primary human fibroblasts were found to expose an oxidized form of membrane-bound vimentin (i.e., malondialdehyde-modified vimentin), specifically recognized by a natural polyreactive IgM antibody that likely enables phagocytosis of SnCs by macrophages [46]. Regarding the mechanisms of direct killing activity, NK cells have been studied in more detail [47]. The ability of NK cells to target SnCs is mediated by the activation of the NKG2D and DNAM-1 receptors. The corresponding stress-induced ligands are strongly upregulated by SnCs following both OIS and TIS [5]. Efficient killing of SnCs by NK cells requires granule exocytosis and not death receptor signaling [48]. Notably, NKG2D and DNAM-1 engagement also triggers the production of IFN-γ that might boost macrophage activity, possibly aiding the clearance of SnCs [49]. Senescent tumor cell clearance by NK cells has been thoroughly investigated in the context of multiple myeloma by our group. Myeloma cells treated with low doses of therapeutic agents commonly used in the management of patients with multiple myeloma, such as doxorubicin, melphalan, and bortezomib, up-regulate NK cell-activating ligands for the NKG2D and DNAM-1 receptors, increasing NK cell degranulation in vitro and in vivo, an effect preferentially associated with SnCs arrested in the G2 phase of the cell cycle [50, 51]. Genotoxic agents initiate a redox-dependent DDR that increases MICA and PVR NK cell activating ligands on senescent multiple myeloma cells through the transcriptional factor E2F1, indicating the immunogenic senescence as a key mechanism promoting the clearance of drug-treated tumor cells by innate effector lymphocytes [49]. Moreover, drug-induced senescent multiple myeloma cells can cross-present IL-15/IL-15RA complex on their membrane, thus stimulating activation and proliferation of NK cells [22].
NK-mediated cytotoxicity has also been attributed to the expression of ICAM-1 on lung tumor cells following TIS [44] and to direct intercellular CDC42 protein transfer from SnCs to NK cells in the preneoplastic pancreas, which leads to actin rearrangement and thus NK cell activation [52].
iii) antigenicity/adjuvanticity properties. As previously outlined, senescent hepatocytes expressing MHC class II and CD86 mediate CD4+ T lymphocyte activation, stimulating a macrophage response against pre-malignant senescent hepatocytes [39]. In this context, recent findings highlighted the specific contribution of the adaptive immune system. By investigating different models of senescence (drug-induced, p53-driven in fibroblasts, melanoma, and pancreatic adenocarcinoma), Serrano and colleagues demonstrated a type I IFN-dependent upregulation of MHC-I molecules on SnCs, along with an elevated senescence-associated epitope presentation. This led to CD8+ T lymphocyte activation as revealed by CD25 and CD69 expression. In addition, alarmins (ATP, calreticulin) released from SnCs were accompanied by greater dendritic cell maturation (MHC-II, CD80, and CD86 expression) than that observed in the immunogenic cell death. Remarkably, autologous antigen-specific tumor infiltrating lymphocytes were restimulated by senescence induction [53]. In the model of hepatocyte transformation by oncogenic RAS and senescence induction by p53 reactivation, Scott Lowe et al. reported complementary findings. The authors found a stark switch from an immunosuppressive tumor environment characterized by exhausted T lymphocytes to an immunocompetent setting with functional CD8+ T lymphocytes and active macrophages following senescence induction attributing these changes to a cell-surface proteome remodeling of SnCs, which includes the increase of antigen-presenting machinery and heightened responsiveness to IFN-γ [54]. Taken together, these studies emphasize that senescence potently enhances the immunogenicity of cancer cells and supports the pioneering work of Weichselbaum and colleagues that suggests to use SnCs as cancer vaccination agents [55], as for the loading of dendritic cell-based vaccines. Furthermore, the finding that senescence acts as an immunostimulatory response to drugs better than the immunogenic cell death gives rise to the possibility of dosage lowering, with the aim of reducing side effects. The identification of senescence-specific antigens in cancer cells leads to hypothesize the existence of senescence-associated antigens in senescent aged tissues with the possibility of tailoring anti-aging therapies. Molecular details on the immunopeptidome generation are still missing, but a fascinating idea is a possible link with the enhanced mobilization of retrotransposable elements that characterizes senescent cells and the following cGAS-STING pathway stimulation that in turn supports the MHC I machinery [56, 57]. As addressed below in a separate section, upregulation of the inhibitory molecule PD-L1 and PD-L2 can also occur on SnCs and this accounts for the failure of anti-tumor immunity [58]. The overall immunogenicity of a senescent tumor cell is thus a fine balance that reflects the intrinsic history of the single cell, its surrounding milieu in term of cytokines and chemokines and the compartmentalization of infiltrating immune cells in the tumor.
Senescence within the “equilibrium phase”
In cancer immunoediting, the “equilibrium phase” is when the expansion of transformed cells is arrested and maintained in a state of functional dormancy by immunity. If during the “elimination phase” the innate and adaptive immune system is unable to completely destroy the tumor, it converts cancer cells in a quiescent mass, resulting in a state of equilibrium between the developing tumor and the action of the immune system, which may occur over a period of years. In this phase the tumor dormancy is specifically controlled by adaptive immunity [59, 60] (Fig. 2A). Conceivably, the same immune functions that keep cancer cells in a “not expanding” status also provide the selective pressure responsible for the outgrowth of tumor cells once these cells have acquired immune evasive mutations.
Thus, the “equilibrium phase” is a condition that may occur before tumor development at the primary site (referred to as “equilibrium 1”), after the spreading out of single cells at the metastatic sites referred to as “equilibrium 2”) or following therapies that lead to residual tumor mass (referred to as “equilibrium 3”) (Fig. 2B). Senescence and senescence-escape could serve as one mechanism of tumor dormancy and disease recurrence. Specifically, tumor cell senescence can represent a state of dormancy due to cell cycle arrest and apoptosis-resistance, whilst the SASP can establish a dormancy-promoting environment for “cancer stem cells”.

SASP into the different steps of cancer immunoediting
(A) Cancer immunoediting is a dynamic process between tumor cells and the immune system and it has been outlined in the 3Es. In the first phase of elimination, innate and adaptive immunity destroys developing tumors before they become clinically evident. If this step is performed successfully the tumor cells are removed. If, however, the tumor cell variants are not destroyed, the equilibrium phase can be entered, in which tumor growth is confined by immunological mechanisms in a state of functional dormancy. In this phase tumor cell variants may emerge as a consequence of constant immune selection pressure and may enter into the escape phase, in which their growth is no longer blocked by immunity and tumor becomes clinically detectable. (B) SASP can lead to the clearance or the protection of cancer cells, favoring cell dormancy and tumor recurrence. In the “elimination phase” SnCs are efficiently eliminated by both innate and adaptive immune cells. Initially SASP released by oncogene-induced senescent tumor cells can recruit immune cells into the tumor bed favoring the elimination of cancer cells. During this phase some cancer cells may remain and contribute to the establishment of an equilibrium between the tumor and the immune system. In this “equilibrium phase 1”, the immune system holds the tumor in a state of functional dormancy and senescent cells may contribute by releasing a plethora of different factors conditioning the TME. During this phase there is a balance between anti-tumor and tumor promoting cytokines and due to constant immune pressure, tumor cell variants evolve that resist immune recognition and drive the following “escape phase”. As a further layer of complexity some variants that have acquired the ability to metastasize leave the primary tumor (“equilibrium phase 2”), and senescent tumor cells can be found within metastatic lesions. In the “escape phase” senescent tumor cells release a pro-inflammatory SASP and may re-entry into the cell cycle contributing to a clinically detectable tumor that can be treated by therapy. Following treatment, a new “equilibrium phase 3” can be achieved in which it is possible to observe a residual tumor mass. The final use of senotherapy can lead to complete recovery.
Abbreviation: TRM (Tissue resident memory cells); MDSC (Myeloid-derived suppressor cells)
Created with BioRender.com with granted permission and license
Senescence as a form of in-depth cell dormancy
By definition, a ‘dormant cancer cell’ requires that the cell can reawaken and proliferate to repopulate a tumor. In this regard, while cellular senescence was initially believed to be irreversible, recent studies have shown that SnCs can re-enter the cell cycle in response to appropriate stimuli [61,62,63,64,65] and contribute to delayed relapse. This further supports the concept that senescence may represent one form of “in-depth dormancy”, whereby cancer cells evade the direct cytotoxic impact of therapy. Indeed, SnCs, like dormant tumor cells, are resistant to apoptosis in response to therapies that target proliferative cells. Unlike quiescent cells, they are unresponsive to growth factors and release paracrine signaling by the SASP that can impact the TME, thus sustaining delayed relapse and metastasis [66]. An additional point to consider concerning cancer cell dormancy is the accumulation of genetic and epigenetic changes selected in the tumor variants. This mechanism can leave a track of the immune system pressure [2]. Cell-cycle re-entry in the form of senescence escape may be a driver mechanism to interrupt this state of immune dormancy. Indeed, it is now clear that senescent tumor cells can resume proliferation through cell-autonomous and non-cell-autonomous mechanisms [43, 67,68,69]. Accordingly, a recent paper reveals that epigenetic memory defines post-SnCs in colorectal cancer, raising the possibility of early detection of lesions susceptible to cancer progression [70].
Reversible senescence has been mostly observed following TIS upon drug removal [63, 71,72,73]. Interestingly, endoreplication events leading to polyploidy are common after TIS [74] and there are studies pointing out that polyploidy may represent a prerequisite for succeeding in cell cycle re-entry [75, 76]. Indeed, senescent polyploid cells give rise to cycling euploid daughter cells by budding or asynchronous division [77]. It is conceivable that polyploidization supports the energetic and biosynthetic requirements of senescent cells from a transcriptional point of view. Furthermore, escaped cells often show more aggressive tumorigenic features, with increased stem-like markers or epithelial to mesenchymal transition traits [78, 79]. For comprehensive reviews on the topic see [80,81,82]. Mechanistically, senescence escape has been also linked to a lack of persistent DDR activation or not sustained mTOR signaling [83, 84]. Of course, it remains difficult to clearly distinguish between a regrowth from a locked cell cycle arrest (true senescence escape) or just a reprogramming from a not fully achieved senescent state (senescence bypass).
The concept that dormant cancer cells can remain dormant for many years suggests that tumor-specific immune memory acts as the key player in inducing and maintaining tumor dormancy via continuous durable pressure. In a model of T cell antigen-driven multistage carcinogenesis, Müller-Hermelink and colleagues found that specific Th1 cells were able to induce a state of tumor dormancy through a coordinated interaction between the cytokines IFN-γ and TNF [85]. Paradoxically, in the absence of both cytokines, the same T cells significantly promoted angiogenesis and multistage carcinogenesis [85]. In a different experimental setting, Th1 cell immunity arrested cancer progression through IFN-γ and TNF-induced senescence, the activation of p16INK4a, and downstream Rb hypo-phosphorylation at serine 795 [86]. Hence it is imperative to support endogenous anti-tumor immune responses through strategies aimed at reinvigorating antitumor immunity, or approaches capable of reversing tumor-induced immune tolerance [87, 88]. Of note, since IFN-γ and TNF induce senescence in numerous murine and human cancers, this may be a general mechanism for arresting cancer progression and maintaining senescent dormancy [89, 90].
The mechanisms by which dormant tumor cells preclude the recruitment or function of activated T cells remain largely unknown, but a leading hypothesis is that dormant cells persist likely due to poor interaction with endogenous antigen-specific T cells, a relatively rare population [91, 92]. However, in a model of triple-negative breast cancer (TNBC), despite close proximity to effector T cells, dormant tumor cells are not eliminated and support a CD4+ and FoxP3+ Treg-rich microenvironment. Secreted factors and DKK3, the Wnt pathway antagonist, were identified as a crucial mediators of this effect [93]. Additionally, immune evasion might be involved in cellular dormancy. As a matter of fact dormant leukemia cells express PD-L1, a mechanism of T-cell inhibition [94]. It is possible that senescent dormant cells may adopt similar escape mechanisms.
The outcome of the equilibrium phase depends on the balance between the strength of endogenous anti-tumor immunity and the tolerance mechanisms developed by tumor cells [2]. The frequency of various immune cell subsets among the tumor-infiltrating lymphocytes (TILs) from the dormant and progressive sarcomas have been analyzed in a model of induced mouse sarcoma [95]. Indeed, it has been shown that the microenvironment permits the survival of tumor cells without significant proliferation through a fine balance between antitumor immune cells and immunosuppressive cells [95]. A higher frequency of intratumoral cytotoxic T lymphocytes (CTLs), NK cells and γδ-T cells has been related to the maintenance of immune-mediated dormancy although the recruitment mechanism were not clarified.
Keeping dormancy
Many studies revealed the importance of chronic inflammation in driving the reactivation of dormant cells [66, 96, 97]. However, context- and timing-dependent mixture of inflammatory cytokines, proteases, and stemness factors released by SnCs may stabilizes the senescence state in an autocrine manner, also affecting neighboring cells in a paracrine manner. Indeed, senescence can be maintained and reinforced by factors such as IL-1α and IL-6 whereas multiple SASP components, including TGF-β family ligands, VEGF, CCL2 and CCL20, can transmit non-cell autonomous growth arrest, referred to as paracrine senescence, to their neighbors [98, 99].
Intriguingly, in a recent paper, the authors through a study of integrating time-resolved gene expression and epigenomic profiles of cells escaping from oncogenic RAS-induced senescence in colorectal cancer, reveal that escape from OIS is accompanied by a gradual downregulation of SASP-associated genes and re-expression of genes that promote cell-cycle progression, confirming the role of a SASP-driven dormancy [70]. The excessive production of reactive oxygen species (ROS) by “dormant” senescent cells may also contribute to the maintenance of cellular senescence, via a positive self-amplifying loop, which constantly generates DNA damage [100]. Furthermore, a proinflammatory TME can promote macrophages to produce more ROS further amplifying this loop [101].
This evidence of a clinical-relevant dormant state of tumor cells comes from different in vitro and ex vivo models [102, 103], and from clinical data derived from late recurrence or relapse in cancer patients [104]. Nevertheless, the identity of dormant tumor cells remains elusive and studies directly identifying dormant tumor cells as senescent are scarce. However, the latest findings on the possibility of reversal of the senescent phenotype and the ability of senescent cells to release proinflammatory cytokines strongly support the idea that these cells are likely involved in cancer dormancy, escape, and recurrence.
Senescence contribution to cancer progression: the “escape phase”
Different mechanisms of escaping immune control have been described in cancer progression mainly involving: (i) prevention of immune detection, (ii) prevention of immune activation and (iii) activation of immune suppression. Importantly, in the context of anti-cancer immune responses, these three mechanisms can occur simultaneously or in distinct phases.
Senescence-driven mechanisms of escaping anticancer immune control
Induction of senescence and secretion of SASP can be regulated over the time during cancer progression, reshaping TME potentially promoting tumor growth and metastasis induction [105, 106] (Fig. 2B). In this regard, early SASP factors, such as TGF-β1 and -β3, can be immunosuppressive [107, 108] whilst the SASP secreted in a later time (deep senescence) often consists of an array of factors and proinflammatory cytokines, including the classical IL-1β, IL-6 and CXCL8, able to attract and activate a wide range of immune effector cells (e.g. CTLs, B-cells, neutrophils) [109], thus favoring the elimination of senescent cells, as previously described in the ‘‘senescence surveillance” process [110]. Noteworthy, in the case of IL-6, this cytokine can be also produced by dysregulated senescent stromal cells and has been shown to recruit myeloid suppressive cells to inhibit T-cell activation and response against malignant cells [111]. Moreover, an additional mechanism related to the persistent secretion of IL-6 by SnCs is the upregulation of HLA-E expression, able to impair their elimination by NK cells and CD8 + T cells via the inhibitory receptor NKG2A, thus inhibiting senescent cancer cell surveillance despite the presence of immune cell infiltrates [112]. A cogent example of this dual behavior of senescent cancer cells during progression has been provided by Eggert et.al [40]. where the authors described how, in HCC progression, senescence-induced immune surveillance requires the initial recruitment and maturation of CCR2+ myeloid cells; however, if SnCs are not cleared, fully established senescent HCC cells can repress the maturation of the recruited myeloid precursors, able to block the activity of NK cells and to promote tumor growth. This contradictory phenomenon can be explained by the plasticity of CCR2+ myeloid cells, upon entry into their end-organ tissue, depending on the microenvironment and cytokine milieu they can find [113, 114]. Noteworthy, senescence in peritumoral tissue of HCC patients has been associated with bad prognosis and survival of human HCC patients, and this was correlated with high CCL2 expression, accumulation of myeloid cells and low NK cell gene activity [40]. An additional example of the critical roles played by different microenvironment/cytokine milieu on myeloid cells regards macrophage polarization. Senescent hepatocytes/stellate cells have been shown to drive the polarization of macrophages toward an M1 phenotype, effective in containing liver tumor growth, through a SASP regulated by persistent p21 (Cdkn1a) expression [115]. In an opposite way, the activity of p53-deficient stellate cells, which cannot enter the senescent state, has been shown to produce a SASP able to promote M2 macrophage differentiation, thus contributing to the proliferation of premalignant tumor hepatic cells [36].
In obesity-related liver carcinoma, a different model of SASP-mediated immunosuppression, it was demonstrated that the cooperation between the hepatic translocation of the obesity-induced gut microbial component lipoteichoic acid (LTA), and the production of the obesity-induced gut microbial metabolite, deoxycholic acid, enhanced induction of SASP and the activity of COX2 in hepatic stellate cells (HSCs), by a Toll-like receptor 2 mechanism. Increased COX2-mediated prostaglandin E2 (PGE2) production suppressed anti-cancer immune response via the activity of the PTGER4 receptor, increasing formation of metastasis [116]. Thus, these findings broaden the molecular complexity associated with senescence-induced changes in the TME, highlighting that lipid mediators, such as PGs, are also produced by SnCs and act as immunomodulating SASP factors. In the context of TME, Krtolica and colleagues described for the first time that the phenomenon of cellular senescence occurs also in mesenchymal cells, revealing the important concept that cellular senescence, despite protecting against cancer in young adults, may promote cancer progression in aged organisms [106]. The SASP components released by senescent stroma include different cytokines and chemokines (e.g. CXCL8 and CXCL3), different growth factors, including amphiregulin (AREG), and are mainly involved in inducing and accelerating disease progression via paracrine actions [37, 105, 117, 118]. In particular, AREG, an epidermal growth factor (EGF) receptor ligand, is involved in the progression of many cancer types [119, 120]. Stromal cells induced to senescence by genotoxic treatments release AREG that confers increased resistance of prostate cancer cells against mitoxantrone, a DNA-targeting chemotherapeutic, and simultaneously induces the PD-L1 expression in tumors via a pathway that functionally involves EGFR and its downstream elements [121]. Cancer stage may also play a role in the ability of stromal SASP to influence tumor fate. SASP released from H202-treated mesenchymal stromal cells blocks proliferation and induces senescence of immortalized prostate cells while metastatic prostatic cancer cells appeared insensitive to SASP [122]. Furthermore, the immunomodulatory effect of senescent stroma is often correlated to the accumulation of MDSCs [23, 111].
Regarding the suppression of immune activating pathways as immune escape mechanism, it should be observed that persistent SnCs accumulate in cancer patients and are found in inflamed/damaged tissues, premalignant lesions and after chemo/radiotherapies [105, 123,124,125]. Here, the establishment of a senescence program in persistent SnCs has been shown to subvert immune clearance through the activity of MMP-dependent shedding of selected NKG2D-Ls and suppression of NKG2D-mediated immunosurveillance [126,127,128] leading to SnCs persistence, as described in cancer epithelial cells and in residual tumors from patients with breast and prostate cancer who underwent genotoxic chemotherapy [127]. Interestingly, the activity of p53 and p16 tumor suppressors was not required to induce or maintain the expression of NKG2D-Ls and MMPs in response to senescence-inducing genotoxic stresses after radiotherapy and chemotherapy [127]. Furthermore, the senescence-associated evasion process was restricted to genotoxic-resistant tumors, suggesting that a tailored senolytic therapy could eliminate deleterious resistant SnCs, avoiding possible side effects. Here, the elimination of persistent SnCs by pharmacological strategies has shown beneficial effects in mouse models [129,130,131] and approaches designed to boost the immune recognition of persistent SnCs could represent a promising strategy. These aspects will be further discussed.
Immunosenescence as further player in the “escape phase”
Immunosenescence is defined as a dysregulation of immune cell function that contributes to the increased susceptibility to several age-related diseases (including cancer). However, the senescence of the immune system can also occur independently by the age. Indeed, the accumulation of dysfunctional senescent T-cell types have been found in relatively young patients with certain types of cancers, including lung cancers, colorectal cancer, head and neck cancer, ovarian cancer and hepatocellular carcinoma [132,133,134,135,136,137]. In this regard, malignant tumors can promote T cell exhaustion and senescence leading to a defective effector function in the TME [136,137,138]. These two important dysfunctional states that share overlapping characteristics, mostly the loss of proliferative potential and CD28 surface expression [139], favor cancer progression. What is currently widely recognized as a major distinguishing difference is that exhausted T cells are characterized by elevated expression of a panel of inhibitory receptors (e.g. PD-1, CTLA-4, TIM-3) [140,141,142] while senescent T cells, together with the high expression of SA-β-Gal [143,144,145], dramatically upregulate additional senescence-associated markers, including CD57, CD45RA, and killer cell lectin-like receptor subfamily G member 1 (KLRG-1) [146, 147]. In addition, unlike exhausted T cells, senescent T cells present decreased expression of functional molecules related to cytotoxic activity, such as perforin and granzyme B, in vivo [148], and acquire a particular SASP, generating elevated amounts of proinflammatory and strongly suppressive cytokines, such as IL-10 and TGF-β [143] that contribute to the functionally hyporesponsive states of the immune system observed in the TME during cancer progression. In addition to this, in recent years, several studies have demonstrated that tumor-associated Treg cells can directly induce T cell senescence in the TME [149, 150]. Indeed, naturally occurring human CD4+CD25hiFoxP3+ Treg cells can selectively modulate p38 and ERK1/2 MAPK signaling pathways, suppressing T cell proliferation through induction of senescence on responding naive and effector T cells [143]. Treg cells also display predominant dependence on glucose metabolism compared with effector T cells thus increasing the glucose consumption that leads to cell senescence and suppression in responder T cells [150]. Interestingly, the TLR8 signaling selectively inhibits glucose uptake resulting in reversal of Treg suppression opening the possibility of identifying new cancer treatment strategies [151]. In vitro studies on human T cells have also found that induction of the DNA damage response, caused by metabolic competition, is the main cause of Treg-mediated induction of senescence on responder T cells. Indeed, the authors demonstrated that the stress-induced ERK1/2 and p38 MAPK signaling pathways functionally cooperating with the STAT1/STAT3 transcription factors, can control Treg-induced T-cell senescence [149].
Some characteristics of senescence in human effector memory CD8+ T cells that re-express the naive T cell marker CD45RA (EMRA), such as mitochondrial disruption and ROS accumulation are mediated by p38 MAPK signaling and are reversible [152]. In this scenario, the possibility to revert the senescent CD8+ phenotype is important, as the number of EMRA CD8+ T cells increases in metastatic melanoma patients [153].
Altogether these studies suggest that T-cell senescence is an alternative novel mechanism utilized by malignant tumors for immune evasion and provide the rationale for tumor immunotherapy. Finally, an additional layer of complexity is the observation that, during aging, there is an accumulation of CD8+ T cells with the senescence phenotype that acquire NK function, thus compensating for reduced TCR-mediated classical T cell function and involved in tumor elimination [154]. Indeed, CD27−CD28−CD8+ T cells with the senescence phenotype can acquire a NK function upregulating the expression of NKRs such as NKG2D, NKG2A and CD16 and are involved in tumor elimination in vitro and in vivo [155]. Accordingly, these observations have been confirmed in a B16 melanoma lung metastasis murine model, where it has been demonstrated that Menin-deficient senescent CD8+ T cells from aged mice [156, 157], play a role in antitumor immunity by acquiring NK cell-like innate immune functions [158]. Nonetheless, the role of senescent CD8+ T cells in tumor immunity remains to be fully elucidated.
Tracking the dynamics of cellular senescence via the cGAS/STING axis
In the last few years, endogenous DNA sensing through the cGAS/STING pathway has emerged as an important regulator of senescence and SASP. In this regard, the relationship between the cGAS/STING axis and senescence can be viewed as an example of the complex crosstalk between senescence induction and anti-cancer immune response taking into consideration its contribution to cancer “immunoediting”. Accumulation of double strand DNA (dsDNA) in the cytoplasm of senescent cancer cells leads to the activation of the stress sensor pathway mediated by the cyclic guanosine monophosphate-adenosine monophosphate (cGAMP) synthase (cGAS) and the adaptor stimulator of interferon genes (STING). Upon binding to dsDNA, cGAS drives the production of cGAMP, a second messenger detected by STING on the endoplasmic reticulum (ER) membrane. After that, STING transfers to the Golgi apparatus and recruits TANK-binding kinase 1 (TBK1) and IκB kinase to activate IFN regulatory factor 3 (IRF3) and NF-κB, respectively, giving rise to the production of type I interferons (IFNs) and inflammatory cytokines and chemokines [159, 160] (Fig. 3).
Senescent cells are characterized from the presence of CCFs, micronuclei and mitochondrial DNA (mtDNA) that represent important sources of immunostimulatory self-DNA able to activate the cGAS/STING pathway, thereby regulating the SASP and facilitating paracrine senescence [161,162,163,164]. Gluck and colleagues found that diverse triggers of cellular senescence, including oxidative stress, oncogene signaling, irradiation, and pro-senescent drugs (i.e.: palbociclib) depend on cGAS/STING signaling to drive the production of several SASP components [162]. As such, mouse embryonic fibroblasts deficient in cGAS exhibited reduced signs of senescence [165] and defects in cGAS or STING alter SASP composition [166]. In line with these evidences, Omer and colleagues reported that genetic depletion or chemical inhibition of the factor Ras GTPase activating protein-binding protein 1 (G3BP1) which impairs the association of cGAS with cytoplasmic DNA, prevents the expression of SASP factors without affecting cell commitment to senescence [167, 168].
As stated above, SASP is a key feature of senescent cells that mainly acts to reinforce cellular senescence in a non-cell autonomous manner. Through the auto and paracrine effects of the SASP, senescent cells modulate their own cellular fate and influence the surrounding microenvironment including immune cell functions and their recruitment. For instance, type I IFNs strongly contribute to the generation of a senescent phenotype [169, 170]. In particular, prolonged exposure to IFN-β induced a p53-dependent cellular senescence program in normal human fibroblasts [169]. The importance of cGAS-STING pathway-induced SASP and its role in cancer immune surveillance has been extensively described in several experimental models. For instance, in an in vivo model of OIS, Dou and colleagues have demonstrated that STING is essential for RAS-induced SASP and for the immune-mediated clearance of cancer cells [161]. Similarly, in an ovarian tumor model, the induction of SASP mediated by the GAS/STING pathway was shown to be decisive in promoting T cell recruitment into tumors [171]. Also, selective recruitment and activation of CD8+ T cells into mismatch repair-deficient colorectal cancers strictly depends on CCL5 and CXCL10 chemokines due to endogenous activation of cGAS/STING and type I IFN signaling by damaged DNA [172]. Another consideration to be made concerns the production of the second messenger cGAMP by tumor cells and its diffusion in the tumor microenvironment with consequent activation of STING in other cell types. Accordingly, it has been shown that cancer cell intrinsic expression of cGAS is critical for cancer immune surveillance since cGAMP produced by cancer cells is transferred to surrounding DCs and promotes type I IFN production in a STING dependent fashion [173].
In this scenario, the cGAS/STING pathway contributes to the elimination phase acting as a bridge connecting innate and adaptive immune response through an immunostimulatory SASP. On the other hand, continuous cGAS/STING activation over time, following accumulation of cytosolic dsDNA in senescent cancer cells, determines the persistence of a pro-inflammatory and pro-tumoral SASP favoring the generation of immunosuppressive populations that negatively affect cancer immunosurveillance (Fig. 3).
In addition to this, it is also important to highlight the recurrent inactivation of cGAS/STING pathway in a wide variety of cancers [163, 165, 174, 175] suggesting a potential additional immune escape mechanism driven by modulation of the cGAS/STING pathway. Nonetheless, the impact of cGAS/STING inactivation in the crosstalk between senescence and cancer immune surveillance remains to be fully elucidated.
Overall, these studies emphasize multifaceted roles of cGAS/STING pathway in the induction, maintenance and persistence of senescence and its remarkable influence on immune cell response through SASP-related components. Therefore, it will be of interest to extend the field of application of drugs targeting cGAS or STING developed for cancer immunotherapy [176] as novel regulators of senescence cell fate.

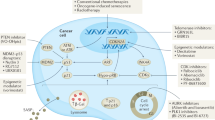
cGAS/STING pathway as driver of senescence and immunosurveillance
(1) Upon binding to cytosolic dsDNA, cGAS drives the production of cGAMP that is detected by STING on ER membrane. Then, STING transfers to the Golgi apparatus and recruits TBK1 and IκB kinases to activate IRF3 and NF-κB, respectively, giving rise to the production of type I IFNs and a plethora of inflammatory cytokines and chemokines that are secreted in the extracellular milieu representing the SASP components. (2) SASP factors promote autocrine and paracrine senescence, moreover they contribute to immune cell recruitment and activation favoring the elimination of cancer senescent cells. (3) Sustained cGAS/STING activation over time, following accumulation of cytosolic dsDNA in senescent cancer cells, determines the persistence of a pro-inflammatory and pro-tumoral SASP favoring the generation of immunosuppressive populations that negatively affect cancer immunosurveillance.
Abbreviations: Treg (T regulatory cells); MDSC (Myeloid Suppressor Cells); iDC (immature Dendritic Cells)
Created with BioRender.com with granted permission and license
Modulation of senescence for cancer immunotherapy
Accumulation and persistence of SnCs in cancer patients has been proven to be detrimental to the organism. Selective removal of senescent cells, namely senotherapy, has been proposed as a promising adjuvant approach to eliminate the adverse effects of SnCs [177,178,179]. Senotherapy can be pursued through different strategies including: (i) the usage of senolytics, i.e. drugs targeting senescent cells; (ii) modulation or inhibition of SASP (senomorphics); (iii) immune cell-mediated clearance of senescent cells. The use of senotherapy in combination with current cancer therapies represents the so-called “one-two punch” approach in which cancer cells are forced to senescence by pro-senescence therapies and then killed by senolytic drugs or by the action of the immune system. Albeit metabolically active, senescent cells are cell cycle arrested and, therefore, cellular senescence has been viewed as a desirable outcome during cancer treatment, provided that SASP adverse effects are abrogated by senomorphics. Herein we will focus above all on approaches aimed at enhancing the recognition and elimination of SnCs by immune cells.
Senolytic drugs and senomorphics
Briefly, the development of senolytic drugs is mainly based on the inhibition of biological pathways essential for the survival of senescent cells. This includes the targeting of the BCL-2 family members, p53/p21 axis, PI3K/AKT pathway, receptor tyrosine kinases, and HIF-1α and HSP90 proteins. For instance, Navitoclax/ABT-263 or ABT737 compounds target BCL-2 family members and are effective to induce cell death in SnCs. However, these compounds have been shown to be highly harmful acting also on non-SnCs [180]. In a different way, HSP90 inhibitors, recently identified as new class of senolytic agents in a drug screening approach, are immunomodulators with potent anti-tumor activity [181]. In this regard we and others have demonstrated that HSP90 inhibitors induce the expression of NKG2D ligands on cancer cells that become more sensitive to NK cell mediated attack [182, 183].
Novel therapeutic strategies aimed at preventing the detrimental activity of SnCs are focused on the inhibition or modulation of SASP without killing SnCs by use of the so-called “senomorphics”. Distinct approaches have been carried out so far to reprogram or abrogate SASP that is regulated at epigenetic, transcriptional and post-transcriptional levels. The majority of senomorphics target key SASP regulators including chromatin readers (i.e.: BRD4, MLL1, HMGB2), JAK2/STAT3, cGAS/STING, mTOR, p38MAPK, and NF-kB [12, 177,178,179].
A large body of evidence has described the importance of epigenetic regulation in the control of SASP. As such, inhibition of the BET protein BRD4 reduced the production of IL-1α, IL-1β, IL-6, and CXCL8 in senescent fibroblasts [42]. In addition, suppression of other chromatin modifiers including mixed-lineage leukemia protein 1 (MLL1) and high-mobility group box 2 (HMGB2) [184, 185] hampered SASP inflammatory factors without altering the senescence phenotype.
Through an RNAi screen, the splicing factor polypyrimidine tract binding protein 1 (PTBP1) was identified as potent SASP modulator as its depletion suppresses NF-kB pathway and blunts pro-inflammatory cytokine secretion in various cell types exposed to different senescence-inducing stimuli. Moreover, PTBP1 knockout mice in a model of liver cancer, decreased immune cell infiltrate [186]. Pharmacological inhibition of the JAK2/STAT3 pathway in Pten-deficient senescent prostate tumors modified SASP composition, leading to an anti-tumor immune response that enhances chemotherapy efficacy [187]. The role of JAK/STAT pathway in the modulation of SASP was further explored in senescent preadipocytes and umbilical vein endothelial cells (HUVEC) [188]. It was discovered that different JAK inhibitors reduced the production of some cytokines including IL-6, CXCL8, CCL2, CXCL1 and in vivo administration of the JAK1/2 inhibitor ruxolitinib reduced systemic and adipose tissue inflammation in aged mice [188].
As already mentioned, the cGAS-STING pathway plays a central role in the establishment of senescence and SASP. For instance, G3BP1 depletion or its pharmacological inhibition impairs the cGAS-pathway preventing the expression of SASP factors (i.e.: IL-6, TNFα, IFNβ, CXCL8) without affecting the senescence phenotype [168]. Thus, the usage of cGAS or STING inhibitors could represent an alternative approach to modulate the release of SASP components avoiding their accumulation over time [189] (Fig. 3).
In a drug screen to find novel SASP regulators, rapamycin, a selective inhibitor of the mTOR complex 1 (mTORC1), abrogated the induction of a number of SASP components through a mechanism dependent on downregulation of mitogen-activated protein kinase-activated protein kinase 2 (MAP-KAPK2 or MK2), a downstream effector of p38, thereby reducing breast tumor growth in mice [190]. It should be noted that mTOR can affect SASP at several levels; an elegant study by Laberge and colleagues reported that rapamycin modulates SASP by reducing IL-1α mRNA translation and as consequence IL1R1 signaling leading to a decreased NF-κB transcriptional activity. In this context, the NF-κB-dependent expression of IL-6 and CXCL8 was suppressed in senescent cells [191]. As matter of fact, in a model of OIS into human lung embryonic fibroblasts, IL-1a knockdown was shown to weaken the late proinflammatory arm of SASP controlled by NF-κB without altering cell cycle arrest [192]. Moreover, ablation of the SASP through the deletion of IL-1α gene in a mouse model of pancreatic cancer impairs tumor progression [192].
Based on the above findings, selective removal of one or more SASP components may represent a promising strategy to reduce SASP detrimental effects since monoclonal antibodies and other agents targeting inflammatory cytokines and their receptors are clinically approved for inflammatory disorders. These include drugs targeting IL-1α/IL-1β/IL1R (Anakinra, Canakinumab, Rilonacept) [193, 194], TNFα (Infliximab, Etanercept) [195], and IL-6/IL-6R (Tocilizumab, Siltuximab) [196]. Interestingly, new nanotechnologies settled on the employment of galactose-modified nanoparticles have been recently proposed to increase the specificity of drug delivery into senescent cells [197, 198]. Table 1 recapitulates some experimental models of senescence used to study SASP modulation.
Studies on senomorphics still lack of completeness, as most of the research has largely employed senescence models derived from tumor suppressors loss of function (i.e. p53, PTEN) or from OIS. It is not supposed that similar results could be obtained in models of TIS. Additional work is urgently needed to elucidate how modulation of SASP in different model of senescence and in different cancer contexts may impact tumor immunosurveillance.
Harnessing the immune cell mediated clearance of SnCs
As previously discussed, NK cells strongly contribute to the elimination of senescent cells in different cancer models, thus NK cell engaging therapies such as therapeutic antibodies or adoptive NK cell transfer may further increase the clearance of SnCs [199]. Indeed, combined high-dose radiotherapy and adoptive NK cell transfer improve tumor control over monotherapies in xenografted mice. Noteworthy, the authors showed that CXCL8, as a SASP component, was responsible for NK cell recruitment and infiltration to the tumor site [200]. In a different therapeutic strategy, targeting the inhibitory receptor NKG2A that binds the non-classical MHC-class Ib molecule HLA-E, could be employed as a senolytic strategy to boost NK cell activity. As stated before, HLA-E was found upregulated on the cell surface of senescent fibroblasts exposed to the SASP-related cytokine IL-6. Inhibition of HLA-E expression through RNA interference enhanced NK cell-mediated cytotoxicity against SnCs [112] suggesting the usage of monalizumab, a therapeutic antibody targeting NKG2A, in cancer patients [201].
An additional relevant therapeutic target to consider derived from the observation that SnCs can evade NK cell mediated surveillance through the shedding of NKG2DLs [126,127,128]. Hence, strategies aimed at stabilizing NKG2DLs on the cell surface of SnCs look promising to foster NK cell senescence surveillance. For instance, antibodies blocking MICA/B protease-mediated cleavage have been developed and their effectiveness has been assessed in pre-clinical models [202]. In another study, Zingoni and coworkers demonstrated that the combined use of chemotherapeutic drugs and metalloproteinase inhibitors strengthened NK cell-mediated killing of senescent multiple myeloma cells, preserving the expression of MIC molecules on the cell surface [126].
A fast-developing field in cancer immunotherapy relies on the search of cell surface proteins specifically or highly expressed by SnCs that can be employed to design antibodies and chimeric antigen receptors (CAR). Of interest, by mass spectrometry analysis, dipeptidyl peptidase 4 (DPP4 or CD26) was identified as a surface marker of senescent human fibroblasts. Moreover, DPP4+ SnCs opsonized with an anti-DPP4 specific antibody were handled as NK cell targets in an antibody-dependent cell-mediated cytotoxicity (ADCC) assay [203].
Amor and colleagues identified the urokinase-type plasminogen activator receptor (uPAR) as a SnC-surface marker and showed that uPAR-specific CAR T cells can efficiently eliminate lung cancer cells induced into senescence in response to combined therapeutic treatment (i.e. MEK plus CDK4/6 inhibitors). Further, in a xenograft model of lung cancer, uPAR-specific CAR T cells selectively killed SnCs in tumor tissues and extended the survival of mice bearing lung adenocarcinoma [204]. Interestingly, as already mentioned, NKG2DLs are highly expressed by SnCs and have been proposed as target molecules for CAR T cell therapy. A recent study demonstrated that NKG2D-CAR T cells recognized and killed NKG2DL+ SnCs in aged mice. Furthermore, the use of murine NKG2D-CAR T cells in two independent mouse models of aging, reversed senescence-associated phenotypes, highlighting their potential in treating aging and age-related diseases, including cancer [205]. These reports indicate that T-CAR technology applied to the recognition of senescent tumor cells could boost therapies based on the induction of senescence. Novel approaches aimed at eliminating dysfunctional senescent T lymphocytes may also contribute to enhance cancer immunosurveillance [149, 151]. For example, a vaccine targeting CD153 has been found to deplete senescent T lymphocytes from high-fat diet-induced obese mice [206].
Targeting the immune checkpoint inhibitors (ICi) PD-1 and PDL-1/2
A large body of evidence has described the upregulation of the immune checkpoint molecule PD-L1 [58, 207,208,209] and PD-L2 [210] in SnCs leading to an impairment of T cell mediated immune response.
Accordingly, PD-L1 targeting combined with chemotherapy could be effective to potentiate the clearance of SnCs. In this regard, a recent study reported that the combined use of PARPi (talazoparib) and CDK4/6i (palbociclib) potently triggered TIS in colorectal cancer cells and promoted T cell-mediated anti-tumor immune response. In this context, characterization of SASP components revealed the presence of type I IFNs and some interferon-stimulated genes (ISGs) like CCL5 and CXCL10 chemokines as result of the activation of cGAS/STING signaling pathway. The addition of the immune checkpoint inhibitor anti-PD-L1 further improved the clearance of senescent cancer cells in immunocompetent mice [209]. In a Trp53 WT mouse mammary tumor model, chemotherapy followed by treatment with anti-PD-L1, stimulated a substantial accumulation of T cells within the tumor and a concomitant anti-tumor immune response. In addition, single cell analysis data showed that senescent PD-L1+ cells are characterized by enhanced IFNγ signaling (Stat1, Irf1, and Lgals9) and display higher PD-L1 expression compared to non-senescent cells [58]. Accordingly, upregulation of PD-L1 expression was previously shown to be regulated by both type I and type II interferons [211] and it is interesting to note that independently from the trigger (i.e.: replication exhaustion, activated oncogene HRasV12, DNA-damaging agent etoposide, or ionizing irradiation), PD-L1 is upregulated in all forms of senescence in primary human diploid lung fibroblasts [208].
Several studies provided evidence that secretome produced by senescent cells could contribute to the recruitment of PD-1+ T lymphocytes within the tumor. Remarkably, using a pre-clinical mouse model of breast cancer brain metastasis (BCBM), Uceda-Castro and colleagues discovered that doxorubicin-induced senescence in BCBM tumor cells triggers a significant recruitment of PD-1+ CD8+ T lymphocytes to BCBM lesions thus promoting the efficacy of anti-PD-1 treatment [212].
Moreover, in a preclinical mouse model of KRAS-mutant pancreatic ductal adenocarcinoma, chemotherapy-induced senescent cancer cells led to the secretion of pro-angiogenic SASP factors, which favored the intratumoral accumulation of CD8+ T cells, thereby sensitizing tumors to PD-1 checkpoint blockade [45]. Taken together these studies highlight how anticancer immunotherapies combined with ICi reinforce the efficacy of senotherapy as well as its outcome.
In Fig. 4, immunotherapeutic strategies involved in the clearance of SnCs are illustrated.

Immune cell-mediated clearance of senescent cells
a) Adoptive NK cell transfer combined with radio and chemotherapy. b) Approaches aimed at fostering NK cell recognition and killing of senescent cells including: (i) stabilization of NKG2DLs on senescent cell surface by blocking the protease-mediated cleavage through iMMPs or mAbs targeting the MICA/B shedding domain; (ii) targeting CD94/NKG2A inhibitory receptor using anti-NKG2A therapeutic mAb (i.e.: monalizumab); (iii) ADCC triggered by anti-DDP4 mAb. c) Enhancement of T cell mediated functions by targeting PD-1/PDL-1 axis; generation of uPAR specific CAR-T cells; d) depletion of senescent T lymphocytes following αCD153 vaccination
Abbreviations: iMMPs (metalloproteinase inhibitors); ADCC (Antibody dependent cellular cytotoxicity); NKG2DLs (NKG2D ligands)
Created with BioRender.com with granted permission and license
Conclusions
Cancer immunoediting theory represents the conceptualization of the complex and highly dynamic interaction that exists between neoplastic growth and the immune system. Unfortunately, some pieces of this picture are missing. Cellular senescence is one of this, as its impact on tumor progression is still unclear. Initially described as a defense mechanism against malignant transformation, its contribution to inflammation can definitely favor cancer progression. In both cases cellular senescence has an intimate connection with the immune system. As debated in the present review, there is strong evidence of the role of senescence in shaping the elimination and escape phases of cancer immunoediting, with implications that should not be neglected in tailoring the upcoming therapeutic strategies. The equilibrium phase, instead, has been less investigated so far due to the lack of experimental models and technical difficulties, but the discovery of dormant cancer cells and the new methodologies about single cell profiling are shedding light on this aspect so relevant for the biology of tumors. In this context, the connection between dormancy and senescence is extremely interesting and approaches currently used for targeting SnCs could be extended to dormant cancer cells. It should be noted that findings derived from different experimental models or settings could not drive to similar conclusions. A factor of complexity is given by the high variable spectrum of immune responses which characterize different types of tumors, spanning from hematological to solid tumors or slow to fast growing cancers. Furthermore, the senescent phenotype can affect not only cancer cells but also cells of the TME impacting on the mechanisms of tumor growth (e.g. epithelial to mesenchymal transition) and drug resistance. SnCs are nowadays considered a key component of premalignant lesions and tumor mass due to oncogenic and other cancer-related stresses, where they can act either as antitumor barrier or inflammatory agent, as stated above. Relevant for anticancer therapies, senescence is a common outcome of different genotoxic drugs and molecularly targeted agents routinely adopted in clinical settings, with uncertain effects on therapy efficacy. Senescence may serve as one mechanism of drug resistance and tumor dormancy, thus contributing to cancer relapse, as senescent cells share characteristics with dormant cancer cells. For all these reasons, cellular senescence deeply affects cancer immunoediting all along the way, pushing the action of the immune system into even opposite directions. Uncovering the underlying mechanisms that link senescence to cancer immunoediting is thus mandatory to depict the complex immune landscape that characterizes tumor progression and consequently adopt successful actions.
Data availability
No datasets were generated or analysed during the current study.
Abbreviations
- ADCC:
-
Antibody Dependent Cellular Cytotoxicity
- AREG:
-
Aphiregulin
- BET:
-
Bromo and Extra Terminal domain
- CAR:
-
Chimeric Antigen Receptor
- cGAS:
-
Cyclic GMP-AMP Synthase
- CCFs:
-
Cytoplasmic Chromatin Fragments
- CTLs:
-
Cytotoxic T lymphocytes
- DDR:
-
DNA Damage Response
- dsDNA:
-
Double Strand DNA
- EGCG:
-
Epigallocatechin gallate
- EGF:
-
Epidermal Growth Factor
- ER:
-
Endoplasmic Reticulum
- HCC:
-
Hepatocellular Carcinoma
- HPCs:
-
Hepatic Stellate Cells
- ICi:
-
Immune Check Point Inhibitors
- ISGs:
-
Interferon Stimulated Genes
- KLRG-1:
-
Killer Cell Lectin-Like Receptor Subfamily G Member 1
- LTA:
-
Lipoteichoic Acid
- MM:
-
Multiple Myeloma
- MDSC:
-
Myeloid-derived suppressor cells
- NK:
-
Natural Killer
- NKG2DL:
-
NKG2D Ligand
- OIS:
-
Oncogene-Induced Senescence
- PGE2:
-
Prostaglandin E2
- ROS:
-
Reactive Oxygen Species
- SASP:
-
Senescence-Associated Secretory Phenotype
- SnCs:
-
Senescent Cancer Cells
- STING:
-
Stimulator of Interferon Genes
- TBK:
-
TANK-Binding Kinase
- TILs:
-
Tumor-Infiltrating Lymphocytes
- TIS:
-
Therapy-Induced Senescence
- TME:
-
Tumor Microenvironment
- TNA:
-
Tumor Neoantigens
- TRM:
-
Tissue resident memory cells
References
Dunn GP, Bruce AT, Ikeda H, Old LJ, Schreiber RD. Cancer immunoediting: from immunosurveillance to tumor escape. Nat Immunol [Internet]. 2002;3:991–8. https://doi.org/10.1038/ni1102-991
Schreiber RD, Old LJ, Smyth MJ. Cancer immunoediting: integrating immunity’s roles in cancer suppression and promotion. Science. 2011;331:1565–70.
BURNET M. Cancer: a biological approach. III. Viruses associated with neoplastic conditions. IV. Practical applications. Br Med J. 1957;1:841–7.
McHugh D, Gil J. Senescence and aging: causes, consequences, and therapeutic avenues. J Cell Biol. 2018;217:65–77.
Sagiv A, Burton DGA, Moshayev Z, Vadai E, Wensveen F, Ben-Dor S, et al. NKG2D ligands mediate immunosurveillance of senescent cells. Aging. 2016;8:328–44.
Karin O, Agrawal A, Porat Z, Krizhanovsky V, Alon U. Senescent cell turnover slows with age providing an explanation for the Gompertz law. Nat Commun. 2019;10:5495.
HAYFLICK L, MOORHEAD PS. The serial cultivation of human diploid cell strains. Exp Cell Res. 1961;25:585–621.
López-Otín C, Blasco MA, Partridge L, Serrano M, Kroemer G. Hallmarks of aging: an expanding universe. Cell. 2023;186:243–78.
Hanahan D. Hallmarks of Cancer: New dimensions. Cancer Discov. 2022;12:31–46.
Gorgoulis V, Adams PD, Alimonti A, Bennett DC, Bischof O, Bishop C, et al. Cellular Senescence: defining a path Forward. Cell. 2019;179:813–27.
Ou H-L, Hoffmann R, González-López C, Doherty GJ, Korkola JE, Muñoz-Espín D. Cellular senescence in cancer: from mechanisms to detection. Mol Oncol. 2021;15:2634–71.
Cuollo L, Antonangeli F, Santoni A, Soriani A. The Senescence-Associated Secretory phenotype (SASP) in the Challenging Future of Cancer Therapy and Age-Related diseases. Biology (Basel). 2020;9.
Kowald A, Passos JF, Kirkwood TBL. On the evolution of cellular senescence. Aging Cell. 2020;19:e13270.
Kumari R, Jat P. Mechanisms of Cellular Senescence: cell cycle arrest and Senescence Associated Secretory phenotype. Front Cell Dev Biol. 2021;9:1–24.
Takasugi M. Emerging roles of extracellular vesicles in cellular senescence and aging. Aging Cell [Internet]. 2018;17:e12734. https://doi.org/10.1111/acel.12734
Abbas M, Jesel L, Auger C, Amoura L, Messas N, Manin G, et al. Endothelial microparticles from Acute Coronary Syndrome patients induce premature coronary artery endothelial cell aging and thrombogenicity: role of the Ang II/AT1 Receptor/NADPH oxidase-mediated activation of MAPKs and PI3-Kinase pathways. Circulation. 2017;135:280–96.
Takasugi M, Okada R, Takahashi A, Virya Chen D, Watanabe S, Hara E. Small extracellular vesicles secreted from senescent cells promote cancer cell proliferation through EphA2. Nat Commun [Internet]. 2017;8:1–11. https://doi.org/10.1038/ncomms15728
Weilner S, Schraml E, Wieser M, Messner P, Schneider K, Wassermann K, et al. Secreted microvesicular miR-31 inhibits osteogenic differentiation of mesenchymal stem cells. Aging Cell. 2016;15:744–54.
Vulpis E, Cuollo L, Borrelli C, Antonangeli F, Masuelli L, Cippitelli M et al. Doxorubicin-mediated miR-433 expression on Exosomes promotes Bystander Senescence in multiple myeloma cells in a DDR-Independent manner. Int J Mol Sci. 2023;24.
Vulpis E, Loconte L, Peri A, Molfetta R, Caracciolo G, Masuelli L, et al. Impact on NK cell functions of acute versus chronic exposure to extracellular vesicle-associated MICA: dual role in cancer immunosurveillance. J Extracell Vesicles. 2022;11:e12176.
Soriani A, Vulpis E, Cuollo L, Santoni A, Zingoni A. Cancer extracellular vesicles as novel regulators of NK cell response. Cytokine Growth Factor Rev. 2020;51:19–26.
Borrelli C, Ricci B, Vulpis E, Fionda C, Ricciardi MR, Petrucci MT, et al. Drug-Induced senescent multiple myeloma cells elicit NK Cell proliferation by direct or exosome-mediated IL15 trans-presentation. Cancer Immunol Res. 2018;6:860–9.
Guan X, LaPak KM, Hennessey RC, Yu CY, Shakya R, Zhang J, et al. Stromal senescence by prolonged CDK4/6 inhibition potentiates tumor growth. Mol Cancer Res. 2017;15:237–49.
Pardella E, Pranzini E, Nesi I, Parri M, Spatafora P, Torre E et al. Therapy-Induced Stromal Senescence Promoting Aggressiveness of Prostate and Ovarian Cancer. Cells [Internet]. 2022;11. Available from: https://www.mdpi.com/2073-4409/11/24/4026
Kanehira M, Fujiwara T, Nakajima S, Okitsu Y, Onishi Y, Fukuhara N et al. A Lysophosphatidic Acid Receptors 1 and 3 Axis Governs Cellular Senescence of Mesenchymal Stromal Cells and Promotes Growth and Vascularization of Multiple Myeloma. Stem Cells [Internet]. 2016;35:739–53. https://doi.org/10.1002/stem.2499
Sikora E, Bielak-Zmijewska A, Mosieniak G. A common signature of cellular senescence; does it exist? Ageing Res Rev. 2021;71:101458.
Vitale I, Sistigu A, Manic G, Rudqvist N-P, Trajanoski Z, Galluzzi L. Mutational and Antigenic Landscape in Tumor Progression and Cancer Immunotherapy. Trends Cell Biol. 2019;29:396–416.
Schmitt CA, Wang B, Demaria M. Senescence and cancer — role and therapeutic opportunities. Nat Rev Clin Oncol. 2022;19:619–36.
Xue W, Zender L, Miething C, Dickins RA, Hernando E, Krizhanovsky V et al. Senescence and tumour clearance is triggered by p53 restoration in murine liver carcinomas. Nature [Internet]. 2007;445:656–60. https://doi.org/10.1038/nature05529
Ventura A, Kirsch DG, McLaughlin ME, Tuveson DA, Grimm J, Lintault L, et al. Restoration of p53 function leads to tumour regression in vivo. Nature. 2007;445:661–5.
Chien Y, Scuoppo C, Wang X, Fang X, Balgley B, Bolden JE, et al. Control of the senescence-associated secretory phenotype by NF-κB promotes senescence and enhances chemosensitivity. Genes Dev. 2011;25:2125–36.
Perkins ND. Integrating cell-signalling pathways with NF-κB and IKK function. Nat Rev Mol Cell Biol [Internet]. 2007;8:49–62. https://doi.org/10.1038/nrm2083
Huang W-C, Ju T-K, Hung M-C, Chen C-C. Phosphorylation of CBP by IKKα Promotes Cell Growth by Switching the Binding Preference of CBP from p53 to NF-κB. Mol Cell [Internet]. 2007;26:75–87. Available from: https://www.sciencedirect.com/science/article/pii/S1097276507001190
Amanda K, Frank Julia I-J, Leu YZKDTNAK-SMH, Murphy ME. The Codon 72 Polymorphism of p53 Regulates Interaction with NF-κB and Transactivation of Genes Involved in Immunity and Inflammation. Mol Cell Biol [Internet]. 2011;31:1201–13. https://doi.org/10.1128/MCB.01136-10
Pavlakis E, Stiewe T. p53’s Extended Reach: The Mutant p53 Secretome. Biomolecules [Internet]. 2020;10. Available from: https://www.mdpi.com/2218-273X/10/2/307
Lujambio A, Akkari L, Simon J, Grace D, Tschaharganeh DF, Bolden JE, et al. Non-cell-autonomous tumor suppression by p53. Cell. 2013;153:449–60.
Coppé JP, Patil CK, Rodier F, Sun Y, Muñoz DP, Goldstein J et al. Senescence-associated secretory phenotypes reveal cell-nonautonomous functions of oncogenic RAS and the p53 tumor suppressor. PLoS Biol. 2008;6.
Pribluda A, Elyada E, Wiener Z, Hamza H, Goldstein RE, Biton M et al. A Senescence-Inflammatory Switch from Cancer-Inhibitory to Cancer-Promoting Mechanism. Cancer Cell [Internet]. 2013;24:242–56. Available from: https://www.sciencedirect.com/science/article/pii/S153561081300281X
Kang T-W, Yevsa T, Woller N, Hoenicke L, Wuestefeld T, Dauch D, et al. Senescence surveillance of pre-malignant hepatocytes limits liver cancer development. Nature. 2011;479:547–51.
Eggert T, Wolter K, Ji J, Ma C, Yevsa T, Klotz S, et al. Distinct functions of Senescence-Associated Immune responses in Liver Tumor Surveillance and Tumor Progression. Cancer Cell. 2016;30:533–47.
Iannello A, Thompson TW, Ardolino M, Lowe SW, Raulet DH. p53-dependent chemokine production by senescent tumor cells supports NKG2D-dependent tumor elimination by natural killer cells. J Exp Med. 2013;210:2057–69.
Tasdemir N, Banito A, Roe J-S, Alonso-Curbelo D, Camiolo M, Tschaharganeh DF, et al. BRD4 connects enhancer remodeling to Senescence Immune Surveillance. Cancer Discov. 2016;6:612–29.
Yu Y, Schleich K, Yue B, Ji S, Lohneis P, Kemper K, et al. Targeting the Senescence-Overriding Cooperative activity of structurally unrelated H3K9 demethylases in Melanoma. Cancer Cell. 2018;33:322–e3368.
Ruscetti M, Leibold J, Bott MJ, Fennell M, Kulick A, Salgado NR, et al. NK cell-mediated cytotoxicity contributes to tumor control by a cytostatic drug combination. Science. 2018;362:1416–22.
Ruscetti M, Morris JP 4th, Mezzadra R, Russell J, Leibold J, Romesser PB, et al. Senescence-Induced Vascular Remodeling creates therapeutic vulnerabilities in Pancreas Cancer. Cell. 2020;181:424–e44121.
Frescas D, Roux CM, Aygun-Sunar S, Gleiberman AS, Krasnov P, Kurnasov OV, et al. Senescent cells expose and secrete an oxidized form of membrane-bound vimentin as revealed by a natural polyreactive antibody. Proc Natl Acad Sci U S A. 2017;114:E1668–77.
Antonangeli F, Zingoni A, Soriani A, Santoni A. Senescent cells: living or dying is a matter of NK cells. J Leukoc Biol. 2019;105:1275–83.
Sagiv A, Biran A, Yon M, Simon J, Lowe SW, Krizhanovsky V. Granule exocytosis mediates immune surveillance of senescent cells. Oncogene [Internet]. 2013;32:1971–7. https://doi.org/10.1038/onc.2012.206
Soriani A, Iannitto ML, Ricci B, Fionda C, Malgarini G, Morrone S, et al. Reactive oxygen species- and DNA damage response-dependent NK cell activating ligand upregulation occurs at transcriptional levels and requires the transcriptional factor E2F1. J Immunol. 2014;193:950–60.
Soriani A, Zingoni A, Cerboni C, Iannitto ML, Ricciardi MR, Di Gialleonardo V, et al. ATM-ATR-dependent up-regulation of DNAM-1 and NKG2D ligands on multiple myeloma cells by therapeutic agents results in enhanced NK-cell susceptibility and is associated with a senescent phenotype. Blood. 2009;113:3503–11.
Antonangeli F, Soriani A, Ricci B, Ponzetta A, Benigni G, Morrone S, et al. Natural killer cell recognition of in vivo drug-induced senescent multiple myeloma cells. Oncoimmunology. 2016;5:e1218105.
Biran A, Perelmutter M, Gal H, Burton DGA, Ovadya Y, Vadai E, et al. Senescent cells communicate via intercellular protein transfer. Genes Dev. 2015;29:791–802.
Marin I, Boix O, Garcia-Garijo A, Sirois I, Caballe A, Zarzuela E, et al. Cellular Senescence is immunogenic and promotes Antitumor Immunity. Cancer Discov. United States; 2023. pp. 410–31.
Chen H-A, Ho Y-J, Mezzadra R, Adrover JM, Smolkin R, Zhu C, et al. Senescence Rewires Microenvironment sensing to facilitate Antitumor Immunity. Cancer Discov. United States; 2023. pp. 432–53.
Meng Y, Efimova EV, Hamzeh KW, Darga TE, Mauceri HJ, Fu Y-X, et al. Radiation-inducible immunotherapy for cancer: senescent tumor cells as a cancer vaccine. Mol Ther. 2012;20:1046–55.
Anthony R, Colombo HKE, Ramsingh G. Senescence induction universally activates transposable element expression. Cell Cycle [Internet]. 2018;17:1846–57. https://doi.org/10.1080/15384101.2018.1502576
De Cecco M, Ito T, Petrashen AP, Elias AE, Skvir NJ, Criscione SW, et al. L1 drives IFN in senescent cells and promotes age-associated inflammation. Nature. 2019;566:73–8.
Shahbandi A, Chiu FY, Ungerleider NA, Kvadas R, Mheidly Z, Sun MJS et al. Breast cancer cells survive chemotherapy by activating targetable immune-modulatory programs characterized by PD-L1 or CD80. Nat. Cancer. Springer US; 2022.
Koebel CM, Vermi W, Swann JB, Zerafa N, Rodig SJ, Old LJ, et al. Adaptive immunity maintains occult cancer in an equilibrium state. Nature. 2007;450:903–7.
Teng MWL, Vesely MD, Duret H, McLaughlin N, Towne JE, Schreiber RD, et al. Opposing roles for IL-23 and IL-12 in maintaining occult cancer in an equilibrium state. Cancer Res. 2012;72:3987–96.
Aguirre-Ghiso JA. Models, mechanisms and clinical evidence for cancer dormancy. Nat Rev Cancer. 2007;7:834–46.
Santos-de-Frutos K, Djouder N. When dormancy fuels tumour relapse. Commun Biol. 2021;4:747.
Roberson RS, Kussick SJ, Vallieres E, Chen S-YJ, Wu DY. Escape from therapy-induced accelerated cellular senescence in p53-null lung cancer cells and in human lung cancers. Cancer Res. 2005;65:2795–803.
Saleh T, Gewirtz DA. Considering therapy-induced senescence as a mechanism of tumour dormancy contributing to disease recurrence. Br J Cancer. 2022;126:1363–5.
Pluquet O, Abbadie C, Coqueret O. Connecting cancer relapse with senescence. Cancer Lett. 2019;463:50–8.
Truskowski K, Amend SR, Pienta KJ. Dormant cancer cells: programmed quiescence, senescence, or both? Cancer Metastasis Rev [Internet]. 2023;42:37–47. https://doi.org/10.1007/s10555-022-10073-z
Patel PL, Suram A, Mirani N, Bischof O, Herbig U. Derepression of hTERT gene expression promotes escape from oncogene-induced cellular senescence. Proc Natl Acad Sci U S A. 2016;113:E5024–33.
Zampetidis CP, Galanos P, Angelopoulou A, Zhu Y, Polyzou A, Karamitros T, et al. A recurrent chromosomal inversion suffices for driving escape from oncogene-induced senescence via subTAD reorganization. Mol Cell. 2021;81:4907–e49238.
Milanovic M, Fan DNY, Belenki D, Däbritz JHM, Zhao Z, Yu Y et al. Senescence-associated reprogramming promotes cancer stemness. Nature [Internet]. 2018;553:96–100. https://doi.org/10.1038/nature25167
Martínez-Zamudio RI, Stefa A, Nabuco Leva Ferreira Freitas JA, Vasilopoulos T, Simpson M, Doré G, et al. Escape from oncogene-induced senescence is controlled by POU2F2 and memorized by chromatin scars. Cell Genomics. 2023;3:100293.
Saleh T, Tyutyunyk-Massey L, Murray GF, Alotaibi MR, Kawale AS, Elsayed Z, et al. Tumor cell escape from therapy-induced senescence. Biochem Pharmacol. 2019;162:202–12.
Elmore LW, Di X, Dumur C, Holt SE, Gewirtz DA. Evasion of a single-step, chemotherapy-induced senescence in breast cancer cells: implications for treatment response. Clin cancer Res off J Am Assoc Cancer Res. 2005;11:2637–43.
Fleury H, Malaquin N, Tu V, Gilbert S, Martinez A, Olivier MA et al. Exploiting interconnected synthetic lethal interactions between PARP inhibition and cancer cell reversible senescence. Nat Commun [Internet]. 2019;10. https://doi.org/10.1038/s41467-019-10460-1
Puig P-E, Guilly M-N, Bouchot A, Droin N, Cathelin D, Bouyer F, et al. Tumor cells can escape DNA-damaging cisplatin through DNA endoreduplication and reversible polyploidy. Cell Biol Int. 2008;32:1031–43.
Mosieniak G, Sliwinska MA, Alster O, Strzeszewska A, Sunderland P, Piechota M et al. Polyploidy Formation in Doxorubicin-Treated Cancer Cells Can Favor Escape from Senescence. Neoplasia [Internet]. 2015;17:882–93. Available from: https://www.sciencedirect.com/science/article/pii/S1476558615001451
Wang Q, Wu PC, Dong DZ, Ivanova I, Chu E, Zeliadt S, et al. Polyploidy road to therapy-induced cellular senescence and escape. Int J Cancer. 2013;132:1505–15.
Bojko A, Staniak K, Czarnecka-Herok J, Sunderland P, Dudkowska M, Śliwińska MA et al. Improved Autophagic Flux in Escapers from Doxorubicin-Induced Senescence/Polyploidy of Breast Cancer Cells. Int J Mol Sci [Internet]. 2020;21. Available from: https://www.mdpi.com/1422-0067/21/17/6084
Gosselin K, Martien S, Pourtier A, Vercamer C, Ostoich P, Morat L, et al. Senescence-associated oxidative DNA damage promotes the generation of neoplastic cells. Cancer Res. 2009;69:7917–25.
Achuthan S, Santhoshkumar TR, Prabhakar J, Nair SA, Pillai MR. Drug-induced Senescence Generates Chemoresistant Stemlike Cells with Low Reactive Oxygen Species*. J Biol Chem [Internet]. 2011;286:37813–29. Available from: https://www.sciencedirect.com/science/article/pii/S0021925820508028
Moiseeva O, Guillon J, Ferbeyre G, Senescence. A program in the road to cell elimination and cancer. Semin Cancer Biol [Internet]. 2022;81:48–53. Available from: https://www.sciencedirect.com/science/article/pii/S1044579X20302777
Saleh T, Carpenter VJ, Bloukh S, Gewirtz DA. Targeting tumor cell senescence and polyploidy as potential therapeutic strategies. Semin Cancer Biol [Internet]. 2022;81:37–47. Available from: https://www.sciencedirect.com/science/article/pii/S1044579X20302704
Sikora E, Czarnecka-Herok J, Bojko A, Sunderland P. Therapy-induced polyploidization and senescence: Coincidence or interconnection? Semin Cancer Biol [Internet]. 2022;81:83–95. https://doi.org/10.1016/j.semcancer.2020.11.015
Fumagalli M, Rossiello F, Clerici M, Barozzi S, Cittaro D, Kaplunov JM, et al. Telomeric DNA damage is irreparable and causes persistent DNA-damage-response activation. Nat Cell Biol. 2012;14:355–65.
Walters HE, Deneka-Hannemann S, Cox LS. Reversal of phenotypes of cellular senescence by pan-mTOR inhibition. Aging. 2016;8:231–44.
Müller-Hermelink N, Braumüller H, Pichler B, Wieder T, Mailhammer R, Schaak K, et al. TNFR1 signaling and IFN-gamma signaling determine whether T cells induce tumor dormancy or promote multistage carcinogenesis. Cancer Cell. 2008;13:507–18.
Braumüller H, Wieder T, Brenner E, Aßmann S, Hahn M, Alkhaled M, et al. T-helper-1-cell cytokines drive cancer into senescence. Nature. 2013;494:361–5.
Yamada A, Sasada T, Noguchi M, Itoh K. Next-generation peptide vaccines for advanced cancer. Cancer Sci. 2013;104:15–21.
Melero I, Grimaldi AM, Perez-Gracia JL, Ascierto PA. Clinical development of immunostimulatory monoclonal antibodies and opportunities for combination. Clin cancer Res off J Am Assoc Cancer Res. 2013;19:997–1008.
Homann L, Rentschler M, Brenner E, Böhm K, Röcken M, Wieder T. IFN-γ and TNF induce senescence and a distinct senescence-Associated Secretory phenotype in Melanoma. Cells. 2022;11.
Kandhaya-Pillai R, Yang X, Tchkonia T, Martin GM, Kirkland JL, Oshima J. TNF-α/IFN-γ synergy amplifies senescence-associated inflammation and SARS-CoV-2 receptor expression via hyper-activated JAK/STAT1. Aging Cell. 2022;21:e13646.
Goddard ET, Linde MH, Srivastava S, Klug G, Shabaneh TB, Iannone S, et al. Immune evasion of dormant disseminated tumor cells is due to their scarcity and can be overcome by T cell immunotherapies. Cancer Cell. 2024;42:119–e13412.
Corthay A, Bakacs T, Thangavelu G, Anderson CC. Tackling cancer cell dormancy: Insights from immune models, and transplantation. Semin Cancer Biol [Internet]. 2022;78:5–16. Available from: https://www.sciencedirect.com/science/article/pii/S1044579X21000250
Trotter TN, Dagotto CE, Serra D, Wang T, Yang X, Acharya CR et al. Dormant tumors circumvent tumor-specific adaptive immunity by establishing a Treg-dominated niche via DKK3. JCI Insight. 2023;8.
Saudemont A, Quesnel B. In a model of tumor dormancy, long-term persistent leukemic cells have increased B7-H1 and B7.1 expression and resist CTL-mediated lysis. Blood. 2004;104:2124–33.
Wu X, Peng M, Huang B, Zhang H, Wang H, Huang B, et al. Immune microenvironment profiles of tumor immune equilibrium and immune escape states of mouse sarcoma. Cancer Lett. 2013;340:124–33.
Park S-Y, Nam J-S. The force awakens: metastatic dormant cancer cells. Exp Mol Med. 2020;52:569–81.
Morales-Valencia J, David G. The origins of cancer cell dormancy. Curr Opin Genet Dev [Internet]. 2022;74:101914. Available from: https://www.sciencedirect.com/science/article/pii/S0959437X22000235
Acosta JC, Banito A, Wuestefeld T, Georgilis A, Janich P, Morton JP et al. A complex secretory program orchestrated by the inflammasome controls paracrine senescence. Nat Cell Biol [Internet]. 2013;15:978–90. https://doi.org/10.1038/ncb2784
Acosta JC, O’Loghlen A, Banito A, Guijarro MV, Augert A, Raguz S, et al. Chemokine signaling via the CXCR2 receptor reinforces senescence. Cell. 2008;133:1006–18.
Passos JF, Nelson G, Wang C, Richter T, Simillion C, Proctor CJ, et al. Feedback between p21 and reactive oxygen production is necessary for cell senescence. Mol Syst Biol. 2010;6:347.
Saha J, Sarkar D, Pramanik A, Mahanti K, Adhikary A, Bhattacharyya S. PGE2-HIF1α reciprocal induction regulates migration, phenotypic alteration and immunosuppressive capacity of macrophages in tumor microenvironment. Life Sci [Internet]. 2020;253:117731. Available from: https://www.sciencedirect.com/science/article/pii/S0024320520304793
Sistigu A, Musella M, Galassi C, Vitale I, De Maria R. Tuning Cancer Fate: Tumor Microenvironment’s role in Cancer Stem Cell Quiescence and Reawakening. Front Immunol. 2020;11:2166.
Udagawa T. Tumor dormancy of primary and secondary cancers. APMIS. 2008;116:615–28.
Gao X-L, Zhang M, Tang Y-L, Liang X-H. Cancer cell dormancy: mechanisms and implications of cancer recurrence and metastasis. Onco Targets Ther. 2017;10:5219–28.
Demaria M, O’Leary MN, Chang J, Shao L, Liu S, Alimirah F, et al. Cellular Senescence promotes adverse effects of Chemotherapy and Cancer Relapse. Cancer Discov. 2017;7:165–76.
Krtolica A, Parrinello S, Lockett S, Desprez PY, Campisi J. Senescent fibroblasts promote epithelial cell growth and tumorigenesis: a link between cancer and aging. Proc Natl Acad Sci U S A. 2001;98:12072–7.
Hoare M, Ito Y, Kang T-W, Weekes MP, Matheson NJ, Patten DA, et al. NOTCH1 mediates a switch between two distinct secretomes during senescence. Nat Cell Biol. 2016;18:979–92.
Ito Y, Hoare M, Narita M. Spatial and temporal control of Senescence. Trends Cell Biol. 2017;27:820–32.
Romashin DD, Rusanov AL, Kozhin PM, Karagyaur MN, Tikhonova OV, Zgoda VG, et al. Impact of p53 knockdown on protein dataset of HaCaT cells. Data Br. 2022;42:108274.
Kubuschok B, Neumann F, Breit R, Sester M, Schormann C, Wagner C, et al. Naturally occurring T-cell response against mutated p21 ras oncoprotein in pancreatic cancer. Clin cancer Res off J Am Assoc Cancer Res. 2006;12:1365–72.
Ruhland MK, Loza AJ, Capietto A-H, Luo X, Knolhoff BL, Flanagan KC, et al. Stromal senescence establishes an immunosuppressive microenvironment that drives tumorigenesis. Nat Commun. 2016;7:11762.
Pereira BI, Devine OP, Vukmanovic-Stejic M, Chambers ES, Subramanian P, Patel N, et al. Senescent cells evade immune clearance via HLA-E-mediated NK and CD8(+) T cell inhibition. Nat Commun. 2019;10:2387.
Gabrilovich DI, Nagaraj S. Myeloid-derived suppressor cells as regulators of the immune system. Nat Rev Immunol. 2009;9:162–74.
Gabrilovich DI, Ostrand-Rosenberg S, Bronte V. Coordinated regulation of myeloid cells by tumours. Nat Rev Immunol. 2012;12:253–68.
Sturmlechner I, Zhang C, Sine CC, van Deursen E-J, Jeganathan KB, Hamada N, et al. p21 produces a bioactive secretome that places stressed cells under immunosurveillance. Science. 2021;374:eabb3420.
Loo TM, Kamachi F, Watanabe Y, Yoshimoto S, Kanda H, Arai Y, et al. Gut microbiota promotes obesity-Associated Liver Cancer through PGE(2)-Mediated suppression of Antitumor Immunity. Cancer Discov. 2017;7:522–38.
Sun Y, Campisi J, Higano C, Beer TM, Porter P, Coleman I et al. Treatment-induced damage to the tumor microenvironment promotes prostate cancer therapy resistance through WNT16B. Nat Med [Internet]. 2012;18:1359–68. https://doi.org/10.1038/nm.2890
Obenauf AC, Zou Y, Ji AL, Vanharanta S, Shu W, Shi H, et al. Therapy-induced tumour secretomes promote resistance and tumour progression. Nature. 2015;520:368–72.
Xu Q, Chiao P, Sun Y. Amphiregulin in Cancer: New Insights for Translational Medicine. Trends in Cancer [Internet]. 2016;2:111–3. Available from: https://www.sciencedirect.com/science/article/pii/S2405803316000327
Zaiss DMW, van Loosdregt J, Gorlani A, Bekker CPJ, Gröne A, Sibilia M et al. Amphiregulin Enhances Regulatory T Cell-Suppressive Function via the Epidermal Growth Factor Receptor. Immunity [Internet]. 2013;38:275–84. Available from: https://www.sciencedirect.com/science/article/pii/S1074761313000022
Xu Q, Long Q, Zhu D, Fu D, Zhang B, Han L, et al. Targeting amphiregulin (AREG) derived from senescent stromal cells diminishes cancer resistance and averts programmed cell death 1 ligand (PD-L1)-mediated immunosuppression. Aging Cell. 2019;18:1–25.
Alessio N, Aprile D, Squillaro T, Di Bernardo G, Finicelli M, Melone MA, et al. The senescence-associated secretory phenotype (SASP) from mesenchymal stromal cells impairs growth of immortalized prostate cells but has no effect on metastatic prostatic cancer cells. Aging. 2019;11:5817–28.
Gilbert LA, Hemann MT. DNA damage-mediated induction of a chemoresistant niche. Cell. 2010;143:355–66.
Sanoff HK, Deal AM, Krishnamurthy J, Torrice C, Dillon P, Sorrentino J, et al. Effect of cytotoxic chemotherapy on markers of molecular age in patients with breast cancer. J Natl Cancer Inst. 2014;106:dju057.
Jeon H-Y, Kim J-K, Ham SW, Oh S-Y, Kim J, Park J-B, et al. Irradiation induces glioblastoma cell senescence and senescence-associated secretory phenotype. Tumour Biol J Int Soc Oncodevelopmental Biol Med. 2016;37:5857–67.
Zingoni A, Cecere F, Vulpis E, Fionda C, Molfetta R, Soriani A, et al. Genotoxic stress induces Senescence-Associated ADAM10-Dependent release of NKG2D MIC ligands in multiple myeloma cells. J Immunol. 2015;195:736–48.
Muñoz DP, Yannone SM, Daemen A, Sun Y, Vakar-Lopez F, Kawahara M et al. Targetable mechanisms driving immunoevasion of persistent senescent cells link chemotherapy-resistant cancer to aging. JCI Insight. 2019;5.
Zingoni A, Vulpis E, Loconte L, Santoni A. NKG2D ligand shedding in response to stress: role of ADAM10. Front Immunol. 2020;11:447.
Baker DJ, Wijshake T, Tchkonia T, LeBrasseur NK, Childs BG, van de Sluis B, et al. Clearance of p16Ink4a-positive senescent cells delays ageing-associated disorders. Nature. 2011;479:232–6.
Baker DJ, Childs BG, Durik M, Wijers ME, Sieben CJ, Zhong J, et al. Naturally occurring p16(Ink4a)-positive cells shorten healthy lifespan. Nature. 2016;530:184–9.
Baar MP, Brandt RMC, Putavet DA, Klein JDD, Derks KWJ, Bourgeois BRM, et al. Targeted apoptosis of senescent cells restores tissue homeostasis in response to Chemotoxicity and Aging. Cell. 2017;169:132–e14716.
Meloni F, Morosini M, Solari N, Passadore I, Nascimbene C, Novo M, et al. Foxp3 expressing CD4 + CD25 + and CD8 + CD28- T regulatory cells in the peripheral blood of patients with lung cancer and pleural mesothelioma. Hum Immunol. 2006;67:1–12.
Ye S-W, Wang Y, Valmori D, Ayyoub M, Han Y, Xu X-L, et al. Ex-vivo analysis of CD8 + T cells infiltrating colorectal tumors identifies a major effector-memory subset with low perforin content. J Clin Immunol. 2006;26:447–56.
Webb JR, Wick DA, Nielsen JS, Tran E, Milne K, McMurtrie E, et al. Profound elevation of CD8 + T cells expressing the intraepithelial lymphocyte marker CD103 (alphaE/beta7 integrin) in high-grade serous ovarian cancer. Gynecol Oncol. 2010;118:228–36.
Tsukishiro T, Donnenberg AD, Whiteside TL. Rapid turnover of the CD8(+)CD28(-) T-cell subset of effector cells in the circulation of patients with head and neck cancer. Cancer Immunol Immunother. 2003;52:599–607.
Montes CL, Chapoval AI, Nelson J, Orhue V, Zhang X, Schulze DH, et al. Tumor-induced senescent T cells with suppressor function: a potential form of tumor immune evasion. Cancer Res. 2008;68:870–9.
Li H, Wu K, Tao K, Chen L, Zheng Q, Lu X, et al. Tim-3/galectin-9 signaling pathway mediates T-cell dysfunction and predicts poor prognosis in patients with hepatitis B virus-associated hepatocellular carcinoma. Hepatology. 2012;56:1342–51.
Ye J, Peng G. Controlling T cell senescence in the tumor microenvironment for tumor immunotherapy. Oncoimmunology. 2015;4:e994398.
Lian J, Yue Y, Yu W, Zhang Y. Immunosenescence: a key player in cancer development. J Hematol Oncol. 2020;13:151.
Wherry EJ, Kurachi M. Molecular and cellular insights into T cell exhaustion. Nat Rev Immunol. 2015;15:486–99.
Duraiswamy J, Kaluza KM, Freeman GJ, Coukos G. Dual blockade of PD-1 and CTLA-4 combined with tumor vaccine effectively restores T-cell rejection function in tumors. Cancer Res. 2013;73:3591–603.
Sakuishi K, Apetoh L, Sullivan JM, Blazar BR, Kuchroo VK, Anderson AC. Targeting Tim-3 and PD-1 pathways to reverse T cell exhaustion and restore anti-tumor immunity. J Exp Med. 2010;207:2187–94.
Ye J, Huang X, Hsueh EC, Zhang Q, Ma C, Zhang Y, et al. Human regulatory T cells induce T-lymphocyte senescence. Blood. 2012;120:2021–31.
Ye J, Ma C, Hsueh EC, Eickhoff CS, Zhang Y, Varvares MA, et al. Tumor-derived γδ regulatory T cells suppress innate and adaptive immunity through the induction of immunosenescence. J Immunol. 2013;190:2403–14.
Debacq-Chainiaux F, Erusalimsky JD, Campisi J, Toussaint O. Protocols to detect senescence-associated beta-galactosidase (SA-betagal) activity, a biomarker of senescent cells in culture and in vivo. Nat Protoc. 2009;4:1798–806.
Brenchley JM, Karandikar NJ, Betts MR, Ambrozak DR, Hill BJ, Crotty LE, et al. Expression of CD57 defines replicative senescence and antigen-induced apoptotic death of CD8 + T cells. Blood. 2003;101:2711–20.
Heffner M, Fearon DT. Loss of T cell receptor-induced Bmi-1 in the KLRG1(+) senescent CD8(+) T lymphocyte. Proc Natl Acad Sci U S A. 2007;104:13414–9.
Yang OO, Lin H, Dagarag M, Ng HL, Effros RB, Uittenbogaart CH. Decreased perforin and granzyme B expression in senescent HIV-1-specific cytotoxic T lymphocytes. Virology. 2005;332:16–9.
Liu X, Mo W, Ye J, Li L, Zhang Y, Hsueh EC, et al. Regulatory T cells trigger effector T cell DNA damage and senescence caused by metabolic competition. Nat Commun. 2018;9:249.
Li L, Liu X, Sanders KL, Edwards JL, Ye J, Si F, et al. TLR8-Mediated metabolic control of human Treg function: a mechanistic target for Cancer Immunotherapy. Cell Metab. 2019;29:103–e1235.
Ye J, Ma C, Hsueh EC, Dou J, Mo W, Liu S, et al. TLR8 signaling enhances tumor immunity by preventing tumor-induced T-cell senescence. EMBO Mol Med. 2014;6:1294–311.
Henson SM, Lanna A, Riddell NE, Franzese O, Macaulay R, Griffiths SJ, et al. p38 signaling inhibits mTORC1-independent autophagy in senescent human CD8+ T cells. J Clin Invest. 2014;124:4004–16.
Valmori D, Scheibenbogen C, Dutoit V, Nagorsen D, Asemissen AM, Rubio-Godoy V, et al. Circulating tumor-reactive CD8(+) T cells in melanoma patients contain a CD45RA(+)CCR7(-) effector subset exerting ex vivo tumor-specific cytolytic activity. Cancer Res. 2002;62:1743–50.
Vallejo AN, Mueller RG, Hamel DLJ, Way A, Dvergsten JA, Griffin P, et al. Expansions of NK-like αβT cells with chronologic aging: novel lymphocyte effectors that compensate for functional deficits of conventional NK cells and T cells. Ageing Res Rev. 2011;10:354–61.
Pereira BI, De Maeyer RPH, Covre LP, Nehar-Belaid D, Lanna A, Ward S, et al. Sestrins induce natural killer function in senescent-like CD8(+) T cells. Nat Immunol. 2020;21:684–94.
Yamada T, Kanoh M, Nabe S, Yasuoka T, Suzuki J, Matsumoto A, et al. Menin plays a critical role in the regulation of the Antigen-Specific CD8 + T cell response upon Listeria infection. J Immunol. 2016;197:4079–89.
Suzuki J, Yamada T, Inoue K, Nabe S, Kuwahara M, Takemori N et al. The tumor suppressor menin prevents effector CD8 T-cell dysfunction by targeting mTORC1-dependent metabolic activation. Nat Commun [Internet]. 2018;9:3296. https://doi.org/10.1038/s41467-018-05854-6
Kakuda T, Suzuki J, Matsuoka Y, Kikugawa T, Saika T, Yamashita M. Senescent CD8(+) T cells acquire NK cell-like innate functions to promote antitumor immunity. Cancer Sci. 2023;114:2810–20.
Chen C, Xu P. Cellular functions of cGAS-STING signaling. Trends Cell Biol. 2023;33:630–48.
Hopfner K-P, Hornung V. Molecular mechanisms and cellular functions of cGAS-STING signalling. Nat Rev Mol Cell Biol. 2020;21:501–21.
Dou Z, Ghosh K, Vizioli MG, Zhu J, Sen P, Wangensteen KJ, et al. Cytoplasmic chromatin triggers inflammation in senescence and cancer. Nature. 2017;550:402–6.
Glück S, Guey B, Gulen MF, Wolter K, Kang T-W, Schmacke NA, et al. Innate immune sensing of cytosolic chromatin fragments through cGAS promotes senescence. Nat Cell Biol. 2017;19:1061–70.
Mackenzie KJ, Carroll P, Martin C-A, Murina O, Fluteau A, Simpson DJ, et al. cGAS surveillance of micronuclei links genome instability to innate immunity. Nature. 2017;548:461–5.
Victorelli S, Salmonowicz H, Chapman J, Martini H, Vizioli MG, Riley JS, et al. Apoptotic stress causes mtDNA release during senescence and drives the SASP. Nature. 2023;622:627–36.
Yang H, Wang H, Ren J, Chen Q, Chen ZJ. cGAS is essential for cellular senescence. Proc Natl Acad Sci U S A. 2017;114:E4612–20.
Walker MM, Kim S, Crisler WJ, Nguyen K, Lenz LL, Cambier JC, et al. Selective loss of responsiveness to exogenous but not endogenous cyclic-dinucleotides in mice expressing STING-R231H. Front Immunol. 2020;11:238.
Omer A, Di Marco S, Gallouzi I-E. The senescence-associated secretory phenotype as a driver of tumor growth: does G3BP1 hold the key? Mol Cell Oncol. 2021;8:1850161.
Omer A, Barrera MC, Moran JL, Lian XJ, Di Marco S, Beausejour C et al. G3BP1 controls the senescence-associated secretome and its impact on cancer progression. Nat Commun [Internet]. 2020;11:4979. https://doi.org/10.1038/s41467-020-18734-9
Moiseeva O, Mallette FA, Mukhopadhyay UK, Moores A, Ferbeyre G. DNA damage signaling and p53-dependent senescence after prolonged beta-interferon stimulation. Mol Biol Cell. 2006;17:1583–92.
Yu Q, Katlinskaya YV, Carbone CJ, Zhao B, Katlinski KV, Zheng H, et al. DNA-damage-induced type I interferon promotes senescence and inhibits stem cell function. Cell Rep. 2015;11:785–97.
Paffenholz SV, Salvagno C, Ho Y-J, Limjoco M, Baslan T, Tian S et al. Senescence induction dictates response to chemo- and immunotherapy in preclinical models of ovarian cancer. Proc Natl Acad Sci U S A. 2022;119.
Mowat C, Mosley SR, Namdar A, Schiller D, Baker K. Anti-tumor immunity in mismatch repair-deficient colorectal cancers requires type I IFN-driven CCL5 and CXCL10. J Exp Med. 2021;218.
Schadt L, Sparano C, Schweiger NA, Silina K, Cecconi V, Lucchiari G, et al. Cancer-Cell-intrinsic cGAS expression mediates Tumor Immunogenicity. Cell Rep. 2019;29:1236–e12487.
Xia T, Konno H, Ahn J, Barber GN. Deregulation of STING Signaling in Colorectal Carcinoma constrains DNA damage responses and correlates with tumorigenesis. Cell Rep. 2016;14:282–97.
Bu Y, Liu F, Jia Q-A, Yu S-N. Decreased expression of TMEM173 predicts poor prognosis in patients with Hepatocellular Carcinoma. PLoS ONE. 2016;11:e0165681.
Xuan C, Hu R. Chemical Biology perspectives on STING agonists as Tumor Immunotherapy. ChemMedChem. 2023;202300405.
Chibaya L, Snyder J, Ruscetti M. Senescence and the tumor-immune landscape: implications for cancer immunotherapy. Semin Cancer Biol. 2022;86:827–45.
Wang L, Lankhorst L, Bernards R. Exploiting senescence for the treatment of cancer. Nat Rev Cancer. 2022;22:340–55.
Zhang J-W, Zhang D, Yu B-P. Senescent cells in cancer therapy: why and how to remove them. Cancer Lett. 2021;520:68–79.
Ovadya Y, Krizhanovsky V. Strategies targeting cellular senescence. J Clin Invest. 2018;128:1247–54.
Fuhrmann-Stroissnigg H, Ling YY, Zhao J, McGowan SJ, Zhu Y, Brooks RW et al. Identification of HSP90 inhibitors as a novel class of senolytics. Nat Commun [Internet]. 2017;8:422. https://doi.org/10.1038/s41467-017-00314-z
Fionda C, Soriani A, Malgarini G, Iannitto ML, Santoni A, Cippitelli M. Heat shock protein-90 inhibitors increase MHC class I-related chain A and B ligand expression on multiple myeloma cells and their ability to trigger NK cell degranulation. J Immunol. 2009;183:4385–94.
Böll B, Eltaib F, Reiners KS, von Tresckow B, Tawadros S, Simhadri VR, et al. Heat shock protein 90 inhibitor BIIB021 (CNF2024) depletes NF-kappaB and sensitizes Hodgkin’s lymphoma cells for natural killer cell-mediated cytotoxicity. Clin cancer Res off J Am Assoc Cancer Res. 2009;15:5108–16.
Aird KM, Iwasaki O, Kossenkov AV, Tanizawa H, Fatkhutdinov N, Bitler BG, et al. HMGB2 orchestrates the chromatin landscape of senescence-associated secretory phenotype gene loci. J Cell Biol. 2016;215:325–34.
Capell BC, Drake AM, Zhu J, Shah PP, Dou Z, Dorsey J, et al. MLL1 is essential for the senescence-associated secretory phenotype. Genes Dev. 2016;30:321–36.
Georgilis A, Klotz S, Hanley CJ, Herranz N, Weirich B, Morancho B, et al. PTBP1-Mediated alternative splicing regulates the inflammatory secretome and the pro-tumorigenic effects of senescent cells. Cancer Cell. 2018;34:85–e1029.
Toso A, Revandkar A, Di Mitri D, Guccini I, Proietti M, Sarti M, et al. Enhancing chemotherapy efficacy in Pten-deficient prostate tumors by activating the senescence-associated antitumor immunity. Cell Rep. 2014;9:75–89.
Xu M, Tchkonia T, Ding H, Ogrodnik M, Lubbers ER, Pirtskhalava T et al. JAK inhibition alleviates the cellular senescence-associated secretory phenotype and frailty in old age. Proc Natl Acad Sci [Internet]. 2015;112:E6301–10. Available from: https://www.pnas.org/doi/abs/https://doi.org/10.1073/pnas.1515386112
Zhang Q, Chen C, Xia B, Xu P. Chemical regulation of the cGAS-STING pathway. Curr Opin Chem Biol [Internet]. 2022;69:102170. Available from: https://www.sciencedirect.com/science/article/pii/S1367593122000552
Herranz N, Gallage S, Mellone M, Wuestefeld T, Klotz S, Hanley CJ et al. mTOR regulates MAPKAPK2 translation to control the senescence-associated secretory phenotype. Nat Cell Biol [Internet]. 2015;17:1205–17. https://doi.org/10.1038/ncb3225
Laberge R-M, Sun Y, Orjalo AV, Patil CK, Freund A, Zhou L, et al. MTOR regulates the pro-tumorigenic senescence-associated secretory phenotype by promoting IL1A translation. Nat Cell Biol. 2015;17:1049–61.
Lau L, Porciuncula A, Yu A, Iwakura Y, David G. Uncoupling the Senescence-Associated Secretory phenotype from cell cycle exit via Interleukin-1 inactivation unveils its Protumorigenic Role. Mol Cell Biol. 2019;39.
Arnold DD, Yalamanoglu A, Boyman O. Systematic review of Safety and Efficacy of IL-1-Targeted Biologics in Treating Immune-mediated disorders. Front Immunol. 2022;13.
Malcova H, Milota T, Strizova Z, Cebecauerova D, Striz I, Sediva A, et al. Interleukin-1 blockade in Polygenic Autoinflammatory disorders: where are we now? Front Pharmacol. 2021;11:1–21.
Lopetuso LR, Cuomo C, Mignini I, Gasbarrini A, Papa A. Focus on Anti-Tumour Necrosis Factor (TNF)-α-Related Autoimmune Diseases. Int J Mol Sci [Internet]. 2023;24. Available from: https://www.mdpi.com/1422-0067/24/9/8187
Aliyu M, Zohora FT, Anka AU, Ali K, Maleknia S, Saffarioun M, et al. Interleukin-6 cytokine: an overview of the immune regulation, immune dysregulation, and therapeutic approach. Int Immunopharmacol. 2022;111:109130.
Muñoz-Espín D, Rovira M, Galiana I, Giménez C, Lozano-Torres B, Paez-Ribes M et al. A versatile drug delivery system targeting senescent cells. EMBO Mol Med. 2018;10.
Paez-Ribes M, González-Gualda E, Doherty GJ, Muñoz-Espín D. Targeting senescent cells in translational medicine. EMBO Mol Med. 2019;11:e10234.
Shin E, Bak SH, Park T, Kim JW, Yoon S-R, Jung H, et al. Understanding NK cell biology for harnessing NK cell therapies: targeting cancer and beyond. Front Immunol. 2023;14:1192907.
Walle T, Kraske JA, Liao B, Lenoir B, Timke C, von Halbach B. Radiotherapy orchestrates natural killer cell dependent antitumor immune responses through CXCL8. Sci Adv. 2022;8:eabh4050.
Ruggeri L, Urbani E, André P, Mancusi A, Tosti A, Topini F, et al. Effects of anti-NKG2A antibody administration on leukemia and normal hematopoietic cells. Haematologica. 2016;101:626–33.
Ferrari de Andrade L, Tay RE, Pan D, Luoma AM, Ito Y, Badrinath S, et al. Antibody-mediated inhibition of MICA and MICB shedding promotes NK cell-driven tumor immunity. Science. 2018;359:1537–42.
Kim KM, Noh JH, Bodogai M, Martindale JL, Yang X, Indig FE, et al. Identification of senescent cell surface targetable protein DPP4. Genes Dev. 2017;31:1529–34.
Amor C, Feucht J, Leibold J, Ho Y-J, Zhu C, Alonso-Curbelo D, et al. Senolytic CAR T cells reverse senescence-associated pathologies. Nature. 2020;583:127–32.
Yang D, Sun B, Li S, Wei W, Liu X, Cui X, et al. NKG2D-CAR T cells eliminate senescent cells in aged mice and nonhuman primates. Sci Transl Med. 2023;15:eadd1951.
Yoshida S, Nakagami H, Hayashi H, Ikeda Y, Sun J, Tenma A et al. The CD153 vaccine is a senotherapeutic option for preventing the accumulation of senescent T cells in mice. Nat Commun [Internet]. 2020;11:2482. https://doi.org/10.1038/s41467-020-16347-w
Wang TW, Johmura Y, Suzuki N, Omori S, Migita T, Yamaguchi K et al. Blocking PD-L1–PD-1 improves senescence surveillance and ageing phenotypes. Nature [Internet]. 2022;611:358–64. https://doi.org/10.1038/s41586-022-05388-4
Onorati A, Havas AP, Lin B, Rajagopal J, Sen P, Adams PD, et al. Upregulation of PD-L1 in Senescence and Aging. Mol Cell Biol. 2022;42:e0017122.
Wang T, Liu W, Shen Q, Tao R, Li C, Shen Q, et al. Combination of PARP inhibitor and CDK4/6 inhibitor modulates cGAS/STING-dependent therapy-induced senescence and provides one-two punch opportunity with anti-PD-L1 therapy in colorectal cancer. Cancer Sci. 2023;114:4184–201.
Chaib S, López-Domínguez JA, Lalinde M, Prats N, Marin I, Meyer K et al. The efficacy of chemotherapy is limited by intratumoural senescent cells that persist through the upregulation of PD-L2. bioRxiv [Internet]. 2022;2022.11.04.501681. Available from: https://www.biorxiv.org/content/https://doi.org/10.1101/2022.11.04.501681v1%0Ahttps://www.biorxiv.org/content/10.1101/2022.11.04.501681v1.abstract
Garcia-Diaz A, Shin DS, Moreno BH, Saco J, Escuin-Ordinas H, Rodriguez GA, et al. Interferon receptor signaling pathways regulating PD-L1 and PD-L2 expression. Cell Rep. United States; 2019. p. 3766.
Uceda-Castro R, Margarido AS, Cornet L, Vegna S, Hahn K, Song J-Y, et al. Re-purposing the pro-senescence properties of doxorubicin to introduce immunotherapy in breast cancer brain metastasis. Cell Rep Med. 2022;3:100821.
Acknowledgements
Not applicable.
Funding
This research was funded by AIRC-IG 2021 (ID 25680), AIRC-IG 2023 (ID 29244), PRIN-2022 (20223RRASS), Sapienza- University of Rome (RG12218166D295B0 and RG123188A3D8C26E), CNR project FOE-2021 DBA.AD005.225, the European Union - NextGenerationEU through the Italian Ministry of University and Research under PNRR - M4C2-I1.3 Project PE_00000019 “HEAL ITALIA”, European Union - Next Generation EU, in the context of The National Recovery and Resilience Plan, Investment Partenariato Esteso PE8 “Conseguenze e sfide dell’invecchiamento”, Project Age-It (Ageing Well in an Ageing Society).
Author information
Authors and Affiliations
Contributions
AZ, FA, SS, ASa, MC, ASo contributed to write the manuscript, collected the related papers and designed the figures. All authors reviewed the final version of the article before giving their approval for publication.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
All authors have approved to publish this manuscript.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Zingoni, A., Antonangeli, F., Sozzani, S. et al. The senescence journey in cancer immunoediting. Mol Cancer 23, 68 (2024). https://doi.org/10.1186/s12943-024-01973-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12943-024-01973-5