
Abstract
Analogous to DNA methylation and histone modification, RNA modification, as another epigenetic layer, plays an important role in many diseases, especially in tumours. As the most common form of RNA modification, m6A methylation has attracted increasing research interest in recent years. m6A is catalysed by RNA methyltransferases METTL3, METTL14 and WTAP (writers), m6A is removed by the demethylases FTO and ALKBH5 (erasers) and interacts with m6A-binding proteins, such as YT521-B homology (YTH) domain-containing proteins. This article reviews recent studies on methylation modification of m6A in gastrointestinal tract cancers.
Similar content being viewed by others

Introduction
In recent years, epigenetics has attracted the attention of the research community. Epigenetics is a study of reversible, inheritable phenotypes that do not involve changes in nuclear DNA sequences [1]. Although the full scope of epigenetics has not yet been determined, it is generally defined as chemical modification that mainly includes DNA and RNA methylation, histone modification, noncoding RNA modification and chromatin rearrangement. In epigenetic modification, DNA methylation and histone modification have been well studied. For example, 5-methylcytosine methylation in DNA has affected gene expression in many tumours. Significant advances have been achieved in recent years in the study of methylated drugs, such as demethylation drugs Decitabine and Azacitidine and histone deacetylase inhibitor Sedamine, which provides additional strategies for treatment of clinical diseases [2, 3]. In addition to DNA and histone methylation, another level of epigenetic regulation, namely, RNA methylation, has become a hot topic in biosciences over the past decade. Common RNA methylation sites include 5-methylcytosine (m5C), 7-methylguanosine (m7G), m1G, m2G, m6G, N1-methyladenosine (m1A) and m6A. m5C modification promotes splicing and translation [4]. m1G, m2G and m1A modifications at the first or second codon repress protein synthesis [5,6,7], and tRNA m7G methylation is required for mRNA translation into proteins [8]. m6A is the most common among various RNA modifications [9] and has critical roles in cancer pathogenesis. In this review, we focus on the relationship between RNA m6A methylation and gastrointestinal cancer, especially their role, mechanism and potential clinical application as biomarkers and therapeutic targets for gastrointestinal cancer.
RNA m6A methylation
More than 100 kinds of chemical modifications of RNA, including mRNAs, rRNAs, tRNAs, snRNAs and snoRNAs, have been identified in organisms [10]. Among them, m6A, discovered in the 1970s, is the most abundant internal modification of mRNA in most eukaryotes [11] and involves almost all stages of RNA life cycle, including RNA transcription, exporting through nuclear translation and degradation [12,13,14,15]. About 0.1–0.4% of adenosine in isolated RNA is modified by m6A in mammals [16]. Transcriptome-wide research reveals that m6A modification may affect more than 7000 mRNAs in individual transcriptomes of mammalian cells. m6A modifications are enriched in the 3′-untranslated regions (UTRs) near the stop codons of mRNAs and with a consensus sequence of RRACH (R = G or A; H = A, C, or U) [17] (Fig. 1). Highly conserved m6A is widely present in most eukaryotic species (from yeast, plants and fruit flies to mammals) and viral mRNAs and plays a key regulatory role in post-transcriptional mRNA processing and metabolism. Several lncRNAs also accept m6A modification [18].

Chemical modification of eukaryotic mRNA. Schematic diagram of common chemical modifications of eukaryotic mRNA transcripts
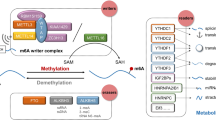
Similar to DNA and histone methylation, m6A modifications are dynamic and reversible and exert biological effects that are mainly mediated by ‘writers’, ‘erasers’ and ‘reader’ proteins (Fig. 2). Reversibility means that RNA can be methylated under the action of methyltransferases and demethylated under the action of demethylases; this phenomenon is called dynamic balance. Writers traditionally consist of methyltransferase-like 3 and 14 proteins (METTL3 and METTL14) and their cofactors WTAP (Wilms tumour suppressor-1-associated protein) [19,20,21]. METTL3 and METTL14 contain an S-adenosylmethionine-binding motif. METTL3 and METTL14 are co-located in nuclear spots and form stable complexes in a 1:1 ratio [22]. METTL3 is a major catalytic enzyme with functions reminiscent of the N6-adenine methyltransferase system [23]. METTL14 is a pseudomethyltransferase that stabilizes METTL3 and recognizes target RNA [24]. WTAP is the main regulatory component of the m6A methylation complex. WTAP interacts with METTL3 and METTL14 and helps them to be localised in nuclear spots [20]. Writers also include methyltransferase-like 16 (METTL16) [25], KIAA1429 [21] and RBM15 [26]. Demethylation, which is the removal of methyl groups, is also important. Demethylation is achieved by another enzyme family called demethylases (erasers), mainly including FTO and ALKBH5. FTO can sequentially oxidise m6A into N6-hydroxymethyladeosine and N6-formyladenosine, which are moderately stable and can be hydrolysed into adenine. Given that FTO was identified as the first RNA demethylase [27], RNA methylation has gradually gained research attention. ALKBH5, an FTO homologue [28], ensures the equilibrium of m6A modification in the transcriptome. In addition to writers and erasers, another important group is readers, which can recognise these modifications, bind to them, and carry out different biological functions [29]. Readers can be recognised by proteins containing the YT521B homology (YTH) domain. The YTH domain in human cells, including YTH domain family (YTHDF1–3), YTH domain-containing 1 (YTHDC1) and YTH domain-containing 2 (YTHDC2), have conserved m6A binding domain and preferentially bind to m6A-modified RNA in RRm6ACH consensus sequence [11]. Moreover, IGF2BP1–3 is a common reader. YTHDF2, the first characteristic m6A reader, can result in the localisation of binding mRNA to the decay site of RNA [30]. YTHDF1 was initially demonstrated to bind to m6A sites around the stop codon and improve RNA translation efficiency by interacting with translation initiation factor eif3 [31]. YTHDF3 plays a fine-tuning role in the RNA accessibility of YTHDF1 and YTHDF2.

Mechanism of m6A methylation m6A-RNA methylation is regulated by its “writer”, “eraser” and “reader”, the writer refers to the m6A-methylase complex, mainly including METTL3, METTL14, METTL16 and WTAP. The eraser is an m6A-demethylase complex, including FTO and ALKBH5. The reader is a protein that binds to m6A and includes proteins containing the YTH domain (YTHDF1, YTHDF2, YTHDF3, YTHDC1, YTHDC2), which involving splicing, export, translation, RNA decay
Roles of RNA m6A in gastrointestinal tract cancers
Emerging evidence suggests that m6A is closely associated with the progression of gastrointestinal cancer, including its tumourigenesis, metastasis and angiogenesis. Herein, we briefly review recent studies of m6A methylation in gastrointestinal tract cancers (Table 1).
Liver cancer
Diverse and reversible m6A modification on RNAs emerges as a new layer of regulation in liver cancer. In the study of Ma et al. [32], the expression level of m6A in HCC decreased firstly, especially in metastatic HCC. METTL14 is an important factor for the aberrant expression of m6A. The decrease in METTL14 expression was proven to be related to HCC metastasis in vivo and in vitro. In addition, the target genes of METTL14 during HCC metastasis have been explored. One of these genes is microRNA 126 (micro-126), which plays an important role in cancer metastasis by acting as a tumour inhibitor. The reduction of METTL14 interacts with microprocessor protein DGCR8 and positively regulates pri-microRNA-126 process in an m6A-dependent manner, resulting in reduced expression of microRNA-126. Yang and coworkers [33] elucidated that miR-145 can increase the m6A level by downregulating YTHDF2 in hepatocellular carcinoma cells. YTHDF2 can recognise mRNA m6A site to regulate mRNA degradation [30, 43]. In a previous work [33], miR-145 can inhibit the proliferation of HepG2 cells, suggesting that miR-145-increased methylated mRNAs may be involved in cell apoptosis. Chen and colleagues [44] demonstrated that the expression of METTL3 in human liver cancer is frequently up-regulated and contributes to the progression of liver cancer. METTL3 inhibits the expression of SOCS2 in liver cancer through the m6A-YTHDF2-dependent mechanism. Studies have shown that SOCS2 acts as a tumour suppressor but only inhibits the proliferation, migration or drying of different cancer types, including leukaemia, oral squamous cell carcinoma and, more recently HCC [45]. Cheng et al. [46] showed that m6A modification also plays an important role in HCC.
Gastric cancer (GC)
GC is one of the most common malignancies worldwide [47]. Among Chinese men, the morbidity of GC is second. The mortality of GC in the Chinese crowd is also in second [48]. Lin and collaborators [34] found that METTL3, as an oncogene, might be a potential target for treatment of human GC. Zhang and coworkers [35] demonstrated that reduced m6A methylation activates oncogenic Wnt/PI3K-AKT signalling and promotes malignant phenotypes in GC cells. Zhang et al. [49] showed that NEAT1 acts as a LncRNA and undergoes m6A methylation in GC. As an eraser, ALKBH5 reduced the level of m6A methylation of NEAT1, but the expression of ALKBH5 and NEAT1 was positively correlated. Therefore, NEAT1 is upregulated with decreasing m6A methylation of NEAT1; this phenomenon promotes the malignant phenotype of GC because NEAT1 can act as a scaffold to affect EZH2 expression, tumour invasion and metastasis. Liu et al. [36] found that METTL3 is a factor of poor prognosis in patients with GC, and the expression level of METTL3 is related to tumour stage and grade. In addition, METTL3 knockdown reduced α-smooth muscle actin. Hence, METTL3 could serve as an oncogene in the tumourigenesis of GC.
Colorectal cancer (CRC)
CRC is one of the most common malignant tumours in humans and has increasing mortality and morbidity worldwide [50]. Although the level of medical treatment continues to improve, the 5-year relative survival rate of patients is only 64.9% [51]. Li et al. [37] found that METTL3 is highly expressed in metastatic CRC, and its downstream gene is SOX2. SOX2 is recognised as cancer stem-like cells (CSCs) [52]. CSCs are a group of tumour cells with self-renewal ability and multidirectional differentiation potential and related to strong possibility of tumour occurrence and metastasis [53]. Mechanistically, the methylation of SOX2 transcripts in CRC cells, especially in the CD region, leads to the specific recognition of IGF2BP2 by m6A readers, thereby preventing the degradation of SOX2 mRNA. In contrast to Li’s conclusion, Deng [38] believed that METTL3 plays an anti-oncogene role in CRC and is a favourable prognostic factor in CRC. Specifically, METTL3 targets downstream p38/ERK signalling pathways. Nishizawa and colleagues [39] elucidated that over-expression of YTHDF1 in CRC cells compared with that in noncancer cells is associated with malignant phenotype and poor prognosis. Moreover, c-myc, an oncogenic transcription factor, activates the transcriptional expression of the YTHDF1 gene. Bai and coworkers [40] found that the mRNA and protein levels of YTHDF1 are overexpressed in CRC. In addition, the expression level of YTHDF1 is related to tumour depth and size. Knockdown of YTHDF1 inhibits the activity of the Wnt/beta-catenin pathway, of which the Wnt pathway component can lead to CRC.
Pancreatic cancer
Pancreatic neoplasms are one of the few cancers with a mortality rate approaching 100% [54]. The aetiology and screening tests for this highly lethal disease are not well defined; as such, identifying genetic factors that contribute to the development of this cancer is important [55]. Chen and colleagues [41] found that the expression level of YTHDF2 in pancreatic cancer tissues is higher than that in normal tissues at the mRNA and protein levels; moreover, YTHDF2 is an independent influencing factor for the deterioration of patients’ condition. YTHDF2 knockdown promoted the expression of YAP, suggesting that YTHDF2 may regulate EMT through YAP signalling. A large number of studies have shown that EMT can promote the migration and proliferation of tumour cells. Tang [42] demonstrated that FTO, a primary demethylase in vivo, was overexpressed in pancreatic cancer cells compared with that in normal pancreatic epithelial cells. Knockdown of FTO resulted in impaired proliferation and increased apoptosis of pancreatic cancer cells. These observations suggest that FTO is essential for the proliferation of pancreatic cancer cells.
Potential application of RNA m6A in gastrointestinal tract cancers
RNA m6A as biomarker in gastrointestinal tract cancers
An increasing number of studies have shown that m6A may have great potential as a biomarker for diagnosis, prognosis prediction and therapeutic evaluation of digestive tract tumours. Zhou et al. [56] firstly found that the abnormal expression of METTL3 or YTHDF1 is associated with overall survival in HCC and confirmed its protein expression. Moreover, METTL3 and YTHDF1 were upregulated in HCC. The low METTL3/YTHDF1 group had better prognosis. In summary, the combination of METTL3 and YTHDF1 can be used as biomarker to reflect the malignant degree of liver cancer and evaluate its prognosis. Zhao et al. [57] found that low YTHDF1 expression level is associated with improved survival of patients with HCC. Therefore, YTHDF1 may be a marker of HCC.
In GC, the abnormal expression of FTO has significant prognostic value; hence, FTO may play important roles in GC progression and metastasis [58]. Similarly to the report of Li, Xu et al. [59] concluded that FTO expression might play an important role in promoting the occurrence of GC and might be an important molecular marker for the diagnosis and prognosis of this disease. They found that the expression level of FTO is related to poor differentiation, lymph node metastasis, TNM staging and poor prognosis. Therefore, FTO expression may play an important role in promoting the occurrence of GC and may be an important molecular marker for the diagnosis and prognosis of patients with GC. In pancreatic cancer, ALKBH5 is an independent prognostic factor [60].
RNA m6A as therapeutic targets in in gastrointestinal tract cancers
The important roles of m6A in gastrointestinal cancers suggest that it may be exploited as a therapeutic target. For treatment of common digestive tract tumours, radiotherapy and chemotherapy have a pivotal position; however, chemo- and radioresistance lead to unsatisfactory treatment outcomes. Studies have shown that m6A methylation is important in treatment of tumours, especially in targeted therapy. In the study of Zhu et al. [61], impaired autophagy degradation of lncRNA ARHGAP5-AS1 in chemoresistant cancer cells may promote chemoresistance. The transcription of ARHGAP5 in the nucleus is activated by recruiting METTL3, and the m6A modification of ARHGAP5 is stimulated to stabilise the mRNA of ARHGAP5 in the cytoplasm. Therefore, m6A methylation may play an important role in the chemotherapeutic resistance of GC and deserves further study and exploration. Taketo and coworkers [62] established METTL3-knockdown pancreatic cancer cell line by using short hairpin RNA. METTL3-depleted cells showed higher sensitivity to anticancer reagents, such as Gemcitabine, 5-Fluorouracil, Cisplatin and irradiation. In summary, METTL3 is associated with therapeutic resistance and a potential therapeutic target for pancreatic cancer. A previous study suggested that YTHDF1 is a potential therapeutic target for anti-cancer immunotherapy [63]. The down-regulation of the YTHDF1 gene can inhibit tumour proliferation and sensitivity to the exposure of fluorouracil, oxaliplatin and other anticancer drugs [39]. Chen et al. [64] found that rhein, an FTO inhibitor, prevents FTO from binding to m6A substrates by binding to the active site of FTO, thereby increasing the cell content of m6A in mRNA.
Discussion
RNA epigenetics has become a hot topic in recent years. Among more than 100 different RNA modifications, m6A is the most abundant and influences the processing of lncRNA and miRNA. m6A methylation plays an important role in digestive tract tumours. This article describes the role, mechanism and application of m6A from the perspective of different tumours in the gastrointestinal tract. However, the current specific mechanism for m6A in cancer is unclear because m6A methylation has the function of a double-edged sword. High expression levels of m6A may lead to the development of certain tumours, but lack of m6A modification may lead to the progression of other tumours. (i) For the same tumour, different researchers presented inconsistent results on the expression level of m6A because of differences in research samples used. Ma et al. [32] believed that the expression level of m6A in liver cancer is reduced, but Yang et al. [33] reported inconsistent results. (ii) m6A expression in different tumours has different meanings. (iii) For the same tumour, the same molecule, conclusions may be quite different. For example, for the mechanism of METTL3, Li [37] and Deng [38] reported contradicting results. (iv): For the same tumour, the same molecule, different researchers state contrasting results on its downstream genes and present different mechanisms [37,38,39,40]. Previous differences have been noted, but most m6A-modified RNAs undergo, rapid journal from RNA processing to degradation and m6A controls cell differentiation and pluripotency, which are both associated with cancer progression.
Inflammation is a major cause of liver cancer, GC, CRC and pancreatic cancer. Lipopolysaccharide (LPS) is the main component of the cell wall of Gram-negative bacteria and one of the most effective stimulants of innate immunity. LPS can stimulate macrophages to secrete various inflammatory cytokines, including TNF-α, IL-6 and IL-1β, by activating NF-κB and MAPK pathways, further aggravating the immune response [65]. Yu et al. [66] found the increased expression of YTHDF2 in LPS-stimulated RAW 264.7 cells (RAW 264.7 cells are widely used as a suitable macrophage model for research on inflammation). In addition, inhibition of YTHDF2 enhances the mRNA expression of MAP2K4 and MAP4K4 and triggers the activation of p38, ERK and NF- κB signalling, thereby promoting the expression of TNF-α, IL-1 β, IL-6 and IL-12. These results suggest that YTHDF2 may play a regulatory role in LPS-induced macrophage inflammatory responses. This study provides a potential target for anti-inflammatory therapy and new insights into the mechanisms of inflammation.
At present, most studies on m6A methylation report its mechanism and role as a tumour marker. However, few studies have focused on drug therapy. In addition, Future prospects still need to be further explored. Firstly, m6A regulates the detailed and precise function of each component in the regulation of cells. Secondly, as a tumour marker, the specificity and sensitivity of m6A need further studies. Thirdly, many studies have shown that m6A regulatory factors and related pathways can be used as therapeutic targets; however, no specific response has been reported in clinical practice. Future research will demonstrate the feasibility of using m6A alone or in combination with other therapies to treat diseases, such as tumours. Fourthly, other components of m6A methylation and demethylation and effectors should be identified. For instance, the functions of few m6A eraser components (e.g. ALKBH1, ALKBH4, ALKBH6 and ALKBH7) remain unknown and need to be determined.
Conclusion
Given that m6A methylation plays an important role in a variety of digestive tract tumors, m6A modification can serve as a diagnostic/prognostic target. Due to various related factors, the results of many researchers are sometimes contradictory. This requires more multi-center and large-scale research to further explore, so as to lay a foundation for accurate treatment of human tumors.
Availability of data and materials
Not applicable.
Abbreviations
- ALKBH5:
-
A-ketoglutarate-dependent dioxygenase alkB homolog 5
- CRC:
-
Colorectal cancer
- DGCR8:
-
DiGeorge Syndrome Crisis Area Gene 8
- EMT:
-
Epithelial-Mesenchymal Transition
- FTO:
-
Obesity-associated protein
- HCC:
-
Hepatic cell carcinoma
- m6A:
-
N6-methyladenosine
- METTL14:
-
Methyltransferase-like 14
- METTL3:
-
Methyltransferase-like 3
- SOCS2:
-
Suppressor Of Cytokine Signalling 2
- WTAP:
-
Wilms tumor suppressor-1-associated protein
- YTHDC1:
-
YTH domain-containing 1
- YTHDC2:
-
YTH domain-containing 2
- YTHDF1:
-
YTH N6-methyladenosine RNA binding protein 1
- YTHDF2:
-
YTH N6-methyladenosine RNA binding protein 2
- YTHDF3:
-
YTH N6-methyladenosine RNA binding protein 3
References
Ng RK, Gurdon JB. Epigenetic inheritance of cell differentiation status. Cell Cycle. 2008;7(9):1173–7.
Wouters BJ, Delwel R. Epigenetics and approaches to targeted epigenetic therapy in acute myeloid leukemia. Blood. 2016;127(1):42–52.
Li Y, Chen K, Zhou Y, et al. A new strategy to target acute myeloid leukemia stem and progenitor cells using chidamide, a histone deacetylase inhibitor. Curr Cancer Drug Targets. 2015;15(6):493–503.
Xing J, Yi J, Cai X, et al. NSun2 promotes cell growth via elevating cyclin-dependent kinase 1 translation. Mol Cell Biol. 2015;35(23):4043–52.
Dominissini D, Nachtergaele S, Moshitch-Moshkovitz S, et al. The dynamic N(1)-methyladenosine methylome in eukaryotic messenger RNA. Nature. 2016;530(7591):441–6.
Safra M, Sas-Chen A, Nir R, et al. The m1A landscape on cytosolic and mitochondrial mRNA at single-base resolution. Nature. 2017;551(7679):251–5.
You C, Dai X, Wang Y. Position-dependent effects of regioisomeric methylated adenine and guanine ribonucleosides on translation. Nucleic Acids Res. 2017;45(15):9059–67.
Lin S, Liu Q, Lelyveld VS, et al. Mettl1/Wdr4-mediated m7G tRNA methylome is required for normal mRNA translation and embryonic stem cell self-renewal and differentiation. Mol Cell. 2018;71(2):244–55 e5.
Batista PJ. The RNA Modification N6-methyladenosine and its implications in human disease. Genomics Proteomics Bioinformatics. 2017;15(3):154–63.
Boccaletto P, Machnicka MA, Purta E, et al. MODOMICS: a database of RNA modification pathways. 2017 update. Nucleic Acids Res. 2018;46(D1):D303–7.
Roundtree IA, Evans ME, Pan T, et al. Dynamic RNA modifications in gene expression regulation. Cell. 2017;169(7):1187–200.
Adhikari S, Xiao W, Zhao YL, et al. m6A: signaling for mRNA splicing. RNA Biol. 2016;13(9):756–9.
Genenncher B, Durdevic Z, Hanna K, et al. Mutations in cytosine-5 tRNA methyltransferases impact mobile element expression and genome stability at specific DNA repeats. Cell Rep. 2018;22(7):1861–74.
Ke S, Alemu EA, Mertens C, et al. A majority of m6A residues are in the last exons, allowing the potential for 3’UTR regulation. Genes Dev. 2015;29(19):2037.
Ke S, Pandya-Jones A, Saito Y, et al. m6A mRNA modifications are deposited in nascent pre-mRNA and are not required for splicing but do specify cytoplasmic turnover. Genes Dev. 2017;31(10):990–1006.
Desrosiers R, Friderici K, Rottman F. Identification of methylated nucleosides in messenger RNA from Novikoff hepatoma cells. Proc Natl Acad Sci U S A. 1974;71(10):3971–5.
Dominissini D, Moshitch-Moshkovitz S, Schwartz S, et al. Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq. Nature. 2012;485(7397):201–6.
Meyer KD, Saletore Y, Zumbo P, et al. Comprehensive analysis of mRNA methylation reveals enrichment in 3’UTRs and near stop codons. Cell. 2012;149(7):1635–46.
Liu J, Yue Y, Han D, et al. A METTL3-METTL14 complex mediates mammalian nuclear RNA N6-adenosine methylation. Nat Chem Biol. 2014;10(2):93–5.
Ping XL, Sun BL, Wang L, et al. Mammalian WTAP is a regulatory subunit of the RNA N6-methyladenosine methyltransferase. Cell Res. 2014;24(2):177–89.
Schwartz S, Mumbach MR, Jovanovic M, et al. Perturbation of m6A writers reveals two distinct classes of mRNA methylation at internal and 5′ sites. Cell Rep. 2014;8(1):284–96.
Wang P, Doxtader KA, Nam Y. Structural basis for cooperative function of METTL3 and METTL14 methyltransferases. Mol Cell. 2016;63(2):306–17.
Barbieri I, Tzelepis K, Pandolfini L, et al. Promoter-bound METTL3 maintains myeloid leukaemia by m6A-dependent translation control. Nature. 2017;552(7683):126–31.
Weng H, Huang H, Wu H, et al. Mettl14 inhibits hematopoietic stem/progenitor differentiation and promotes leukemogenesis via mRNA m6A modification. Cell Stem Cell. 2018;22(2):191–205e199.
Pendleton KE, Chen B, Liu K, et al. The U6 snRNA m(6) a methyltransferase METTL16 regulates SAM synthetase intron retention. Cell. 2017;169(5):824–835e14.
Patil DP, Chen CK, Pickering BF, et al. M(6) A RNA methylation promotes XIST-mediated transcriptional repression. Nature. 2016;537(7620):369–73.
Jia G, Fu Y, Zhao X, et al. N6-methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO. Nat Chem Biol. 2011;7(12):885–7.
Zheng G, Dahl JA, Niu Y, et al. ALKBH5 is a mammalian RNA demethylase that impacts RNA metabolism and mouse fertility. Mol Cell. 2013;9(1):18–29.
Torres IO, Fujimori DG. Functional coupling between writers, erasers and readers of histone and DNA methylation. Curr Opin Struct Biol. 2015;35:68–75.
Wang X, Lu Z, Gomez A, et al. N6-methyladenosine-dependent regulation of messenger RNA stability. Nature. 2014;505:117–20.
Wang X, Zhao BS, Roundtree IA, et al. N(6)-methyladenosine modulates messenger RNA translation efficiency. Cell. 2015;161(6):1388–99.
Ma JZ, Yang F, Zhou CC, et al. METTL14 suppresses the metastatic potential of hepatocellular carcinoma by modulating N6-Methyladenosine-dependent primary MicroRNA processing. Hepatology. 2017;65(2):529–43.
Yang Z, Li J, Feng G, et al. MicroRNA-145 Modulates N6-methyladenosine Levels by Targeting the 3′-Untranslated mRNA Region of the N6-methyladenosine–binding YTH Domain Family 2. Protein J Biol Chem. 2017;292(9):3614–23.
Lin S, Liu J, Jiang W, et al. METTL3 promotes the proliferation and mobility of gastric cancer cells. Open Med (Wars). 2019;14:25–31.
Zhang C, Zhang M, Ge S, et al. Reduced m6A modification predicts malignant phenotypes and augmented Wnt/PI3K-Akt signaling in gastric cancer. Cancer Med. 2019;8(10):4766–81.
Liu T, Yang S, Sui J, et al. Dysregulated N6-methyladenosine methylation writer METTL3 contributes to the proliferation and migration of gastric cancer. J Cell Physiol. 2020;235(1):548–62.
Li T, Hu PS, Zuo Z, et al. METTL3 facilitates tumor progression via an m6A-IGF2BP2-dependent mechanism in colorectal carcinoma. Mol Cancer. 2019;18(1):112.
Deng R, Cheng Y, Ye S, et al. m6A methyltransferase METTL3 suppresses colorectal cancer proliferation and migration through p38/ERK pathways. Onco Targets Ther. 2019;12:4391–402.
Nishizawa Y, Konno M, Asai A, et al. Oncogene c-Myc promotes epitranscriptome m6A reader YTHDF1 expression in colorectal cancer. Oncotarget. 2017;9(7):7476–86.
Bai Y, Yang C, Wu R, et al. YTHDF1 regulates Tumorigenicity and Cancer stem cell-like activity in human colorectal carcinoma. Front Oncol. 2019;9:332.
Chen J, Sun Y, Xu X, et al. YTH domain family 2 orchestrates epithelial-mesenchymal transition/proliferation dichotomy in pancreatic cancer cells. Cell Cycle. 2017;16(23):2259–71.
Tang X, Liu S, Chen D, et al. The role of the fat mass and obesity-associated protein in the proliferation of pancreatic cancer cells. Oncol Lett. 2019;17(2):2473–8.
Du H, Zhao Y, He J, et al. YTHDF2 destabilizes m6A-containing RNA through direct recruitment of the CCR4-NOT deadenylase complex. Nat Commun. 2016;7:12626.
Chen M, Wei L, Law CT, et al. RNA N6-methyladenosine methyltransferase-like 3 promotes liver cancer progression through YTHDF2-dependent posttranscriptional silencing of SOCS2. Hepatology. 2018;67(6):2254–70.
Cui M, Sun J, Hou J, et al. The suppressor of cytokine signaling 2 (SOCS2) inhibits tumor metastasis in hepatocellular carcinoma. Tumour Biol. 2016;37(10):13521–31.
Cheng X, Li M, Rao X, et al. KIAA1429 regulates the migration and invasion of hepatocellular carcinoma by altering m6A modification of ID2 mRNA. Onco Targets Ther. 2019;12:3421–8.
Torre LA, Bray F, Siegel RL, et al. Global cancer statistics, 2012. CA Cancer J Clin. 2015;65(2):87–108.
Chen W, Zheng R, Baade PD, et al. Cancer statistics in China, 2015. CA Cancer J Clin. 2016;66(2):115–32.
Zhang J, Guo S, Piao HY, et al. ALKBH5 promotes invasion and metastasis of gastric cancer by decreasing methylation of the lncRNA NEAT1. J Physiol Biochem. 2019;75(3):379–389.
Bray F, Ferlay J, Soerjomataram I, et al. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424.
Siegel R. Desantis, Jemal a. colorectal cancer statistics, 2014. CA Cancer J Clin. 2014;64(2):104–17.
Justilien V, Walsh MP, Ali SA, et al.The PRKCI and SOX2 oncogenes are coamplified and cooperate to activate Hedgehog signaling in lung squamous cell carcinoma.Send toCancer Cell. 2014;25(2):139–151.
Ricci-Vitiani L, Lombardi DG, Pilozzi E, et al. Identification and expansion of human colon-cancer-initiating cells. Nature. 2007;445(7123):111–5.
Lowenfels AB, Maisonneuve P. Epidemiology and risk factors for pancreatic cancer. Best Pract Res Clin Gastroenterol. 2006;20(2):197–209.
Wei P, Tang H, Li D. Insights into pancreatic cancer etiology from pathway analysis of genome-wide association study data. PLoS One. 2012;7(10):e46887.
Zhou Y, Yin Z, Hou B, et al. Expression profiles and prognostic significance of RNA N6-methyladenosine-related genes in patients with hepatocellular carcinoma: evidence from independent datasets. Cancer Manag Res. 2019;11:3921–31.
Zhao X, Chen Y, Mao Q, et al. Overexpression of YTHDF1 is associated with poor prognosis in patients with hepatocellular carcinoma. Cancer Biomark. 2018;21(4):859–68.
Li Y, Zheng D, Wang F, Xu Y, et al. Expression of Demethylase genes, FTO and ALKBH1, is associated with prognosis of gastric Cancer. Dig Dis Sci. 2019;64(6):1503–13.
Xu D, Shao W, Jiang Y, et al. FTO expression is associated with the occurrence of gastric cancer and prognosis. Oncol Rep. 2017;38(4):2285–92.
Cho SH, Ha M, Cho YH, et al. ALKBH5 gene is a novel biomarker that predicts the prognosis of pancreatic cancer: a retrospective multicohort study. Ann Hepatobiliary Pancreat Surg. 2018;22(4):305–9.
Zhu L, Zhu Y, Han S, et al. Impaired autophagic degradation of lncRNA ARHGAP5-AS1 promotes chemoresistance in gastric cancer. Cell Death Dis. 2019;10(6):383.
Taketo K, Konno M, Asai A, et al. The epitranscriptome m6A writer METTL3 promotes chemo- and radioresistance in pancreatic cancer cells. Int J Oncol. 2018;52(2):621–9.
Han D, Liu J, Chen C, et al. Anti-tumour immunity controlled through mRNA m6A methylation and YTHDF1 in dendritic cells. Nature. 2019;566(7743):270–4.
Chen B, Ye F, Yu L, et al. Development of cell-active N6-methyladenosine RNA demethylase FTO inhibitor. J Am Chem Soc. 2012;134(43):17963e17971.
Zhao BB, Guo HJ, Liu Y, et al. A novel benzoxazole derivative, exhibits anti-inflammatory properties via inhibiting GSK3beta activity in LPS-induced RAW264.7 macrophages. J Cell Biochem. 2018;119(7):5382–90.
Yu R, Li Q, Feng Z. m6A Reader YTHDF2 Regulates LPS-Induced Inflammatory Response. Int J Mol Sci. 2019;20(6):E1323.
Acknowledgements
This study was supported by Jiangsu Provincial Key Research and Development Special Fund (BE2015666), Jiangsu Innovative team leading talent fund (CXTDC2016006), Jiangsu Natural Science Foundation (BK20171304), Jiangsu six high peak talent fund (WSW-205,) Jiangsu 333 talent fund (BRA2016140) and Suqian Science and Technology Support Project (S201522).
Funding
Jiangsu Provincial Key Research and Development Special Fund (BE2015666), Jiangsu Innovative team leading talent fund (CXTDC2016006), Jiangsu Natural Science Foundation (BK20171304), Jiangsu six high peak talent fund (WSW-205,) Jiangsu 333 talent fund (BRA2016140) and Suqian Science and Technology Support Project (S201522).
Author information
Authors and Affiliations
Contributions
YF provided direction and guidance throughout the preparation of this manuscript. B-bH wrote and edited the manuscript. X-yW, Z-jG, and HZ reviewed and made significant revisions to the manuscript. X-YG and CZ collected and prepared the related papers. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Hu, Bb., Wang, Xy., Gu, XY. et al. N6-methyladenosine (m6A) RNA modification in gastrointestinal tract cancers: roles, mechanisms, and applications. Mol Cancer 18, 178 (2019). https://doi.org/10.1186/s12943-019-1099-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12943-019-1099-7