
Abstract
Background
Circular RNA Itchy E3 ubiquitin protein ligase (Circ-ITCH) is significantly down-regulated in various kinds of tumors, however, the mechanisms of action and functions of circITCH gene in prostate cancer (PC) are still under investigation. The mail goal of this research was to study the functional role of Circ-ITCH gene in prostate cancer and to illuminate the function role of circ-ITCH gene in prostate cancer by targeting miR-17-5p/HOXB13.
Methods
RT-qPCR was applied to measure the expression level of circ-ITCH and miR-17-5p in PC cell lines and tissues. CCK-8, colony formation, Brdu incorporation labeling and flow cytometry assays were applied to detect the effects of circ-ITCH and miR-17-5p on proliferation and cell apoptosis. Target gene prediction and screening, luciferase reporter gene assays were utilized to assess downstream target genes of miR-17-5p and Circ-ITCH. The protein and expression of HOXB13 gene were measured by Western blotting and RT-qPCR.
Results
CircITCH was significantly reduced in PC cell lines and tissues. Low circITCH expression level was highly related with preoperative PSA, tumor stage and Gleason score. Overexpression of circITCH can inhibit the malignant phenotype of prostate cancer. There was a high negative relationship between the expression level of microRNA-17-5p and circITCH in PC tissues, however, there existed a positive relationship between the expression of HOXB13 and circITCH. CircITCH acted as a sponge of miR-17-5p to increase HOXB13 gene expression. In addition, miR-17-5p overexpression or HOXB13 silencing can reduce the carcinogenic effects of circICCH in prostate cancer.
Conclusion
CircITCH promoted prostate cancer progression by regulating the HOXB13/miR-17-5p axis, and circITCH have a potential usage as therapeutic target for PC tumors.
Similar content being viewed by others

Background
PC (short for prostate cancer) is a popular malignant tumor in men [1, 2]. Its mortality rate ranks second in cancer in Europe and the United States. Its incidence is also the second among all malignant tumors in men worldwide [1, 3]. For the past few years, the occurrence rate of prostate cancer in males has increased year by year, which is closely related to the continuous growth of life expectancy, the aging of the population, the change of diet structure and the continuous improvement of diagnostic techniques [4,5,6]. Prostate cancer poses a heavily threat to the health and life of men, particularly in the middle and late stages. After endocrine therapy, most of the patients eventually progress to CRPC (castration-resistant prostate cancer), which is insensitive to radiotherapy and chemotherapy. So far, there is no effective treatment, which is a worldwide problem. At present, there are many methods for treating prostate cancer, including surgical treatment, hormone therapy, and radiation therapy [7,8,9]. Although these treatment methods are widely used in clinical practice, there are many limitations in terms of efficacy or prognosis. Therefore, the current research hotspot is to find a biomarker with sensitivity and specificity for early diagnosis and cure of prostate cancer.
The development and progression of prostate cancer is complicated, including changes in molecular genetics and epigenetics. Research on the pathogenesis of prostate cancer has never stopped, from proteomics to genomics, from DNA to RNA, from encoding RNA to non-coding RNA [10, 11]. Although researchers have put a lot of effort into the study of prostate cancer, the mechanism of action for the pathogenesis of prostate cancer has not been unveiled. With the advancement of science and technology, non-coding RNAs that cannot encode proteins after transcription gradually enter the field of view.
Recent studies have shown that circRNAs are a kind of single-stranded closed circular RNA, which are characterized by stable structure, tissue-specific and conservative evolution [12, 13]. Recent studies have gradually revealed the function of circRNAs, for example, circCDR1as has about 70 miR-7 binding sites, which can regulate EGFR expression by adsorbing miR-7 [14]. Circ_001569 can promote the invasion and proliferation of colorectal tumor cells by adsorbing mi R-145 to regulate the expression level of target E2F5 gene [15]. In general, in recent years, it has been found that circRNAs mainly function as competitive endogenous RNA (ce RNA) or MI RNA sponge. In addition, CIRC RNAs also play a role in regulating selective splicing and gene transcription, regulating parental gene expression and transcription translation [16, 17]. Cir-ITCH is located on human chromosome 20, 20q11.22. It is homologous to the RNA sequence of ITCH, a protein-coding gene. It usually spans 1–5 exons [18]. Cir-ITCH was first found to be low-expressed in esophageal cancer, then in colon cancer, hepatocellular carcinoma and lung cancer [19, 20]. At present, the mechanism of action of cir-ITCH in the progression and development of prostate cancer is still under investigation. To solve these problems, this study was set to study the effect of cir-ITCH gene on the apoptosis and proliferation of prostate cancer by detecting the expression of cir-ITCH in PC cell lines and tissues, and further interpret the functional role of cir-ITCH in regulating the apoptosis and proliferation of prostate cancer cells. To explain the pathogenesis of PC maybe offer an effective and precise therapeutic target.
Methods
Patient organization
The study was authorized by the Research Ethics Committee of The Jintan Hospital Affiliated with Jiangsu University. Informed consent was gained from all the patients. PC tissues and matched nearby normal tissues were obtained from 52 patients who received surgical resection of PC between Feb. 2014 to Apr. 2018. After excision of the prostate tissue, all tissue samples were stored in liquid nitrogen. Histopathological results validated the diagnosis of prostate cancer.
Cell culture and cell transfection
Human PC cell lines (C4-2, LNCaP, DU145, 22Rv1, PC-3 and VcaP) and human prostate epithelial cell line (RWPE-1) were obtained from the Chinese Academy of Sciences Cell Bank (Shanghai, China). Human prostate cancer cell lines were maintained in RPMI-1640 medium. Human prostate epithelial cell line (RWPE-1) was maintained in Eagle medium with 10% FBS (fetal bovine serum) (Invitrogen).
The fragment of 873 bp cDNA was inserted into a PLCDH-cir plasmid vector (Ribobio, China) to construct a circ-ITCH overexpression vector. A lentivirus was packaged by Hanbio Company (Shanghai, China). Transfection procedures were performed as described previously. The si-HOXB13, miR-17-5p mimetic and the responding vector controls (GenePharma, China) were transfected into PC cells with Lipofectamine 2000 reagents (Invitrogen, USA). The sequence of si-HOXB13 was as follows: 5′-TAAGAAAGCATGTGTGAACTA-3′.
Quantitative reverse transcription PCR (qRT-PCR)
qRT-PCR measure was applied with SYBR Premix Ex Taq reagents (TaKaRa, China). The expression levels of each gene was detected and normalized to the U6 snRNA or GAPDH using the Ct method of 2−ΔΔ. Total RNA was enriched from PC tissues or cells lines with TRIzol reagent. cDNA (Complementary DNA) was generated according to total RNA with SuperScript III. The gene expression of circRNA/mRNA and miRNA was determined by LightCyclerTM 480 system. The internal controls of miRNA and mRNA/circRNA were U6 and GAPDH, respectively. The oligonucleotides are as shown in Table 1.
Animal research
A total of 24 Female BALB/c nude mice, 6 weeks old, were randomly grouped into 4 groups, 6 mice in each group. Prostate cells were with a transfection of control vector and circ-ITCH. Prostate cancer cells with a density of 1 × 107 tumor cells per mice were subcutaneously injected into the lateral flank of female athymic BALB/C nude mice. The size of tumor was detected twice every a week using a caliper and the formula V = (width2 × length)/2 was used. One month after the subcutaneous injection, the nude mice were sacrificed and the last total weight of tumor was detected. Animal studies were applied in strictly comply with the institutional ethics guidelines for animal experiments approved by the The Jintan Hospital Affiliated with Jiangsu University Animal Management Committee.
IHC staining: Frozen sections from mouse xenografts tumor were stained with Ki67 primary antibodies. Protein level was detected by an I-View Detection chemistry system and a NexES automated stainer (Ventana Medical Systems, AZ).
CCK-8 method
Transfected cells were placed into wells of 96-well plates at a cell density of 1 × 103 cells/well. Three wells were replicated in each group and 10 μL/well of CCK8 was added to each well of cells for the last 2 h of incubation. Finally, the OD value of the cell liquid was measured by an enzyme-linked immunosorbent assay.
Colony formation
The transfected cells (1 × 103 cells per well) were seeded into a 6-well plate to culture for 24 h. After the cells were exposed to ADR for 24 h, each well was washed, the culture medium was exchanged, and the cells were maintained for 8 days. Finally, colonies were counted after fixation for 10 min with 10% formaldehyde, and incubated with crystal violet (0.1%) for 10 min. Cell viability was analyzed using GraphPad Prism 6 (GraphPad Sofware, USA).
BrdU incorporation labeling
The transfected cells (1 × 103 cells per well) were seeded in a 6-well plate to adhere within 24 h. After transfection for 48 h, the cell proliferation assay was conducted with the BrdU cell growth assay kit.
Apoptosis detection
The transfected cells were adhered to the wall and washed two times with cooled PBS, then mixed up in 100 μL volume of working buffer. The assay kit of Annexin V-FITC-PI (Roche Applied Science, Germany) was utilized for cell staining following the vendor guidance. Apoptosis results were run by a FACS cytometer (Can II, BD Biosciences, USA).
Luciferase assay
The pmirGLO miRNA target expression vector containing dual luciferase was used in the 3′-untranslated region (UTR) luciferase reporter gene assay (Promega, USA). Cells were placed into a 24-well plate at a cell density of 0.4 million cells per mL, after that cells were cotransfected by Lipofectamine 2000 (Invitrogen) with HOXB13 and circITCH mimics and wild type or mutant sequences. Cells were collected 48 h after transfection, after that, a dual luciferase reporter assay system from premega was used to assess firefly or Renilla luciferase activities and calculated as ratio of firefly to Renilla luciferase activity.
Western blot
After the total protein was cleaved using a lysis buffer containing 1% PMSF, the concentration of protein was measured with a BCA Protein Detection Kit (Sigma, USA). The lysate was applied to a 12% polyacrylamide gel for electrophoresis experiments, and after electrophoresis, it was transferred to a PVDF membrane. Membranes were blocked using 5% milk and incubated with anti-HOXB13 antibody (Proteintech, Chicago, IL, USA) over a night at 4–8 °C. After that, it was incubated with anti-rabbit IgG antibody (diluted in 1/5000 n; GE, Little) for 2–3 h at 37 °C temperature. Commerical GAPDH antibody reagent (diluted in 1/2500; Bioworld) was applied as an internalcontrol standard. The kit of ECL Western blotting (Amersham, UK) was used to measure all protein bands.
Statistical method
The monitoring data were analyzed by SPSS19.0 statistical software. The data analysis data was showed as mean ± SD (mean ± standard deviation). The data analysis between the 2 groups was applied by t test. The data analysis between 2 more groups was applied with ANOVA method (one-way variance analysis), and the LSD test was conducted for the next analysis. Kaplan–Meier plot was utilized to analyze the survival curves. P < 0.05 means that there was statistically significant difference.
Results
CircITCH was significantly over expressed and indicated poor prognosis in prostate cancer
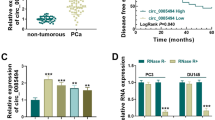
qRT-PCR method was applied to measure the expression of circITCH gene in PC cell lines and tissues to study the potential biological role of circITCH gene in the development of prostate cancer. The results were shown in Fig. 1a, circITCH expression levels were significantly reduced in prostate cancer tissues samples with a comparison to mated nearby normal tissues (P < 0.05, n = 52). CircITCH expression levels were significantly reduced in PC cell lines with a comparison to human prostate epithelial cell line RWPE-1 (Fig. 1b). As shown in Fig. 1c, in the PC cell lines, the expression level of circITCH gene was significantly increased compared with a comparison to the control group (P < 0.05). Because the cell line C4-2 is a subline of LNCaP, LNCaP and DU145 were selected for further studies. In addition, it was significantly associated with preoperative PSA, Gleason score, and tumor stage in the circITCH low expression group compared with that in the circITCH high expression group (Table 2). More importantly, low circITHC expression was significantly related to poor overall survival (P < 0.01; Fig. 1d). The data indicated that circITCH had a potential carcinogenic effect in prostate cancer.

Expression level of circITCH gene in prostate cancer. a Expression of circITCH in tissues of PC tumor and matched nearby normal tissues (sample number = 52). b Expression level of circITCH gene in PC tumor cell lines (C4-2, LNCaP, DU145, PC-3, 22Rv1 and VcaP) and human prostate epithelial cell line (RWPE-1). c Interference effector expression of circITCH targeting vectors of circITCH and ITCH in LNCaP and DU145cell lines. d Kaplan–Meier plot of PC patients with low (n = 26) or high (n = 26) circITCH expression. *P < 0.05, **P < 0.01
Overexpression of circITCH inhibited prostate cancer growth in vivo
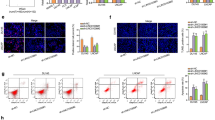
Further analysis in animal experiments was conducted to investigate the carcinogenic effects of circITCH in prostate cancer. Results were shown in Fig. 2a–c, circITCH overexpression inhibited tumor growth and decreased tumor volume. The IHC results of Ki-67 showed that the positive rate of Ki-67 expression in the circITCH overexpression group was significantly lower than control group (Fig. 2d, p < 0.05). These data indicated that overexpression of circITCH can inhibit the development of prostate cancer.

Overexpression of circITCH inhibited PC tumor growth in nude mice model. a Representative images of tumors after subcutaneous injection of different prostate cancer cells. b Tumor size in nude mice. c Tumor weight in nude mice. d IHC staining results (Ki-67). Scale bar = 20 μm. ***P < 0.001, **P < 0.01, *P < 0.05
Overexpression of circITCH suppressed the proliferation and induced the apoptosis in prostate cancer cells
Next the functional role of circITCH gene in prostate cancer by cell assay was tested. The CCK-8 assay showed that the cell livability of the overexpressed circITCH group was significantly decreased in LNCaP and DU145 cells with a comparison to that in control group (P < 0.05, Fig. 3a). Colony formation assay results showed that the circITCH overexpression group showed a significant decrease in cell colony forming ability in DU145 and LNCaP cells (P < 0.05, Fig. 3b). The Brdu incorporation labeling results showed that the cell proliferation of the circITCH overexpression group was significantly decreased in DU145 and LNCaP cells (P < 0.05, Fig. 3c). Flow cytometry results showed that the apoptosis rate of circITCH overexpression in DU145 and LNCaP cells was significantly increased (Fig. 3d). These data indicated that overexpression of circITCH can suppress the malignant phenotype of PC cells.

Overexpression of circITCH inhibited the malignant phenotype of PC cells. a Cell livability assay. b Cell growth assay. c Determination of cell colony forming ability. d Determination of apoptotic rate. ***P < 0.001, **P < 0.01, *P < 0.05
CircITCH served as a sponge of miR-17-5p
By searching the network tool Starbase, it was predicted that circITCH might have a putative binding site for miR-17-5p (Fig. 4a). MT and WT miR-17-5p-3′-UTR was cloned and luciferase activity was analyzed to confirm whether circITCH can directly bind to miR-17-5p. As shown in Fig. 4b, circITCH overexpression significantly reduced luciferase activity of WT miR-17-5p 3′ UTR (P < 0.01), but luciferase activity against MT miR-17-5p-3′UTR had no significant changes. In addition, the expression yield of miR-17-5p gene was significantly reduced in the circITCH group with a comparison to control group (Fig. 4c, P < 0.05). Furthermore, miR-17-5p expression yield were significantly up-regulated in PC tissues with a comparison to matched normal tissues (P < 0.05) (Fig. 4d). There was also a clearly negative relationship between the expression level of miR-17-5p and circITCH in PC tissues (P < 0.001, r = − 0.533, Fig. 4e). These data showed that circITCH may get its functional role through spongiform miR-17-5p.

CircITCH can be used as a sponge for miR-17-5p. a Putative targeting sites for miR-17-5p and circITCH, b assessment of luciferase activity. c Expression yield of miR-17-5p in cells overexpressing circITCH. d Expression levels of miR-17-5p gene in PC tissues and nearby normal tissues (tissues number = 52). e Pearson correlation analysis of miR-17-5p and circITCH in PC tissues (tissues number = 52) (P < 0.001, r = − 0.533). **P < 0.01, ***P < 0.001
CircITCH sponges and sequesters miR-17-5p to upregulate HOXB13 expression
By searching the network tool Starbase, HOXB13 was calculated to be a potential target protein for miR-17-5p (Fig. 5a). To check whether miR-17-5p can bind directly to HOXB13, WT or MT HOXB13-3′-UTR was cloned and luciferase activity was analyzed. As indicated in Fig. 5b, miR-17-5p over expression significantly reduced luciferase activity of WT HOXB13-3′UTR (P < 0.01), but there was no significant change in luciferase activity of MT HOXB13-3′UTR. In addition, compared with that in the control group, the protein and mRNA expression yields of HOXB13 gene in the miR-17-5p overexpression group were significantly decreased, and the protein and mRNA expression yeilds of HOXB13 in the circITCH overexpression group were significantly increased (P < 0.05), while miR-17-5p-overexpression was able to reverse the increase in the protein and mRNA expression yields of HOXB13 induced by circITCH overexpression (Fig. 5c, d). Furthermore, it was found that circITCH expression was highly related to HOXB13 expression in PC tissues (P < 0.001, r = 0.508) (Fig. 5e). These data showed that circITCH promoted the expression of the tumor suppressor HOXB13 by acting as a sponge of miR-17-5p in prostate cancer.

CircITCH upregulated HOXB13 expression by sponge miR-17-5p. a Putative binding regions for HOXB13 3′-UTR and miR-17-5p. b Measurement of luciferase activity. c Analysis of HOXB13 mRNA expression levels. d Analysis of HOXB13 protein expression levels. e Correlation analysis of HOXB13 and circITCH in prostate cancer tissues (sample numbers = 52) (r = − 0.508, P < 0.001. *Control group, #vs miR-17-5p simulation group. **P < 0.01, ##P < 0.01
MiR-17-5p overexpression or HOXB13 silencing effectively reversed circITCH-induced inhibition of prostate cancer progression
In order to further confirm that circITCH exerted a biological role in prostate cancer by regulating the HOXB13/miR-17-5p axis, the circITCH expression vector and the miR-17-5p mimic vector were co-transfected into PC cells. The results of CCK-8 assay showed that compared with that in the control group, the viability of circITCH overexpression group was significantly decreased in LNCaP and DU145 cells, and miR-17-5p overexpression and HOXB13 silencing could significantly reverse the effect of the ircITCH overexpression on viability (P < 0.05) (Fig. 6a). The results of Brdu incorporation assay showed that the cell proliferation of LNCaP and DU145 cells was significantly decreased in circITCH overexpression group, and miR-17-5p overexpression and HOXB13 silencing could significantly reverse the effect of ircITCH overexpression on cell proliferation (P < 0.05) (Fig. 6b). The results of colony formation assay showed that the colony forming ability of circITCH overexpression group was significantly decreased in LNCaP and DU145 cells, and miR-17-5p overexpression and HOXB13 silencing could significantly reverse the effect of ircITCH overexpression on cell colony formation ability (P < 0.05) (Fig. 6c). The data of flow cytometry indicated that the apoptosis rate of circITCH overexpression in DU145 and LNCaP cells was significantly increased, and miR-17-5p overexpression and HOXB13 silencing could significantly reverse the effect of circITCH overexpression on apoptosis ability (P < 0.05) (Fig. 6d). In a conclusion, these data demonstrated that HOXB13 silencing or miR-17-5p overexpression reversed the invasive phenotype of circITCH-induced prostate cancer cells.

HOXB13 silencing or miR-17-5p overexpression effectively reversed the promotion of circITCH-induced prostate cancer progression. a CCK-8 assay of DU145 and LNCaP cells co-transfected with control or circITCH vector and si-HOXB13 or miR-17-5p mimic. b BrdU incorporation assay of DU145 and LNCaP cells co-transfected with control or circITCH vector and si-HOXB13 or miR-17-5p mimic. c Colony formation assay of DU145 and LNCaP cells co-transfected with control or circITCH vector and si-HOXB13 or miR-17-5p mimic. d Flow cytometry assay of DU145 and LNCaP cells co-transfected with control or circITCH vector and si-HOXB13 or miR-17-5p mimic. #vs circITCH group, *vs control vector group, ##P < 0.01, **P < 0.01
Discussion
Cancer of prostate is a usual tumor of the male genitourinary system, and its incidence rate is not only the highest in Europe and the United States, but also shows a significant upward trend [21, 22]. Early hormone-dependent prostate cancer can achieve the purpose of reducing tumors and lowering blood PSA through endocrine therapy, but gradually evolves into hormone-independent prostate cancer after treatment, leading to the death of patients eventually from hormone-insensitive cells [23, 24]. The molecular mechanism of the progression and development of hormone-independent prostate tumor has not comprehension. Therefore, it is very necessary to study the molecular mechanism of hormone-independent prostate tumorigenesis and provide a gist for clinical protection and treatment of hormone-independent prostate tumor.
Non-coding RNA (ncRNA) is a biomolecule that is ubiquitous in organisms and plays an important role in regulating life activities [25, 26]. Circular RNA (circRNA) is a type of endogenous ncRNA molecule that is widely and diversely present in a variety of biological cells and regulates gene expression [27, 28]. CircRNA has a key function role in gene regulation, such as miRNA adsorption as miRNA sponge, regulation of related gene expression, expression of peptides, and so on [29, 30]. As a new regulatory endogenous RNA, circRNA has attracted more and more attention in the development of tumors and has become a new research hotspot [31, 32]. It has been reported that many circRNAs are abnormally expressed in different tumors types such as gastric cancer, colon cancer and esophageal cancer, and exert an significant role in tumor growth, invasion and migration [33, 34]. Studies have found that the expression levels of circ MYLK in PCA samples and PCA cells are significantly higher than that in the normal tissues and normal prostate cells. Overexpression of circRNA MYLK promotes proliferation, invasion and migration of PCA cells, while is circRNA MYLK accelerates apoptosis of PCA cells. In addition, the study has also found that si circRNA MYLK affects the function of PCA cells by targeting mir-29a [35]. In esophageal, colon and lung cancer, cir-ITCH binds to MI R-7/mi R-17/mi R-214 through molecular sponge to form cir-ITCH-mi RNA-7/mi R-17/mi R-214-ITCH regulatory signal axis, up-regulates the expression of ITCH, promotes the ubiquitination degradation of DVL2, inhibits the classical Wnt/β-catenin signaling pathway, and thus inhibiting the growth of cancer cells [19, 20]. Other articles have showed that the expression of cir-ITCH in bladder cancer is also down-regulated. Overexpression of cir-ITCH can suppress the growth, invasion and migration of bladder cancer cells, and further confirm that cir-ITCH targets miR-17 and miR-224, up-regulates the target genes p21 and PTEN of mi R-17 and mi R-224, thus suppressing the growth, invasion, migration and metastasis of cancer cells [36]. In this study, it was found that the expression of cir-ITCH was reduced in breast cancer cell lines and tissues, which was consistent with the above reported results, and confirmed that its expression level was significantly correlated with preoperative PSA, Gleason score and tumor stage. Furthermore, in vivo experiments showed that circITCH overexpression inhibited tumor growth rate and tumor volume. In vitro experiments showed that up-regulation of circITCH significantly suppressed the growth, invasion and migration of prostate cancer cells and induced apoptosis. These data suggested that overexpression of circITCH can effectively delay the progression of prostate cancer.
The study found that circRNA contains a large number of MERs that act as competitive endogenous RNA (ceRNA) and become a new member of competitive endogenous RNA [37]. How to form a regulatory network between circRNAs and miRNAs to maintain balance and regulate life activities of the organism needs further research and exploration. Some study found that using a TaqMan-based reverse transcriptase polymerase chain reaction assay, the relationship between cir-ITCH and ESCC was analyzed in a total of 684 ESCC and paired adjacent non-tumor tissue samples from eastern and southern China. We found that cir-ITCH expression was usually low in ESCC compared to the peritumoral tissue. The functional relevance of cir-ITCH was further examined by biochemical assays. As sponge of miR-7, miR-17, and miR-214, cir-ITCH might increase the level of ITCH [20]. In recent years, the relationship between miRNAs and tumors has become a research hotspot. In general, the decline or loss of prostate cancer gene expression is considered to be a tumor inhibitor gene. The expression of microRNA17 is generally increased in cancer cells, and it is highly expressed in all detected solid tumors. It is currently believed that miRNA-17-5p is widely involved in tumor proliferation, migration, apoptosis and growth [38]. Studies have found that patients with higher miR-17-5p expression level have lower survival rates overall and there is a highly negative relation between p130 and mir-17-5p expression [39]. This study suggested that circITCH can bind to target miR-17-5p, and the expression level of miR-17-5p was significantly elevated in PC tissues. And a high negative relationship was found between the expression of miR-17-5p and circITCH in PC tissues. This indicated that circITCH may exert its biological function through spongy miR-17-5p, which was consistent with the literature reports.
CircITCH can act on multiple target genes like p63, p73, Notch1, Wnt/pathway [19, 36]. A study has shown that ITCH degrades the phosphorylated form of disheveled (Dvl) via the proteasome pathway, thus inhibiting the action of the canonical Wnt pathway [20]. The expression of HOXB13 is mostly in the posterior structure of the embryo and plays an important role in the development of the spinal cord, digestive tract and genitourinary system [40]. HOXB13 is a recently discovered gene closely related to the growth and development of normal prostate tissue and the occurrence and progression of prostate cancer. HOXB13 also plays a variety of functions during the progression of prostate cancer [41, 42]. On the one hand, it is believed that the high expression of HOXB13 can inhibit the activation of AR, regulate the cell cycle and inhibit the growth of prostate cancer, on the other hand, HOXB13 is thought to promote the invasion and metastasis of prostate cancer [43]. This study indicated that miR-17-5p-overexpression reversed the increase in protein and mRNA expression yeilds of HOXB13 induced by circITCH overexpression. Furthermore, circITCH expression was positively correlated with HOXB13 expression in prostate cancer tissues. It was shown that circITCH enhanced the expression of the tumor suppressor HOXB13 by acting as a sponge of miR-17-5p in prostate cancer.
Conclusion
CircITCH promoted prostate cancer growth and invasion and metastasis by regulating miR-17-5p/HOXB13 axis, suggesting that circITCH may be a potential therapeutic target for PC. It provided experimental basis for the study of clinical prognosis of the tumor and further targeted intervention therapy.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Change history
30 April 2024
This article has been retracted. Please see the Retraction Notice for more detail: https://doi.org/10.1186/s12935-024-03344-y
Abbreviations
- PC:
-
prostate cancer
- CRPC:
-
castration-resistant prostate cancer
- ceRNA:
-
endogenous RNA
- UTR:
-
3′-untranslated region
- circRNA:
-
circular RNA
References
Scher HI, Fizazi K, Saad F, Taplin ME, Sternberg CN, Miller K, et al. Increased survival with enzalutamide in prostate cancer after chemotherapy. N Engl J Med. 2012;367(13):1187.
Vordermark D. Quality of life and satisfaction with outcome among prostate-cancer survivors. N Engl J Med. 2008;26(5):565–6.
Montgomery RB, Mostaghel EA, Vessella R, Hess DL, Kalhorn TF, Higano CS, et al. Maintenance of intratumoral androgens in metastatic prostate cancer: a mechanism for castration-resistant tumor growth. Can Res. 2008;68(11):4447–54.
Viktorov AA, Zharinov GM, Neklasova NJ, Morozova EE. Prediction of life expectancy for prostate cancer patients based on the kinetic theory of aging of living systems. Adv Gerontol. 2017;7(4):290–5.
Kalra S, Basourakos S, Abouassi A, Achim M, Volk RJ, Hoffman KE, et al. The implications of ageing and life expectancy in prostate cancer treatment. Nat Rev Urol. 2016;13(5):289–95.
Daskivich TJ, Lai J, Dick AW, Setodji CM, Hanley JM, Litwin MS, et al. Variation in treatment associated with life expectancy in a population-based cohort of men with early-stage prostate cancer. Cancer. 2015;120(23):3642–50.
Walz J, Graefen M, Huland H. Basic principles of anatomy for optimal surgical treatment of prostate cancer. World J Urol. 2007;25(1):31–8.
Akaza H, Chang SJ, Chen KK, Esuvaranathan K, Fujioka T, Hirao Y, et al. The 2nd conference on Asian trends in prostate cancer hormone therapy. Gan to Kagaku Ryoho Cancer Chemother. 2004;31(8):1285–95.
Pisansky TM, Hunt D, Gomella LG, Amin MB, Balogh AG, Chinn DM, et al. Duration of androgen suppression before radiotherapy for localized prostate cancer: radiation therapy oncology group randomized clinical trial 9910. J Urol. 2016;195(1):96.
Mateo J, Carreira S, Sandhu S, Miranda S, Mossop H, Perezlopez R, et al. DNA-repair defects and olaparib in metastatic prostate cancer. N Engl J Med. 2016;195(4):925–8.
Chung S, Nakagawa H, Uemura M, Piao L, Ashikawa K, Hosono N, et al. Association of a novel long non-coding RNA in 8q24 with prostate cancer susceptibility. Cancer Sci. 2011;102(1):245–52.
Chen LL, Yang L. Regulation of circRNA biogenesis. RNA Biol. 2015;12(4):381–8.
Glažar P, Papavasileiou P, Rajewsky N. circBase: a database for circular RNAs. Rna-a Publ Rna Soc. 2014;20(11):1666–70.
Zhang X, Yang D, Wei Y. Overexpressed CDR1as functions as an oncogene to promote the tumor progression via miR-7 in non-small-cell lung cancer. Oncotargets Ther. 2018;11:3979–87.
Xie H, Ren X, Xin S, Lan X, Lu G, Yuan L, et al. Emerging roles of circRNA_001569 targeting miR-145 in the proliferation and invasion of colorectal cancer. Oncotarget. 2016;7(18):26680–91.
Jeck WR, Sorrentino JA, Wang K, Slevin MK, Burd CE, Liu J, et al. Circular RNAs are abundant, conserved, and associated with ALU repeats. Rna-a Publ Rna Soc. 2013;19(2):141–57.
Zhang S, Zhu D, Li H, Li H, Feng C, Zhang W. Characterization of circRNA-associated-ceRNA networks in a senescence-accelerated mouse prone 8 brain. Mol Ther. 2017;25(9):2053.
Huang G, Hua Z, Shi Y, Wu W, Cai H, Chen X. cir-ITCHPlays an inhibitory role in colorectal cancer by regulating the Wnt/β-catenin pathway. PLoS ONE. 2015;10(6):e0131225.
Wan L, Zhang L, Fan K, Cheng ZX, Sun QC, Wang JJ. Circular RNA-ITCH suppresses lung cancer proliferation via inhibiting the Wnt/Î2-CATENIN PATHWAY. Biomed Res Int. 2016;2016(1):1579490.
Li F, Zhang L, Li W, Deng J, Zheng J, An M, et al. Circular RNA ITCH has inhibitory effect on ESCC by suppressing the Wnt/β-catenin pathway. Oncotarget. 2015;6(8):6001–13.
Keating NL, O’Malley AJ, Freedland SJ, Smith MR. Diabetes and cardiovascular disease during androgen deprivation therapy: observational study of veterans with prostate cancer. J Natl Cancer Inst. 2012;104(19):1518–23.
Hieronymus H. Integrative genomic profiling of human prostate cancer. Cancer Cell. 2010;18(1):11–22.
Shaw G, Oliver RT. Intermittent hormone therapy and its place in the contemporary endocrine treatment of prostate cancer. Surg Oncol. 2009;18(3):275–82.
Rick FG, Block NL, Schally VA. An update on the use of degarelix in the treatment of advanced hormone-dependent prostate cancer. Oncol Targets Ther. 2013;6:391–402.
Mattick JS, Makunin IV. Non-coding RNA. Hum Mol Genet. 2006;15 Spec No 1(suppl_1):R17.
Esteller M. Non-coding RNAs in human disease. Nat Rev Genet. 2011;12(12):861–74.
Memczak S, Jens M, Elefsinioti A, Torti F, Krueger J, Rybak A, et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature. 2013;495(7441):333–8.
Salzman J, Gawad C, Wang PL, Lacayo N, Brown PO. Circular RNAs are the predominant transcript isoform from hundreds of human genes in diverse cell types. PLoS ONE. 2012;7(2):e30733.
Kulcheski FR, Christoff AP, Margis R. Circular RNAs are miRNA sponges and can be used as a new class of biomarker. J Biotechnol. 2016;238:42–51.
Toit AD. RNA: circular RNAs as miRNA sponges. Nat Rev Mol Cell Biol. 2013;14(4):195.
Yang W, Du WW, Li X, Yee AJ, Yang BB. Foxo3 activity promoted by non-coding effects of circular RNA and Foxo3 pseudogene in the inhibition of tumor growth and angiogenesis. Oncogene. 2015;35(30):3919.
Du WW, Fang L, Yang W, Wu N, Awan FM, Yang Z, et al. Induction of tumor apoptosis through a circular RNA enhancing Foxo3 activity. Cell Death Differ. 2016;24(2):357.
Pan H, Li T, Jiang Y, Pan C, Ding Y, Huang Z, et al. Overexpression of circular RNA ciRS-7 abrogates the tumor suppressive effect of miR-7 on gastric cancer via PTEN/PI3 K/AKT signaling pathway. J Cellular Biochem. 2018;119(1):440–6.
Zhong Y, Du Y, Xue Y, Mo Y, Fan C, Fang X, et al. Circular RNAs function as ceRNAs to regulate and control human cancer progression. Mol Cancer. 2018;17(1):79.
Dai Y, Li D, Chen X, Tan X, Gu J, Chen M, et al. Circular RNA myosin light chain kinase (MYLK) promotes prostate cancer progression through modulating Mir-29a expression. Med Sci Monit Int Med J Exp Clin Res. 2018;24:3462–71.
Yang C, Yuan W, Yang X, Li P, Wang J, Han J, et al. Circular RNA circ-ITCH inhibits bladder cancer progression by sponging miR-17/miR-224 and regulating p21, PTEN expression. Mol Cancer. 2018;17(1):19.
Xing Z, Min Z, Rong Y, Zhao W, Hu X, Gan J. Identification and comparison of novel circular RNAs with associated co-expression and competing endogenous RNA networks in pulmonary tuberculosis. Oncotarget. 2017;8(69):113571–82.
Fan M, Sethuraman A, Brown M, Sun W, Pfeffer LM. Systematic analysis of metastasis-associated genes identifies miR- 17-5p as a metastatic suppressor of basal-like breast cancer. Breast Cancer Res Treat. 2014;146(3):487–502.
Hossain A, Kuo MT, Saunders GF. Mir-17-5p regulates breast cancer cell proliferation by inhibiting translation of AIB1 mRNA. Mol Cell Biol. 2006;26(21):8191–201.
Ewing CM, Ray AM, Lange EM, Zuhlke KA, Robbins CM, Tembe WD, et al. Germline mutations in HOXB13 and prostate-cancer risk. N Engl J Med. 2012;366(2):141–9.
Walsh PC. Re: germline mutations in HOXB13 and prostate cancer risk. J Urol. 2012;187(4):1266–7.
Edwards S, Campbell C, Flohr P, Shipley J, Giddings I, Tepoele R, et al. Expression analysis onto microarrays of randomly selected cDNA clones highlights HOXB13 as a marker of human prostate cancer. BRJ Cancer. 2005;92(2):376–81.
Jung C, Kim RS, Lee SJ, Wang C, Jeng MH. HOXB13 homeodomain protein suppresses the growth of prostate cancer cells by the negative regulation of T-cell factor 4. Can Res. 2004;64(9):3046–51.
Acknowledgements
Not applicable.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
XGW: study concepts, study design, literature research, experimental studies, manuscript preparation and editing; RW: definition of intellectual content, literature research experimental studies, manuscript preparation and editing; ZW: literature research, experiments work and manuscript writing; PDB: data acquisition, statistical analysis. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Ethical approval was obtained from the Ethics Committee of The Jintan Hospital Affiliated with Jiangsu University. All procedures performed in studies involving human participants or animals were in accordance with the ethical standards of the institutional and national research committee. Written informed consent was obtained from all individual patients included in the study.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article has been retracted. Please see the retraction notice for more detail: https://doi.org/10.1186/s12935-024-03344-y
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Wang, X., Wang, R., Wu, Z. et al. RETRACTED ARTICLE: Circular RNA ITCH suppressed prostate cancer progression by increasing HOXB13 expression via spongy miR-17-5p. Cancer Cell Int 19, 328 (2019). https://doi.org/10.1186/s12935-019-0994-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12935-019-0994-8