
Abstract
Background
Wastewater treatment plants contribute approximately 6% of anthropogenic methane emissions. Methanotrophs, capable of converting methane into polyhydroxybutyrate (PHB), offer a promising solution for utilizing methane as a carbon source, using activated sludge as a seed culture for PHB production. However, maintaining and enriching PHB-accumulating methanotrophic communities poses challenges.
Results
This study investigated the potential of Methylosinus trichosporium OB3b to bioaugment PHB-accumulating methanotrophic consortium within activated sludge to enhance PHB production. Waste-activated sludges with varying ratios of M. trichosporium OB3b (1:0, 1:1, 1:4, and 0:1) were cultivated. The results revealed substantial growth and methane consumption in waste-activated sludge with M. trichosporium OB3b-amended cultures, particularly in a 1:1 ratio. Enhanced PHB accumulation, reaching 37.1% in the same ratio culture, indicates the dominance of Type II methanotrophs. Quantification of methanotrophs by digital polymerase chain reaction showed gradual increases in Type II methanotrophs, correlating with increased PHB production. However, while initial bioaugmentation of M. trichosporium OB3b was observed, its presence decreased in subsequent cycles, indicating the dominance of other Type II methanotrophs. Microbial community analysis highlighted the successful enrichment of Type II methanotrophs-dominated cultures due to the addition of M. trichosporium OB3b, outcompeting Type I methanotrophs. Methylocystis and Methylophilus spp. were the most abundant in M. trichosporium OB3b-amended cultures.
Conclusions
Bioaugmentation strategies, leveraging M. trichosporium OB3b could significantly enhance PHB production and foster the enrichment of PHB-accumulating methanotrophs in activated sludge. These findings contribute to integrating PHB production in wastewater treatment plants, providing a sustainable solution for resource recovery.
Graphical abstract

Similar content being viewed by others

Background
Methane emissions from wastewater treatment plants
Methane (CH4) stands out as a significant greenhouse gas, contributing to annual emissions of 576 Tg CH4 [1]. Anthropogenic methane sources, including agriculture, solid waste, coal mining, and wastewater treatment plants, collectively account for up to 60% of global methane emissions. Among these sources, the wastewater treatment plant sector is particularly notable, contributing 5–8% of methane emissions, followed by livestock (31%), oil and gas (26%), landfills (14%), and coal mining (11%) [2]. In the United States, centralized wastewater treatment plants are estimated to produce approximately 10.9 ± 7.0 MMT CO2-eq methane annually, with 72% attributed to plants equipped with anaerobic digestion [3].
Recent studies have explored diverse methodologies for valorizing methane into high-value-added products, such as heat and power generation, biopolymer production (e.g., polyhydroxybutyrate), and biofuel synthesis [4,5,6]. Incorporating polyhydroxybutyrate (PHB) production into wastewater treatment plants has emerged as a cost-effective alternative for methane valorization [7, 8]. PHB, a biopolymer fully degradable in natural environments, including soil, oceans, and landfills [9], presents a sustainable avenue. The substantial activation energy of C-H bonds (439 kJ/mol) in methane presents a formidable barrier to chemical oxidation processes [10]. Leveraging methanotrophs as biological catalysts offers a promising solution to reduce high energy requirements and convert methane into PHB.
PHB production in methanotrophs
Methanotrophs, a subset of methylotrophs, utilize methane as a carbon and energy source for cell growth and biosynthesis [11]. These bacteria are classified into three phyla: Gammaproteobacteria, Alphaproteobacteria, and Verrucomicrobia [12,13,14]. Within proteobacteria, methanotrophs are further categorized into Type I and Type II based on the assimilation pathway of formaldehyde. Type I methanotrophs use the ribulose monophosphate (RuMP) cycle, whereas Type II methanotrophs use the serine cycle or the ethylmalonyl-CoA cycle (EMCP) [15,16,17]. Type II methanotrophs are notable for their production of PHB, through the serine pathway.
Unlike Type I methanotrophs, which have received some attention regarding PHB production, the majority of studies have focused on Type II methanotrophs [18, 19]. Notably, Type II methanotrophs, including Methylocystis, Methylosinus, Methylocapsa, and Methylocella, can yield substantial amounts of PHB [20], making them more efficient candidates for PHB production compared to Type I methanotrophs.
In the serine pathway, methane is ultimately condensed into acetyl-CoA, which enters two distinctive pathways based on conditions: the PHB cycle and the tricarboxylic acid (TCA) cycle. The PHB cycle is initiated with acetyl-CoA under nutrient-limited, typically nitrogen-restricted, conditions [4]. The TCA cycle operates under nutrient-sufficient conditions to derive energy sources. Key enzymes in the methanotrophic PHB cycle include beta-ketothiolase (phaA), acetoacetyl-CoA reductase (phaB), and PHB polymerase (phaC). During PHB synthesis, acetyl-CoA is condensed to acetoacetyl-CoA by phaA, subsequently reduced to (R)-3-hydroxybutyryl-CoA by phaB. Finally, phaC converts this precursor to PHB [9, 19].
PHB production process involves two stages: cell growth and PHB accumulation [21]. In the first stage, rapid methanotroph proliferation occurs under nutrient-rich conditions, generating ample active biomass. The primary goal of this stage is to achieve a high concentration of active biomass to maximize PHB production. Subsequently, in the production stage, PHB granules are synthesized within cells under nitrogen-limited conditions, impeding cellular proliferation. Optimizing nutritional and environmental conditions enhances PHB production efficiency. Nevertheless, the selection of proficient PHB-producing methanotrophs characterized by elevated biomass concentration, PHB content, and expedited growth rates stands as a pivotal determinant in this process [22]. In addition, contamination resistance and adaptability are crucial indicators for industrial-scale PHB production with methanotrophs.
Selection of Type II methanotrophic consortium
The production of PHB is heavily reliant on the enrichments of Type II methanotrophs, which possess the capability to synthesize intracellular PHB. Integrating PHB production seamlessly into wastewater treatment plants and harnessing waste resources such as biogas can be achieved by utilizing activated sludge to enrich cultures dominated by Type II methanotrophs. This approach eliminates the sterilization costs associated with operating pure methanotroph bioreactors [22, 23]. The inclusion of mixed consortia, comprising methylotrophs and heterotrophs, may stimulate the microbial growth of methanotrophs either by consuming toxic metabolites (e.g., methanol and formate) or providing essential byproducts (e.g., quinone, pyridoxine, and vitamin B12) [24, 25]. However, constructing stable and effective microbial communities for PHB production, particularly with Type II methanotrophic bacteria, remains a challenge.
Various approaches have been explored to adjust environmental and nutritional factors (e.g., pH, methane concentration, nitrogen source, and copper) for selecting Type II methanotrophs in activated sludge. Although altering methane concentrations affects microbial population structure, it often results in the dominance of Type I methanotrophs over Type II methanotrophs [26]. Nitrogen-selection methods, such as using N2 gas with low O2 concentration, have been employed to form Type II methanotrophic enrichments. However, slow growth has been observed compared to other nitrogen sources (i.e., ammonium and nitrate) [27, 28]. Efficient PHB production requires faster growth, high biomass concentration, and elevated PHB content. Strategies have been employed to counter ammonium inhibition and prevent the invasion of Type I methanotrophs, leveraging the higher resistance of Type II methanotrophs to ammonium toxicity under elevated concentrations [29,30,31]. Despite the progress, selecting Type II methanotrophs still demands a considerable timeframe and may result in an unstable microbial community structure.
This study aims to assess the potential of M. trichosporium OB3b in activated sludge for bioaugmenting Type II methanotrophs in a mixed culture, with a focus on PHB production. This approach aims to establish robust Type II methanotrophic enrichments and expedite the start-up time. The presence of methanotrophs was confirmed by monitoring microbial growth and gas consumption (i.e., methane and oxygen). Digital quantitative polymerase chain reaction was employed to track the bioaugmentation of Type II methanotrophs and M. trichosporium OB3b. Microbial community composition was analyzed through 16S rRNA gene amplicon sequencing to identify accompanying methylotrophs and heterotrophs. Quantitative analysis of PHB was performed by gas chromatography equipped with a flame ionization detector to demonstrate the capability of PHB production. This approach holds promise for implementation in wastewater treatment plants to enhance PHB production for waste resource recovery, encompassing methane and activated sludge. Consequently, it could represent a definitive strategy for improving the resilience and stability of obtaining Type II methanotrophic enrichment.
Methods
Bacterial strain and culture medium
Activated sludge samples were collected from the second sediment aerobic basin located at the Daejeon wastewater treatment plant in Republic of Korea (36.38179°N, 127.40677°E). Upon immediate transfer to the laboratory, the samples were filtered with a 100 μm cell strainer to remove large materials. Methylosinus trichosporium OB3b, obtained from Prof. Sukhwan Yoon’s lab at KAIST (Daejeon, Republic of Korea), was precultured before commencing the experiments.
For cultivation, the ammonia mineral salt medium (AMS) was chosen due to its superior ability to accumulate polyhydroxybutyrate (PHB) compared to nitrate mineral salt medium (NMS) [32,33,34]. The AMS medium contained (per 1 L): 1 g of MgSO4.7H2O, 0.2 g of CaCl2 · 2H2O, 0.1 mL of a 3.8% (w/v) Fe(III)-EDTA solution, 0.5 mL of a 0.1% (w/v) Na2MoO4·2H2O solution, and 1 mL of a trace element solution. The trace element solution contained (per L): 0.5 g of FeSO4 ·7H2O, 0.4 g of ZnSO4·7H2O, 0.02 g of MnCl2·7H2O, 0.05 g of CoCl2· 6H2O, 0.01 g of NiCl2· 6H2O, 0.015 g of H3BO3 and 0.25 g of EDTA [35].
In 160 mL serum bottles, 43 mL of AMS medium was distributed and sealed with butyl rubbers and aluminum caps. After autoclaving, the media were supplemented with 0.5 mL of 10x vitamin stock, 0.5 mL of 0.4 M of phosphate buffer solution, and 1 mL of 0.5 mM CuCl2·2H2O solution. The 10x vitamin stock contained (per 1 L): 20 mg of biotin, 20 mg of folic acid, 50 mg of thiamine HCl, 50 mg of Ca pantothenate, 1 mg of vitamin B12, 50 mg of riboflavin, and 50 mg of nicotinamide. The 10x vitamin stock was filter-sterilized using a 0.22-µm syringe filter (Advantec, cellulose-acetate membrane). The phosphate buffer solution, containing 13 g of KH2PO4 and 31 g of Na2HPO4·7H2O per 500 mL, was used for pH adjustment to 6.8.
Methanotrophic enrichments and PHB accumulation
M. trichosporium OB3b (referred to as OB3b hereafter), initially with an optical density of 0.2, was supplemented with activated sludge (referred to as AS) in serum bottles based on a biomass ratio. The inoculation ratios of OB3b to AS (w/w) were 1:0, 1:1, 1:4, and 0:1. A pure culture of M. trichosporium OB3b was used as a control to evaluate PHB production and methanotrophic activity (refer to Fig. 1 for an experiment schematic). These pure cultures were not subjected to repetitive cycles to minimize the risk of contamination.

Schematic diagram of growth cycle and PHB accumulation for methanotrophic mixed cultures
Fresh AMS medium (45 mL) was inoculated with 5 mL of activated sludge with and without OB3b. For continuous growth, 10% of the previously grown culture was transferred into fresh AMS medium every 3 d to enrich the Type II methanotrophic culture. After the growth cycle, the residual culture was centrifuged at 3,500 rpm for 25 min to form a pellet. The pellet was resuspended in 5 mL of PHB medium and then injected into 45 mL of PHB medium. The PHB medium contained the same components as AMS medium but lacked ammonium. The PHB accumulation cycle was performed under nitrogen-depleted conditions for 3 d. CH4 and O2 (99.999%, Cheil Gas Industry Co., Republic of Korea) were supplied to the 110 mL of headspace and maintained at a 1:1 (v/v) ratio. All cultures were conducted in triplicate, agitated horizontally at 150 rpm at 30 ℃ in a shaking incubator.
Analytical method
CH4 and O2 concentrations were analyzed using gas chromatography (GC) (Agilent 8890, USA) equipped with Porapak Q and MolSieve 5 A columns and a thermal conductivity detector (TCD). The injector temperature was set at 200 ℃, with argon gas serving as the carrier gas. The oven temperature program follows as: 50 ℃ for 2 min, and 150 ℃ for 2 min. For gas monitoring, 150 µL of gas in the headspace was sampled by a Hamilton-gas-tight syringe and directly injected into the GC.
Microbial growth was monitored by measuring the optical density at 600 nm by a UV-Vis spectrophotometer (Thermo Scientific, Genesys-180, USA).
PHB quantification
The PHB content was measured by GC following established protocols [36]. After the PHB accumulation cycle, all liquid cultures were harvested, centrifuged at 3,500 rpm for 25 min, and the pelletized cells were freeze-dried, weighed, and transferred into 12 mL glass vials. Each sample underwent cell lysis, with the addition of 2 mL chloroform and 2 mL acidified methanol containing 3% sulfuric acid and benzoic acid (0.25 mg/ mL methanol). Benzoic acid and 3-hydroxybutyric acid salt (Sigma-Aldrich) were used as internal and external standards, respectively. Sealed with Teflon-lined plastic caps, all samples were heated at 100 ℃ for 3.5 h and then cooled to room temperature. To separate the organic and aqueous phases, 1 mL of deionized water was added to each sample, followed by vortexing for 30 s. The organic phase was extracted and analyzed by GC (Agilent 8890, USA) equipped with an HP-5 column and flame ionization detector (FID). The oven temperature program follows as: 160 °C for 4 min, 200 °C for 4 min, and 275 °C for 6 min.
Digital polymeric chain reaction and microbial community analysis
Cell suspensions were collected from cycle 1, cycle 4, and cycle 12 after 72 h of growth incubation to monitor microbial community shifts using digital PCR (dPCR) and 16S rRNA gene amplicon sequencing. DNA was extracted using the DNeasy PowerSoil Pro kit (Qiagen, Hilden, Germany) following the manufacturer’s protocol. Copy concentrations of 16S rRNA bacteria, Type II methanotrophs, and M. trichosporium OB3b were quantified by dPCR, employing specific target primers (Table S1). dPCR reaction mixtures were prepared using the QIAcuity EvaGreen PCR kit (Qiagen, Hilden, Germany), as per the manufacturer’s protocol and loaded onto a QIAcuity Nanoplate 26 K 24-well. The reactions were analyzed in a QIAcuity ONE 2-Plex system (Qiagen, Hilden, Germany) using the following thermal cycling program: an initial heat activation step at 95 ℃ for 2 min, 40 cycles of amplification (denaturation at 95 ℃ for 15 s, annealing at 55 ℃ (for total bacteria) and 58 ℃ (for Type II methanotrophs and M. trichosporium OB3b) for 15 s, final extension at 72 ℃ for 15 s), and a cooling step at 40 ℃ for 5 min [37].
To assess the microbial composition of the cultures, the V3-V4 region of the 16S rRNA gene was amplified and sequenced on the Illumina Miseq platform (Macrogen Inc., Seoul, Republic of Korea).
Results and discussions
Growth characteristics of M. trichosporium OB3b-amended cultures
Activated sludges with M. trichosporium OB3b (OBAS) were incubated at varying inoculation ratios. Biotic controls, consisting of activated sludge (AS-only) and pure M. trichosporium OB3b (OB3b-only), were employed to assess methanotrophic activity and PHB synthesis capabilities relative to M. trichosporium OB3b-amended cultures (OBAS (1:4), and OBAS (1:1)). Optical density at 600 nm was used to monitor the microbial growth. Microbial growth was monitored using optical density at 600 nm, and cycles 1, 4, and 12 were analyzed for microbial growth, CH4 consumption, and O2 consumption (Fig. 2).

Microbial growth curves, CH4, and O2 consumption were monitored at Cycle 1 (A, D, G), Cycle 4 (B, E, H), and Cycle 12 (C, F, I) in OBAS and AS-only cultures
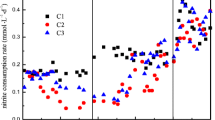
In Cycle 1, OBAS cultures, particularly OBAS (1:1), exhibited higher optical density compared to OB3b-only cultures, reaching a final optical density of 0.659 ± 0.056. Approximately 31% of CH4 and 40% of O2 were consumed within 72 h in OBAS (1:1) cultures, with a CH4 to O2 consumption ratio of 1.29, slightly lower than that observed in previous studies [19, 38,39,40]. AS-only and OBAS (1:4) cultures displayed lower optical densities of 0.371 ± 0.150, and 0.176 ± 0.031, respectively. Both cultures experienced a prolonged lag phase (within 50 h) and consumed about 8% and 11% of CH4, respectively. This suggests that AS-only and OBAS (1:4) cultures did not result in methanotrophic enrichments due to their lack of adaptation to the presence of excess methane.
In Cycle 4, initially injected CH4 was completely consumed in all cultures, accompanied with oxygen depletion. Compared to Cycle 1, faster growth and CH4 consumption were observed in OBAS (1:4) and AS-only cultures, indicating the enrichment of methanotrophs. However, AS-only cultures exhibited poor growth in Cycle 12, suggesting lower methanotrophic activity and a smaller portion of methanotrophs in the microbial community. This result indicated that the enrichment of Type II methanotrophs in AS-only cultures may not have been successful in maintaining the robust PHB-accumulating methanotrophic consortia over successive cycles. The optical density of OBAS cultures (1:1, and 1:4) in Cycle 12 increased to 0.807 ± 0.049, and 0.829 ± 0.031, respectively. Lag phases of each culture were within 18 h, reaching an exponential phase faster than Cycle 1. Notably, CH4 and O2 in OBAS (1:4) were completely consumed within 48 h, and cultures were refilled with methane and oxygen mixtures (1:1 v/v). No methane oxidation was observed after refilling. The methane-to-oxygen consumption ratio of OBAS (1:1, and 1:4) was calculated as 1.02 and 1.28, respectively. Previous studies reported that 1.5–2.0 moles of oxygen were required to oxidize 1 mol of methane in methanotrophic enrichments and M. trichosporium OB3b [36, 38, 39, 41]. Significant amounts of CH4 and O2 consumption were observed in OBAS cultures, even exceeding the levels seen in OB3b-only cultures.
PHB accumulation of M. trichosporium OB3b-amended cultures
PHB accumulation tests were conducted over a 72 h period following the growth cycle to evaluate the PHB production capability. Cells harvested during this period were transferred into a nitrogen-depleted medium to induce PHB accumulation in the presence of excess methane. PHB content was analyzed at cycles 1,4, and 12 (Fig. 3).

PHB concentration (mg/L), biomass concentration (mg/L), and PHB content (%) in OBAS, AS-only cultures, and OB3b-only cultures
In the case of AS-only culture, no PHB accumulation was observed throughout all cycles, suggesting that Type II methanotrophs did not dominate in the enriched cultures. Conversely, Type I methanotrophs may have dominated, as evidenced by methane consumption in Sect. 3.1. The prevalence of Type II methanotrophs is crucial for effective PHB production. PHB contents in OBAS (1:1) cultures significantly and consistently increased from 5.3 ± 1.2% to 37.1 ± 5.3%, alongside an augmented biomass concentration. This suggests that the dominance of Type II methanotrophs, including M. trichosporium OB3b, in OBAS (1:1) cultures substantially occurred. OBAS (1:4) cultures accumulated up to 6.4 ± 1.8% (Cycle 1), and 15.1 ± 1.0% (Cycle 12) of PHB, with higher contents observed as the number of cycles increased. Notably, the addition of M. trichosporium OB3b had a significant impact on PHB content, as evidenced by the observed higher PHB contents. The maximum PHB content in OBAS (1:1) was comparable to the mixed methanotrophic consortia in previous studies, where Methylocystis-dominated cultures achieved up to 39% of PHB accumulation [36]. Similarly, PHB contents of 37.8% and 40% were recorded in mixed methanotrophic consortia [33, 42]. In comparison to OB3b-only cultures, higher PHB contents were obtained in OB3b-amended cultures, indicating the synergistic effect of mixed methanotrophic consortia [43].
In the context of PHB production, biomass concentration, and PHB content were closely related, affecting the PHB concentration. The concentration of PHB in cells, determined through the analysis of cell dry weight and PHB content, revealed that OBAS (1:1) cultures exhibited the most substantial biomass and PHB concentrations, amounting to 271 ± 1.0 mg/L and 95.7 ± 9.2 mg/L, respectively (Fig. 3). These values were 2.30 and 2.65 times higher than those of OB3b-only cultures, respectively, implying that non-methanotrophs in mixed methanotrophic consortia might support their microbial growth. Similar increasing trends were observed in OB3b-amended cultures concerning PHB concentration, biomass concentration, and PHB content. Consequently, the addition of M. trichosporium OB3B significantly enhanced the capability of PHB production (Fig. 3).
Quantification of Type II methanotrophs and M. trichosporium OB3b
To investigate the dominance of Type II methanotrophs and M. trichosporium OB3b in all cultures, dPCR was employed to quantify the microbial population. Copy concentrations of the bacterial 16S rRNA gene were measured to determine the ratio of Type II methanotrophs and M. trichosporium OB3b in the enrichments, with Type II methanotrophs representing PHB accumulating methanotrophs. 16S rRNA copy concentration of Type II methanotrophs referred as Type II methanotrophs, 16S rRNA copy concentration of bacteria referred as total bacteria, and pmoA gene copy concentration of M. trichosporium OB3b referred as M. trichosporium OB3b in this section.
The initial 16S rRNA gene copy concentration of Type II methanotrophs in each culture under initial condition (0 h in Cycle 1) was 1.7 × 104 copies/µL, 1.4 × 104 copies/µL, and 5.1 × 103 copies/µL for OBAS (1:1, and 1:4) cultures and AS-only cultures (Fig. S1 and Fig. S2). The 16 S rRNA gene copy concentration of Type II methanotrophs increased gradually from 2.1 × 104 copies/µL to 3.5 × 104 copies/µL in OBAS (1:1) cultures (Fig. 4A). Importantly, the Type II methanotrophs to total bacteria ratio in Cycle 12 (44%) was twice as high as in Cycle 1 (22%), and 3.4 times higher than the initial condition (13%) (refer to Fig. 4A and Fig. S2). This result aligns with the PHB production discussed in Sect. 3.2. For OBAS (1:4) cultures, the 16S rRNA gene copy concentration of Type II methanotrophs in OBAS (1:4) cultures remained in the range between 1.2 × 104 copies/µL and 1.3 × 103 copies/µL, notably slightly higher in Cycle 4 and Cycle 12. However, the Type II methanotrophs to total bacteria ratio exhibited notable differences, showing 20% at the beginning, decreasing to 5%, and then increasing to 16% (Fig. 4). In contrast, AS-only culture showed that the proportion of Type II methanotrophs to total bacteria was approximately 1%. This outcome explains why AS-only culture accumulated a lower PHB content. In the OBAS (1:1) cultures, specifically, the relatively higher initial abundance of M. trichosporium OB3b expedited the establishment of conditions conducive to the prevalence of Type II methanotrophs. Conversely, in the 1:4 ratio condition, this establishment for favoring the dominance of PHB-accumulating methanotrophs was delayed. This resulted in an initial decline in Type II methanotrophs in Cycle 4, followed by a subsequent increase. Consequently, the initial M. trichosporium OB3b inoculation could have significantly influenced the establishment period of microbial community composition and targeted community composition, which can synthesize and utilize PHB.

The gene copy concentrations of total bacteria, Type II methanotrophs (A), and M. trichosporium OB3b (B) in OBAS and AS-only cultures. The percentage of type II methanotrophs (A) and M. trichosporium OB3b (B) were monitored. Below 100 copies/µL was considered a limit of detection (LOD)
The most intriguing observations were made in the pmoA gene copy concentration of M. trichosporium OB3b. The pmoA gene copy concentrations of M. trichosporium OB3b in cycle 1 were 2.9 × 104 copies/µL and 1.2 × 104 copies/µL in OBAS (1:1, and 1:4), respectively. The M. trichosporium OB3b to total bacteria ratio in Cycle 1 were 30% and 20%, respectively, identical to the results of Type II methanotrophs gene copy concentration. Under the initial conditions (0 h in Cycle 1), the M. trichosporium OB3b to total bacteria in OBAS (1:1, and 1:4) were 13%, and 5%, respectively (Fig. S2). This implies that bioaugmentation of M. trichosporium OB3b occurred in the first cycle. On the contrary, the M. trichosporium gene copy concentrations of AS-only cultures were below the detection limit (100 copies/µL), indicating the absence of M. trichosporium OB3b. Both OBAS cultures (1:1, and 1:4) decreased to 16 copies/µL and 384 copies/µL after cycle 1, respectively, whereas the copy numbers of OBAS (1:1) were below the detection limit. The percentages of M. trichosporium OB3b were almost 0% in all cultures after Cycle 1. Increased gene copy concentrations of Type II methanotrophs were anticipated for the successful bioaugmentation of M. trichosporium OB3b. However, the results of M. trichosporium OB3b gene copy concentration suggest that the effective bioaugmentation of M. trichosporium OB3b was only achieved after Cycle 1, rather than at the conclusion of Cycle 12. M. trichosporium OB3b-amended cultures might be dominated by other Type II methanotrophs (e.g., Methylocystis, Methylocella, and Methylocapsa).
Microbial community analysis
To qualitatively analyze microbial compositions, 16S rRNA amplicon sequencing was conducted. At the phylum level, Proteobacteria dominated, accounting for over 70% of OBAS cultures and activated sludge. Notably, the genera Methylocystis and Methylosinus emerged as representative PHB-accumulating methanotrophs. In OBAS cultures (1:1, and 1:4), the Methylosinus genus accounted for 39.39% and 28.75%, respectively, with higher prevalence in the initial stages. However, across repeating cycles, the Methylosinus genus’s relative abundance declined, giving way to the Methylocystis genus (Fig. 5). Post-bioaugmentation with M. trichosporium OB3b, the Methylocystis genus dominated at 49.17% in OBAS (1:1) cultures, the highest observed across all cultures. This observation correlates with the highest PHB contents noted in OBAS cultures.

Taxonomy relative abundance of OBAS and AS-only cultures at the phylum level (A), and genus level (B). Relative abundances below 1% are classified as ‘Others’
In the OBAS (1:1) culture, the proportion of Type II methanotrophs initially starts at 39.68% in Cycle 1 and later increases to 49.17% by Cycle 12. Concurrently, Type I methanotrophs, initially present in Cycle 1, became undetectable as cycles progressed. Likewise, the OBAS (1:4) culture exhibited Type II methanotrophs constituting 25.13% of the population by Cycle 12, with Type I methanotrophs also undetectable at the conclusion of the observation period (Fig. 5, Fig. S3, and Table 1). The invasion of Type I methanotrophs was not observed in the last cycle of OBAS cultures.
The consistent observations across both cultures imply a progressive increase in Type II methanotrophs coupled with a declining presence of Type I methanotrophs throughout repeated cycles. The stable maintenance of the microbial community structure corresponds with the predominance of Type II methanotrophs (Fig. 5 and Fig. S3). This indicates the successful establishment of a Type II methanotrophs-dominated culture. In contrast, AS-only cultures were dominated by Type I methanotrophs, constituting 33.11% (Cycle 1), 29.05% (Cycle 4), and 10.87% (Cycle 12) (Table 1) This outcome aligns with previous studies where Type I methanotrophs often predominate over Type II methanotrophs due to their higher affinity for methane [28, 30, 44]. Methyloparacoccus and Methylomonas were the most abundant in AS-only cultures (Fig. 5, and Table 2).
The relative abundance of Type II methanotrophs in AS-only cultures remained below 10%, suggesting that Type II methanotrophs require extended adaptation periods for their enrichment. From the perspective of methylotrophs, Methylophilus, a non-methanotrophic methylotroph capable of utilizing methanol, was frequently observed in methanotrophic enrichments in previous studies [33, 45,46,47,48,49]. Methanotrophic enrichments coexisting with methylotrophs and heterotrophs can eliminate toxic metabolites (e.g., methanol and formate) from methanotrophs’ methane oxidation. Additionally, Methylocystis- and Methylophilus-dominated cultures accumulated up to 59.4% of PHB during a more extended period of methane adaptation compared to this study [33].
Taxonomic classification and relative abundance of methanotrophs and methylotrophs are summarized (Tables 1 and 2). Importantly, the addition of M. trichosporium OB3b led to the outcompetition of Type I methanotrophs and successful enrichment into Type II methanotrophs-dominated cultures. Although initially M. trichosporium OB3b was dominant, Methylocystis spp. eventually became prevalent due to their versatility in oxygen and methane requirements. Methylocystis spp. is particularly robust under low methane conditions [50,51,52]. This is related to particulate methane monooxygenase A, which is the key enzyme of methane oxidation. In our batch feeding cycle, the batch supply of methane and oxygen, may be a critical factor favoring the growth of these methanotrophs after the methane consumption of M. trichosporium OB3b. Previous studies reported that nitrogen selection (i.e., ammonium inhibition), cyclic methane, and nitrogen starvation were involved in securing Type II methanotrophic enrichments, especially Methylocystis-dominated cultures [29, 30, 36, 53]. However, in our research, the bioaugmentation of M. trichosporium might play a vital role for enriching PHB-accumulating methanotrophic enrichments.
Conclusion
This study investigated the impact of incorporating pure methanotrophs into activated sludge, examining methane oxidation, PHB production, gene copy concentrations, and microbial community dynamics. Quantitative PHB accumulation tests demonstrated that only activated sludge bioaugmented with M. trichosporium OB3b has the capability of PHB production. Maximum PHB contents and concentration of 37% and 96 mg/L, respectively, were observed, significantly surpassing those in activated sludge without M. trichosporium OB3b. Quantification of M. trichosporium OB3b and Type II methanotrophs unveiled that the bioaugmentation of M. trichosporium OB3b occurred in the initial stages, triggering the involvement of other Type II methanotroph for efficient PHB production. Microbial community analysis suggested that Methylocystis-dominated cultures played a pivotal role in synthesizing PHB. To validate microbial shifts in activated sludge with M. trichosporium OB3b, metabolite analysis is recommended to confirm the influences of M. trichosporium OB3b in activated sludge. This approach could be an effective strategy to enrich Type II methanotrophs-dominated culture by bioaugmentation with M. trichosporium OB3b in an activated sludge system for enhancing PHB production and rapidly securing the PHB-accumulating methanotrophic enrichments.
Data availability
No datasets were generated or analysed during the current study.
References
Saunois M, Stavert AR, Poulter B, Bousquet P, Canadell JG, Jackson RB, et al. The global methane budget 2000–2017. Earth Syst Sci Data. 2020;12(3):1561–623. https://doi.org/10.5194/essd-12-1561-2020
Ocko IB, Sun TY, Shindell D, Oppenheimer M, Hristov AN, Pacala SW, et al. Acting rapidly to deploy readily available methane mitigation measures by sector can immediately slow global warming. Environ Res Lett. 2021;16(5). https://doi.org/10.1088/1748-9326/abf9c8
Song CH, Zhu JJ, Willis JL, Moore DP, Zondlo MA, Ren ZJ. Methane emissions from municipal wastewater collection and treatment systems. Environ Sci Technol. 2023;57(6):2248–61. https://doi.org/10.1021/acs.est.2c04388
Strong PJ, Laycock B, Mahamud SNS, Jensen PD, Lant PA, Tyson G, et al. The opportunity for high-performance biomaterials from methane. Microorganisms. 2016;4(1). https://doi.org/10.3390/microorganisms4010011
Jawaharraj K, Shrestha N, Chilkoor G, Dhiman SS, Islam J, Gadhamshetty V. Valorization of methane from environmental engineering applications: a critical review. Water Res. 2020;187. https://doi.org/10.1016/j.watres.2020.116400
Strong PJ, Xie S, Clarke WP. Methane as a resource: can the methanotrophs add value? Environ Sci Technol. 2015;49(7):4001–18. https://doi.org/10.1021/es504242n.
Levett I, Birkett G, Davies N, Bell A, Langford A, Laycock B, et al. Techno-economic assessment of poly-3-hydroxybutyrate (PHB) production from methane—the case for thermophilic bioprocessing. J Environ Chem Eng. 2016;4:3724–33. https://doi.org/10.1016/j.jece.2016.07.033. 4, Part A).
AlSayed A, Fergala A, Eldyasti A. Sustainable biogas mitigation and value-added resources recovery using methanotrophs intergrated into wastewater treatment plants. Rev Environ Sci Bio. 2018;17(2):351–93. https://doi.org/10.1007/s11157-018-9464-3
Tan GYA, Chen CL, Li L, Ge L, Wang L, Razaad IMN, et al. Start a research on biopolymer polyhydroxyalkanoate (PHA): a review. Polymers-Basel. 2014;6(3):706–54. https://doi.org/10.3390/polym6030706
Nguyen DTN, Lee OK, Nguyen TT, Lee EY. Type II methanotrophs: a promising microbial cell-factory platform for bioconversion of methane to chemicals. Biotechnol Adv. 2021;47. https://doi.org/10.1016/j.biotechadv.2021.107700
Hanson RS, Hanson TE. Methanotrophic bacteria. Microbiol Rev. 1996;60(2):439–71. https://doi.org/10.1128/mr.60.2.439-471.1996
Bowman JP, Sly LI, Nichols PD, Hayward AC. Revised taxonomy of the methanotrophs: description of methylobacter gen. nov., emendation of methylococcus, validation of methylosinus and methylocystis species, and a proposal that the family methylococcaceae includes only the group I methanotrophs. Int J Syst Evol Microbiol. 1993;43(4):735–53. https://doi.org/10.1099/00207713-43-4-735
Semrau JD, DiSpirito AA, Yoon S. Methanotrophs and copper. FEMS Microbiol. Rev. 2010;34(4):496–531. https://doi.org/10.1111/j.1574-6976.2010.00212.x
Schmitz RA, Peeters SH, Versantvoort W, Picone N, Pol A, Jetten MSM, et al. Verrucomicrobial methanotrophs: ecophysiology of metabolically versatile acidophiles. FEMS Microbiol Rev. 2021;45(5). https://doi.org/10.1093/femsre/fuab007
Jiang W, Villamor DH, Peng HD, Chen J, Liu L, Haritos V, et al. Metabolic engineering strategies to enable microbial utilization of C1 feedstocks. Nat Chem Biol. 2021;17(8):845–55. https://doi.org/10.1038/s41589-021-00836-0
Kalyuzhnaya MG, Puri AW, Lidstrom ME. Metabolic engineering in methanotrophic bacteria. Metab Eng. 2015;29:142–52. https://doi.org/10.1016/j.ymben.2015.03.010
Lee OK, Hur DH, Nguyen DTN, Lee EY. Metabolic engineering of methanotrophs and its application to production of chemicals and biofuels from methane. Biofuel Bioprod Bior. 2016;10(6):848–63. https://doi.org/10.1002/bbb.1678
Helm J, Wendlandt KD, Jechorek M, Stottmeister U. Potassium deficiency results in accumulation of ultra-high molecular weight poly‐β‐hydroxybutyrate in a methane‐utilizing mixed culture. J Appl Microbiol. 2008;105(4):1054–61. https://doi.org/10.1111/j.1365-2672.2008.03831.x.
Pieja AJ, Sundstrom ER, Criddle CS. Poly-3-Hydroxybutyrate metabolism in the type II Methanotroph OBBP. Appl Environ Microbiol. 2011;77(17):6012–9. https://doi.org/10.1128/Aem.00509-11
Pieja AJ, Rostkowski KH, Criddle CS. Distribution and selection of poly-3-hydroxybutyrate production capacity in methanotrophic proteobacteria. Microb Ecol. 2011;62(3):564–73. https://doi.org/10.1007/s00248-011-9873-0
Karthikeyan OP, Chidambarampadmavathy K, Cirés S, Heimann K. Review of sustainable methane mitigation and biopolymer production. Crit Rev Environ Sci Technol. 2015;45(15):1579–610. https://doi.org/10.1080/10643389.2014.966422.
Alcantara JMG, Distante F, Storti G, Moscatelli D, Morbidelli M, Sponchioni M. Current trends in the production of biodegradable bioplastics: the case of polyhydroxyalkanoates. Biotechnol Adv. 2020;42. https://doi.org/10.1016/j.biotechadv.2020.107582
Tan D, Wang Y, Tong Y, Chen G-Q. Grand challenges for industrializing polyhydroxyalkanoates (PHAs). Trends Biotechnol. 2021;39(9):953–63. https://doi.org/10.1016/j.tibtech.2020.11.010
Ho A, de Roy K, Thas O, De Neve J, Hoefman S, Vandamme P, et al. The more, the merrier: heterotroph richness stimulates methanotrophic activity. Isme J. 2014;8(9):1945–8. https://doi.org/10.1038/ismej.2014.74
Stock M, Hoefman S, Kerckhof FM, Boon N, De Vos P, De Baets B, et al. Exploration and prediction of interactions between methanotrophs and heterotrophs. Res Microbiol. 2013;164(10):1045–54. https://doi.org/10.1016/j.resmic.2013.08.006
López JC, Quijano G, Pérez R, Muñoz R. Assessing the influence of CH4 concentration during culture enrichment on the biodegradation kinetics and population structure. J Environ Manage. 2014;146:116–23. https://doi.org/10.1016/j.jenvman.2014.06.026.
Rostkowski KH, Pfluger AR, Criddle CS. Stoichiometry and kinetics of the PHB-producing type II methanotrophs methylosinus trichosporium OB3b and methylocystis parvus OBBP. Bioresour Technol. 2013;132:71–7. https://doi.org/10.1016/j.biortech.2012.12.129.
Pfluger AR, Wu W-M, Pieja AJ, Wan J, Rostkowski KH, Criddle CS. Selection of type I and type II methanotrophic proteobacteria in a fluidized bed reactor under non-sterile conditions. Bioresour Technol. 2011;102(21):9919–26. https://doi.org/10.1016/j.biortech.2011.08.054.
Fergala A, AlSayed A, Eldyasti A. Behavior of type II methanotrophic bacteria enriched from activated sludge process while utilizing ammonium as a nitrogen source. Int Biodeterior Biodegradation. 2018;130:8–16. https://doi.org/10.1016/j.ibiod.2018.03.010
Fergala A, AlSayed A, Eldyasti A. Factors affecting the selection of PHB accumulating methanotrophs from waste activated sludge while utilizing ammonium as their nitrogen source. J Chem Technol Biot. 2018;93(5):1359–69. https://doi.org/10.1002/jctb.5502
Hoefman S, van der Ha D, Boon N, Vandamme P, De Vos P, Heylen K. Niche differentiation in nitrogen metabolism among methanotrophs within an operational taxonomic unit. Bmc Microbiol. 2014;14. https://doi.org/10.1186/1471-2180-14-83
Zaldívar Carrillo JA, Stein LY, Sauvageau D. Defining nutrient combinations for optimal growth and polyhydroxybutyrate production by Methylosinus trichosporium OB3b using response surface methodology. Front Microbiol. 2018;9. https://doi.org/10.3389/fmicb.2018.01513
Salem R, Soliman M, Fergala A, Audette GF, ElDyasti A. Screening for methane utilizing mixed communities with high polyhydroxybutyrate (phb) production capacity using different design approaches. Polymers. 2021;13(10):1579. https://doi.org/10.3390/polym13101579
Lazic M, Gudneppanavar R, Whiddon K, Sauvageau D, Stein LY, Konopka M. In vivo quantification of polyhydroxybutyrate (PHB) in the alphaproteobacterial methanotroph, sp. Rockwell Appl Microbiol Biotechnol. 2022;106(2):811–9. https://doi.org/10.1007/s00253-021-11732-x
Whittenb.R, Phillips KC, Wilkinso.Jf. Enrichment, isolation and some properties of methane-utilizing bacteria. J Gen Microbiol. 1970;61:205–&. https://doi.org/10.1099/00221287-61-2-205
Myung J, Galega WM, Van Nostrand JD, Yuan T, Zhou J, Criddle CS. Long-term cultivation of a stable methylocystis-dominated methanotrophic enrichment enabling tailored production of poly (3-hydroxybutyrate-co-3-hydroxyvalerate). Bioresour Technol. 2015;198:811–8. https://doi.org/10.1016/j.biortech.2015.09.094
Yu H, Ko D, Lee C. Continuous cultivation of mixed-culture microalgae using anaerobic digestion effluent in photobioreactors with different strategies for adjusting nitrogen loading rate. Bioresour Technol. 2023;387. https://doi.org/10.1016/j.biortech.2023.129650
Knapp CW, Fowle DA, Kulczycki E, Roberts JA, Graham DW. Methane monooxygenase gene expression mediated by methanobactin in the presence of mineral copper sources. Proc Natl Acad Sci U S A. 2007;104(29):12040–5. https://doi.org/10.1073/pnas.0702879104
Rodríguez Y, Firmino PIM, Arnáiz E, Lebrero R, Muñoz R. Elucidating the influence of environmental factors on biogas-based polyhydroxybutyrate production by CSC1. Sci. Total Environ. 2020;706. https://doi.org/10.1016/j.scitotenv.2019.135136
Amabile C, Abate T, Chianese S, Musmarra D, Muñoz R. Assessing the performance of synthetic co-cultures during the conversion of methane into poly(3-hydroxybutyrate). Chem Eng J. 2023;476. https://doi.org/10.1016/j.cej.2023.146646
Bordel S, Rodríguez Y, Hakobyan A, Rodríguez E, Lebrero R, Muñoz R. Genome scale metabolic modeling reveals the metabolic potential of three type II methanotrophs of the genus. Metab Eng. 2019;54:191–9. https://doi.org/10.1016/j.ymben.2019.04.001
Myung J, Wang ZY, Yuan T, Zhang P, Van Nostrand JD, Zhou JZ, et al. Production of nitrous oxide from nitrite in stable type II methanotrophic enrichments. Environ Sci Technol. 2015;49(18):10969–75. https://doi.org/10.1021/acs.est.5b03385
Goers L, Freemont P, Polizzi KM. Co-culture systems and technologies: taking synthetic biology to the next level. J R Soc Interface. 2014;11(96). https://doi.org/10.1098/rsif.2014.0065
Knief C, Kolb S, Bodelier PLE, Lipski A, Dunfield PF. The active methanotrophic community in hydromorphic soils changes in response to changing methane concentration. Environ Microbiol. 2006;8(2):321–33. https://doi.org/10.1111/j.1462-2920.2005.00898.x
Pérez R, Cantera S, Bordel S, García-Encina PA, Muñoz R. The effect of temperature during culture enrichment on methanotrophic polyhydroxyalkanoate production. Int Biodeterior Biodegradation. 2019;140:144–51. https://doi.org/10.1016/j.ibiod.2019.04.004
Luangthongkam P, Strong PJ, Mahamud SNS, Evans P, Jensen P, Tyson G, et al. The effect of methane and odd-chain fatty acids on 3-hydroxybutyrate (3HB) and 3-hydroxyvalerate (3HV) synthesis by a Methylosinus-dominated mixed culture. Bioresour Bioprocess. 2019;6(1). https://doi.org/10.1186/s40643-019-0285-1
Zhang TT, Wang XW, Zhou JT, Zhang Y. Enrichments of methanotrophic-heterotrophic cultures with high poly-β-hydroxybutyrate (PHB) accumulation capacities. J Environ Sci. 2018;65:133–43. https://doi.org/10.1016/j.jes.2017.03.016
Kulkarni PP, Chavan SB, Deshpande MS, Sagotra D, Kumbhar PS, Ghosalkar AR. Enrichment of Methylocystis dominant mixed culture from rice field for PHB production. J Biotechnol. 2022;343:62–70. https://doi.org/10.1016/j.jbiotec.2021.11.007
Cardoso LOB, Karolski B, Gracioso LH, Borrego BB, Do Nascimento CAO, Perpetuo EA. Enrichment of Methylosinus-dominant consortia from mangroves for polyhydroxybutyrate (PHB) production. J Environ Chem Eng. 2022;10(5). https://doi.org/10.1016/j.jece.2022.108490
Knief C. Diversity and habitat preferences of cultivated and uncultivated aerobic methanotrophic bacteria evaluated based on pmoA as molecular marker. Front Microbiol. 2015;6:170928. https://doi.org/10.3389/fmicb.2015.01346.
Knief C, Dunfield PF. Response and adaptation of different methanotrophic bacteria to low methane mixing ratios. Environ Microbiol. 2005;7(9):1307–17. https://doi.org/10.1111/j.1462-2920.2005.00814.x.
Safitri AS, Hamelin J, Kommedal R, Milferstedt K. Engineered methanotrophic syntrophy in photogranule communities removes dissolved methane. Water Res X. 2021;12:100106. https://doi.org/10.1016/j.wroa.2021.100106.
Pieja AJ, Sundstrom ER, Criddle CS. Cyclic, alternating methane and nitrogen limitation increases PHB production in a methanotrophic community. Bioresour Technol. 2012;107:385–92. https://doi.org/10.1016/j.biortech.2011.12.044.
Funding
This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIT) (2022R1A4A3029607, 2022R1F1A1064064, and RS-2023-00209472) and Korean Ministry of Land, Infrastructure and Transport (MOLIT) as Innovative Talent Education Program for Smartcity. This research was also funded by the Ministry of Oceans and Fisheries, Korea (20200104).
Author information
Authors and Affiliations
Contributions
HE and JM conceived and designed the study. HE performed cultivation, gas analysis, and PHB analysis. HE and DK performed the digital PCR analysis and interpreted the data. HE wrote orginal draft. HE, CL, and JM wrote and revised the manuscript. All authors read and approved to submit the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
All authors approved the manuscript.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Eam, H., Ko, D., Lee, C. et al. Methylosinus trichosporium OB3b bioaugmentation unleashes polyhydroxybutyrate-accumulating potential in waste-activated sludge. Microb Cell Fact 23, 160 (2024). https://doi.org/10.1186/s12934-024-02442-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12934-024-02442-w