
Abstract
Background
Trehalose is related to several types of stress responses, especially freezing response in baker’s yeast (Saccharomyces cerevisiae). It is desirable to manipulate trehalose-related genes to create yeast strains that better tolerate freezing-thaw stress with improved fermentation capacity, which are in high demand in the baking industry.
Results
The strain overexpressing MAL62 gene showed increased trehalose content and cell viability after prefermention-freezing and long-term frozen. Deletion of NTH1 in combination of MAL62 overexpression further strengthens freezing tolerance and improves the leavening ability after freezing-thaw stress.
Conclusions
The mutants of the industrial baker’s yeast with enhanced freezing tolerance and leavening ability in lean dough were developed by genetic engineering. These strains had excellent potential industrial applications.
Similar content being viewed by others


Background
Frozen dough technology has been used in bakery industry to provide consumers with high-quality fresh bakery and convenience. However, cellular macromolecules, including proteins, nucleic acids and lipids of the yeast used in frozen dough, could be seriously damaged under the freezing and the subsequent thawing treatments, leading to inhibition of cell growth, cell viability and the leavening ability [1].
A great body of knowledge is already available regarding the molecular responses of the baker’s yeast (Saccharomyces cerevisiae) to frozen dough-associated stresses [2]. Among other molecules, trehalose has been highlighted due to its main function as a protective molecule in stress response [3]. This effect is achieved either by protecting membrane integrity through the union with phospholipids [4], or by preserving the native conformation of proteins and preventing aggregation of partially denatured proteins [5].
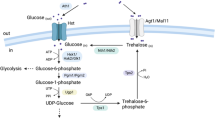
When yeast cells suffer from freezing stress, they accumulate large amounts of trehalose [6]. The accumulation is mainly induced by the classical the UDPG-dependent trehalose synthesis pathway, or referred as system I. It contains a trehalose-6-phosphate synthase encoded by TPS1 [7], a trehalose-6-phosphate phosphatase encoded by TPS2 [8] and a trehalose-synthesis protein complex encoded by TSL1 [9]. In addition, an alternative trehalose synthesis pathway, called ADPG-dependent trehalose synthesis pathway or the system II, has been proposed [10, 11]. It is specifically linked to maltose utilization.
Maltose metabolism in yeast depends on at least one of the five unlinked MAL loci (MAL1 through MAL4 and MAL6). A typical MAL locus consists of a MALx1 (MALxT) gene (where x is the locus), encoding maltose permease, a MALx2 (MALxS) gene, coding for alpha-glucosidase (maltase), and a MALx3 (MALxR) gene, encoding a positive regulatory protein [9]. It is reported that the expression of any one of the MAL loci in MAL-constitutive strains could elicit a maltose-induced trehalose accumulation [11]. Studies have shown that maltose and trehalose seem to share a common regulating mechanism [17, 18]. The maltose permease has been considered the rate-limiting enzyme in the MAL genes induction and maltose metabolism [4, 6, 12]. Hence, attempts to increase the trehalose content by system II had been concentrated on the modification of maltose permease or the entire MAL gene cluster [19–21]. However, recent studies showed that the alpha-glucosidase (maltase) is more important than maltose permease in maltose metabolism and leavening ability of baker’s yeast in lean dough [22–24]. In addition, the system II might be dependent of the system I, due to the fact that the system II is completely prevented when TPS1, a key gene in system I, is deleted [12].
Trehalose degradation could also be induced under certain stress [13, 14]. The best characterized trehalase is the neutral trehalase encoded by the NTH1 gene, which is induced by stress, such as heat. Nth1p is involved in thermos-tolerance and hydrolyzes intracellular trehalose into glucose [15, 16]. Deletion of NTH1 results in accumulation of trehalose, and heat sensitivity.
To better understand the role of trehalose in freezing tolerance of baker’s yeast in lean dough, and its possible mechanism, we investigated the effects of overexpression of MAL62, the gene encoding an alpha glucosidase, and deletion of NTH1 gene, on trehalose accumulation and on the freezing tolerance and leavening ability of baker’s yeast in lean dough.
Methods
Strains, plasmids and growth conditions
The genetic properties of all S. cerevisiae strains and plasmids used in the present study are summarized in Table 1. The BY14a was selected as a high leavening capacity haploid from 32 clones derived from the diploid BY14 strain, which has been maintained at the Tianjin Key Laboratory of Industrial Microbiology, Tianjin University of Science and Technology.
Recombinant DNA was amplified in Escherichia coli DH5a. Transformants were grown in Luria–Bertani medium (10 g/L tryptone, 5 g/L yeast extract, and 10 g/L NaCl) with 100 mg/L ampicillin. The plasmid was obtained using Plasmid Mini Kit II (D6945, Omega, USA).
The yeast strain was grown at 30 °C in yeast extract peptone dextrose (YEPD) medium (10 g/L yeast extract, 20 g/L peptone, and 20 g/L dextrose). Approximately 800 mg/L of G418 was added to the YEPD plates for selecting Geneticin (G418)-resistant transformants. After cultivation in YEPD for 24 h, 20 mL of the cell culture was inoculated into 200 mL of cane molasses medium (5 g/L yeast extract, 0.5 g/L (NH4)2SO4, and 12° Brix cane molasses) at the initial OD600 = 0.4 and cultivated for 24 h at 30 °C with 180 rpm rotary shaking to the final OD600 = 1.8. Cells were harvested through centrifugation (4 °C, 1500×g, 5 min) and were washed twice with sterile water at 4 °C for the succeeding fermentation experiments. To investigate the degradation of trehalose during prefermentation and the freezing tolerance, a modified the low sugar model liquid dough (LSMLD) medium was used [17]. The modified medium contains 2.5 g/L (NH4)2SO4, 5 g/L urea, 16 g/L KH2PO6, 5 g/L Na2HPO4, 0.6 g/L MgSO4, 22.5 mg/L nicotinic acid, 5 mg/L Ca-pantothenate, 2.5 mg/L thiamine, 1.25 g/L pyridoxine, 1 mg/L riboflavin, and 0.5 mg/L folic acid and carbon sources (33.25 g/L maltose with 5 g/L glucose).
Plasmid construction and yeast transformation
Genomic yeast DNA was prepared from the industrial baker’s yeast BY14a using a yeast DNA kit (D3370-01, Omega, Norcross, GA, USA). Table 2 shows the PCR primers used in this study.
Plasmid Yep-PMK (Yep-PGK1-MAL62-KanMX), an episomal plasmid with MAL62 under the control of the constitutive yeast phosphoglycerate Kinase gene (PGK1) promoter (PGK1 P ) and terminator (PGK1 T ), was constructed as follows: a KpnI/BamHIKanMX fragment, which was the dominant selection marker during yeast conversion, was amplified through PCR using pUG6 as template with Kan-U and Kan-D primers, and was cloned to the Yep352 vector to construct the empty plasmid Yep-K (Yep-KanMX). A XhoI fragment of MAL62 amplified with MAL62-U and MAL62-D primers from the genomes of the parental strain BY14a was inserted into the PGK1 fragment of pPGK1 vector and resulted in plasmid pPGKM. Then, the BamHI fragment of PGKM (the entire PGK1 and the inserted MAL62) amplified with PGK-U and PGK-D from pPGKM was cloned to Yep-K to produce the final plasmid Yep-PMK.
Baker’s yeast transformation was achieved through lithium acetate/PEG method [18]. The deletion cassette of NA-loxP-KanMX-loxP-NB was amplified with N-S and N-X and transformed into the industrial baker’s yeast BY14a. The fragment was integrated into the chromosome at the NTH1 locus of BY14a by homologous recombination to construct the NTH1 deletion strain. The selection of NTH1 deletion strain was performed using the YEPD medium supplemented with 800 mg/L geneticin (G418). After selection, recombinant strains were verified with the primers N-S, K-S and N-X, K-X. Cre recombinase was expressed and KanMX was excised after introducing the plasmid pSH-Zeocin into the NTH1 deletion strain, thus resulting in B-NTH1. The respective transformation plasmids Yep-K, Yep-PMK were then transformed to select the G418-resistant strains BY14a + K, B-NTH1 + K, B + MAL62 and B-NTH1 + MAL62. BY14a + K and B-NTH1 + K were BY14a and B-NTH1 carrying the vector Yep-K, respectively, used as a blank control to demonstrate any possible effect of the empty vector. The transformants were then verified by PCR using the primers Kan-U and Kan-D.
Assay of the intracellular trehalose content
Fresh yeast cells were dried overnight at 85 °C to calculate the cell dry weight (CDW). Trehalose was extracted from 0.1 g of fresh yeast cells (previously washed with distilled water twice) with 4 mL of 0.5 mol/L cold trichloroacetic acid and the extract was employed for measuring the trehalose content as described previously [19, 20]. Experiments were conducted three times.
Determination of neutral trehalase activity
The activities of neutral trehalase in crude extracts were measured as described previously [21]. The liberated glucose was analyzed by HPLC employing an Aminex HPX-87H column with 5 mmol/L H2SO4 as the mobile phase at a flow rate of 0.6 mL/min at 65 °C. One unit of trehalase activity was defined as the amount of trehalase producing 1.0 μm glucose per min under assay conditions. The specific trehalase activity was expressed as the units per gram CDW. Experiments were conducted three times.
Determination of Tps1 (trehalose-6-phosphate synthase) activity
Tps1 activity was measured as described previously [22]. The trehalose-6-phosphate formed during the reaction was quantitatively determined using the Anthrone method [19]. One unit of Tps1 activity was defined as the amount of Tps1 producing 1.0 μm 6-phosphate-trehalose per min under assay conditions. The specific Tps1 activity was expressed as the units per gram CDW. Experiments were conducted three times.
Determination of alpha-glucosidase activity
Crude extracts were prepared using the Salema-Oom method to determine enzyme activities [23]. Alpha-glucosidase were determined following the Houghton-Larsen method [24]. Standard errors were less than 10 %.
Determination of the cell viability of baker’s yeast after freezing and thaw
For the freeze–thaw stress, yeast cells were harvested from the cane molasses medium and inoculated into the LSMLD medium at 30 °C for 25 min. One milliliter of cell culture was shifted to −20 °C and at 5 min intervals for different prefermentation time periods. After freezing for 1–3 week, the frozen suspensions were thawed at 30 °C for 30 min then diluted and plated on YEPD plates for 2 days. Cell viability was determined by the percentage of the number of colonies after stressing relative to the number of colonies before stress. Three independent experiments were performed.
Determination of leavening ability
The leavening ability of yeast cells was assayed by measuring the CO2 production in lean dough. Lean dough was composed of 280 g of standard flour, 150 mL of water, 4 g of salt, and 9 g of fresh yeast. The dough was evenly and rapidly stirred for 5 min at 30 ± 0.2 °C then divided into pieces (50 g each) and placed in a fermentograph box 171 (Type JM451, Sweden). CO2 production was recorded at 30 °C for 120 min. Experiments were conducted three times.
To assay the leavening ability after freeze–thaw, the mixed dough was stored at −20 °C. After freezing for 1 week, the frozen dough was thawed at 30 °C for 30 min, and the CO2 production was assayed for 120 min at 30 °C. Experiments were conducted at least thrice.
Statistical analysis
Data were expressed as mean ± SD and were accompanied by the number of experiments independently performed. Differences among all the strains were analyzed using ANOVA. P < 0.05 were considered statistically significant. The differences between the transformants and the parental strain were confirmed by Student’s t test. Differences at P < 0.05 were considered statistically significant.
Results
Overexpression of MAL62 enhances the Tps1 activity and intracellular trehalose content of baker’s yeast
Previous studies have reported that the MAL gene has a positive effect on the activity of Tps1, a trehalose-6-phosphate synthase that synthesizes trehalose under stress conditions [12]. We first tested if the Tps1 activity is affected by MAL62 overexpression. As shown in Table 3, overexpression of MAL62 (in both B + MAL62 and B-NTH1 + MAL62) significantly increased the Tps1 activity (P < 0.05). The alpha-glucosidase activities of these two strains were also increased significantly (Table 3). These results suggest that overexpression of MAL62 induces trehalose production.
To further confirm this, we measured and compared the trehalose levels in different strains. We found that all six strains (BY14a, B-NTH1, B + MAL62, B-NTH1 + MAL62, BY14a + K and B-NTH1 + K) had similar growth curves. Cells entered exponential phase 3 h after inoculation, and stationary phase 10 h after inoculation (data not shown). Our results showed in strains overexpressing MAL62 (B + MAL62 and B-NTH1 + MAL62), trehalose started to accumulate in late exponential stage at a rate of 21.9 mg/h/g CDW. In contrast, in strains having no MAL62 overexpression (BY14a, B-NTH1, BY14a + K and B-NTH1 + K), trehalose accumulation started only in stationary phase and at a lower rate (19.1 mg/h/g CDW) (Fig. 1).

Trehalose accumulation during growth of the six S. cerevisiae strains in cane molasses medium. BY14a + K and B-NTH1 + K were BY14a and B-NTH1 carrying the vector Yep-K, respectively, used as a blank control to demonstrate any possible effect of the empty vector. Data are average of three independent experiments, and error bars represent ± SD
MAL62 overexpression does not affect the rate of trehalose degradation
To examine if MAL62 overexpression or NTH1 is involved in trehalose degradation, we compared the neutral trehalase activity and the degradation rate of intracellular trehalose among the six strains. As shown in Table 3, the B + MAL62 strain had a similar neutral trehalase activity compared to its control (BY14a and BY14a + K), suggesting that overexpression of MAL62 did not affect the trehalose degradation. This is further confirmed by direct measurement of the intracellular trehalose content (Fig. 2), which showed a similar degradation rate among B + MAL62, BY14a and BY14a + K. In addition, both the neutral trehalase activity and the rate of trehalose degradation were significantly lower in all NTH1 deletion strains (B-NTH1, B-NTH1 + K and B-NTH1 + MAL62) (Table 3, Fig. 2), regardless whether MAL62 was overexpressed or not. These results suggest that NTH1, but not MAL62, is important for trehalose degradation.

Content of intracellular trehalose during cultivation in LSMLD (prefermentation). Data are averages of three independent experiments, and error bars represent ± SD
High trehalose content increases viability of yeast cells after freezing
Although a number of reports have shown that the degradation of trehalose during prefermentation is necessary [25], the residual intracellular trehalose is still considered to be important to freezing tolerance of yeast [26, 27]. Hereby, we assessed the cell viability of the six strains to investigate the effect of MAL62 overexpression and/or NTH1 deletion on the freezing tolerance of yeasts after prefermentation and 7 d freezing.
As shown in Fig. 3, the cell viability of strains with MAL62 overexpression (B + MAL62 and B-NTH1 + MAL62) was significantly higher than the other strains before prepermentation (time = 0 min). Cell viability of all strains decreased as prefermentation time increased. 25 min after prefermentation, the cell viability of the strain with both MAL62 overexpression and NTH1 deletion (B-NTH1 + MAL62) was significantly higher than other strains (ANOVA, P < 0.05). The cell viability of strains with either MAL62 overexpression or NTH1 deletion remained in the middle, while the control strains (BY14a and BY14a + K) had the lowest viability, dropping from about 80 % to about 40 %. The cell viability is in agreement with the trehalose content (Fig. 2), which showed that 25 min after prefermentation, the B-NTH1 + MAL62 had the highest trehalose content (95 mg/g CDW) and the controls (BY14a and BY14a + K) had the lowest (about 55 mg/g CDW). These results suggest that the residual trehalose content has a positive correlation with the viability of yeast cells after prefermentation and freezing [28].

Cell viability of strains after prefermentation for different time periods in LSMLD and frozen for 7 d at −20 °C. Data are average of three independent experiments, and error bars represent ± SD
Overexpression of MAL62 or deletion of NTH1 confers long-term freezing tolerance of baker’s yeast
In order to access the long-term freezing tolerance of the NTH1-deletion and/or the MAL62-overexpression strains, we examined the trehalose content before freezing and the cell viability 21d after freezing (Fig. 4). As shown in Fig. 4, both the trehalose content and the cell viability were significantly higher in strains with MAL62 overexpression (B + MAL62 and B-NTH1 + MAL62) (ANOVA, P < 0.05). Compared with the control (BY14a and BY14a + K), deletion of NTH1 alone (B-NTH1 and B-NTH1 + K) also induced a higher trehalose content and higher cell viability, which is in agreement with previous studies [26, 29].

Intracellular trehalose content (before frozen) and cell viability (after frozen) of strains before or after 21d frozen. Data are average of three independent experiments, and error bars represent ± SD
Overexpression of MAL62 and deletion of NTH1 enhance the fermentation characteristics of baker’s yeast exposed to freezing-thaw stress
Leavening ability is an important fermentation characteristic of baker’s yeast used in frozen dough. We next explored the possible effects of MAL62 overexpression and NTH1 deletion on the leavening ability after freezing and thaw by measuring the CO2 production. Our results showed that freezing-thaw caused a reduction of CO2 production in all strains (comparing Fig. 5a with 5b). However, either before or after freezing-thaw, overexpression of MAL62 (B + MAL62 and B-NTH1 + MAL62) significantly enhanced the CO2 production (ANOVA, P < 0.05). NTH1 deletion alone had no effect on CO2 production before freezing-thaw (Fig. 5a) but enhanced the CO2 production after freezing-thaw (Fig. 5b). Interesting, MAL62 overexpression and NTH1 deletion (B-NTH1 + MAL62) had a lower CO2 production than MAL62 overexpression alone (B + MAL62) before freezing-thaw, but the CO2 production was higher after the freezing-thaw, suggesting that MAL62 overexpression and NTH1 deletion provide the best enhancement on leavening ability upon freezing-thaw stress.

CO2 production of yeast in lean dough before and after freeze–thaw. CO2 production was measured before (a) and after (b) freeze–thaw stress. Data are average of three independent experiments, and error bars represent ± SD
Discussion
Biological macromolecules and membranes are liable to denaturation under freezing conditions [30], Freezing also causes the formation of intracellular ice crystals, which are harmful to cells. It has been suggested that trehalose could act as a stabilizer of cellular membranes and proteins under freezing stress [28]. Previous studies have reported that the modification of the whole MAL gene cluster is necessary to elicit trehalose synthesis [31, 32]. In this study, we demonstrated that the single-gene-overexpression of MAL62 in industrial baker’s yeast is capable of increasing trehalose accumulation and cell viability under freezing stress. Trehalose formation in MAL62 overexpressing strains (B + MAL62 and B-NTH1 + MAL62) was earlier and faster than the controls (Fig. 1), suggesting the positive effects on the intracellular trehalose content and freezing tolerance (Figs. 2, 4). Moreover, although MAL62 overexpression had little effect on protecting trehalose against degradation during prefermentation, the cell viability assay showed that the MAL62 overexpression could protect cells against freezing stress after prefermentation. This is in line with a previous report [33], showing that the trehalose accumulation before the induction of stress was more important for stress tolerance.
One explanation is that MAL62 overexpression enhances the activity of Tps1. This hypothesis relies on the fact that maltose constitutive genes could partially relieve Tps1 from the catabolite repression [34], and the alpha-glucosidase (coded by gene MAL62) is the rate-limiting factor in maltose metabolism [35]. Our result is consistent with this hypothesis, since Tps1 activity could increase when the alpha-glucosidase activity was enhanced by MAL62 overexpression (Table 3). Another explanation is that the existence of adenosine-diphosphoglucose (ADPG)-dependent trehalose synthase, which requires ADPG instead of UDPG as donor of glucose units for trehalose synthesis [36]. Since the expression of ADPG-pyrophosphorylase gene and MAL genes shared the common regulation, any of the MAL gene products either by means of control at the transcription level, or by acting directly on enzyme activity could regulate the activity of the ADPG-pyrophosphorylase activity [11]. Hence, overexpression of MAL62 alone could increase intracellular trehalose content and bring about further enhancements in freezing tolerance.
The fermentation characteristics of baker’s yeast as a strong correlation with the tolerance in stress conditions [37]. After exposure to freeze–thaw stress, response to the environmental change involved in rapid accumulation of relevant protectants and rapid production of enzymes related to stress-protective effect [7, 19]. In this work, we found that the freezing tolerance and the fermentation characteristics of the double mutant (B-NTH1 + MAL62) were significantly enhanced than that of either single mutant (B-NTH1 or B + MAL62) after the freezing-thaw stress (Figs. 4, 5b). In addition, we found that NHT1 deletion (B-NTH1, B-NTH1 + K and B-NTH1 + MAL62) induced a low neutral trehalase (Table 3), which caused a lower level of trehalose degradation. High activity of trehalose synthase (+MAL62) [38] and low activity of neutral trehalase (−NTH1) increase the intracellular trehalose level [26], which explains why the double mutant (B-NTH1 + MAL62) provides the best freezing tolerance and fermentation characteristics [39].
In summary, our study showed that MAL62 overexpression and NTH1 deletion in baker’s yeast significantly enhanced the freezing tolerance and fermentation characteristics, which is in high demand in the frozen dough baking industry [40]. On the basis of our findings, it is also possible to lengthen the storage period of frozen dough through overexpression of the MAL62 with NTH1 deletion. It provides valuable insights for breeding novel stress-tolerant and fast-fermented baker’s yeast strains that are useful for baking industry.
Conclusion
The results of this study show that overexpression of MAL62 was an effective way of increasing trehalose content and cell viability after prefermention-freezing and long-term frozen. Deletion of NTH1 in combination of MAL62 overexpression could further strengthen freezing tolerance and improve the leavening ability after freezing-thaw stress. Furthermore, the single-gene-overexpression of MAL62 in industrial baker’s yeast is capable of increasing trehalose accumulation, therefore, promoting cell viability and the leavening ability of baker’s yeast in lean dough under freezing stress. Hence, such baker’s yeast has excellent commercial and industrial applications.
References
Sasano Y, Haitani Y, Hashida K, Ohtsu I, Shima J, Takagi H. Simultaneous accumulation of proline and trehalose in industrial baker’s yeast enhances fermentation ability in frozen dough. J Biosci Bioeng. 2012;113:592–5.
Tanaka F, Ando A, Nakamura T, Takagi H, Shima J. Functional genomic analysis of commercial baker’s yeast during initial stages of model dough-fermentation. Food Microbiol. 2006;23:717–28.
Elbein AD, Pan Y, Pastuszak I, Carroll D. New insights on trehalose: a multifunctional molecule. Glycobiology. 2003;13:17R–27R.
Crowe JH, Hoekstra FA, Crowe LM. Anhydrobiosis. Annu Rev Physiol. 1992;54:579–99.
Singer MA, Lindquist S. Multiple effects of trehalose on protein folding in vitro and in vivo. Mol Cell. 1998;1:639–48.
Blomberg A. Metabolic surprises in Saccharomyces cerevisiae during adaptation to saline conditions: questions, some answers and a model. FEMS Microbiol Lett. 2000;182:1–8.
Attfield PV. Stress tolerance: the key to effective strains of industrial baker’s yeast. Nat Biotechnol. 1997;15:1351–7.
De Virgilio C, Burckert N, Bell W, Jeno P, Boller T, Wiemken A. Disruption of TPS2, the gene encoding the 100-kDa subunit of the trehalose-6-phosphate synthase/phosphatase complex in Saccharomyces cerevisiae, causes accumulation of trehalose-6-phosphate and loss of trehalose-6-phosphate phosphatase activity. Eur J Biochem/FEBS. 1993;212:315–23.
Vuorio OE, Kalkkinen N, Londesborough J. Cloning of two related genes encoding the 56-kDa and 123-kDa subunits of trehalose synthase from the yeast Saccharomyces cerevisiae. Eur J Biochem/FEBS. 1993;216:849–61.
Paschoalin VM, Costa-Carvalho VL, Panek AD. Further evidence for the alternative pathway of trehalose synthesis linked to maltose utilization in Saccharomyces. Curr Genet. 1986;10:725–31.
Ferreira JC, Thevelein JM, Hohmann S, Paschoalin VM, Trugo LC, Panek AD. Trehalose accumulation in mutants of Saccharomyces cerevisiae deleted in the UDPG-dependent trehalose synthase-phosphatase complex. Biochim Biophys Acta. 1997;1335:40–50.
Petit T, Francois J. Accumulation of trehalose in Saccharomyces cerevisiae growing on maltose is dependent on the TPS1 gene encoding the UDPglucose-linked trehalose synthase. FEBS Lett. 1994;355:309–13.
Garre E, Pérez-Torrado R, Gimeno-Alcañiz JV, Matallana E. Acid trehalase is involved in intracellular trehalose mobilization during postdiauxic growth and severe saline stress in Saccharomyces cerevisiae. FEMS Yeast Res. 2009;9:52–62.
Parrou JL, Teste MA, Francois J. Effects of various types of stress on the metabolism of reserve carbohydrates in Saccharomyces cerevisiae: genetic evidence for a stress-induced recycling of glycogen and trehalose. Microbiology. 1997;143(6):1891–900.
Shima J, Takagi H. Stress-tolerance of baker’s-yeast (Saccharomyces cerevisiae) cells: stress-protective molecules and genes involved in stress tolerance. Biotechnol Appl Biochem. 2009;53:155–64.
Ando A, Nakamura T, Murata Y, Takagi H, Shima J. Identification and classification of genes required for tolerance to freeze-thaw stress revealed by genome-wide screening of Saccharomyces cerevisiae deletion strains. FEMS Yeast Res. 2007;7:244–53.
Panadero J, Randez-Gil F, Prieto JA. Validation of a flour-free model dough system for throughput studies of baker’s yeast. Appl Environ Microbiol. 2005;71:1142–7.
Gietz RD, Woods RA. Transformation of yeast by lithium acetate/single-stranded carrier DNA/polyethylene glycol method. Meth Enzymol. 2002;350:87–96.
Lewis JG, Learmonth RP, Watson K. Role of growth phase and ethanol in freeze-thaw stress resistance of Saccharomyces cerevisiae. Appl Environ Microbiol. 1993;59:1065–71.
Trevelyan WE, Harrison JS. Studies on yeast metabolism. 5. The trehalose content of baker’s yeast during anaerobic fermentation. Biochem J. 1956;62:177–83.
Jules M, Guillou V, Francois J, Parrou JL. Two distinct pathways for trehalose assimilation in the yeast Saccharomyces cerevisiae. Appl Environ Microbiol. 2004;70:2771–8.
Hottiger T, Schmutz P, Wiemken A. Heat-induced accumulation and futile cycling of trehalose in Saccharomyces cerevisiae. J Bacteriol. 1987;169:5518–22.
Salema-Oom M, de Sousa HR, Assuncao M, Goncalves P, Spencer-Martins I. Derepression of a baker’s yeast strain for maltose utilization is associated with severe deregulation of HXT gene expression. J Appl Microbiol. 2011;110:364–74.
Houghton-Larsen J, Brandt A. Fermentation of high concentrations of maltose by Saccharomyces cerevisiae is limited by the COMPASS methylation complex. Appl Environ Microbiol. 2006;72:7176–82.
Sebollela A, Louzada PR, Sola-Penna M, Sarone-Williams V, Coelho-Sampaio T, Ferreira ST. Inhibition of yeast glutathione reductase by trehalose: possible implications in yeast survival and recovery from stress. Int J Biochem Cell Biol. 2004;36:900–8.
Shima J, Hino A, Yamada-Iyo C, Suzuki Y, Nakajima R, Watanabe H, et al. Stress tolerance in doughs of Saccharomyces cerevisiae trehalase mutants derived from commercial baker’s yeast. Appl Environ Microbiol. 1999;65:2841–6.
Izawa S, Sato M, Yokoigawa K, Inoue Y. Intracellular glycerol influences resistance to freeze stress in Saccharomyces cerevisiae: analysis of a quadruple mutant in glycerol dehydrogenase genes and glycerol-enriched cells. Appl Microbiol Biotechnol. 2004;66:108–14.
Tan H, Dong J, Wang G, Xu H, Zhang C, Xiao D. Enhanced freeze tolerance of baker’s yeast by overexpressed trehalose-6-phosphate synthase gene (TPS1) and deleted trehalase genes in frozen dough. J Ind Microbiol Biotechnol. 2014;41:1275–85.
Cui-ying Z, Dong-guang X, Ye L. Influence of trehalose accumulation on response to freeze stress in baker’s yeast. 4th International Conference on Bioinformatics and Biomedical Engineering (iCBBE 2010). 2010.
Griffiths JB. Effect of hypertonic stress on mammalian cell lines and its relevance to freeze-thaw injury. Cryobiology. 1978;15:517–29.
Plourde-Owobi L, Durner S, Goma G, Francois J. Trehalose reserve in Saccharomyces cerevisiae: phenomenon of transport, accumulation and role in cell viability. Int J Food Microbiol. 2000;55:33–40.
Peixoto DN, Panek AD. The involvement of hexokinases in trehalose synthesis. Biochem Mol Biol Int. 1999;47:873–80.
Mahmud SA, Hirasawa T, Shimizu H. Differential importance of trehalose accumulation in Saccharomyces cerevisiae in response to various environmental stresses. J Biosci Bioeng. 2010;109:262–6.
Kopp M, Muller H, Holzer H. Molecular analysis of the neutral trehalase gene from Saccharomyces cerevisiae. J Biol Chem. 1993;268:4766–74.
Zhang C-Y, Lin X, Song H-Y, Xiao D-G. Effects of MAL61 and MAL62 overexpression on maltose fermentation of baker’s yeast in lean dough. World J Microbiol Biotechnol. 2015;31:1241–9.
Paschoalin VM, Silva JT, Panek AD. Identification of an ADPG-dependent trehalose synthase in Saccharomyces. Curr Genet. 1989;16:81–7.
Sasano Y, Haitani Y, Ohtsu I, Shima J, Takagi H. Proline accumulation in baker’s yeast enhances high-sucrose stress tolerance and fermentation ability in sweet dough. Int J Food Microbiol. 2012;152:40–3.
Ming-Zhe A, Yue-Qin T, Mitsumasu K, Ze-Shen L, Shigeru M, Kenji K. Enhanced thermotolerance for ethanol fermentation of Saccharomyces cerevisiae strain by overexpression of the gene coding for trehalose-6-phosphate synthase. Biotechnol Lett. 2011;33:1367–74.
Perez-Torrado R, Matallana E. Enhanced fermentative capacity of yeasts engineered in storage carbohydrate metabolism. Biotechnol Prog. 2015;31:20–4.
Sasano Y, Haitani Y, Hashida K, Oshiro S, Shima J, Takagi H. Improvement of fermentation ability under baking-associated stress conditions by altering the POG1 gene expression in baker’s yeast. Int J Food Microbiol. 2013;165:241–5.
Gueldener U, Heinisch J, Koehler GJ, Voss D, Hegemann JH. A second set of loxP marker cassettes for Cre-mediated multiple gene knockouts in budding yeast. Nucl Acid Res. 2002;30:e23.
Lilly M, Lambrechts MG, Pretorius IS. Effect of increased yeast alcohol acetyltransferase activity on flavor profiles of wine and distillates. Appl Environ Microbiol. 2000;66:744–53.
Authors’ contributions
XS carried out the experiments and drafted the manuscript. MYW and SNL participated in the plasmid and strain construction. ZHF and WBZ assisted in fermentation experiments. CYZ and DGX conceived the study and reviewed the final manuscript. All authors read and approved the final manuscript.
Acknowledgements
The current study was financially supported by the National Natural Science Foundation of China (31571809) and the Research Fund for the Doctoral Program of Tianjin Agricultural University (2013N04).
Competing interests
The authors declare that they have no competing interests.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Sun, X., Zhang, CY., Wu, MY. et al. MAL62 overexpression and NTH1 deletion enhance the freezing tolerance and fermentation capacity of the baker’s yeast in lean dough. Microb Cell Fact 15, 54 (2016). https://doi.org/10.1186/s12934-016-0453-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12934-016-0453-3