
Abstract
Inflammation and immune processes underlie pulmonary hypertension progression. Two main different activated phenotypes of macrophages, classically activated M1 macrophages and alternatively activated M2 macrophages, are both involved in inflammatory processes related to pulmonary hypertension. Recent advances suggest that macrophages coordinate interactions among different proinflammatory and anti-inflammatory mediators, and other cellular components such as smooth muscle cells and fibroblasts. In this review, we summarize the current literature on the role of macrophages in the pathogenesis of pulmonary hypertension, including the origin of pulmonary macrophages and their response to triggers of pulmonary hypertension. We then discuss the interactions among macrophages, cytokines, and vascular adventitial fibroblasts in pulmonary hypertension, as well as the potential therapeutic benefits of macrophages in this disease. Identifying the critical role of macrophages in pulmonary hypertension will contribute to a comprehensive understanding of this pathophysiological abnormality, and may provide new perspectives for pulmonary hypertension management.
Similar content being viewed by others

Introduction
Pulmonary hypertension (PH) is a pathophysiological abnormality characterized by a progressive increase in mean pulmonary arterial pressure (mPAP) over 20 mmHg [1]. PH has been classified into five clinical groups based on clinical and hemodynamic parameters, namely pulmonary arterial hypertension (group 1), PH associated with left heart disease (group 2), PH associated with lung diseases or hypoxemia (group 3), PH caused by chronic thrombotic or embolic disease (group 4), and miscellaneous PH (group 5) [1]. PH is regarded as a major threat to global health issues due to its high prevalence and poor prognosis and is estimated to affect 1% of the global population [2]. The etiologic processes of PH vary from congenital heart diseases, rheumatic heart diseases and infections, which are common in less-developed areas, to respiratory diseases and left heart disorders, which usually affect patients in developed areas [2]. The primary goal of PH treatment is to delay or even reverse this pathophysiological disorder that causes progressive deterioration of the lungs and other target organs, leading to unpredictable and refractory outcomes [1]. Hence, the marked etiological heterogeneity of PH demands precise and targeted medical intervention, which highlights the need for a comprehensive understanding of the pathogenesis of PH.
Pulmonary vascular remodelling is characterized as an important histopathological alteration in PH. Abnormal vascular remodelling of precapillary arterioles, occlusive intimal lesions, and concentric arterial wall thickening have been observed in PH patients. These histopathological alterations are associated with increased pulmonary vascular resistance [3]. A variety of factors, such as genetic context, vascular endothelial cells (ECs), vascular smooth muscle cells (VSMCs), fibroblasts, platelets and inflammatory cells, contribute to the pathogenesis and progression of PH, suggesting the involvement of the whole vessel wall [4]. Recently, alterations in soluble mediator levels as well as perivascular infiltration of immune cells, such as monocytes/macrophages, neutrophils and lymphocytes in established PH have been demonstrated [3, 5,6,7]. Lekva et al. reported that an elevated level of sCD163, a monocyte/macrophage biomarker, was associated with worse outcomes in PH patients [8]. These results implicate that inflammation plays an important role in PH progression. The infiltration of macrophages occurs mainly in perivascular areas and is involved in the pathogenesis of PH by coordinating the initiation and resolution of pulmonary inflammation [6]. Since macrophages play a critical role in pulmonary vascular remodelling, this review aimed to provide new perspectives for further understanding the pathogenesis and therapeutic strategies for this catastrophic condition. Group 2, Group 3, and Group 4 PH are often included and discussed in their primary conditions, such as left heart dysfunction or chronic lung diseases. Therefore, we focused mainly on Group 1 (pulmonary arterial hypertension) in this review.
Origin of macrophages in the lung during PH
There are two types of tissue-resident macrophages in the lung, alveolar macrophages (AMs) and interstitial macrophages (IMs). AMs originate from embryonic liver mononuclear cells, reside on alveoli and airways and maintain immune homeostasis [9]. IMs originate from circulating monocytes and participate in adaptive immune responses via interactions with interstitial T lymphocytes [10]. Mononuclear phagocytes are recruited to the lung during infection and then shift toward different subtypes [11]. It is currently accepted that macrophages can be classified into two main subgroups: the classically activated type (M1), which promotes the inflammatory response via the synthesis and release of proinflammatory cytokines; or the alternatively activated type (M2), which promotes inflammation resolution and tissue repair [12]. Montani et al. reported the involvement of bone marrow-derived CD117 + cells in human idiopathic PH [13]. Circulating monocytes are recruited to the lungs when chemokine levels increase, where they replace resident interstitial macrophages and subsequently participate in vascular remodelling [14]. Therefore, monocytes are recruited and then react to microenvironmental changes in the lungs during external or internal stimuli, and function as key cellular mediators to coordinate inflammatory reactions, differentiation and polarization into different active subtypes [15].
Macrophages infiltrate vessels during PH, contributing to vascular remodelling
The remodelling of pulmonary arterial vessels is essential for PH progression, and leads to a progressive increase in mPAP [3]. Arterial remodelling requires the accumulation of resident pulmonary vessel cells and inflammatory cells, which suggests the involvement of all three layers. Specific pathological features, such as fibrosis alterations, can be detected via histopathological assessment at the early stage of PH progression [3]. Both external and internal stimuli, such as genetic mutations, hypoxia, cold exposure, air pollution, and respiratory infection, can initiate immune responses and therefore lead to the proliferation of vascular cells, autoantibody formation, and dysregulated immunity [16,17,18,19,20]. Histopathological evidence has suggested that perivascular infiltration of inflammatory cells is common in PH and precedes structural remodelling in vessels [5, 6, 17, 21, 22]. In idiopathic PH, the level of immune infiltration is higher for CD8 + T cells, resting memory CD4 + T cells, γδ T cells, M1/M2 macrophages and resting mast cells; and lower for monocytes, neutrophils and naïve CD4 + T cells [23,24,25]. These results suggest that immune imbalance contributes to remodelling in PH.
Hypoxia-induced (sugen, a vascular endothelial growth factor antagonist, is used simultaneously in many studies) and monocrotaline (MCT)-induced experimental PH models, which are similar to group one pulmonary arterial hypertension, have been widely adopted [26, 27]. Compared with human PH samples, experimental PH models provide richer information about macrophage infiltration. Monocyte numbers increase in both the bone marrow and the blood when hypoxic exposure is prolonged [28]. After one day of exposure to hypoxia, the IL-1β and oncostatin M regulation-related subtypes of macrophages are the most abundant groups in the lungs of mice, which are often associated with proinflammatory effects. Later, new subtypes that are overrepresented with mitochondrial dysfunction, oxidative phosphorylation and the EIF2 signalling pathway emerge after seven days of hypoxic exposure; these subtypes are involved in anti-inflammatory effects and tissue repair [29]. As Pugliese et al. reported previously, during the early period of hypoxic exposure, macrophages accumulate around pulmonary vessels, exhibit a hypoxic response and release proinflammatory cytokines; subsequently, the perivascular accumulation of macrophages decreases and demonstrates the tissue repair and anti-inflammatory programming states [30]. Inhibiting the early accumulation of monocytes and macrophages has been reported to effectively ameliorate the right ventricular burden and Fulton index in hypoxic mouse models [31]. The genetic/pharmacological intervention of several key signalling pathways in macrophages can protect against experimental PH progression [32,33,34,35,36,37,38,39,40,41]. These potential targets function via different mechanisms, such as the alleviation of macrophage-related pulmonary inflammation and the inhibition of macrophage activation and polarization. Additionally, the depletion of AMs reportedly attenuates hypoxia-induced PH in rats [42]. These genetic or pharmacological targets are summarized in Table 1.
Recently, unexpected macrophage infiltration has been detected in the right ventricle in MCT- or hypoxia-induced rat models, leading to inflammation in the right heart [43, 44]. Therefore, the inflammatory response might not only affect the lungs and pulmonary vessels, the heart can also suffer from similar pathophysiological alterations. The similar macrophage infiltration in the heart is important too, because right heart dysfunction and failure are major risk factors for poor prognosis in PH patients. Interestingly, the depletion of macrophages reportedly induces experimental PH in male mice but not in female mice [45], suggesting a potential association between sex and immune cells in PH. This is quite interesting, since PH is more likely to be a female predominant disease with a female-to-male ratio of approximately 4:1, while PH tends to be more severe in males [46]. Oestrogen is believed to mediate the protective effect of female sex in PH via oestrogen receptor-β, based on evidence from MCT-induced rat PH models treated with oestrogen [47]. The genetic variation in oestrogen metabolites and the canonical/noncanonical TGF-β signalling pathways regulated by sex hormones might explain the female predominance in PH [48, 49]. For example, oestrogen inhibition induced by aromatase inhibitors can reduce the development of PH in bmpr2-mutant mice [49]. Oestrogen can regulate the expression of TGF-β receptors and signalling modulators, such as endoglin and TGF-β3, at the transcriptional and protein levels [48]. Other sex hormones, including progestogen, androgen and anti-Müllerian hormone might also contribute to the sex-related differences in PH. Therefore, oestrogen metabolism and oestrogen receptors are regarded as promising therapeutic targets in PH [50]. Additionally, elevated aromatase expression is elevated in the special phenotype of female pulmonary SMCs, suggesting a higher level of estradiol synthesis and thus contributing to female PH susceptibility [51]. Regulatory T-cell function has been demonstrated to be a protective factor in female rodent models of PH, that Treg deficiency results in more severe PH in females than in males [52]. The Y chromosome was reported to be protective against the development of hypoxia-induced PH [53]. However, oestradiol supplementation can also limit several maladaptive processes in the right ventricles associated with PH, such as pro-apoptotic signalling, oxidative stress and activation of pro-inflammatory cytokines, which might prevent right ventricle remodelling [49]. Until now, no sex difference was found in macrophage polarization [45], and there is no direct evidence that macrophages act differently in different sexes.
Macrophages react to PH triggers and polarize to different subgroups
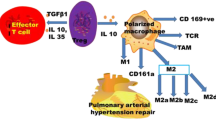
As discussed previously, macrophages infiltrate the perivascular area during PH. Monocyte-derived macrophages are recruited to the lung and then differentiate in response to microenvironmental changes during pulmonary damage; these cells are involved in lung injury and repair [15, 54]. The time- and compartment-specific activation of lung macrophages was reported by Pugliese et al. [30]. During the early stage of hypoxic exposure, macrophages accumulate vastly around pulmonary arterioles instead of the alveolar space, with upregulation of mTORC1 signalling, glycolysis, and oxidative phosphorylation in both AMs and IMs. The expression of IL-1, IL-5, CCL-5, and EGFR is also increased in both AMs and IMs during the early stage of hypoxic exposure. When hypoxia is prolonged, the perivascular accumulation of macrophages decreases and presents different compartment-specific activation, that perivascular IMs tend to signal against inflammation and promote tissue repair/remodelling while AMs still exhibit high proinflammatory-related signalling [30]. A typical change in macrophage subgroups in the experimental PH model is that M1-polarized macrophages predominate in the early phase after stimulation; then, M2-polarized macrophages accumulate in the lung with an increase in the M2/M1 ratio to almost ten after four weeks of induction [28, 55, 56]. M1 macrophages are regarded as proinflammatory components, while M2 macrophages are often associated with anti-inflammatory and reparative effects [45]. However, interruption of the M1 subgroup can also effectively protect against PH [38]. Additionally, a rebalanced M2/M1 ratio could be observed in alleviated PH models [57]. Changes in macrophage activation and the M1/M2 ratio are presented in Fig. 1.

PH triggers induce macrophage activation and polarization. During the early stage, recruited and resident macrophages polarize to the M1 type in response to external stimuli. Then M2 type dominates the main polarization type of macrophages in the lung for tissue repair, as stimuli are constantly present. Macrophage activation and polarization are complex and consecutive processes, that cannot be simply regarded as separate and contradictory processes involved in PH progression
The activation of macrophages in the hypoxia-induced experimental PH model has been reported to be related to alterations in several signalling pathways and cytokines, such as hypoxia-inducible factor 1 (HIF-1), IL-6, NF-κB, HIMF and IL-6 [58]. During the early phase of hypoxia, the increased expression of Ythdf2 in AMs promotes the degradation of m6A-modified Hmox1 mRNA and the activation of macrophages; consequently, levels of anti-inflammatory mediators (such as IL-10) are evaluated and promote the proliferation of VSMCs, leading to PH progression [59]. The deletion of Ythdf2 in bone marrow-derived macrophages (BMDMs) leads to the promotion of the M1 subtype by enhancing MAPK and NF-κB signalling and the inhibition of the M2 subtype by upregulating p53 expression [60]. This result provides another potential explanation for why the absence of Ythdf2 in macrophages protects PH. IMs express thrombospondin-1 after hypoxic exposure, thus promoting hypoxic PH progression via TGF-β activation [61].
Human RELM-β, which is homologous to the rodent RELM-α (also known as HIMF or FIZZ1) and is regarded as a marker of the M2 subtype, has been reported to be upregulated in the proliferative stage in hypoxia-induced PH [62]. HIMF-positive signals are mainly colocalized with perivascular macrophages [63]. When exposed to chronic hypoxia, RELM-α emerges in macrophages and subsequently promotes the activation and release of HMGB1, while the DAMP receptor RAGE is required for HMGB1 function and maintenance of inflammation. Hence, an autocrine positive feedback loop is established in macrophages, resulting in continuous vascular inflammation and proliferation of VSMCs in PH [64]. The hypoxia-induced increase in RELM-β could also inhibit membrane KCNK3 expression via PLC activity and endocytosis [65], which have been reported to contribute to local inflammatory response, vascular remodelling and proliferation [66]. The IL-4 signalling pathway can synergistically enhance the HIMF-induced expression of vascular endothelial growth factor (VEGF) and monocyte chemoattractant protein-1 (MCP-1), which is also associated with angiogenesis in pulmonary microvessels [67, 68].
Hypoxia-inducible factor (HIF) is a major regulator of oxygen homeostasis that initiates a series of hypoxic responses, including changes in vascular tone and proliferation, cellular metabolism, autophagy and cellular survival [34]. HIMF induces the recruitment of macrophages to the angiogenesis area in the HIF-1-dependent manner and the production of IL-1 by perivascular macrophages and VSMCs in the HIF-1α-dependent manner [62]. Deletion of HIF-1α in BMDMs reduces right heart remodelling and pulmonary macrophage infiltration in hypoxia-induced PH models, suggesting the involvement of HIF-1 in PH progression [33]. It has also been reported that decreased perivascular macrophage infiltration in hypoxia-induced PH models was associated with the alleviation of pulmonary vascular muscularization [33]. Leptin, a HIF-dependent peptide hormone, is reportedly upregulated in pulmonary vessels, leading to abnormal monocyte/macrophage activation, perivascular macrophage accumulation, and IL-6 overexpression [69, 70]. Hence, interfering with macrophage migration seems to be a promising target for PH treatment. A previous report indicated that HIF-1α is expressed at higher levels in M1 macrophages, and HIF-2α is expressed at higher levels in M2 macrophages [71]. The suppression of HIF-2α can alleviate hypoxia-induced PH [35]. Active arginase can reduce the supply of L-arginine, which is needed for NO synthase [72], thus decreasing NO production in macrophages. Arginase 1 expression in macrophages is induced by HIF-2α [73]. The protective effect of HIF-2α suppression in PH might be related to M2 macrophage dysfunction, because NO contributes to vascular resistance regulation and M2 polarization contributes to vascular remodelling.
In the MCT-induced rat model, Bruton’s tyrosine kinase (BTK), a nonreceptor tyrosine kinase of the Tec family that is linked to B-cell proliferation and survival, is upregulated in the lung and colocalized with CD68 + macrophages [36]. BTK inhibitors can prevent M1 polarization and PH progression in experimental PH models [36]. In addition, deficiency of legumain (also known as asparaginyl endopeptidase) in macrophages has been reported to attenuate hypoxia-induced PH in mouse models [74].
Activation and metabolic reprogramming in macrophages
Leukotriene B4 (LTB4) is released by activated macrophages and can induce pulmonary arterial endothelial apoptosis and extensive vascular injury [32]. LTB4 can also promote the proliferation, migration, and differentiation of fibroblasts through both the activation of p38 MAPK signalling and the upregulation of Nox4 [75]. Nox4 and reactive oxygen species (ROS) are detected in the adventitia and overlap with fibroblast markers (fibroblast activating proteins) and a monocyte marker (CD11b) [37]. Nox4-derived ROS activate the transient receptor potential melastatin 2 and therefore enhance the proliferation and migration of VSMCs in pulmonary arteries [76]. M1 macrophage infiltration in the right ventricle with enhanced NOD-like receptor thermal protein domain associated protein 3 (NLRP3) expression and activation results in right heart dysfunction [43]. Stimulators of interferon genes in macrophages induce PH progression via the activation of NLRP3 signalling transduction [77]. Moreover, prostaglandin D2 released by macrophages inhibits smooth muscle cell proliferation and induces vasodilation [78]. Under particular conditions, extracellular vesicles that are released by macrophages in certain infections carry high levels of TGF-β1, which is associated with increased pulmonary arterial systolic pressure [79].
When exposed to inflammatory stimuli or chronic hypoxia, macrophages are activated and then undergo aerobic glycolysis [80], which in turn governs macrophage function [81]. Metabolic reprogramming occurs in various cells in PH, including VECs, VSMCs, fibroblasts and immune cells, and plays a synergistic role with other PH hallmarks such as proliferation, apoptosis resistance and inflammation [82]. In macrophages, this metabolic shift is usually reversible and might involve the synthesis and secretion of immune mediators [83]. The metabolic shifts are mainly induced by HIF families and prolong survival under stress [84]. For example, HIF-1α is induced in M1-macrophages, while glycolytic and pentose phosphate pathways are also enhanced; in M2-macrophages, HIF-2α is elevated, while fatty acid oxidation level and mitochondrial respiratory chain activity are also increased [85]. These metabolic shifts are important for vascular diseases [86]. Metabolite alterations and the mitochondrial electron transport chain distribution characterize reprogrammed macrophages [87]. Chronic hypoxia might lead to a prolonged glycolytic shift even when the stimulus has been removed [84]. Due to the angiogenesis and chronically inflammatory nature of PH, several studies have attempted to alleviate PH by interfering with metabolic reprogramming. G6PD activity inhibitors decreased the accumulation of macrophages in hypoxic PH mice [28]. The specific deficiency of Pfkfb3 in myeloid cells, that Pfkfb3 is a critical enzyme of macrophage glycolysis, protects mice from PH and decreases the levels of growth factors and proinflammatory cytokines in experimental PH models [88]. When exposed to IL-4 and hypoxia, the level of insulin receptor substrate 2, a critical regulator of cellular energy homeostasis, is decreased in macrophages and subsequently contributes to chronic inflammation and vascular dysfunction [89]. SIRT1-mTOR/HIF-1α signalling can promote M2-type differentiation by blocking glycolysis and can reduce the recruitment of inflammatory cells [90, 91]. Macrophages reprogrammed by HIF-2α can produce several specific cytokines, such as IL-6, thus protecting organs from injury [92]. The activation of mTOR and HIF-1α can be induced by many factors, such as infection and the circadian clock [93, 94]. However, proinflammatory cytokine production within macrophages might not be glycolytic reprogramming-dependent [95]. In Table 2; Fig. 2, we summarize the published studies on experimental PH that focused on macrophages. Although metabolic reprogramming is an important characteristic of activated macrophages, it can occur in many other cellular components, such as fibroblasts and vascular endothelial cells, during PH progression.

Metabolic reprogramming in macrophages exposed to hypoxia and inflammation. Both HIF-1α and HIF-2α were increased by hypoxia, leading to different responses in macrophages. Glycolysis usually characterizes M1 macrophages and suppresses M2 polarization, while HIF-2α promotes cell survival and the expression of specific cytokines, such as IL-6. Circulating cytokines such as IL-4, and SIRT1 can also affect metabolic reprogramming in macrophages by activating or inhibiting mTOR signalling
Interaction of macrophages with soluble mediators
The expression of many immune mediators is altered in PH, including cytokines (such as IL-1β, IL-6, IL-10, and IL-18), chemokines and their receptors (such as CCL2 -CCR2, CCL5-CCR5, CXCL12, CX3CL1, LTB4), HIMF, and complement [7, 104]. Exosomes from mesenchymal stem cells can also regulate pulmonary inflammation via modulation of macrophage function [15, 57]. These soluble components are involved in both the inflammatory response and tissue repair, as well as interactions between different cells. Several proinflammatory or anti-inflammatory biomarkers have been applied to predict the outcome of PH clinically or experimentally [105]. In Table 3, we summarize the origins of these mediators and the cell types affected. Most mediators are produced by both macrophages and other cellular components (such as fibroblasts or smooth muscle cells) during PH progression. The levels of several chemokines, such as CCL2 and CCL5, are reportedly increased in proinflammatory fibroblasts and can induce macrophage transmigration. Together, these mediators, which have various origins, regulate PH progression.
IL-6
IL-6 has diverse cellular origins, such as fibroblasts, macrophages, pulmonary arterial smooth muscle cells, and Th1 lymphocytes [6, 7, 16]. In the very early stage of hypoxia or MCT exposure, increased levels of both IL-6 protein and mRNA can be detected in experimental PH model lungs [106, 107]. IL-6 plays a complex role in inflammation. IL-6 has both a proinflammatory effect by stimulating IL-4 production in Th2 cells and an anti-inflammatory effect by inhibiting IFN-γ production in Th1 cells [16]. Fibroblasts subjected to hypoxic conditions exhibit resistance to apoptosis in response to environmental changes [82]. The IL-6/STAT-3 signalling contributes to macrophage activation in hypoxia-induced PH models [108]. The IL-6/IL-21 axis is involved in M2 polarization, and IL-6 blockade inhibits M2 polarization in the hypoxia-exposed mouse model of PH [108]. Macrophage-derived IL-6 has been reported to promote pulmonary vascular remodelling [34]. The pharmacologic blockade or mRNA degradation of IL-6 in AMs can decrease right ventricular systolic pressure and alleviate PH progression in hypoxic mouse models [41].
IL-10
CD4 + Th2 cells, B lymphocytes and M2 polarized macrophages produce IL-10 [109]. IL-10 inhibits the proliferation of smooth muscle cells and the synthesis of proinflammatory cytokines in macrophages and Th1 lymphocytes [16]. Upregulation of IL-10 expression in PH rat models alleviated inflammatory infiltration and VMSC proliferation, as well as mPAP and right ventricular hypertrophy [110]. The protective effect of increased IL-10 levels in PH models was reported with alterations in other PH-related cytokines [31]. Hence, quantifying the protective effect of IL-10 in PH seems unlikely. Nevertheless, IL-10 is still a promising target for PH intervention.
IL-1
IL-1 is produced by diverse cell types, including monocytes, fibroblasts, T lymphocytes, neutrophils, and even pulmonary arterial smooth muscle cells [6, 7, 29, 30]. Upregulation of IL-1 occurs in the very early period of PH initiation and progression in response to hypoxia in macrophages [29]. Through the IL-1β/IL-1R1/MyD88 pathway, macrophages induce the proliferation of VSMCs in pulmonary arteries, suggesting that IL-1β is involved in pulmonary vascular remodelling in experimental PH [111].
IL-8 and migration inhibitory factor
IL-8 is expressed by macrophages or injured vascular cells. In the MCT-induced PH models, neutrophil infiltration and proinflammatory mediator expression are reduced by the upregulation of IL-8 in VECs [112]. The migration inhibitory factor (MIF) produced by T-cell lymphocytes is increased and activated in idiopathic PH patients, while an antagonist of MIF can partially reverse the development of experimental PH [113]. MIF functions as a main proinflammatory cytokine and VSMC proliferation promoter, and MIF expression and secretion levels are elevated in fibroblasts, monocytes, and endothelial cells after infection or hypoxia [114].
Chemokine
CCL2-CCR2 and CCL5-CCR5 are essential for the initiation and amplification of VSMCs in PH pathogenesis [115]. Increased expression of CCL2 and its ligand CCR2 was observed in the lungs of hypoxic PH models [40]. Proinflammatory fibroblasts in the vascular adventitia under hypoxic conditions express high levels of CCL2, CCL5, and CXCL12 [6, 116]. CCR2 is essential for the recruitment and development of M1-polarized macrophages, while increased CCL5 induces transmigration, adhesion, and activation [6, 11, 116]. Bordenave et al. reported a marked alleviation in distributed pulmonary hemodynamics and structural disorders in both the lung and heart when chemokine CXCL12 was neutralized in rat PH models [117]. Furthermore, CXCL12 neutraligand administration (chalcone 4, LIT-927, and AMD3100) decreased macrophage infiltration in the lungs of PH rat models [117]. In addition, the expression of CX3CR1 and its ligand CX3CL1 has been reported to increase in the lungs of hypoxia-induced PH models [40]. The inhibition of the CX3CL1-CX3CR1 signalling pathway effectively attenuates pulmonary inflammation and arterial remodelling, thus leading to a certain degree of improvement in hemodynamics [14]. Genetic deletion or pharmacological inhibition of CX3CR1 prevents hypoxia-induced PH by regulating monocyte recruitment, macrophage polarization and VSMC proliferation, which are associated with a changed balance between the M1 and M2 phenotypes [40]. These findings highlight the close interaction between CX3CL1 and macrophages, suggesting that the CX3CL1-CX3CR1 signalling pathway is a potential therapeutic target for PH.
Although these mediators are involved in the promotion or prevention of PH pathogenesis and progression, it is unwise to simply define them as “harmful” or “beneficial”. There is a complex and ingenious collaboration between mediators and cellular components. Cytokines and other inflammatory factors are synthesized and released by both macrophages and other cellular components, and subsequently activate or repress specific downstream signalling pathways, which in turn initiate pro- and/or anti-inflammatory responses and induce the activation or repression of other inflammation-related cells, such as immune cells, fibroblasts, ECs, and even smooth muscle cells. For example, PH patients with moderate cytokine levels have the best prognosis, while groups with the strongest or lowest immune signals have more severe clinical symptoms and worse outcomes [118], despite disordered circular inflammation factors being quite common in PH patients.
Interactions of macrophages with other cellular components in PH
Cellular abnormalities, including pulmonary vascular endothelium dysfunction, VSMC and adventitial fibroblast accumulation in arteries, and innate/adaptive immune system dysregulation, are critical promotors in PH [17]. Pulmonary ECs are transformed into proinflammatory phenotypes, then produce and release multiple cytokines and chemokines, leading to changes in endothelial communication between other resident vascular cells and circulating cells. Endothelial dysfunction and environmental stress induce metabolic alterations and proliferation in pulmonary VSMCs and fibroblasts. Moreover, perivascular inflammatory infiltration and circular cytokine alterations can be detected in the early stage of PH, suggesting that the immune system responds to microenvironmental changes and external stimuli even before substantial vascular remodelling occurs [17, 119]. These changes interact with other PH promotors.
Histological evidence has indicated concentric or eccentric intima-media thickening in muscularized precapillary arteries in individuals with established PH [3], which highlights the involvement of ECs and smooth muscle cells. ECs are essential for the initiation of PH. HIF-2α is activated in ECs under hypoxia, which is required for prominent proinflammatory genes Sdf1 (CXCL12), CXCR4, ICAM1 and VCAM1; then monocytes/macrophages and other circulation-/bone marrow-derived cells are recruited to lung at the early stage of PH development [35, 120]. Inhibition of membrane KCNK3 expression induced by hypoxia occurs mainly in VSMCs and pulmonary ECs, resulting in localized inflammation and vascular remodelling [65, 66]. Neutrophil extracellular traps are also involved in endothelial dysfunction and vascular homeostasis, which is attributed to proinflammation, pro-thrombosis, and the induction of NF-κB [121]. VEC ferroptosis in MCT-induced experimental PH induces HMGB1 release, leading to the upregulation of TLR4 expression in macrophages and triggering the inflammatory response via the HMGB1/TLR4/NLRP3 inflammasome signalling pathway [122]. Aryl hydrocarbon receptors in endothelial cells induce PH in rat models by upregulating inflammatory signals and increasing the accumulation of CD4 + cells [123]. The production of granulocyte-macrophage colony-stimulating factor in pulmonary artery VECs prevents chronically hypoxia-induced PH in mice and is upregulated when reducing BMPR2 expression [124]. In addition, decreased peroxiredoxin 6 in VECs induces the release of HMGB1 and activation of the TLR4/NLRP3 signalling pathway, thus promoting MCT-induced PH in rats [125]. The upregulated salusin-β in VECs and macrophages in PH rats has been reported to be a main contributor to both atherosclerosis and myocardial ischaemic disease, and promotes pulmonary VEC dysfunction via the activation of NF-κB signalling, thus inducing pulmonary inflammation and vascular remodelling [126].
Some factors expressed by activated macrophages can regulate vascular cell function. The proliferation of pulmonary VSMCs can be upregulated by VEGF and platelet-derived growth factor (PDGF), which are expressed by accumulated macrophages in pulmonary vessels [68, 99]. Moreover, pulmonary VSMC proliferation can be inhibited by prostaglandin D2 released by macrophages [78]. The proliferative phenotype of VSMC exhibits HIF-1α activation and enhanced glycolysis in normoxic conditions in experimental PH models [127]. Since the accumulation of VSMCs is very common in PH, the inhibition of smooth muscle cell proliferation and migration via regulating IL-1β, HIF or Nox4 has been reported to be a promising and effective therapeutic target for PH [37, 38, 69, 76]. In addition, the expression of MMP-1 and MMP-10 in M1-polarized macrophages increased in both PH patients and PH rat models, and the increased expression of MMP-1 and MMP-10 can promote the proliferation and promigratory phenotypes of VSMCs [96]. VSMC-derived TGF-β regulates the phenotypes of macrophages via p38-MAPK-dependent signalling, which in turn promotes VSMC proliferation [128]. Additionally, VSMC-derived multipotent vascular progenitor cells can differentiate into multiple phenotypes, such as mature VSMCs, resident macrophages, and endothelial-like cells [129]. These vascular progenitor cells might be derived from VSMCs and located at the normal pulmonary artery muscular-unmuscular border [130], or they can be derived from distinct PW1 + cells in perivascular zones [131].
Another typical characteristic of pulmonary vascular remodelling in PH is the presence of the proinflammatory fibroblast phenotype, as proposed by the outside-in hypothesis of pathological vascular remodelling [132]. When exposed to external stimuli such as hypoxia, fibroblasts produce increased levels of lactic acid, succinate, citrulline, IL-6, and other inflammation-related mediators, thus leading to microenvironmental alterations and anti-inflammatory effects to enhance proliferation and apoptosis resistance [82]. These proinflammatory phenotype fibroblasts exhibit increased HDAC I activity and express high levels of several typical products, such as classic cytokines (IL-1 and IL-6), macrophage chemoattractants (CCL2, CXCL12, and CCL5), the macrophage growth factor GM-CSF, and the adhesion protein VCAM-1, thus inducing the migration, adhesion, and activation of monocytes [6]. In addition, the MiR-124/PTBP1/PKM axis has been reported to be associated with metabolic reprogramming in proinflammatory fibroblasts [133]. Galectin-3 is also expressed in fibroblasts and serves as a pattern-recognition receptor and danger-associated molecular pattern in macrophages, leading to M2 polarization [134].
Notably, fibroblasts can activate the inflammatory phenotype and immune response of BMDMs, and increase aerobic glycolysis in BMDMs [116]. In addition to cytokines and chemokines, fibroblasts can also release extracellular vesicles [135], which can provide ROS [37]. The altered microenvironment forces macrophages to react. On the one hand, increased lactic acid, succinate, and IL-6 levels drive STAT3 and HIF-1 activation, thus promoting metabolic reprogramming in macrophages, while increased citrulline serves as a material for arginase in macrophages and maintains its function [82]. On the other hand, extracellular vesicles induce the transformation of phenotypes and promote the release of mediators in BMDMs, such as IL-4, G-CSF, CXCL13, MCP-5, CXCL11, IL-16, IL-17, IL-1β, CCL5, CCL1, MIG, IL-10, IL-23, TNF-α, IL-27, IFN-γ, IL-13, MIP-2, sICAM-1, IL-3, TIMP-1, and CXCL-1 [135]. Metabolic coordination between macrophages and fibroblasts is subsequently established, leading to persistent metabolic alterations and cell activation. In addition to VECs, VSMCs and fibroblasts, which have received the most attention, other cellular components, such as functional T cells, are also reported to play a protective role in vascular remodelling in a macrophage-related manner [136]. The main interactions between macrophages and other vessel cells are presented in Fig. 3.

The interaction between macrophages and other vessel cells in PH pathogenesis. Although macrophages might accelerate PH progression, pulmonary artery remodelling still results from abnormal angiogenesis. VECs, VSMCs and fibroblasts induce the recruitment of macrophages upon injury, while activated macrophages release variable mediators at different stages of inflammation. Influenced by these inflammatory mediators, vessel cells alter to specific phenotypes for survival or proliferation and release specific factors, such as IL-1, TGF-β, PDGF, or chemokines. These factors can regulate macrophage function and polarization in turn. For example, TGF-β contributes to M2-like polarization, while HMGB1 contributes to M1-like polarization. Together, these complex interactions eventually result in vascular remodelling and pulmonary hypertension
Macrophages as promising therapeutic targets for PH
Current medical interventions, which involve a combination of multiple agents, focus on improving the symptoms and prognosis of PH patients [137]. PDE5 inhibitors such as sildenafil, tadalafil, and vardenafil are effective for Group 1 PH and can be delivered by oral or inhaled treatment [138, 139]. In addition, prostacyclin, endothelin receptor antagonists, and guanylate cyclase stimulators are also effective for specific PH groups [140,141,142]. These classical pulmonary hypertension agents may also affect macrophages. Classic PDE5 inhibitors have been reported to inhibit the mobilization and recruitment of bone marrow-derived cells as well as the release of proinflammatory cytokines [143,144,145]. In addition, sGC stimulators (such as Riociguat) modulate liver inflammation via the inhibition of NLRP3 inflammasome-mediated IL-1β production in Kupffer cells [146]. Endothelin receptor antagonists, such as bosentan, macitentan and ambrisentan, suppress cytokines released from AMs and thus present anti-inflammatory potential [147]. Endothelin has been demonstrated to trigger M2 macrophage accumulation and ROS formation [148, 149]. Prostanoids (including epoprostenol, treprostinil, iloprost, beraprost, and selexipag) have been recognized as major regulators of inflammation progression and resolution [150]. A recent study revealed a novel cyclooxygenase/prostaglandin E2 axis-dependent mechanism of HIF-1α-induced TNF-α expression in macrophages [151]. These studies suggest that classic pharmacological agents for PH treatment might influence the immune system to some degree.
In addition to classical PH drugs, new therapeutic targets are emerging. The protein Regnase-1 is encoded by the ZC3H12A gene and is involved in mRNA degradation [152], which suppresses PH progression by degrading the mRNA of IL-6 and PDGF in AMs [41]. The intratracheal administration of Regnase-1-targeting morpholino oligonucleotides, which enhances Regnase-1 expression, has shown therapeutic efficacy by attenuating inflammatory cascades and fibrosis [153]. However, there is a lack of available pharmacological molecules to enhance Regnase-1 expression in vivo. Serum glucocorticoid-regulated kinase 1 (SGK1), a member of the serine/threonine kinase family, is associated with macrophage activation and inflammatory response. Deletion of SGK1 inhibits macrophage infiltration in the lungs of experimental PH models [39]. EMD638683, an SGK1 inhibitor, has been reported to suppress macrophage infiltration and prevent PH progression in MCT-induced rat models [154]. LDL receptor-related protein 1 (LRP1) plays an anti-inflammatory role in several diseases by maintaining cholesterol homeostasis, inhibiting migration, and blocking cytokine release [12]. In experimental PH, the increased expression of LRP1 in macrophages attenuates neointima formation by inducing the degradation of TGF-β2 [155]. LRP1 has also been identified as an integrator of TGF-β1-mediated vascular remodelling in PH. The expression of LRP1 is important to vascular homeostasis. PH induced by LRP1 deficiency can be reversed by pharmacologic PPARγ activation with pioglitazone [156]. Other LDL receptor family members are also potential therapeutic targets for PH [156].
Since HDACs are involved in perivascular fibroblast proliferation and monocyte activation, the inhibition of HDACs is believed to be an effective therapy for PH patients. However, the efficacy of HDACs pharmacological inhibitors in PH remains a controversial issue [6, 133, 157, 158]. Sotatercept, a ligand trap for multiple TGF-β family members, can suppress macrophage infiltration and reverse experimental PH [159]. The neutralization of CXCL12 improved pulmonary hemodynamics and structural disorders in both the lungs and heart in rat PH models [117]. Donepezil is an acetylcholinesterase inhibitor with a parasympathetic activistic effect. By suppressing M2-macrophage activation, donepezil reverses VSMC dysfunction in MCT-induced rat PH models [160]. The antifibrotic agent pirfenidone decreases macrophage IL-1β secretion in vitro [161].
Similar metabolic remodelling occurs in both fibroblasts and macrophages during PH pathogenesis, which indicates that short- or long-term metabolic regulation may potentially affect PH progression [82]. MTOB, an inhibitor of CtBP1, has been reported to attenuate glycolysis and inflammatory gene expression in macrophages and fibroblasts, thus leading to reversion to hypoxia-induced vascular remodelling and perivascular macrophage accumulation [116, 162]. Carbonic anhydrase inhibitors could also modulate AM activation and polarization and restore vascular homeostasis [163]. Molecular hydrogen restored the increased expression of MCP-1 and stromal cell-derived factor-1 in the MCT-induced PH model and thus suppressed adventitial macrophage accumulation [164]. A new series of N-(phenylmethyl)-benzoxazole-2-thiones, which act as MIF antagonists, have been reported to successfully reverse established MCT-induced PH and alleviate hemodynamics [165]. Additionally, Cheng et al. reported that a novel synthesized hybrid can both reduce the proliferation of perivascular cells and alleviate macrophage infiltration, thus attenuating MCT-induced PH in rat models [166]. Several researchers have attempted to prevent PH by injecting tolerogenic macrophages generated from monocytes into athymic nude rats [167]. Dynamin-related protein 1 induces both a polarity shift and inflammatory mediator expression in macrophages after vascular injury, resulting in intimal thickening [168], which is similar to vascular remodelling in PH. These studies suggest that regulating mitochondrial function in macrophages might be a promising therapeutic target for vascular diseases. In addition, several novel methods of drug delivery have been tested, offering the possibility of tissue-selective delivery [169, 170].
Numerous agents are being tested in clinical trials. These novel agents are summarized in Table 4. Although not all agents are specifically designed to target monocytes or macrophages, they might still affect macrophage function. Targeting mTOR signalling in macrophages has been demonstrated to be a potential therapeutic intervention in cardiovascular diseases [171]. AMP-activated protein kinase activation in macrophages suppresses the inflammatory response [172]. Recombinant relaxin peptides delivered by inhaled porous microspheres can suppress macrophage M2 polarization [173]. Dapagliflozin has been previously tested for its ability to attenuate inflammation and regulate macrophage polarization in cardiac fibrosis [174]. Besides the classical TGF-β and SMAD signalling pathways, activin and PDGF receptor-β are associated with macrophage activation and recruitment [175, 176]. Lysyl-tRNA synthetase induces immune responses through the activation of monocytes and macrophages [177]. Recently, a lysyl-tRNA synthetase (KARS1) inhibitor, ZMA001, is being evaluated as a potential PH therapy. The KARS1 inhibitor ZMA001 is designed to block KARS1-dependent infiltration of monocytes/macrophages and inhibit inflammatory responses in vessels (NCT05967299, registration date: August 1, 2023).
Conclusion
Recent advances have revealed the role of macrophages as key regulators of PH pathogenesis. When exposed to PH triggers, macrophages are recruited and then differentiate into different phenotypes at specific time points, inducing perivascular inflammation, endothelial dysfunction, and consequent vascular remodelling. Macrophages are involved in many typical hallmarks of PH, such as smooth muscle cell proliferation and fibroblast activation, which are essential for PH development. Inflammation-related soluble mediators are closely linked to these alterations. M1 and M2 macrophages are commonly treated as signs of different stages in PH pathogenesis. M1 macrophages promote inflammation, while M2 macrophages have anti-inflammatory functions and regulate tissue repair. Interestingly, M2 macrophages are regarded as promotors of advanced PH in most studies due to their wound-healing function, and M1 macrophages play protective roles in PH in specific circumstances [178]. Several novel macrophage targets have been reported in preclinical studies. However, these novel therapeutic targets need to be tested in further investigation. Macrophages are now recognized as candidate therapeutic targets for PH treatment due to their unique role in PH pathogenesis, suggesting a new strategy for preventing and even reversing PH progression.
Data availability
Not applicable.
Abbreviations
- AMs:
-
Alveolar macrophages
- BMDMs:
-
Bone marrow-derived macrophages
- BMPR2:
-
Bone morphogenetic protein receptor 2
- BTK:
-
Bruton’s tyrosine kinase
- CtBP1:
-
C-terminal binding protein 1
- DAMP:
-
Damage-associated molecular pattern
- EIF2:
-
Eukaryotic translation initiation factor 2
- EGFR:
-
Epidermal growth factor receptor
- FIZZ1:
-
Found in inflammatory zone
- G6PD:
-
Glucose-6-phosphate dehydrogenase
- GM-CSF:
-
Granulocyte-macrophage colony-stimulating factor
- HDAC:
-
Histone deacetylase
- HERV-K:
-
Human endogenous retrovirus K
- HIF:
-
Hypoxia-inducible factor
- HIMF:
-
Hypoxia-induced mitogenic factor
- HMGB1:
-
High mobility group box-1 protein
- Hmox1:
-
Heme oxygenase 1
- IMs:
-
Interstitial macrophages
- IRS2:
-
Insulin receptor substrate 2
- KARS1:
-
Lysyl-tRNA synthetase
- KCNK3:
-
Potassium channel subfamily K member 3
- LRP1:
-
LDL receptor-related protein 1
- LTB4:
-
Leukotriene B4
- MAPK:
-
Mitogen-activated protein kinase
- MCP:
-
Monocyte chemoattractant protein
- MCT:
-
Monocrotaline
- MIF:
-
Migration inhibitory factor
- mPAP:
-
Mean pulmonary arterial pressure
- mTOR:
-
Mammalian target of rapamycin
- mTORC1:
-
mTOR complex 1
- NF-κB:
-
Nuclear factor kappa-B
- NLRP3:
-
NOD-like receptor thermal protein domain associated protein 3
- Nox4:
-
NADPH oxidase 4
- P2X7R:
-
P2 × 7 purinergic receptor
- PDE5:
-
Phosphodiesterase 5
- PDGF:
-
Platelet-derived growth factor
- PH:
-
Pulmonary hypertension
- Pfkfb3:
-
6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase 3
- PKM:
-
Pyruvate kinase muscle
- PLC:
-
Phospholipase C
- PTBP1:
-
Polypyrimidine tract binding protein 1
- RAGE:
-
Receptor for advanced glycation end products
- RELM-α:
-
Resistin-like molecule-α
- ROS:
-
Reactive oxygen species
- RUNX1:
-
Runt-related transcription factor 1
- sGC:
-
Soluble guanylate cyclase
- sICAM-1:
-
Soluble intercellular adhesion molecule-1
- SIRT1:
-
Sirtuin 1
- SGK1:
-
Serum glucocorticoid-regulated kinase-1
- STAT-3:
-
Signal transducer and activator of transcription 3
- TIMP-1:
-
Tissue inhibitor of metalloproteinase 1
- TGF-β:
-
Transforming growth factor-β
- TLR4:
-
Toll-like receptor 4
- VCAM-1:
-
Vascular cell adhesion molecule-1
- VECs:
-
Vascular endothelial cells
- VEGF:
-
Vascular endothelial growth factor
- VSMCs:
-
Vascular smooth muscle cells
- Ythdf2:
-
YT521-B homology domain family 2
References
Humbert M, Kovacs G, Hoeper MM, Badagliacca R, Berger RMF, Brida M, Carlsen J, Coats AJS, Escribano-Subias P, Ferrari P et al. 2022 ESC/ERS guidelines for the diagnosis and treatment of pulmonary hypertension. Eur Respir J 2023, 61.
Hoeper MM, Humbert M, Souza R, Idrees M, Kawut SM, Sliwa-Hahnle K, Jing ZC, Gibbs JS. A global view of pulmonary hypertension. Lancet Respir Med. 2016;4:306–22.
Stacher E, Graham BB, Hunt JM, Gandjeva A, Groshong SD, McLaughlin VV, Jessup M, Grizzle WE, Aldred MA, Cool CD, Tuder RM. Modern age pathology of pulmonary arterial hypertension. Am J Respir Crit Care Med. 2012;186:261–72.
Thenappan T, Ormiston ML, Ryan JJ, Archer SL. Pulmonary arterial hypertension: pathogenesis and clinical management. BMJ. 2018;360:j5492.
Rabinovitch M, Guignabert C, Humbert M, Nicolls MR. Inflammation and immunity in the pathogenesis of pulmonary arterial hypertension. Circ Res. 2014;115:165–75.
Li M, Riddle SR, Frid MG, El Kasmi KC, McKinsey TA, Sokol RJ, Strassheim D, Meyrick B, Yeager ME, Flockton AR, et al. Emergence of fibroblasts with a proinflammatory epigenetically altered phenotype in severe hypoxic pulmonary hypertension. J Immunol. 2011;187:2711–22.
Hu Y, Chi L, Kuebler WM, Goldenberg NM. Perivascular inflammation in pulmonary arterial hypertension. Cells 2020, 9.
Lekva T, Gullestad L, Broch K, Aukrust P, Andreassen AK, Ueland T. Distinct patterns of soluble leukocyte activation markers are associated with etiology and outcomes in precapillary pulmonary hypertension. Sci Rep. 2020;10:18540.
Chen S, Yan D, Qiu A. The role of macrophages in pulmonary hypertension: Pathogenesis and targeting. Int Immunopharmacol. 2020;88:106934.
Florentin J, Dutta P. Origin and production of inflammatory perivascular macrophages in pulmonary hypertension. Cytokine. 2017;100:11–5.
Chen L, Zhang Z, Barletta KE, Burdick MD, Mehrad B. Heterogeneity of lung mononuclear phagocytes during pneumonia: contribution of chemokine receptors. Am J Physiol Lung Cell Mol Physiol. 2013;305:L702–711.
He Z, Wang G, Wu J, Tang Z, Luo M. The molecular mechanism of LRP1 in physiological vascular homeostasis and signal transduction pathways. Biomed Pharmacother. 2021;139:111667.
Montani D, Perros F, Gambaryan N, Girerd B, Dorfmuller P, Price LC, Huertas A, Hammad H, Lambrecht B, Simonneau G, et al. C-kit-positive cells accumulate in remodeled vessels of idiopathic pulmonary arterial hypertension. Am J Respir Crit Care Med. 2011;184:116–23.
Florentin J, Coppin E, Vasamsetti SB, Zhao J, Tai YY, Tang Y, Zhang Y, Watson A, Sembrat J, Rojas M, et al. Inflammatory macrophage expansion in Pulmonary Hypertension depends upon mobilization of blood-borne monocytes. J Immunol. 2018;200:3612–25.
Willis GR, Fernandez-Gonzalez A, Reis M, Mitsialis SA, Kourembanas S. Macrophage immunomodulation: the gatekeeper for mesenchymal stem cell derived-exosomes in Pulmonary arterial hypertension? Int J Mol Sci 2018, 19.
Tomaszewski M, Bebnowska D, Hrynkiewicz R, Dworzynski J, Niedzwiedzka-Rystwej P, Kopec G, Grywalska E. Role of the Immune System Elements in Pulmonary arterial hypertension. J Clin Med 2021, 10.
Humbert M, Guignabert C, Bonnet S, Dorfmuller P, Klinger JR, Nicolls MR, Olschewski AJ, Pullamsetti SS, Schermuly RT, Stenmark KR, Rabinovitch M. Pathology and pathobiology of pulmonary hypertension: state of the art and research perspectives. Eur Respir J 2019, 53.
Crosswhite P, Chen K, Sun Z. AAV delivery of tumor necrosis factor-alpha short hairpin RNA attenuates cold-induced pulmonary hypertension and pulmonary arterial remodeling. Hypertension. 2014;64:1141–50.
Liu J, Ye X, Ji D, Zhou X, Qiu C, Liu W, Yu L. Diesel exhaust inhalation exposure induces pulmonary arterial hypertension in mice. Environ Pollut. 2018;237:747–55.
Minakawa K, Yokokawa T, Ueda K, Nakajima O, Misaka T, Kimishima Y, Wada K, Tomita Y, Miura S, Sato Y, et al. Myeloproliferative neoplasm-driving Calr frameshift promotes the development of pulmonary hypertension in mice. J Hematol Oncol. 2021;14:52.
Mercurio V, Cuomo A, Naranjo M, Hassoun PM. Inflammatory mechanisms in the Pathogenesis of Pulmonary arterial hypertension: recent advances. Compr Physiol. 2021;11:1805–29.
Tamosiuniene R, Tian W, Dhillon G, Wang L, Sung YK, Gera L, Patterson AJ, Agrawal R, Rabinovitch M, Ambler K, et al. Regulatory T cells limit vascular endothelial injury and prevent pulmonary hypertension. Circ Res. 2011;109:867–79.
Chen Y, Ouyang T, Yin Y, Fang C, Tang CE, Jiang L, Luo F. Identification of immune-related hub genes and analysis of infiltrated immune cells of idiopathic pulmonary artery hypertension. Front Cardiovasc Med. 2023;10:1125063.
Zeng H, Liu X, Zhang Y. Identification of potential biomarkers and Immune infiltration characteristics in idiopathic pulmonary arterial hypertension using Bioinformatics Analysis. Front Cardiovasc Med. 2021;8:624714.
Li C, Xia J, Yiminniyaze R, Dong L, Li S. Hub genes and Immune Cell Infiltration in Hypoxia-Induced Pulmonary Hypertension: Bioinformatics Analysis and in vivo validation. Comb Chem High Throughput Screen 2023.
Dignam JP, Scott TE, Kemp-Harper BK, Hobbs AJ. Animal models of pulmonary hypertension: getting to the heart of the problem. Br J Pharmacol. 2022;179:811–37.
Dupuis J, Weissmann N. Animal Models of Pulmonary Hypertension. In Textbook of Pulmonary Vascular Disease Edited by Yuan JXJ, Garcia JGN, West JB, Hales CA, Rich S, Archer SL. Boston, MA: Springer US; 2011: 453–458.
Hashimoto R, Gupte SA. G6PD is a critical enabler of hypoxia-induced accumulation of macrophages and platelets in mice lungs and contributor to lung inflammation. Vascul Pharmacol. 2022;144:106976.
Campbell NV, Mickael C, Kumar S, Zhang H, Campbell IL, Gillen AE, Trentin CO, Diener K, Gao B, Kheyfets VO, et al. Single-cell RNA sequencing and binary hierarchical clustering define lung interstitial macrophage heterogeneity in response to hypoxia. Am J Physiol Lung Cell Mol Physiol. 2022;323:L58–68.
Pugliese SC, Kumar S, Janssen WJ, Graham BB, Frid MG, Riddle SR, El Kasmi KC, Stenmark KR. A time- and compartment-specific activation of Lung macrophages in Hypoxic Pulmonary Hypertension. J Immunol. 2017;198:4802–12.
Vergadi E, Chang MS, Lee C, Liang OD, Liu X, Fernandez-Gonzalez A, Mitsialis SA, Kourembanas S. Early macrophage recruitment and alternative activation are critical for the later development of hypoxia-induced pulmonary hypertension. Circulation. 2011;123:1986–95.
Tian W, Jiang X, Tamosiuniene R, Sung YK, Qian J, Dhillon G, Gera L, Farkas L, Rabinovitch M, Zamanian RT, et al. Blocking macrophage leukotriene b4 prevents endothelial injury and reverses pulmonary hypertension. Sci Transl Med. 2013;5:200ra117.
Kojima H, Tokunou T, Takahara Y, Sunagawa K, Hirooka Y, Ichiki T, Tsutsui H. Hypoxia-inducible factor-1 alpha deletion in myeloid lineage attenuates hypoxia-induced pulmonary hypertension. Physiol Rep. 2019;7:e14025.
Pullamsetti SS, Mamazhakypov A, Weissmann N, Seeger W, Savai R. Hypoxia-inducible factor signaling in pulmonary hypertension. J Clin Invest. 2020;130:5638–51.
Hu CJ, Poth JM, Zhang H, Flockton A, Laux A, Kumar S, McKeon B, Mouradian G, Li M, Riddle S et al. Suppression of HIF2 signalling attenuates the initiation of hypoxia-induced pulmonary hypertension. Eur Respir J 2019, 54.
Yu M, Wu X, Peng L, Yang M, Zhou H, Xu J, Wang J, Wang H, Xie W, Kong H. Inhibition of Bruton’s Tyrosine Kinase Alleviates Monocrotaline-Induced Pulmonary Arterial Hypertension by Modulating Macrophage Polarization. Oxid Med Cell Longev 2022, 2022:6526036.
Barman SA, Chen F, Su Y, Dimitropoulou C, Wang Y, Catravas JD, Han W, Orfi L, Szantai-Kis C, Keri G, et al. NADPH oxidase 4 is expressed in pulmonary artery adventitia and contributes to hypertensive vascular remodeling. Arterioscler Thromb Vasc Biol. 2014;34:1704–15.
Rong W, Liu C, Li X, Wan N, Wei L, Zhu W, Bai P, Li M, Ou Y, Li F, et al. Caspase-8 promotes pulmonary hypertension by activating macrophage-Associated inflammation and IL-1beta (interleukin 1beta) production. Arterioscler Thromb Vasc Biol. 2022;42:613–31.
Xi X, Zhang J, Wang J, Chen Y, Zhang W, Zhang X, Du J, Zhu G. SGK1 Mediates Hypoxic Pulmonary Hypertension through Promoting Macrophage Infiltration and Activation. Anal Cell Pathol (Amst) 2019, 2019:3013765.
Amsellem V, Abid S, Poupel L, Parpaleix A, Rodero M, Gary-Bobo G, Latiri M, Dubois-Rande JL, Lipskaia L, Combadiere C, Adnot S. Roles for the CX3CL1/CX3CR1 and CCL2/CCR2 Chemokine systems in Hypoxic Pulmonary Hypertension. Am J Respir Cell Mol Biol. 2017;56:597–608.
Yaku A, Inagaki T, Asano R, Okazawa M, Mori H, Sato A, Hia F, Masaki T, Manabe Y, Ishibashi T, et al. Regnase-1 prevents pulmonary arterial hypertension through mRNA degradation of Interleukin-6 and platelet-derived growth factor in alveolar macrophages. Circulation. 2022;146:1006–22.
Zaloudikova M, Vytasek R, Vajnerova O, Hnilickova O, Vizek M, Hampl V, Herget J. Depletion of alveolar macrophages attenuates hypoxic pulmonary hypertension but not hypoxia-induced increase in serum concentration of MCP-1. Physiol Res. 2016;65:763–8.
Al-Qazazi R, Lima PDA, Prisco SZ, Potus F, Dasgupta A, Chen KH, Tian L, Bentley RET, Mewburn J, Martin AY, et al. Macrophage-NLRP3 activation promotes right ventricle failure in pulmonary arterial hypertension. Am J Respir Crit Care Med. 2022;206:608–24.
Gorr MW, Sriram K, Chinn AM, Muthusamy A, Insel PA. Transcriptomic profiles reveal differences between the right and left ventricle in normoxia and hypoxia. Physiol Rep. 2020;8:e14344.
Zawia A, Arnold ND, West L, Pickworth JA, Turton H, Iremonger J, Braithwaite AT, Canedo J, Johnston SA, Thompson AAR, et al. Altered macrophage polarization induces experimental pulmonary hypertension and is observed in patients with Pulmonary arterial hypertension. Arterioscler Thromb Vasc Biol. 2021;41:430–45.
Badesch DB, Raskob GE, Elliott CG, Krichman AM, Farber HW, Frost AE, Barst RJ, Benza RL, Liou TG, Turner M et al. Pulmonary Arterial Hypertension. Chest 2010, 137:376–387.
Umar S, Iorga A, Matori H, Nadadur RD, Li J, Maltese F, van der Laarse A, Eghbali M. Estrogen rescues preexisting severe pulmonary hypertension in rats. Am J Respir Crit Care Med. 2011;184:715–23.
Wits M, Becher C, de Man F, Sanchez-Duffhues G, Goumans MJ. Sex-biased TGFbeta signalling in pulmonary arterial hypertension. Cardiovasc Res. 2023;119:2262–77.
Cheron C, McBride SA, Antigny F, Girerd B, Chouchana M, Chaumais MC, Jais X, Bertoletti L, Sitbon O, Weatherald J et al. Sex and gender in pulmonary arterial hypertension. Eur Respir Rev 2021, 30.
Dignam JP, Sharma S, Stasinopoulos I, MacLean MR. Pulmonary arterial hypertension: sex matters. Br J Pharmacol 2023.
Mair KM, Wright AF, Duggan N, Rowlands DJ, Hussey MJ, Roberts S, Fullerton J, Nilsen M, Loughlin L, Thomas M, MacLean MR. Sex-dependent influence of endogenous estrogen in Pulmonary Hypertension. Am J Respir Crit Care Med. 2014;190:456–67.
Tamosiuniene R, Manouvakhova O, Mesange P, Saito T, Qian J, Sanyal M, Lin Y-C, Nguyen LP, Luria A, Tu AB, et al. Dominant Role for Regulatory T Cells in protecting females against pulmonary hypertension. Circul Res. 2018;122:1689–702.
Umar S, Cunningham CM, Itoh Y, Moazeni S, Vaillancourt M, Sarji S, Centala A, Arnold AP, Eghbali M. The Y chromosome plays a protective role in experimental hypoxic pulmonary hypertension. Am J Respir Crit Care Med. 2018;197:952–5.
Watanabe S, Markov NS, Lu Z, Piseaux Aillon R, Soberanes S, Runyan CE, Ren Z, Grant RA, Maciel M, Abdala-Valencia H et al. Resetting proteostasis with ISRIB promotes epithelial differentiation to attenuate pulmonary fibrosis. Proc Natl Acad Sci U S A 2021, 118.
Fan Y, Hao Y, Gao D, Li G, Zhang Z. Phenotype and function of macrophage polarization in monocrotaline-induced pulmonary arterial hypertension rat model. Physiol Res. 2021;70:213–26.
Park JB, Suh M, Park JY, Park JK, Kim YI, Kim H, Cho YS, Kang H, Kim K, Choi JH, et al. Assessment of inflammation in pulmonary artery hypertension by (68)Ga-Mannosylated Human serum albumin. Am J Respir Crit Care Med. 2020;201:95–106.
Klinger JR, Pereira M, Del Tatto M, Brodsky AS, Wu KQ, Dooner MS, Borgovan T, Wen S, Goldberg LR, Aliotta JM, et al. Mesenchymal stem cell Extracellular vesicles Reverse Sugen/Hypoxia Pulmonary hypertension in rats. Am J Respir Cell Mol Biol. 2020;62:577–87.
Pugliese SC, Poth JM, Fini MA, Olschewski A, El Kasmi KC, Stenmark KR. The role of inflammation in hypoxic pulmonary hypertension: from cellular mechanisms to clinical phenotypes. Am J Physiol Lung Cell Mol Physiol. 2015;308:L229–252.
Hu L, Yu Y, Shen Y, Huang H, Lin D, Wang K, Yu Y, Li K, Cao Y, Wang Q, et al. Ythdf2 promotes pulmonary hypertension by suppressing Hmox1-dependent anti-inflammatory and antioxidant function in alveolar macrophages. Redox Biol. 2023;61:102638.
Cai L, Li D, Feng Z, Gu X, Xu Q, Li Q. YTHDF2 Regulates Macrophage Polarization through NF-kappaB and MAPK Signaling Pathway Inhibition or p53 Degradation. Dis Markers 2022, 2022:3153362.
Kumar R, Mickael C, Kassa B, Sanders L, Hernandez-Saavedra D, Koyanagi DE, Kumar S, Pugliese SC, Thomas S, McClendon J, et al. Interstitial macrophage-derived thrombospondin-1 contributes to hypoxia-induced pulmonary hypertension. Cardiovasc Res. 2020;116:2021–30.
Johns RA, Takimoto E, Meuchel LW, Elsaigh E, Zhang A, Heller NM, Semenza GL, Yamaji-Kegan K. Hypoxia-inducible factor 1alpha is a critical downstream Mediator for Hypoxia-Induced Mitogenic factor (FIZZ1/RELMalpha)-Induced Pulmonary Hypertension. Arterioscler Thromb Vasc Biol. 2016;36:134–44.
Angelini DJ, Su Q, Yamaji-Kegan K, Fan C, Skinner JT, Poloczek A, El-Haddad H, Cheadle C, Johns RA. Hypoxia-induced mitogenic factor (HIMF/FIZZ1/RELMalpha) in chronic hypoxia- and antigen-mediated pulmonary vascular remodeling. Respir Res. 2013;14:1.
Lin Q, Fan C, Skinner JT, Hunter EN, Macdonald AA, Illei PB, Yamaji-Kegan K, Johns RA. RELMalpha licenses macrophages for damage-Associated Molecular Pattern activation to instigate pulmonary vascular remodeling. J Immunol. 2019;203:2862–71.
Han L, Song N, Hu X, Zhu A, Wei X, Liu J, Yuan S, Mao W, Chen X. Inhibition of RELM-beta prevents hypoxia-induced overproliferation of human pulmonary artery smooth muscle cells by reversing PLC-mediated KCNK3 decline. Life Sci. 2020;246:117419.
Antigny F, Hautefort A, Meloche J, Belacel-Ouari M, Manoury B, Rucker-Martin C, Pechoux C, Potus F, Nadeau V, Tremblay E, et al. Potassium Channel Subfamily K Member 3 (KCNK3) contributes to the development of pulmonary arterial hypertension. Circulation. 2016;133:1371–85.
Yamaji-Kegan K, Su Q, Angelini DJ, Myers AC, Cheadle C, Johns RA. Hypoxia-induced mitogenic factor (HIMF/FIZZ1/RELMalpha) increases lung inflammation and activates pulmonary microvascular endothelial cells via an IL-4-dependent mechanism. J Immunol. 2010;185:5539–48.
Thenappan T, Goel A, Marsboom G, Fang YH, Toth PT, Zhang HJ, Kajimoto H, Hong Z, Paul J, Wietholt C, et al. A central role for CD68(+) macrophages in hepatopulmonary syndrome. Reversal by macrophage depletion. Am J Respir Crit Care Med. 2011;183:1080–91.
Huertas A, Tu L, Thuillet R, Le Hiress M, Phan C, Ricard N, Nadaud S, Fadel E, Humbert M, Guignabert C. Leptin signalling system as a target for pulmonary arterial hypertension therapy. Eur Respir J. 2015;45:1066–80.
Ranchoux B, Nadeau V, Bourgeois A, Provencher S, Tremblay E, Omura J, Cote N, Abu-Alhayja’a R, Dumais V, Nachbar RT, et al. Metabolic syndrome exacerbates pulmonary hypertension due to Left Heart Disease. Circ Res. 2019;125:449–66.
Abe H, Semba H, Takeda N. The roles of Hypoxia Signaling in the Pathogenesis of Cardiovascular diseases. J Atheroscler Thromb. 2017;24:884–94.
Caldwell RW, Rodriguez PC, Toque HA, Narayanan SP, Caldwell RB. Arginase: a multifaceted enzyme important in Health and Disease. Physiol Rev. 2018;98:641–65.
Takeda N, O’Dea EL, Doedens A, Kim JW, Weidemann A, Stockmann C, Asagiri M, Simon MC, Hoffmann A, Johnson RS. Differential activation and antagonistic function of HIF-alpha isoforms in macrophages are essential for NO homeostasis. Genes Dev. 2010;24:491–501.
Bai P, Lyu L, Yu T, Zuo C, Fu J, He Y, Wan Q, Wan N, Jia D, Lyu A. Macrophage-derived Legumain promotes pulmonary hypertension by activating the MMP (Matrix Metalloproteinase)-2/TGF (transforming growth Factor)-beta1 signaling. Arterioscler Thromb Vasc Biol. 2019;39:e130–45.
Qian J, Tian W, Jiang X, Tamosiuniene R, Sung YK, Shuffle EM, Tu AB, Valenzuela A, Jiang S, Zamanian RT, et al. Leukotriene B4 activates pulmonary artery adventitial fibroblasts in Pulmonary Hypertension. Hypertension. 2015;66:1227–39.
Song JL, Zheng SY, He RL, Gui LX, Lin MJ, Sham JSK. Serotonin and chronic hypoxic pulmonary hypertension activate a NADPH oxidase 4 and TRPM2 dependent pathway for pulmonary arterial smooth muscle cell proliferation and migration. Vascul Pharmacol. 2021;138:106860.
Wu DD, Deng Y, Liao J, Xie SS, Meng H, Lan WF. STING mediates SU5416/hypoxia-induced pulmonary arterial hypertension in rats by regulating macrophage NLRP3 inflammasome activation. Immunobiology. 2023;228:152345.
Jia D, Bai P, Wan N, Liu J, Zhu Q, He Y, Chen G, Wang J, Chen H, Wang C, et al. Niacin attenuates pulmonary hypertension through H-PGDS in macrophages. Circ Res. 2020;127:1323–36.
Krishnamachary B, Mahajan A, Kumar A, Agarwal S, Mohan A, Chen L, Hsue PY, Chalise P, Morris A, Dhillon NK. Extracellular vesicle TGF-beta1 is linked to Cardiopulmonary Dysfunction in Human Immunodeficiency Virus. Am J Respir Cell Mol Biol. 2021;65:413–29.
Wu YT, Xu WT, Zheng L, Wang S, Wei J, Liu MY, Zhou HP, Li QF, Shi X, Lv X. 4-octyl itaconate ameliorates alveolar macrophage pyroptosis against ARDS via rescuing mitochondrial dysfunction and suppressing the cGAS/STING pathway. Int Immunopharmacol. 2023;118:110104.
O’Neill LA, Pearce EJ. Immunometabolism governs dendritic cell and macrophage function. J Exp Med. 2016;213:15–23.
D’Alessandro A, El Kasmi KC, Plecita-Hlavata L, Jezek P, Li M, Zhang H, Gupte SA, Stenmark KR. Hallmarks of Pulmonary Hypertension: mesenchymal and inflammatory cell metabolic reprogramming. Antioxid Redox Signal. 2018;28:230–50.
El Kasmi KC, Stenmark KR. Contribution of metabolic reprogramming to macrophage plasticity and function. Semin Immunol. 2015;27:267–75.
Cottrill KA, Chan SY. Metabolic dysfunction in pulmonary hypertension: the expanding relevance of the Warburg effect. Eur J Clin Invest. 2013;43:855–65.
Sadiku P, Walmsley SR. Hypoxia and the regulation of myeloid cell metabolic imprinting: consequences for the inflammatory response. EMBO Rep 2019, 20.
Lian G, Li X, Zhang L, Zhang Y, Sun L, Zhang X, Liu H, Pang Y, Kong W, Zhang T, et al. Macrophage metabolic reprogramming aggravates aortic dissection through the HIF1alpha-ADAM17 pathway(). EBioMedicine. 2019;49:291–304.
Palmieri EM, Gonzalez-Cotto M, Baseler WA, Davies LC, Ghesquiere B, Maio N, Rice CM, Rouault TA, Cassel T, Higashi RM, et al. Nitric oxide orchestrates metabolic rewiring in M1 macrophages by targeting aconitase 2 and pyruvate dehydrogenase. Nat Commun. 2020;11:698.
Wang L, Zhang X, Cao Y, Ma Q, Mao X, Xu J, Yang Q, Zhou Y, Lucas R, Fulton DJ, et al. Mice with a specific deficiency of Pfkfb3 in myeloid cells are protected from hypoxia-induced pulmonary hypertension. Br J Pharmacol. 2021;178:1055–72.
Nakahara M, Ito H, Skinner JT, Lin Q, Tamosiuniene R, Nicolls MR, Keegan AD, Johns RA, Yamaji-Kegan K. The inflammatory role of dysregulated IRS2 in pulmonary vascular remodeling under hypoxic conditions. Am J Physiol Lung Cell Mol Physiol. 2021;321:L416–28.
Liu G, Bi Y, Shen B, Yang H, Zhang Y, Wang X, Liu H, Lu Y, Liao J, Chen X, Chu Y. SIRT1 limits the function and fate of myeloid-derived suppressor cells in tumors by orchestrating HIF-1alpha-dependent glycolysis. Cancer Res. 2014;74:727–37.
Jing C, Castro-Dopico T, Richoz N, Tuong ZK, Ferdinand JR, Lok LSC, Loudon KW, Banham GD, Mathews RJ, Cader Z, et al. Macrophage metabolic reprogramming presents a therapeutic target in lupus nephritis. Proc Natl Acad Sci U S A. 2020;117:15160–71.
Gao RY, Wang M, Liu Q, Feng D, Wen Y, Xia Y, Colgan SP, Eltzschig HK, Ju C. Hypoxia-inducible Factor-2alpha Reprograms Liver macrophages to protect against Acute Liver Injury through the production of Interleukin-6. Hepatology. 2020;71:2105–17.
Goncalves SM, Duarte-Oliveira C, Campos CF, Aimanianda V, Ter Horst R, Leite L, Mercier T, Pereira P, Fernandez-Garcia M, Antunes D, et al. Phagosomal removal of fungal melanin reprograms macrophage metabolism to promote antifungal immunity. Nat Commun. 2020;11:2282.
Alexander RK, Liou YH, Knudsen NH, Starost KA, Xu C, Hyde AL, Liu S, Jacobi D, Liao NS, Lee CH. Bmal1 integrates mitochondrial metabolism and macrophage activation. Elife 2020, 9.
Murugina NE, Budikhina AS, Dagil YA, Maximchik PV, Balyasova LS, Murugin VV, Melnikov MV, Sharova VS, Nikolaeva AM, Chkadua GZ, et al. Glycolytic reprogramming of macrophages activated by NOD1 and TLR4 agonists: no association with proinflammatory cytokine production in normoxia. J Biol Chem. 2020;295:3099–114.
Chi PL, Cheng CC, Hung CC, Wang MT, Liu HY, Ke MW, Shen MC, Lin KC, Kuo SH, Hsieh PP, et al. MMP-10 from M1 macrophages promotes pulmonary vascular remodeling and pulmonary arterial hypertension. Int J Biol Sci. 2022;18:331–48.
Jeong EM, Pereira M, So EY, Wu KQ, Del Tatto M, Wen S, Dooner MS, Dubielecka PM, Reginato AM, Ventetuolo CE, et al. Targeting RUNX1 as a novel treatment modality for pulmonary arterial hypertension. Cardiovasc Res. 2022;118:3211–24.
Gu S, Mickael C, Kumar R, Lee MH, Sanders L, Kassa B, Harral J, Williams J, Hansen KC, Stenmark KR, et al. The role of macrophages in right ventricular remodeling in experimental pulmonary hypertension. Pulm Circ. 2022;12:e12105.
Ntokou A, Dave JM, Kauffman AC, Sauler M, Ryu C, Hwa J, Herzog EL, Singh I, Saltzman WM, Greif DM. Macrophage-derived PDGF-B induces muscularization in murine and human pulmonary hypertension. JCI Insight 2021, 6.
Batool M, Berghausen EM, Zierden M, Vantler M, Schermuly RT, Baldus S, Rosenkranz S, Ten Freyhaus H. The six-transmembrane protein Stamp2 ameliorates pulmonary vascular remodeling and pulmonary hypertension in mice. Basic Res Cardiol. 2020;115:68.
West JD, Chen X, Ping L, Gladson S, Hamid R, Lloyd JE, Talati M. Adverse effects of BMPR2 suppression in macrophages in animal models of pulmonary hypertension. Pulm Circ. 2019;10:2045894019856483.
Yin J, You S, Liu H, Chen L, Zhang C, Hu H, Xue M, Cheng W, Wang Y, Li X, et al. Role of P2X(7)R in the development and progression of pulmonary hypertension. Respir Res. 2017;18:127.
Saito T, Miyagawa K, Chen SY, Tamosiuniene R, Wang L, Sharpe O, Samayoa E, Harada D, Moonen JAJ, Cao A, et al. Upregulation of human endogenous Retrovirus-K is linked to immunity and inflammation in pulmonary arterial hypertension. Circulation. 2017;136:1920–35.
Frid MG, McKeon BA, Thurman JM, Maron BA, Li M, Zhang H, Kumar S, Sullivan T, Laskowsky J, Fini MA, et al. Immunoglobulin-driven complement activation regulates Proinflammatory Remodeling in Pulmonary Hypertension. Am J Respir Crit Care Med. 2020;201:224–39.
Hojda SE, Chis IC, Clichici S. Biomarkers in pulmonary arterial hypertension. Diagnostics (Basel) 2022, 12.
Savale L, Tu L, Rideau D, Izziki M, Maitre B, Adnot S, Eddahibi S. Impact of interleukin-6 on hypoxia-induced pulmonary hypertension and lung inflammation in mice. Respir Res. 2009;10:6.
Tang C, Luo Y, Li S, Huang B, Xu S, Li L. Characteristics of inflammation process in monocrotaline-induced pulmonary arterial hypertension in rats. Biomed Pharmacother. 2021;133:111081.
Hashimoto-Kataoka T, Hosen N, Sonobe T, Arita Y, Yasui T, Masaki T, Minami M, Inagaki T, Miyagawa S, Sawa Y, et al. Interleukin-6/interleukin-21 signaling axis is critical in the pathogenesis of pulmonary arterial hypertension. Proc Natl Acad Sci U S A. 2015;112:E2677–2686.
Gordon S, Martinez FO. Alternative activation of macrophages: mechanism and functions. Immunity. 2010;32:593–604.
Ito T, Okada T, Miyashita H, Nomoto T, Nonaka-Sarukawa M, Uchibori R, Maeda Y, Urabe M, Mizukami H, Kume A, et al. Interleukin-10 expression mediated by an adeno-associated virus vector prevents monocrotaline-induced pulmonary arterial hypertension in rats. Circ Res. 2007;101:734–41.
Parpaleix A, Amsellem V, Houssaini A, Abid S, Breau M, Marcos E, Sawaki D, Delcroix M, Quarck R, Maillard A, et al. Role of interleukin-1 receptor 1/MyD88 signalling in the development and progression of pulmonary hypertension. Eur Respir J. 2016;48:470–83.
Fu J, Chen YF, Zhao X, Creighton JR, Guo Y, Hage FG, Oparil S, Xing DD. Targeted delivery of pulmonary arterial endothelial cells overexpressing interleukin-8 receptors attenuates monocrotaline-induced pulmonary vascular remodeling. Arterioscler Thromb Vasc Biol. 2014;34:1539–47.
Le Hiress M, Tu L, Ricard N, Phan C, Thuillet R, Fadel E, Dorfmuller P, Montani D, de Man F, Humbert M, et al. Proinflammatory signature of the dysfunctional endothelium in Pulmonary Hypertension. Role of the Macrophage Migration Inhibitory Factor/CD74 complex. Am J Respir Crit Care Med. 2015;192:983–97.
Ahmed M, Miller E. Macrophage migration inhibitory factor (MIF) in the development and progression of pulmonary arterial hypertension. Glob Cardiol Sci Pract 2018, 2018:14.
Abid S, Marcos E, Parpaleix A, Amsellem V, Breau M, Houssaini A, Vienney N, Lefevre M, Derumeaux G, Evans S et al. CCR2/CCR5-mediated macrophage-smooth muscle cell crosstalk in pulmonary hypertension. Eur Respir J 2019, 54.
Li M, Riddle S, Kumar S, Poczobutt J, McKeon BA, Frid MG, Ostaff M, Reisz JA, Nemkov T, Fini MA, et al. Microenvironmental Regulation of Macrophage Transcriptomic and Metabolomic profiles in Pulmonary Hypertension. Front Immunol. 2021;12:640718.
Bordenave J, Thuillet R, Tu L, Phan C, Cumont A, Marsol C, Huertas A, Savale L, Hibert M, Galzi JL, et al. Neutralization of CXCL12 attenuates established pulmonary hypertension in rats. Cardiovasc Res. 2020;116:686–97.
Sweatt AJ, Hedlin HK, Balasubramanian V, Hsi A, Blum LK, Robinson WH, Haddad F, Hickey PM, Condliffe R, Lawrie A, et al. Discovery of distinct Immune Phenotypes using machine learning in pulmonary arterial hypertension. Circ Res. 2019;124:904–19.
Zaloudikova M, Vytasek R, Raskova M, Vizek M, Uhlik J, Hampl V. The effect of exposure to hypoxia on superoxide formation by alveolar macrophages is indirect. Life Sci. 2019;236:116864.
Li J, Rao J, Liu Y, Cao Y, Zhang Y, Zhang Q, Zhu D. 15-Lipoxygenase promotes chronic hypoxia-induced pulmonary artery inflammation via positive interaction with nuclear factor-kappab. Arterioscler Thromb Vasc Biol. 2013;33:971–9.
Aldabbous L, Abdul-Salam V, McKinnon T, Duluc L, Pepke-Zaba J, Southwood M, Ainscough AJ, Hadinnapola C, Wilkins MR, Toshner M, Wojciak-Stothard B. Neutrophil Extracellular traps promote angiogenesis: evidence from Vascular Pathology in Pulmonary Hypertension. Arterioscler Thromb Vasc Biol. 2016;36:2078–87.
Xie SS, Deng Y, Guo SL, Li JQ, Zhou YC, Liao J, Wu DD, Lan WF. Endothelial cell ferroptosis mediates monocrotaline-induced pulmonary hypertension in rats by modulating NLRP3 inflammasome activation. Sci Rep. 2022;12:3056.
Masaki T, Okazawa M, Asano R, Inagaki T, Ishibashi T, Yamagishi A, Umeki-Mizushima S, Nishimura M, Manabe Y, Ishibashi-Ueda H et al. Aryl hydrocarbon receptor is essential for the pathogenesis of pulmonary arterial hypertension. Proc Natl Acad Sci U S A 2021, 118.
Sawada H, Saito T, Nickel NP, Alastalo TP, Glotzbach JP, Chan R, Haghighat L, Fuchs G, Januszyk M, Cao A, et al. Reduced BMPR2 expression induces GM-CSF translation and macrophage recruitment in humans and mice to exacerbate pulmonary hypertension. J Exp Med. 2014;211:263–80.
Liao J, Xie SS, Deng Y, Wu DD, Meng H, Lan WF, Dai P. PRDX6-mediated pulmonary artery endothelial cell ferroptosis contributes to monocrotaline-induced pulmonary hypertension. Microvasc Res. 2023;146:104471.
Xu T, Zhang Z, Liu T, Zhang W, Liu J, Wang W, Wang J. Salusin-beta contributes to vascular inflammation associated with pulmonary arterial hypertension in rats. J Thorac Cardiovasc Surg. 2016;152:1177–87.
Marsboom G, Wietholt C, Haney CR, Toth PT, Ryan JJ, Morrow E, Thenappan T, Bache-Wiig P, Piao L, Paul J, et al. Lung (1)(8)F-fluorodeoxyglucose positron emission tomography for diagnosis and monitoring of pulmonary arterial hypertension. Am J Respir Crit Care Med. 2012;185:670–9.
Ostriker A, Horita HN, Poczobutt J, Weiser-Evans MC, Nemenoff RA. Vascular smooth muscle cell-derived transforming growth factor-beta promotes maturation of activated, neointima lesion-like macrophages. Arterioscler Thromb Vasc Biol. 2014;34:877–86.
Majesky MW, Horita H, Ostriker A, Lu S, Regan JN, Bagchi A, Dong XR, Poczobutt J, Nemenoff RA, Weiser-Evans MCM. Differentiated smooth muscle cells generate a subpopulation of Resident Vascular Progenitor cells in the Adventitia regulated by Klf4. Circul Res. 2017;120:296–311.
Sheikh AQ, Misra A, Rosas IO, Adams RH, Greif DM. Smooth muscle cell progenitors are primed to muscularize in pulmonary hypertension. Sci Transl Med. 2015;7:308ra159.
Dierick F, Héry T, Hoareau-Coudert B, Mougenot N, Monceau V, Claude C, Crisan M, Besson V, Dorfmüller P, Marodon G, et al. Resident PW1 + progenitor cells participate in vascular remodeling during Pulmonary arterial hypertension. Circul Res. 2016;118:822–33.
Jolly AJ, Lu S, Strand KA, Dubner AM, Mutryn MF, Nemenoff RA, Majesky MW, Moulton KS, Weiser-Evans MCM. Heterogeneous subpopulations of adventitial progenitor cells regulate vascular homeostasis and pathological vascular remodelling. Cardiovasc Res. 2022;118:1452–65.
Zhang H, Wang D, Li M, Plecita-Hlavata L, D’Alessandro A, Tauber J, Riddle S, Kumar S, Flockton A, McKeon BA, et al. Metabolic and proliferative state of vascular adventitial fibroblasts in pulmonary hypertension is regulated through a MicroRNA-124/PTBP1 (Polypyrimidine Tract Binding Protein 1)/Pyruvate kinase muscle Axis. Circulation. 2017;136:2468–85.
Fulton DJR, Li X, Bordan Z, Wang Y, Mahboubi K, Rudic RD, Haigh S, Chen F, Barman SA. Galectin-3: a harbinger of reactive oxygen species, fibrosis, and inflammation in pulmonary arterial hypertension. Antioxid Redox Signal. 2019;31:1053–69.
Kumar S, Frid MG, Zhang H, Li M, Riddle S, Brown RD, Yadav SC, Roy MK, Dzieciatkowska ME, D’Alessandro A et al. Complement-containing small extracellular vesicles from adventitial fibroblasts induce proinflammatory and metabolic reprogramming in macrophages. JCI Insight 2021, 6.
Gerasimovskaya E, Kratzer A, Sidiakova A, Salys J, Zamora M, Taraseviciene-Stewart L. Interplay of macrophages and T cells in the lung vasculature. Am J Physiol Lung Cell Mol Physiol. 2012;302:L1014–1022.
Ruopp NF, Cockrill BA. Diagnosis and treatment of pulmonary arterial hypertension: a review. JAMA. 2022;327:1379–91.
Barnes H, Brown Z, Burns A, Williams T. Phosphodiesterase 5 inhibitors for pulmonary hypertension. Cochrane Database Syst Rev. 2019;1:CD012621.
Shahin HI, Vinjamuri BP, Mahmoud AA, Shamma RN, Mansour SM, Ammar HO, Ghorab MM, Chougule MB, Chablani L. Design and evaluation of novel inhalable sildenafil citrate spray-dried microparticles for pulmonary arterial hypertension. J Control Release. 2019;302:126–39.
Barnes H, Yeoh HL, Fothergill T, Burns A, Humbert M, Williams T. Prostacyclin for pulmonary arterial hypertension. Cochrane Database Syst Rev. 2019;5:CD012785.
Liu C, Chen J, Gao Y, Deng B, Liu K. Endothelin receptor antagonists for pulmonary arterial hypertension. Cochrane Database Syst Rev. 2021;3:CD004434.
Wardle AJ, Seager MJ, Wardle R, Tulloh RM, Gibbs JS. Guanylate cyclase stimulators for pulmonary hypertension. Cochrane Database Syst Rev 2016, 2016:CD011205.
Favre S, Gambini E, Nigro P, Scopece A, Bianciardi P, Caretti A, Pompilio G, Corno AF, Vassalli G, von Segesser LK, et al. Sildenafil attenuates hypoxic pulmonary remodelling by inhibiting bone marrow progenitor cells. J Cell Mol Med. 2017;21:871–80.
Imai Y, Kariya T, Iwakiri M, Yamada Y, Takimoto E. Sildenafil ameliorates right ventricular early molecular derangement during left ventricular pressure overload. PLoS ONE. 2018;13:e0195528.
Noel S, Panin N, Beka M, Dhooghe B, Huaux F, Leal T. Vardenafil reduces macrophage pro-inflammatory overresponses in cystic fibrosis through PDE5- and CFTR-dependent mechanisms. Clin Sci (Lond). 2017;131:1107–21.
Flores-Costa R, Duran-Guell M, Casulleras M, Lopez-Vicario C, Alcaraz-Quiles J, Diaz A, Lozano JJ, Titos E, Hall K, Sarno R, et al. Stimulation of soluble guanylate cyclase exerts antiinflammatory actions in the liver through a VASP/NF-kappaB/NLRP3 inflammasome circuit. Proc Natl Acad Sci U S A. 2020;117:28263–74.
Gerlach K, Kohler-Bachmann S, Jungck D, Korber S, Yanik S, Knoop H, Wehde D, Rheinlander S, Walther JW, Kronsbein J, et al. Endothelin receptor-antagonists suppress lipopolysaccharide-induced cytokine release from alveolar macrophages of non-smokers, smokers and COPD subjects. Eur J Pharmacol. 2015;768:123–30.
Rafnsson A, Matic LP, Lengquist M, Mahdi A, Shemyakin A, Paulsson-Berne G, Hansson GK, Gabrielsen A, Hedin U, Yang J, Pernow J. Endothelin-1 increases expression and activity of arginase 2 via ETB receptors and is co-expressed with arginase 2 in human atherosclerotic plaques. Atherosclerosis. 2020;292:215–23.
Hartopo AB, Emoto N, Vignon-Zellweger N, Suzuki Y, Yagi K, Nakayama K, Hirata K. Endothelin-converting enzyme-1 gene ablation attenuates pulmonary fibrosis via CGRP-cAMP/EPAC1 pathway. Am J Respir Cell Mol Biol. 2013;48:465–76.
Schmid T, Brune B. Prostanoids and resolution of inflammation - beyond the lipid-mediator class switch. Front Immunol. 2021;12:714042.
Lewis A, Elks PM. Hypoxia induces macrophage tnfa expression via cyclooxygenase and prostaglandin E2 in vivo. Front Immunol. 2019;10:2321.
Uehata T, Akira S. mRNA degradation by the endoribonuclease Regnase-1/ZC3H12a/MCPIP-1. Biochim Biophys Acta. 2013;1829:708–13.
Tse KM, Vandenbon A, Cui X, Mino T, Uehata T, Yasuda K, Sato A, Tsujimura T, Hia F, Yoshinaga M et al. Enhancement of Regnase-1 expression with stem loop–targeting antisense oligonucleotides alleviates inflammatory diseases. Sci Transl Med 2022, 14.
Xi X, Liu S, Shi H, Yang M, Qi Y, Wang J, Du J. Serum–glucocorticoid regulated kinase 1 regulates macrophage recruitment and activation contributing to Monocrotaline-Induced Pulmonary arterial hypertension. Cardiovasc Toxicol. 2014;14:368–78.
Muratoglu SC, Belgrave S, Lillis AP, Migliorini M, Robinson S, Smith E, Zhang L, Strickland DK. Macrophage LRP1 suppresses neo-intima formation during vascular remodeling by modulating the TGF-beta signaling pathway. PLoS ONE. 2011;6:e28846.
Calvier L, Herz J, Hansmann G. Interplay of Low-Density Lipoprotein Receptors, LRPs, and lipoproteins in Pulmonary Hypertension. JACC: Basic Translational Sci. 2022;7:164–80.
Wang D, Zhang H, Li M, Frid MG, Flockton AR, McKeon BA, Yeager ME, Fini MA, Morrell NW, Pullamsetti SS, et al. MicroRNA-124 controls the proliferative, migratory, and inflammatory phenotype of pulmonary vascular fibroblasts. Circ Res. 2014;114:67–78.
Karoor V, Strassheim D, Sullivan T, Verin A, Umapathy NS, Dempsey EC, Frank DN, Stenmark KR, Gerasimovskaya E. The short-chain fatty acid butyrate attenuates pulmonary vascular remodeling and inflammation in Hypoxia-Induced Pulmonary Hypertension. Int J Mol Sci 2021, 22.
Joshi SR, Liu J, Bloom T, Karaca Atabay E, Kuo TH, Lee M, Belcheva E, Spaits M, Grenha R, Maguire MC, et al. Sotatercept analog suppresses inflammation to reverse experimental pulmonary arterial hypertension. Sci Rep. 2022;12:7803.
Qiu H, Zhang Y, Li Z, Jiang P, Guo S, He Y, Guo Y. Donepezil ameliorates pulmonary arterial hypertension by inhibiting M2-Macrophage activation. Front Cardiovasc Med. 2021;8:639541.
Mavrogiannis E, Hagdorn QAJ, Bazioti V, Douwes JM, Van Der Feen DE, Oberdorf-Maass SU, Westerterp M, Berger RMF. Pirfenidone ameliorates pulmonary arterial pressure and neointimal remodeling in experimental pulmonary arterial hypertension by suppressing NLRP3 inflammasome activation. Pulm Circ. 2022;12:e12101.
Li M, Riddle S, Zhang H, D’Alessandro A, Flockton A, Serkova NJ, Hansen KC, Moldovan R, McKeon BA, Frid M, et al. Metabolic reprogramming regulates the proliferative and inflammatory phenotype of adventitial fibroblasts in Pulmonary Hypertension through the Transcriptional Corepressor C-Terminal binding Protein-1. Circulation. 2016;134:1105–21.
Hudalla H, Michael Z, Christodoulou N, Willis GR, Fernandez-Gonzalez A, Filatava EJ, Dieffenbach P, Fredenburgh LE, Stearman RS, Geraci MW, et al. Carbonic anhydrase inhibition ameliorates inflammation and experimental pulmonary hypertension. Am J Respir Cell Mol Biol. 2019;61:512–24.
Kishimoto Y, Kato T, Ito M, Azuma Y, Fukasawa Y, Ohno K, Kojima S. Hydrogen ameliorates pulmonary hypertension in rats by anti-inflammatory and antioxidant effects. J Thorac Cardiovasc Surg. 2015;150:645–e654643.
Le Hiress M, Akagah B, Bernadat G, Tu L, Thuillet R, Huertas A, Phan C, Fadel E, Simonneau G, Humbert M, et al. Design, synthesis, and Biological Activity of New N-(Phenylmethyl)-benzoxazol-2-thiones as Macrophage Migration Inhibitory factor (MIF) antagonists: Efficacies in Experimental Pulmonary Hypertension. J Med Chem. 2018;61:2725–36.
Cheng Y, Gong Y, Qian S, Mou Y, Li H, Chen X, Kong H, Xie W, Wang H, Zhang Y, Huang Z. Identification of a Novel hybridization from Isosorbide 5-Mononitrate and Bardoxolone Methyl with Dual activities of Pulmonary Vasodilation and vascular remodeling inhibition on pulmonary arterial hypertension rats. J Med Chem. 2018;61:1474–82.
Guihaire J, Deuse T, Wang D, Spin JM, Blankenberg FG, Fadel E, Reichenspurner H, Schrepfer S. Immunomodulation Therapy using Tolerogenic macrophages in a Rodent Model of Pulmonary Hypertension. Stem Cells Dev. 2021;30:515–25.
Umezu R, Koga JI, Matoba T, Katsuki S, Wang L, Hasuzawa N, Nomura M, Tsutsui H, Egashira K. Macrophage (Drp1) dynamin-related protein 1 accelerates Intimal Thickening after Vascular Injury. Arterioscler Thromb Vasc Biol. 2020;40:e214–26.
Muraki Y, Yamasaki M, Takeuchi H, Tohyama K, Sano N, Matsuo T. Fluorescent Imaging Analysis for Distribution of Fluorescent Dye labeled- or encapsulated-liposome in Monocrotaline-Induced Pulmonary Hypertension Model Rat. Chem Pharm Bull (Tokyo). 2018;66:270–6.
Kimura S, Egashira K, Chen L, Nakano K, Iwata E, Miyagawa M, Tsujimoto H, Hara K, Morishita R, Sueishi K, et al. Nanoparticle-mediated delivery of nuclear factor kappaB decoy into lungs ameliorates monocrotaline-induced pulmonary arterial hypertension. Hypertension. 2009;53:877–83.
Mangione MC, Wen J, Cao DJ. Mechanistic target of rapamycin in regulating macrophage function in inflammatory cardiovascular diseases. J Mol Cell Cardiol. 2024;186:111–24.
Day EA, Townsend LK, Rehal S, Batchuluun B, Wang D, Morrow MR, Lu R, Lundenberg L, Lu JH, Desjardins EM et al. Macrophage AMPK β1 activation by PF-06409577 reduces the inflammatory response, cholesterol synthesis, and atherosclerosis in mice. iScience 2023, 26.
Qiu S, Fu X, Shi Y, Zang H, Zhao Y, Qin Z, Lin G, Zhao X. Relaxin-Loaded Inhaled Porous microspheres inhibit idiopathic pulmonary fibrosis and improve pulmonary function Post-bleomycin challenges. Mol Pharm. 2023;20:3947–59.
Lee T-M, Chang N-C, Lin S-Z. Dapagliflozin, a selective SGLT2 inhibitor, attenuated cardiac fibrosis by regulating the macrophage polarization via STAT3 signaling in infarcted rat hearts. Free Radic Biol Med. 2017;104:298–310.
Pich-Bavastro C, Yerly L, Di Domizio J, Tissot-Renaud S, Gilliet M, Kuonen F. Activin A–Mediated polarization of Cancer-Associated fibroblasts and macrophages confers resistance to checkpoint immunotherapy in skin Cancer. Clin Cancer Res. 2023;29:3498–513.
Yang P, Manaenko A, Xu F, Miao L, Wang G, Hu X, Guo Z-N, Hu Q, Hartman RE, Pearce WJ, et al. Role of PDGF-D and PDGFR-β in neuroinflammation in experimental ICH mice model. Exp Neurol. 2016;283:157–64.
Park SG, Kim HJ, Min YH, Choi E-C, Shin YK, Park B-J, Lee SW, Kim S. Human lysyl-tRNA synthetase is secreted to trigger proinflammatory response. Proceedings of the National Academy of Sciences 2005, 102:6356–6361.
Morty RE, Nagai H, Kuwahira I, Schwenke DO, Tsuchimochi H, Nara A, Ogura S, Sonobe T, Inagaki T, Fujii Y et al. Pulmonary macrophages Attenuate Hypoxic Pulmonary Vasoconstriction via β3AR/iNOS pathway in rats exposed to chronic intermittent hypoxia. PLoS ONE 2015, 10.
Acknowledgements
Not applicable.
Funding
This work was supported by grants from the National Natural Science Foundation of China (No. 81770095, 81700093, and 8210082163), the Fundamental Research Funds for the Central Universities (No.2042021kf0081) and the Science Fund for Creative Research Groups of the Natural Science Foundation of Hubei Province (No.2020CFA027).
Author information
Authors and Affiliations
Contributions
YZ, BL and MG wrote the main manuscript text, RX and RH provided significant input. NL and QG reviewed and edited the manuscript and approved the final version. All authors have read and approved the manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Zuo, Y., Li, B., Gao, M. et al. Novel insights and new therapeutic potentials for macrophages in pulmonary hypertension. Respir Res 25, 147 (2024). https://doi.org/10.1186/s12931-024-02772-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12931-024-02772-8