
Abstract
Mesenchymal stem cells (MSCs) have been demonstrated to have a great potential in the treatment of several diseases due to their differentiation and immunomodulatory capabilities and their ability to be easily cultured and manipulated. Recent investigations revealed that their therapeutic effect is largely mediated by the secretion of paracrine factors including exosomes. Exosomes reflect biophysical features of MSCs and are considered more effective than MSCs themselves. Alternative approaches based on MSC-derived exosomes can offer appreciable promise in overcoming the limitations and practical challenges observed in cell-based therapy. Furthermore, MSC-derived exosomes may provide a potent therapeutic strategy for various diseases and are promising candidates for cell-based and cell-free regenerative medicine. This review briefly summarizes the development of MSCs as a treatment for human diseases as well as describes our current knowledge about exosomes: their biogenesis and molecular composition, and how they exert their effects on target cells. Particularly, the therapeutic potential of MSC-derived exosomes in experimental models and recent clinical trials to evaluate their safety and efficacy are summarized in this study. Overall, this paper provides a current overview of exosomes as a new cell-free therapeutic agent.
Similar content being viewed by others

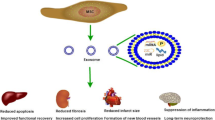
Background
Nowadays, multipotent mesenchymal stem cells (MSCs) have been extensively examined because of their usage in clinical trials. Their effective influence in cellular therapy and regenerative medicine is known for their strong immunosuppressive, immunomodulatory and regenerative activity [1, 2]. In addition, their considerable potential was demonstrated in the treatment of immune-mediated, inflammatory and degenerative diseases [3,4,5,6,7,8,9].
MSCs generally are multipotent, somatic progenitor/stem cells first isolated from adult bone marrow [10, 11] and successfully differentiated from marrow hematopoietic cells according to their adherent nature in in vitro cell lines and fibroblastic morphology. They are able to self-recover and retain variable differentiation potency toward multi-lineages [12, 13]. The International Society for Cellular Therapy has officialy defined minimal criteria for MSCs, following as (a) being plastic-adherent cells, (b) having adipogenic, osteogenic and chondrogenic trilineage mesenchymal differentiation capacity and (c) being positive (> 95%) for surface antigens CD73, CD90 and CD105 and negative (< 2%) for hematopoietic markers CD34, CD45, CD14 or CD11b, CD79α or CD19 and HLA-DR (typical markers of hematopoietic cells) [14]. Human MSCs were described in many tissues (Fig. 1), not only in those of mesodermal origin (bone marrow, bone, adipose, synovial membrane and muscle) but also in skin, heart, lungs, brain, kidneys, thymus, liver and pancreas [14, 15]. Another excellent sources of human MCSs are umbilical cord tissue and placenta [16,17,18]. However, it was revealed that MSCs obtained from various tissues have differences in gene expression, proliferation activity and differentiation potencial. In addition, some variations in surface antigens expression compared to requirements of minimal criteria were reported. Existing variances indicate specific features of MSCs from different tissues and organs or are related with isolation and cultivation protocols [19]. MSCs from different tissues can be cultured prior to clinical use. They can grow easily in the culture dish which leads to an easy manipulation in terms of isolation and cultivation. Subsequently, prepared MSCs suspensions may be introduced intravenously or trough local injection to obtain the required therapeutic effects directly or indirectly [20]. Further characteristics include typical plasticity, intrinsic tropism towards injured or inflammed area (known as homing) and an extensive release of numerous useful growth factors, cytokines and another bioactive soluble factors as important indication of their potential clinical applications in tissue repair and regeneration [21].

The most common sources of MSC isolation. (Created with BioRender.com.)
There is an evidence of tissue alteration by MSCs through secretion of paracrine factors contained in extracellular vesicles (EVs). EVs are a group of cell-derived structures (composed of lipid bilayer membranes), which play an essential role in intercellular communication via transfer of bioactive proteins, lipids and RNAs and represent a potential source for circulating biomarkers of diseases [22]. EVs are generally divided, depending on their biogenesis, into subgroups, like exosomes (40–150 nm in diameters), microvesicles (150–1000 nm in diameter) and apoptotic bodies (50–2000 nm in diameter) [23]. Recent studies suggest possible substitution of the biological MSCs activity with MSC-derived exosomes [24,25,26]. Therefore, exosomes could represent a considerable alternative to cell therapy.
This review is focused on the characterization of MSCs-derived exosomes and their perspective using in cell-free therapeutic applications, as well as on the summarization of important facts about general MSCs´ paracrine secretion.
Paracrine secretion of MSCs
MSCs perform their immunomodulatory activity not only through cell–cell interactions but also via strong paracrine impact. The MSCs´ paracrine effect was firstly described by Heynesworth et al. They notified secretion of a large spectrum of cytokines, chemokines and growth factors by MSCs with possible significant effects on cells in their periphery [27]. However, precise mechanism of action is still unknown and under examination. Numerous studies confirmed that factors secreted by MSCs could regenerate injured myocardium and improve cardiac function in porcine model [28], ameliorate acute renal failure and protect against limb tissue injury [29], promote in vitro and in vivo arteriogenesis [30] or support neovascularization [31].
One of the main pattern representing MSCs secretion of biological factors is by EVs which are classified as membrane vesicles filled with plenty of different proteins, microRNAs or/and messenger RNAs and have been progressively studied as the therapeutic agent in MSCs secretion [32]. The lipid bilayer of EVs encloses their bioactive capacity and protects them from enzymatic degradation. EVs are nowadays defined by their size, sedimentation rate, biogenesis pathway or protein delivery, but most of these parameters are neither terminal nor specific for any of EVs type. They have different structural and biochemical properties depending on their intracellular site of origin, which can affect their given functions [33]. Regardless of their origin, EVs are circular membrane particles possesing the characteristics of the origin cells, containing cytosol. In regard to their intracellular origin and the mechanisms of formation, EVs may be classified as exosomes, microvesicles and apoptotic bodies [23].
Apoptotic bodies are released as products of an appoptotic cell disassembly into subcellular fragments. There is an evidence that EVs generated during apoptosis have an important immunoregulatory role in autoimmunity, infection and cancer [34]. Microvesicles, also called as ectosomes or shedding vesicles, represent a heterogenous population formed by external budding and cleavage of the cell membrane. There is a large volume of phosphatidylserine on their surface and great number of proteins associated with lipid rafts (cholesterol-rich microdomains). Assembling of microvesicles is related to an increase of calcium ions which by calpain activation supports the cytoskeleton reorganization leading to the separation of plasma membrane protrusion from the cortical actin [35, 36]. Microvesicles may contain several plasma proteins depending on the type of the cell they originated and therefore specific markers are required for their identification. The generic marker is Anexin V. CD45 is used to identify leukocyte-derived microvesicles, CD42b/CD31− and CD62P for plateled-derived microvesicles, and CD31+ /CD42−, CD62E and CD144 are used for characterization of endothelial-derived microvesicles [37]. In addition, microvesicles may contain selectins, integrins, metalloproteinases and CD40 ligand [38]. On the other hand, exosomes are smaller and homogenous, have an endosomal origin and are formed by the internal budding of the multivesicular body membrane. The mechanism of their assembling and separation is still unknown [31]. Lipid bilayer of exosomes contains sphingomyelin, phosphatidylserine, phosphatidylcholine, phosphatidylethanolamine, phosphatidylinositol and monosialotetrahex-osylganglioside, which are similar to the cell plasma membrane composition [39]. Considered markers of exosomes are tetraspanins (CD9, CD63, CD81 and CD82), TSG101 (tumour susceptibility gene 101), heat shock proteins HSP70 and HSP90 and ALIX [39].
In general, it was shown that EVs are able to effectively copy the therapeutic effect of MSCs, mainly in tissue repair and regeneration in some preclinical models, e.g. exosomes potentially applied in wound healing and cutaneous regeneration [40], human adult liver stem cells—derived microvesicles increased hepatocyte proliferation associated with an accelerated morphological and functional recovery in a rat model [41] or human bone marrow MSCs—derived microvesicles increased proliferation and reduced apoptosis of tubular cells in a mice model [42].
Exosomes
Presently, the best characterized EVs are exosomes, which secretion into extracellular area by hematopoietic cells, more specifically by reticulocytes, was firstly described in late 1980s [43,44,45]. Initially, exosomes secreted from cells were considered as homeostasis secondary products or cellular waste from cell injury without any significant influence on cells nearby. Nowadays, exosomes are considered as a special agent of intracellular communication, playing a major role in cellular processes including immune response [46], antigen presentation [47] and signal transduction [48]. It was indicated that exosomes are produced and released by various types of healthy cells involving adipocytes, epithelial cells, fibrolasts, neurons, astrocytes and Schwann cells. In addition, they were found in numerous types of body fluids including cerebrospinal, synovial and amniotic fluid, urine, sperm, saliva, blood, ascites, vitreous and brest milk [49].
The biogenesis/formation and secretion of exosomes
In general, the biogenesis of exosomes begins within the endosomal system (Fig. 2) during which early endosomes (generated by internal budding) are unfolded into the late endosomes or multivesicular bodies (MVBs) and the endosomal membrane is invaginated to form intraluminal vesicles (ILVs) in the lumen of the organelles. The MVBs can either fuse with lysosomes to degrade their content or fuse with the plasma membrane to secrete the volume of ILVs as exosomes [50].

Schematic characterization of EVs (exosomes and microvesicles) formation. (Created with BioRender.com.)
The endosomal sorting complex required for transport (ESCRT) machinery is very important for the MVBs/ILVs formation, vesicle budding and protein cargo sorting [51]. Ubiquitin is a relevant signal agent that transports membrane proteins and/or damaged cellular elements to lysosomes for degradation. It is also known as signal molecule for exosomal cargo sorting on the endosome membrane [52]. ESCRT machinery is composed from four multiprotein complexes, namely ESCRT-0, -I, -II, -III and the associated AAA ATPase VPS4 complex [51]. Separation of proteins to MVBs includes segregation of the ubiquitinated proteins into lipid rafts with ESCRT-0. TSG101 (ESCRT-I protein) is able to bind to ubiquitinated cargo proteins and sorts endocytic ubiquitinated cargos into MVBs. Subsequently, ESCRT-II complex is activated, which starts the oligomerization and production of the ESCRT-III complex. This complex is involved in proceeding of the budding process responsible for the sequestration of MVBs proteins, sends the deubiquitinating enzyme to remove the ubiquitin label from the cargo proteins and then sorts them into ILVs. Finally, ESCRT-III complex is separated from MVB membrane by sorting protein VPS4 and is unfolded by ATPase [53, 54]. The precise role of ESCRT machinery in the generating of ILVs secreted later as exosomes is still unclear. In the screening study of RNA interference targeting ESCRT associated proteins in HeLa cells was shown that the depletion of Hrs, TSG101 and STAM1 proteins can reduce the exosomes secretion [55]. It was examined by nanoparticle tracking analysis that knockdown of Hrs reduced exosome secretion from head and neck squamous cell carcinoma cells [56]. Likewise, exosome secretion was increased by knockdown of the ESCRT-III and associated proteins ALIX, VTA1, VPS4B and CHMP4C [55]. Specifically, increase of exosomal level and typical exosomal markers (CD63, HSP70) was confirmed after syndecan – syntenin – ALIX depletion in MCF-7 cells [57].
Alternatively, sorting of exosomal cargo into MVBs and following ILVs formation can occur via ESCRT independent mechanism. Proteolipid protein containing exosomes requires for their secretion ceramide which is able to initiate the exosome budding into MVBs [58]. Expression of tetraspanins (transmembrane proteins rich in exosomes) CD9 and CD82 increased the exosomal release of β-catenin (involved in regulation and organization of cell–cell adhesion and gene transcription) from HEK293 cells [59]. The oligomerization of oligomers could play a significant role in exosome biogenesis based on CD43 exosomal sorting in Jurkat T-cells [60]. Observably, there are various possible mechanisms for separation of bioactive molecules into exosomes, either ESCRT dependent or independent, allow to work depending on the cell type and/or cellular homeostasis. In addition, it was shown that numerous diseases and other pathological conditions enhance exosome secretion. Increased quantity of exosomes were noticed in tumor cells, by progression of inflammation, angiogenesis and coagulation [61,62,63].
Molecular composition of exosomes
The molecular structure of exosomes is related not only to the cell type of origin but also to the microenvironment involving mechanical properties, biochemical impulses and topography, which could influence protein cargo regulation of the secreted exosomes [39]. Exosome secretion and their composition can also be modulated by other environmental factors such as oxygen level, type of disease, mechanical stress or media composition [64].
Exosomes are composed of various macromolecules involving unique lipid and protein structures and nucleic acids (Fig. 3). Exosomes are characterized by abundant amount of miRNAs with majority in the form of pre-miRNAs, which are inactive until their conversion to mature miRNAs [65]. Considering the endosomal origin of exosomes, they contain proteins participating in membrane transport and fusion (e.g. annexins, Rab, flotillin, GTPases), MVBs biogenesis (e.g. ALIX, TSG101) and also proteins associated with lipid microdomains (integrins and tetraspanins). Besides that, another frequently determinated proteins are associated with cytoskeleton (e.g. tubulin, myosin, actin) and metabolism (e.g. GADPH) [54], heat shock proteins (HSC70, HSC90), tissue specific proteins (e.g. MHC II located on the surface of exosomes secreted by dendritic cells or by B-lymphocytes) or proteins specific for cancer cell lines (e.g. glioma EGFR, breast cancer HER2, ovarian cancer CD24) [66].

Exosome´s composition briefing: MHC I, II—major histocompatibility complex I, II); MFGE8 -milk fat globule EGF factor 8 protein; ICAM-1—intercellular adhesion molecule 1; LAMP 1, 2—lysosomal-associated membrane protein 1, 2; proteins involved heat shock proteins (HSP60, HSP70, HSP90), MVB biogenesis proteins (Alix, TSG101, Ubiquitin, Clathrin), cytoskeleton proteins (profilin, cofilin, tubulin, actin, myosin, tropomyosin), signaling proteins (G protein, syntenin, MAPK, ERK ½, Rho); to enzymes belong pyruvate kinase, ATPase, PGK1, GADPH, aldolase, enolase; nucleic acids include mRNA, miRNA, siRNA, tRNA, DNA. (Created with BioRender.com.)
Specifically, numerous studies on the protein and RNA composition of MSC-derived exosomes have been reported. Lai et al. investigated the proteome of HPLC-purified human embryonic stem cells—derived exosomes using mass spectrometry and cytokine array. They identified more than 850 proteins and detected total protein complement of a 20S proteasome with very high reliability [67]. Kang´s group realized proteomic analysis of the nanoscale size-based fractionation of exosomes from human neural stem cells and identified 103 proteins. Results from their study confirmed, that exosomes larger than ∼50 nm were morphologically different from those which were smaller than ∼50 nm [68]. MSC-derived exosomes were found to contain also all five enzymes involved in the ATP synthesis of glycolysis, namely glyceraldehyde 3-phosphate dehydrogenase, phosphoglycerate kinase, phosphoglucomutase, enolase and pyruvate kinase m2 isoform [69]. Furthermore, Arslan´s group detected enzymatically active CD73 in MSC-derived exosomes responsible for the generating of extracellular adenosine from released adenine nucleotides [69]. Exosomes are able to activate adenosine receptors and thus generate adenosine-affected phosphorylation of ERK1/2 and Akt in H9C2 cardiomyocytes [70].
The genetic information in RNAs of exosomes which are endocytosed by acceptor cells is allow to influence the protein expression in those cells. Exosomes contain RNAs mostly in size range less than 700 nt. Chen et al. identified the presence of small RNAs (less than 30 nt) in human embryonic stem cells-derived MSCs´ conditioned medium, which were encapsulated in cholesterol rich phospholipid vesicles [65]. Plethora of miRNAs responsible for post-transcriptional maintainig of gene expression were detected in MSC-derived exosomes which are active in acceptor cells [71] and participate in physiological and pathological processes. Research group of Ratajczak et al. reported that embryonic stem cells – derived exosomes are highly enriched in mRNA (for numerous transcription factors, receptors and cytokines) [72]. Furthermore, Valadi et al. identified different miRNAs including let-7, miR-1, miR-15, miR-16, miR-181 and miR-375 in exosomes isolated from mast-cell line (MC/9), primary bone marrow-derived mast cells (BMMC) and human mast-cell line (HMC-1) [73], which have been suggested to play an important role in exocytosis, tumorigenesis, angiogenesis and haematopoiesis [74]. Ono et al. reported that miR-23b promotes dormancy in breast cancer cells [75]. Exosomal miRNAs derived from umbilical cord MSCs, mainly represented by let-7f, miR-145, miR-199a and miR-221 supported the suppression of hepatitis C virus RNA replication [76]. Results of several sequencing studies also demonstrated, that exosomes isolated from human blood serum and urine contain marked amount of other RNA types, such as tRNA, rRNA, snRNA snoRNA, piRNA and scaRNA [77].
The current studies of the structure and composition of exosomes have relevant importance and are still under examination. Wang et al. compared paracrine functions in vivo and exosomal profiles of human endometrium-, bone marrow- and adipose-derived MSCs in a rat model of myocardial infarction. Analyses of exosomal microRNAs showed that miR-21 expression was improved in exosomes derived from endometrium [78], suggesting that innate differences of various MSC-derived exosomes have substantial influence on their clinical efficacy. The importance of exosomes has long been recognized also due to their capability to transfer important cellular cargoes (proteins, DNA, mRNA, miRNAs) to target cells. Recent evidences suggest that exosomes are involved both in normal physiological functions and in pathological conditions. Deeper understanding of the exosomes content may influence the study of various diseases. Some research groups demonstrated that tetraspanin complexes significantly contributes to selective target binding of exosomes to target cells [79, 80]. Thakur et al. showed that the presence of dsDNA in exosomes represented the whole genomic DNA and could be used for identification of mutations in parental tumor cells. They determined that tumor-derived exosomes carry dsDNA and may be use as a circulating biomarker in the early detection of cancer and metastasis [81]. Liang et al. used engineered exosomes for co-delivery of chemotherapeutic drug 5-fluorouracil and chemoresistance miR-21 inhibitor oligonucleotide to reduce the drug resistance in colorectal carcinoma and thus to improve the efficacy of cancer treatment [82]. Yang et al. demonstrated the capability of brain endothelial cell-derived exosomes to deliver siRNA across the brain-blood barrier in zebrafish and thus inhibit VEGF [83]. Results suggested potential application of natural exosome vesicles in the treatment of brain disease [83]. Raposo et al. showed that both human and murine B-lymphocytes secrete exosomes to induce antigen-specific MHC (major histocompatibility complex) II-restricted T cell responses, reffering to exosome usefulness as biological instruments in immunotherapy [84].
The therapeutic effect and biodistribution of exosomes is also greatly affected by the origin of exosome producing cells. MSC-derived exosomes regarding to their inner properties and source of origin may play a relevant role in their clinical efficiency and represent an ideal delivery system for intermediate processes in specific target cells.
Therapeutic potential of MSC-derived exosomes
MSC-derived exosomes increasingly play an important role in intracellular communication mechanism and tissue repair and their clinical use may supply substantial advantages in comparison with their live cells due to potential to reduce undesirable side effects after application as well as infusional toxicities, uncontrolled cell growth and possible tumor formation. Moreover exosomes transplantation seems to be less risky and may have several advantages in contrast to cell applications. Exosomes are neither able to mutate and duplicate, nor induce metastasis. They have been tested in various animal models (Table 1) for human diseases (e.g. hypoxic pulmonary hypertension [85], acute kidney injury [86], liver fibrosis [87]) and it was detected that their functions are very similar to MSCs. First therapeutic potential of MSC-derived exosomes was described in a Langendorff heart model of acute myocardial ischemia/reperfusion injury in mice, where their cardioprotective effect was identified by myocardial infarct size reducing [88]. In this study, authors identified exosomes as cardioprotective elements in the MSCs´ paracrine secretion [88].
Several preclinical studies compared the beneficial effects of cell therapy based on MSCs and cell-free therapy based on MSC-derived EVs/exosomes and showed that they had similar therapeutic outcomes. Comparative analyses of MSCs and their EVs demonstrated different genetic cargo and protein content that play a significant role in biological processes, including angiogenesis, adipogenesis, apoptosis, regulation of inflammation, blood coagulation and extracellular matrix remodeling. Application of mice adipose MSCs in comparison with its conditioned medium had the same effect on sympotms of chronic colitis mouse model. Clinical symptoms and tissue damages were suppressed in treated mice [89]. Zhi et al. indicated that the application of umbilical cord MSC-derived exosomes (200 µg) resulted in amelioration of clinical symptoms, reduction of colonic damage and decrease of the inflammatory state in mice colitis when compared with MSCs (1 × 106 cells) administration [90]. Shao et al. compared activity of rat bone marrow MSCs and MSC-derived exosomes in a rat acute myocardial infarction model. It was showed a superior beneficial effects of MSC-derived exosomes in contrast to MSCs in cardiac repair. There were observed differences in expression profiles of several miRNAs from that of MSCs detected through miRNA sequence analysis [91]. A recent cutaneous wound model study in rabbits reported that intradermal injection of EVs derived from adipose and bone marrow MSCs were superior to MSCs injection in vivo. Furthermore, adipose MSC-derived EVs enhanced wound healing better than EVs from bone marrow [92]. In the study by Gatti et al. intravenous administration of human bone marrow MSC-derived EVs had the same efficacy as MSCs on the treatment of acute kidney injury in rats by inhibiting apoptosis and stimulating tubular cell proliferation [93]. In an induced experimental autoimmune encephalomyelitis murine model of multiple sclerosis, both human placental MSCs and its MSC-derived EVs showed regenerative effects and prevented oligodendroglia degradation and demyelination [94]. Another preclinical study showed that MSC-derived exosomes could be a promising cell-free therapeutic strategy for the treatment of Alzheimer’s disease. It was demonstrated that 28 days after intervention of mice groups with 10 μg exosomes and 1 × 106 MSCs separately had similar beneficial effects in improvement of neurogenesis and cognitive functions [95].
From the preclinical studies of MSC-derived exosomes therapy to the clinical application, many critical parameters should be resolved and determined, including clarification of important factors and conditions, defining optimal MSC culture conditions and protocols for precise monitoring of exosome formation, isolation, its characterization and storage. The biological effect of MCS-derived exosomes is mainly affected by the source of MSCs. The ideal source would be a high-exosome-yielding cell with a high expansion capacity [96, 97]. Further relevant requirement is the age of the donor tissue considering the exosome production might be indirectly connected with mentioned factor. Isolated exosomes are routinely identified by vesicle size and expression of typically tetraspanin markers CD63, CD9 and CD81. Production of exosomes could be enhanced by changing of several cell cultivation conditions, like increasing of intracellular calcium, or serum starvation. The long lasting donor HEK293 cell cultivation and maintaining cells at acidic pH could results in considerably increased production of exosomes [98]. Pre-conditioning of MSCs with hypoxia [99, 100], cytokines [101, 102] and another biomoleculs or chemicals (e.g. LPS [103], thrombin [104], NO [105], H2O2 [106]) also evoked the increase of exosomes activity, directly or indirectly by increasing MSCs function. Further important requirements for exosome preservation is an adequate storage. Sokolova et al. detected that the exosomes diameter decreased within 4 days at 4 °C and 2 days at 37 °C, indicating a structural change or degradation of exosomes, but storage at − 20 °C did not affect their size [107]. Extensive questions concerning of clinical grade exosomes production in sufficient quantity and of influence of different strategies on exosome potency are still under examination.
Bone marrow MSC-derived exosomes
Improvement of liver regeneration by BM MSC-derived exosomes
The potential of bone marrow (BM) MSC-derived exosomes for the treatment of various disease pathologies seems to be obvious. Rong et al. demonstrated the ability of human BM MSC-derived exosomes to reduce liver fibrosis in a carbon tetrachloride (CCl4)-induced liver fibrosis model of Sprague Dawley (SD) rats through the Wnt/β-catenin pathway. They also indicated the recovery of markers related to improved liver features, increasing hepatocyte regeneration and inhibition of inflammation process (significantly decreased inflammatory cytokines) [108]. Damania et al. studied the capability of rat BM MSC-derived exosomes present in fractionated MSC secretome to reduce liver injury in vitro in both 2D and 3D culture conditions of HepG2 cells and in in vivo rat models of acute liver injury caused by CCl4. Anti-apoptotic, anti-oxidative and prosurvival effects were shown in in vitro models of liver injury. In addition, the exosome rich fraction of conditioned media improved liver regeneration and recovery in vivo [109].
Cardioprotection by BM MSC-derived exosomes
The multiple therapeutic effects of BM MSC-derived exosomes have also been detected in cardiovascular, ischemic and reperfusion diseases. Currently, Chen et al. established significant protection of myocardium against hypertrophy, inhibition of myocardial apoptosis and reduction of cardiac fibrosis by using mice BM MSC-derived exosomes in the murine pressure overload induced cardiac hypertrophy model [110]. Teng et al. in their study hypothesized about a significant role of rat BM MSC-derived exosomes in the cardioprotection through angiogenesis and anti-inflammation in SD rats with acute myocardial infarction. They shown an efficacious action of exosomes in cardiac remodeling post-myocardial infarction in vivo. Accordingly, obtained results indicated that exosomes supported angiogesesis in vitro in human umbilical vein endothelial cell line (HUVEC). Furthermore, the proliferation of CD3 stimulated T-cells was reduced after exosome treatment, which means decrease in proliferation of spleen lymphocytes [111]. The rat myoblast cell line H9c2 was used to study myocardial pathogenic processes as cellular hypoxia‑reoxygenation model. Inhibition effect of cell proliferation, migration and also of suppresion of cardiomyocyte apoptosis during hypoxia-reoxygenation was revealed after rat BM MSC-derived exosome treatment [112]. In addition, quantity of both apoptosis- and autophagy-competent functional proteins and Apaf1 (apoptotic protease activating factor 1) and ATG13 (autophagy-related protein 13) gene expression in these treated H9c2 cells exhibited modulations in accordance with SD rat myocardial ischemia/ reperfusion model. Apaf1 expression was considerably suppressed and ATG13 expression was significantly increased in vivo after exosome treatment. Authors concluded, that myocardial injury associated with myocardial infarction could be inhibited with BM MSC-derived exosomes, alternatively throught regulation of autophagy mechanism [112].
BM MSC-derived exosomes and recovery after stroke and traumatic brain injury
In a stroke model (middle cerebral artery occlusion model) in Wistar rats, Xin et al. indicated that systemic administration of rat BM MSC-derived exosomes significantly enhanced functional recovery and improved neurite remodeling, neurogenesis and angiogenesis [113]. Therefore, exosomes could be effectively used for stroke treatment. Zhang et al. used human BM MSC-derived exosomes for the treatment of experimental traumatic brain injury in controlled cortical impact model in Wistar rats. Similarly, the improvement of functional recovery and promotion of neurovascular remodeling were demonstrated [114]. Administration of BM MSC-derived exosomes could regenerate cognition functions and memory impairment in neurological and neurodegenerative diseases. Exosomes derived from MSCs preconditioned by hypoxia supressed amyloid β accumulation and enhanced the synaptic protein expression in the brains of transgenic APP/PS1 mice (Alzheimer´s disease mice). Furthermore, reduced activation of astrocytes and microglia and changes in levels of inflammatory factors (increase of anti-inflammatoty cytokines IL-4, IL-10 and decrease of pro-inflammatory cytokines TNFα and IL-1β) were observed [115].
Anti-inflammation mediated by BM MSC-derived exosomes
Another promising therapeutic feature of porcine BM MSC-derived exosomes was evaluated by Casado et al. They showed the anti-inflammatory effect of exosomes in porcine model (large white pigs) of antigen-triggered synovitis. The local inflammation in animals caused by intra-articular injection of BSA leads to an elevated level of white blood cells in synovial fluid. Interestingly, there were found no differences of white blood cells in joints after exosome administration, but significant decrease in the lymphocytes accompanied by a noteworthy decline of only one (TNFα) from eight tested inflammatory cytokines in synovial fluid was revealed [116]. It is interesting, that TNFα antagonists (e.g. infliximab, golimumab, etanercept) are generally used for the treatment of rheumatoid arthritis [117].
Influence of BM MSC-derived exosomes on bone regeneration
Osteogenically differentiated human BM MSCs and subsequently derived EVs were used in the study of Martins et al.. They demonstrated, that human BM MSC-derived vesicles have osteoinductive potential characterized by early activation of alkaline phosphatase, early overexpression of the activator bone morphogenetic protein 2, transient increase in expression of Sp7 transcription factor (osterix) and secretion of phosphoprotein 1 (osteopontin) and integrin-binding sialoprotein (bone sialoprotein) [118]. Qin et al. tested the BM MSC-derived exosomes in the regulation of osteoblast activity in vitro and bone regeneration in vivo. Osteoblasts treated with miR-196a exhibited the best osteogenic activity in comparison with miR-27a and miR-206 treatment. Mentioned miRNAs are typical osteogenic related RNAs and are highly enriched in BM MSC-derived exosomes [119]. They also generated calvarial bone defects in SD rats and then applied hydrogel with EVs, which resulted in accelerated bone regeneration and indicated obvious improvement of the defect repair in comparison with hydrogel without EVs [119]. Narayanan et al. confirmed that exosomes from osteogenic human marrow MSCs are able to trigger lineage-specific differentiation of undifferentiated human BM MSCs [120]. Moreover, Shimbo et al. showed that the introduction of synthetic miR-143 into BM MSCs leads to an increase in not only the extracellular miR-143 but also increased secretion of exosomes. Such exosome-formed miR-143 was transferred to human osteosarcoma cell line 143B and caused suppression of their migration. It seems, that BM MSC-derived exosomes are also able to act as effective delivery system [121].
Umbilical cord MSC-derived exosomes
Reduction of liver fibrosis and liver injury by UC MSC-derived exosomes
Umbilical cord (UC) MSCs and their exosomes have also extensive potential in regenerative medicine, but their fundamental mechanism of action is still unknown. Li et al. used UC MSC-derived exosomes to treat CCl4-induced mouse liver fibrosis on Kunmingbai strains mice. It was shown that transplantation of human UC MSC-derived exosomes caused successful decrease of the serum fibrotic marker hyaluronic acid, TGF-β1 and serum aspartate aminotransferase and reduced hepatic inflammation and collagen deposition. Entire improvement after liver injury was confirmed [87]. Likewise, Jiang et al. identified hepatoprotective activities of human UC MSC-derived exosomes throught antioxidant defenses in mouse models (BALB/c female mice) of acute and chronic liver injury and liver tumor induced by CCl4 injection. They detected suppression of the liver tumor development, inhibition of oxidative stress in liver tumor, reduction of oxidative stress, inhibition of apoptosis in liver fibrosis and accordingly, reduction of oxidative stress and inhibition of apoptosis in acute liver injury after human UC MSC-derived exosome administration [71]. Shao et al. described large production of miR-455-3p enriched exosomes by human UC MSCs and their ability to suppress macrofage activation and reduce acute liver injury in mice model by inhibiting IL-6 signaling pathway [122].
Influence of UC MSC-derived exosomes on treatment of kidney injury
Furthermore, Zhou et al. studied the influence of human UC MSC-derived exosomes in SD rat model of kidney injury induced by cisplatin and in rat NRK-52E cells treated with or without cisplatin in vitro. There was indicated activation of the p38MAPK pathway followed by the increase of caspase 3 in NRK-52E cells after cisplatina treatment. Increase of apoptotic cells and oxidative stress were also observed. By contrast, these parameters were significantly reduced after human UC MSC-derived exosome administration. Accordingly, human UC MSC-derived exosomes moderated tubular oxidative damage, suppressed renal cell apoptosis and promoted renal cell proliferation in vivo in rats [123]. The major reason of acute kidney injury is ischemia/reperfusion injury in hospitalized patients. Therefore, Zou et al. in other study showed, that a single intravenous administration of human UC MSC-derived exosomes in rats with acute kidney injury induced by ischemia/reperfusion injury elevated renal capillary vessel density and alleviated renal fibrosis by increase of proangiogenic vascular endothelial growth factor (VEGF). In this process, reduction of HIF-1α (hypoxia inducible factor) was also observed. Exosomes were able to reduce cell apoptosis and improve proliferation after kidney injury [124].
Enhancement of fracture healing and wound healing by UC MSC-derived exosomes
Zhang et al. demonstrated intensive support of cutaneous wound healing and angiogenesis in vivo in a rat model of skin-deep second degree burn wound by human UC MSC-derived exosomes [125]. The Wnt signaling pathway plays an important role in angiogenesis mediated with the endothelial cell proliferation modulation, migration, vascular sprouting and remodeling, and vascular system maturation. UC-MSC-derived exosomes promote the tube formation, proliferation and migration of endothelial cells in vitro. In addition to that, applied exosomes improved angiogenesis by delivering Wnt4 to activation of Wnt/β-catenin in endothelial cells which could be one of the possible mechanism for tissue repair [125]. Likewise, Zhou et al. investigated the role of human UC MSC-derived exosomes in the Wnt signaling and their influence on femoral fracture healing in SD rats. Increase of β-catenin and Wnt3 expression indicating presumed participation of injected exosomes in repairing of the fracture was identified [126]. An important knowledge in this area is the study of Fang et al., in which they found that UC MSC-derived exosomes, especially exosomal microRNAs, decreased scar formation and myofibroblast accumulation in a skin-defect ICR mouse (Swiss-Hauschka mice) and nude mouse (BALB/c-υ) model. Myofibroblast formation can generally result in abnormal scarring. It was shown, that specific exosomal microRNAs (miR-21, miR-23a, miR-125b, and miR-145) inhibited redundant α-smooth muscle actin (α-SMA) and collagen I deposition and also suppressed TGF-β/SMAD2 signaling pathway [127].
UC MSC-derived exosomes relieve bowel diseases
UC MSC-derived exosomes have high potential in the treatment of inflammatory bowel diseases involving chronic inflammation of the gastrointestinal tract, both Crohn´s disease and ulcerative colitis in the future. Mao et al. demonstrated decrease of pro-inflammatory cytokines IL-6, IL-1β, TNF-α expression and increase of anti-inflammatory cytokine IL-10 expression after UC MSC-derived exosomes treatment in inflammatory bowel disease in a mice model. Interestingly, significant inhibition of IL-7 expression was also observed in the colon mucosa tissues and spleens in a mice model [128]. The serum cytokine level of IL-7 is normally increased in inflammatory bowel disease patients [129]. Similarly, single intraperitoneal injection of UC MSC-derived exosomes resulted in a significant reduction of the clinical symptoms and colonic damages in the mouse model of dextran sodium sulfate-induced colitis through suppression of inflammation mechanism [90].
Adipose MSC-derived exosomes
Attenuation of kidney inflammation by AD MSC-derived exosomes
Adipose (AD) MSC-derived exosomes as well as exosomes derived from BM and UC MSCs present a multipotent and rich therapeutic role in the improvement of the injury repair of many tissues. AD MSC-derived exosomes are more abundant and have lower risk of side effects. A single intrarenal delivery of pig AD MSC-derived exosomes in a porcine model of metabolic syndrome and renal artery stenosis resulted in reduction of renal inflammation, enhancement of the reparative macrophages number and elevation of anti-inflammatory cytokine IL-10 expression. Furthermore, exosome administration lowered renal vein level of pro-inflammatory cytokines TNF-α, IL-1β and IL-6 [130]. Results in the study of Eirin et al., established attenuation of renal fibrosis and improvement of stenotic kidney function after AD MSC-derived exosome treatment [130].
Cardioprotection by AD MSC-derived exosomes
It was observed, that AD MSC-derived exosomes are able to protect myocardium against acute ischemia/reperfusion induced necrosis and apoptosis in SD rat myocardial ischemia/reperfusion model. Ischemia/reperfusion injury in rats was accompanied with a remarkable decrease of Bcl-2 and an obvious increase in Bax expression. Both were eliminated after exosome administration. It was also observed, that AD MSC-derived exosomes attenuated hypoxia/reoxygenation induced apoptosis and promoted cell survival in H9c2 cell line [131]. In addition, Cui et al. hypothesized that AD MSC-derived exosome administration could protect ischemic myocardium through activation of Wnt/β-catenin signaling in vivo [131]. Liu et al. determined the protective influence of mouse AD MSC-derived exosomes on cardiomyocytes under oxidative stress in vitro [132].
Potential of AD MSC-derived exosomes for Alzheimer´s disease treatment
Interestingly, Katsuda et al. demonstrated remarkable potential of AD MSC-derived exosomes for Alzheimer´s disease therapy [133]. They showed that AD MSC-derived exosomes exhibited neprilysin specific enzyme activity. Neprilysin is the most essential enzyme that degrade amyloid beta peptide in the brain. In addition, transfer of mentioned exosomes to neuroblastoma N2a cells resulted in a decrease of both intracellular and extracellular amyloid beta peptide grades, suggesting a promising therapeutic approach for exosome-based Alzheimer´s disease treatment [133].
Role of AD MSC-derived exosomes in tumor progression
Recently, the influence of MSC-derived exosomes on tumor progression in both inhibiting and supporting mode was intensively described. Reza et al. indicated that human AD MSC-derived exosomal miRNAs have significant inhibitory influence on the regulation of different ovarian cancer cells [134]. Exosomes collected from human AD MSC-derived conditioned medium inhibited the growth and proliferation of ovarian cancer cells A2780 and SKOV-3. Decreased cell viability and wound healing of cancer cells were also observed after exosome treatment. Furthermore, collected exosomes caused apoptosis by increasing of pro-apoptotic signalling molecules Bax, caspase 3 and caspase 9 and by decreasing of anti-apoptotic bcl-2 protein [134].
Clinical perspectives
Clinical applications using exosome technology as cell-free therapy has become an important field of research over the last years. Currently, 91 clinical trials involving exosomes are listed on www.clinicaltrials.gov. Exosomes used in these trials are mainly derived from several body fluids and are used as early diagnostic tools in prediction of various diseases.
The clinical use of human MSC-derived exosomes is limited due to rigorous resolution of critical parameters involved in the translation process of preclinical studies to the clinical ones. These paramaters include the optimal MSC culture conditions and protocols for exosome production, isolation, and storage with a considerable effect on the uniformity of optimal dose, exosome administration and efficacy evaluation [2, 24]. Various approaches to optimize the therapeutic efficacy of exosomes are being developed. In general, the substantial requirement is a standardization of the classification and extraction method of exosomes from various body fluids, including definition of using of lower biofluid volume, higher purity and yield. Also the identification and better characterization of specific EVs subgroups is needed because different EVs could involve different biological effects. Whereas actual extraction methods of exosomes are too diverse for confirmation of its purity, it is necessary to standardize the protocols and characterization methods before application of exosomes in clinical trials. In addition, determination of the optimal dose, adequate time and appropriate method for exosome administration with maximal targeted efficacy, biological safety and without adverse effects must be confirmed before their clinical use.
Up to date, there are 15 clinical trials related to MSC-derived exosomes, registered on Clinicaltrials.gov, which are summarized in Table 2. Some of these studies have been completed or are recruiting/about to open to accrual. The trial NCT04491240 is focused on the evaluation of safe and effective method of MSC-derived exosomes aerosol inhalation in SARS-CoV-2 associated pneumonia and is only one trial which has been posted the results. Similar issue is performed in the completed pilot clinical trial NCT04276987 where the safety and efficiency of allogenic AD MSC-derived exosomes inhalation in the treatment of patients hospitalized with new coronavirus pneumonia is investigated. In completed trial NCT03562715 peripheral blood exosomes´ miRNA136, miRNA494 and miRNA495 genes expression in comparison to UC MSC conditioned media exosomes in patients with pre-eclampsia (pregnancy complication) was indentified. Based on the received and published data, MSC-derived exosomes are going to be great biological tools for diabetes, stroke, Alzheimer´s Disease and cancer therapy. Actually, it is hopeful to delve deeper into the potential of MSC-exosomes among SARS-CoV-2 pneumonia therapy and provide effective treatments with the highest safety.
Conclusion
MSCs mainly exert their therapeutic effects through the secretion of paracrine factors to reduce inflammation, cellular injury and enhance cell and tissue repair. MSC-derived exosomes probably work in a similar manner and have the capacity to interact with multiple cell types, enabling the cells to recover, repair and regenerate within the tissue. Due to their ability to deliver genetic material, immunomodulatory proteins, enzymes, and growth factors directly to the recipient cells, they also represent an ideal multifunctional delivery system. MSC-derived exosome therapy may be an emerging and a promising tool for the treatment of various diseases, mainly of those with an inflammatory component. Whats more, encouraging results of preclinical and clinical data predicted that MSC-derived exosome treatment could be superior to cell-based therapy in the meaning of safety and versatility.
Availability of data and materials
Not applicable.
Abbreviations
- MSCs:
-
Mesenchymal stem cells
- EVs:
-
Extracellular vesicles
- MVBs:
-
Multivesicular bodies
- ILVs:
-
Intraluminal vesicles
- BM:
-
Bone marrow
- UC:
-
Umbilical cord
- AD:
-
Adipose
- ESCRT:
-
The endosomal sorting complex required for transport
- SD rats:
-
Sprague Dawley rats
References
Galipeau J, Sensebe L. Mesenchymal stromal cells: clinical challenges and therapeutic opportunities. Cell Stem Cell. 2018;22(6):824–33.
Squillaro T, Peluso G, Galderisi U. Clinical trials with mesenchymal stem cells: an update. Cell Transplant. 2016;25(5):829–48.
Tanna T, Sachan V. Mesenchymal stem cells: potential in treatment of neurodegenerative diseases. Curr Stem Cell Res Ther. 2014;9(6):513–21.
Chen XD, Wang SJ, Cao W. Mesenchymal stem cell-mediated immunomodulation in cell therapy of neurodegenerative diseases. Cell Immunol. 2018;326:8–14.
Staff NP, Jones DT, Singer W. Mesenchymal stromal cell therapies for neurodegenerative diseases. Mayo Clin Proc. 2019;94(5):892–905.
Shi YF, Wang Y, Li Q, Liu KL, Hou JQ, Shao CS, et al. Immunoregulatory mechanisms of mesenchymal stem and stromal cells in inflammatory diseases. Nat Rev Nephrol. 2018;14(8):493–507.
Wang LT, Ting CH, Yen ML, Liu KJ, Sytwu HK, Wu KK, et al. Human mesenchymal stem cells (MSCs) for treatment towards immune- and inflammation-mediated diseases: review of current clinical trials. J Biomed Sci. 2016. https://doi.org/10.1186/s12929-016-0289-5.
Katuchova J, Harvanova D, Spakova T, Kalanin R, Farkas D, Durny P, et al. Mesenchymal stem cells in the treatment of Type 1 diabetes mellitus. Endocr Pathol. 2015;26(2):95–103.
Katuchova J, Tothova T, Iannaccone SF, Toporcer T, Harvanova D, Hildebrand T, et al. Impact of different pancreatic microenvironments on improvement in hyperglycemia and insulin deficiency in diabetic rats after transplantation of allogeneic mesenchymal stromal cells. J Surg Res. 2012;178(1):188–95.
Haynesworth SE, Goshima J, Goldberg VM, Caplan AI. Characterization of cells with osteogenic potential from human marrow. Bone. 1992;13(1):81–8.
Lazarus HM, Haynesworth SE, Gerson SL, Rosenthal NS, Caplan AI. Ex vivo expansion and subsequent infusion of human bone marrow-derived stromal progenitor cells (mesenchymal progenitor cells): implications for therapeutic use. Bone Marrow Transplant. 1995;16(4):557–64.
Pittenger MF, Mackay AM, Beck SC, Jaiswal RK, Douglas R, Mosca JD, et al. Multilineage potential of adult human mesenchymal stem cells. Science. 1999;284(5411):143–7.
Caplan AI. Mesenchymal stem cells. J Orthopaedic Res. 1991;9(5):641–50.
Dominici M, Le Blanc K, Mueller I, Slaper-Cortenbach I, Marini FC, Krause DS, et al. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy. 2006;8(4):315–7.
Harvanová D, Tóthová T, Sarišský M, Amrichová J, Rosocha J. Isolation and characterization of synovial mesenchymal stem cells. Folia Biol. 2011;57(3):119–24.
Romanov YA, Svintsitskaya VA, Smirnov VN. Searching for alternative sources of postnatal human mesenchymal stem cells: candidate MSC-like cells from umbilical cord. Stem Cells. 2003;21(1):105–10.
He S, Gleason J, Fik-Rymarkiewicz E, Difiglia A, Bharathan M, Morschauser A, et al. Human placenta-derived mesenchymal stromal-like cells enhance angiogenesis via T cell-dependent reprogramming of macrophage differentiation. Stem Cells. 2017;35(6):1603–13.
Bacenkova D, Rosocha J, Tothova T, Rosocha L, Sarissky M. Isolation and basic characterization of human term amnion and chorion mesenchymal stromal cells. Cytotherapy. 2011;13(9):1047–56.
Kern S, Eichler H, Stoeve J, Kluter H, Bieback K. Comparative analysis of mesenchymal stem cells from bone marrow, umbilical cord blood, or adipose tissue. Stem Cells. 2006;24(5):1294–301.
Fitzsimmons REB, Mazurek MS, Soos A, Simmons CA. Mesenchymal Stromal/Stem Cells in Regenerative Medicine and Tissue Engineering. Stem Cells International. 2018;2018.
Saeedi P, Halabian R, Imani Fooladi AA. A revealing review of mesenchymal stem cells therapy, clinical perspectives and Modification strategies. Stem cell Investigation. 2019;6:34.
Shao H, Im H. New technologies for analysis of extracellular vesicles. Chem Rev. 2018;118(4):1917–50.
Heldring N, Mager I, Wood MJA, Le Blanc K, Andaloussi SEL. Therapeutic potential of multipotent mesenchymal stromal cells and their extracellular vesicles. Hum Gene Ther. 2015;26(8):506–17.
Mendt M, Rezvani K, Shpall E. Mesenchymal stem cell-derived exosomes for clinical use. Bone Marrow Transplant. 2019;54:789–92.
Lou GH, Chen Z, Zheng M, Liu YN. Mesenchymal stem cell-derived exosomes as a new therapeutic strategy for liver diseases. Exp Mol Med. 2017;49:e346.
Liew LC, Katsuda T, Gailhouste L, Nakagama H, Ochiya T. Mesenchymal stem cell-derived extracellular vesicles: a glimmer of hope in treating Alzheimer’s disease. Int Immunol. 2017;29(1):11–9.
Haynesworth SE, Baber MA, Caplan AI. Cytokine expression by human marrow-derived mesenchymal progenitor cells in vitro: effects of dexamethasone and IL-1 alpha. J Cell Physiol. 1996;166(3):585–92.
Min JY, Sullivan MF, Yang Y, Zhang JP, Converso KL, Morgan JP, et al. Significant improvement of heart function by cotransplantation of human mesenchymal stem cells and fetal cardiomyocytes in postinfarcted pigs. Ann Thorac Surg. 2002;74(5):1568–75.
Patschan D, Plotkin M, Goligorsky MS. Therapeutic use of stem and endothelial progenitor cells in acute renal injury: ca ira. Curr Opin Pharmacol. 2006;6(2):176–83.
Kinnaird T, Stabile E, Burnett MS, Lee CW, Barr S, Fuchs S, et al. Marrow-derived stromal cells express genes encoding a broad spectrum of arteriogenic cytokines and promote in vitro and in vivo arteriogenesis through paracrine mechanisms. Circ Res. 2004;94(5):678–85.
Miyahara Y, Nagaya N, Kataoka M, Yanagawa B, Tanaka K, Hao H, et al. Monolayered mesenchymal stem cells repair scarred myocardium after myocardial infarction. Nat Med. 2006;12(4):459–65.
Akyurekli C, Le Y, Richardson RB, Fergusson D, Tay J, Allan DS. A systematic review of preclinical studies on the therapeutic potential of mesenchymal stromal cell-derived microvesicles. Stem Cell Rev Rep. 2015;11(1):150–60.
Thery C, Ostrowski M, Segura E. Membrane vesicles as conveyors of immune responses. Nat Rev Immunol. 2009;9(8):581–93.
Caruso S, Poon IKH. Apoptotic cell-derived extracellular vesicles: more than just debris. Front Immunol. 2018;9:1486.
Pap E, Pallinger E, Pasztoi M, Falus A. Highlights of a new type of intercellular communication: microvesicle-based information transfer. Inflamm Res. 2009;58(1):1–8.
Camussi G, Deregibus MC, Bruno S, Grange C, Fonsato V, Tetta C. Exosome/microvesicle-mediated epigenetic reprogramming of cells. Am J Cancer Res. 2011;1(1):98–110.
Jy W, Horstman LL, Jimenez JJ, Ahn YS. Measuring circulating cell-derived microparticles. J Thromb Haemost. 2004;2(10):1842–51.
Heijnen HFG, Schiel AE, Fijnheer R, Geuze HJ, Sixma JJ. Activated platelets release two types of membrane vesicles: microvesicles by surface shedding and exosomes derived from exocytosis of multivesicular bodies and alpha-granules. Blood. 1999;94(11):3791–9.
Bjorge IM, Kim SY, Mano JF, Kalionis B, Chrzanowski W. Extracellular vesicles, exosomes and shedding vesicles in regenerative medicine—a new paradigm for tissue repair. Biomater Sci. 2017;6(1):60–78.
Hu P, Yang QX, Wang Q, Shi CS, Wang DL, Armato U, et al. Mesenchymal stromal cells-exosomes: a promising cell-free therapeutic tool for wound healing and cutaneous regeneration (vol 7, 38, 2019). Burns & Trauma. 2020;8.
Herrera MB, Fonsato V, Gatti S, Deregibus MC, Sordi A, Cantarella D, et al. Human liver stem cell-derived microvesicles accelerate hepatic regeneration in hepatectomized rats. J Cell Mol Med. 2010;14(6B):1605–18.
Bruno S, Grange C, Deregibus MC, Calogero RA, Saviozzi S, Collino F, et al. Mesenchymal stem cell-derived microvesicles protect against acute tubular injury. J Am Soc Nephrol. 2009;20(5):1053–67.
Johnstone RM, Adam M, Hammond JR, Orr L, Turbide C. Vesicle formation during reticulocyte maturation—association of plasma-membrane activities with released vesicles (exosomes). J Biol Chem. 1987;262(19):9412–20.
Harding C, Heuser J, Stahl P. Receptor-mediated endocytosis of transferrin and recycling of the transferrin receptor in rat reticulocytes. J Cell Biol. 1983;97(2):329–39.
Pan BT, Teng K, Wu C, Adam M, Johnstone RM. Electron-microscopic evidence for externalization of the transferrin receptor in vesicular form in sheep reticulocytes. J Cell Biol. 1985;101(3):942–8.
Greening DW, Gopal SK, Xu R, Simpson RJ, Chen WS. Exosomes and their roles in immune regulation and cancer. Semin Cell Dev Biol. 2015;40:72–81.
Mittelbrunn M, Gutierrez-Vazquez C, Villarroya-Beltri C, Gonzalez S, Sanchez-Cabo F, Gonzalez MA, et al. Unidirectional transfer of microRNA-loaded exosomes from T cells to antigen-presenting cells. Nat Commun. 2011. https://doi.org/10.1038/ncomms1285.
Gangoda L, Boukouris S, Liem M, Kalra H, Mathivanan S. Extracellular vesicles including exosomes are mediators of signal transduction: are they protective or pathogenic? Proteomics. 2015;15(2–3):260–71.
Isola AL, Chen SZ. Exosomes: the messengers of health and disease. Curr Neuropharmacol. 2017;15(1):157–65.
Kalluri R, LeBleu VS. The biology, function, and biomedical applications of exosomes. Science. 2020;367(6478):640.
Henne WM, Stenmark H, Emr SD. Molecular mechanisms of the membrane sculpting ESCRT pathway. Cold Spring Harbor Perspect Biol. 2013;5(9):a016766.
Piper RC, Luzio JP. Ubiquitin-dependent sorting of integral membrane proteins for degradation in lysosomes. Curr Opin Cell Biol. 2007;19(4):459–65.
Zhang Y, Liu YF, Liu HY, Tang WH. Exosomes: biogenesis, biologic function and clinical potential. Cell Biosci. 2019. https://doi.org/10.1186/s13578-019-0282-2.
Urbanelli L, Magini A, Buratta S, Brozzi A, Sagini K, Polchi A, et al. Signaling pathways in exosomes biogenesis. Secretion Fate Genes. 2013;4(2):152–70.
Colombo M, Moita C, van Niel G, Kowal J, Vigneron J, Benaroch P, et al. Analysis of ESCRT functions in exosome biogenesis, composition and secretion highlights the heterogeneity of extracellular vesicles. J Cell Sci. 2013;126(24):5553–65.
Hoshino D, Kirkbride KC, Costello K, Clark ES, Sinha S, Grega-Larson N, et al. Exosome secretion is enhanced by invadopodia and drives invasive behavior. Cell Rep. 2013;5(5):1159–68.
Baietti MF, Zhang Z, Mortier E, Melchior A, Degeest G, Geeraerts A, et al. Syndecan-syntenin-ALIX regulates the biogenesis of exosomes. Nat Cell Biol. 2012;14(7):677–85.
Trajkovic K, Hsu C, Chiantia S, Rajendran L, Wenzel D, Wieland F, et al. Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science. 2008;319(5867):1244–7.
Chairoungdua A, Smith DL, Pochard P, Hull M, Caplan MJ. Exosome release of beta-catenin: a novel mechanism that antagonizes Wnt signaling. J Cell Biol. 2010;190(6):1079–91.
Fang Y, Wu N, Gan X, Yan WH, Morrell JC, Gould SJ. Higher-order oligomerization targets plasma membrane proteins and HIV gag to exosomes. PLoS Biol. 2007;5(6):1267–83.
Janowska-Wieczorek A, Wysoczynski M, Kijowski J, Marquez-Curtis L, Machalinski B, Ratajczak J, et al. Microvesicles derived from activated platelets induce metastasis and angiogenesis in lung cancer. Int J Cancer. 2005;113(5):752–60.
Ailawadi S, Wang XH, Gu HT, Fan GC. Pathologic function and therapeutic potential of exosomes in cardiovascular disease. BBA Mol Basis Dis. 2015;1852(1):1–11.
Mitchell PJ, Welton J, Staffurth J, Court J, Mason MD, Tabi Z, et al. Can urinary exosomes act as treatment response markers in prostate cancer? J Transl Med. 2009;7:4.
Atienzar-Aroca S, Flores-Bellver M, Serrano-Heras G, Martinez-Gil N, Barcia JM, Aparicio S, et al. Oxidative stress in retinal pigment epithelium cells increases exosome secretion and promotes angiogenesis in endothelial cells. J Cell Mol Med. 2016;20(8):1457–66.
Chen TS, Lai RC, Lee MM, Choo ABH, Lee CN, Lim SK. Mesenchymal stem cell secretes microparticles enriched in pre-microRNAs. Nucleic Acids Res. 2010;38(1):215–24.
Simpson RJ, Lim JWE, Moritz RL, Mathivanan S. Exosomes: proteomic insights and diagnostic potential. Expert Rev Proteomics. 2009;6(3):267–83.
Lai RC, Tan SS, Teh BJ, Sze SK, Arslan F, de Kleijn DP, et al. Proteolytic potential of the msc exosome proteome: implications for an exosome-mediated delivery of therapeutic proteasome. Int J Proteomics. 2012;2012:971907.
Kang DJ, Oh S, Ahn SM, Lee BH, Moon MH. Proteomic analysis of exosomes from human neural stem cells by flow field-flow fractionation and nanoflow liquid chromatography-tandem mass spectrometry. J Proteome Res. 2008;7(8):3475–80.
Arslan F, Lai RC, Smeets MB, Akeroyd L, Choo A, Aguor EN, et al. Mesenchymal stem cell-derived exosomes increase ATP levels, decrease oxidative stress and activate PI3K/Akt pathway to enhance myocardial viability and prevent adverse remodeling after myocardial ischemia/reperfusion injury. Stem Cell Res. 2013;10(3):301–12.
Li X, Arslan F, Ren Y, Adav SS, Poh KK, Sorokin V, et al. Metabolic adaptation to a disruption in oxygen supply during myocardial ischemia and reperfusion is underpinned by temporal and quantitative changes in the cardiac proteome. J Proteome Res. 2012;11(4):2331–46.
Deng H, Sun C, Sun YX, Li HH, Yang L, Wu DB, et al. Lipid, protein, and MicroRNA composition within mesenchymal stem cell-derived exosomes. Cell Reprogram. 2018;20(3):178–86.
Ratajczak J, Miekus K, Kucia M, Zhang J, Reca R, Dvorak P, et al. Embryonic stem cell-derived microvesicles reprogram hematopoietic progenitors: evidence for horizontal transfer of mRNA and protein delivery. Leukemia. 2006;20(5):847–56.
Valadi H, Ekstrom K, Bossios A, Sjostrand M, Lee JJ, Lotvall JO. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol. 2007;9(6):654-U72.
Alvarez-Garcia I, Miska EA. MicroRNA functions in animal development and human disease. Development. 2005;132(21):4653.
Ono M, Kosaka N, Tominaga N, Yoshioka Y, Takeshita F, Takahashi RU, et al. Exosomes from bone marrow mesenchymal stem cells contain a microRNA that promotes dormancy in metastatic breast cancer cells. Sci Signal. 2014;7(332):ra63.
Qian X, Xu C, Fang S, Zhao P, Wang Y, Liu H, et al. Exosomal MicroRNAs derived from umbilical mesenchymal stem cells inhibit Hepatitis C virus infection. Stem Cells Transl Med. 2016;5(9):1190–203.
Li M, Zeringer E, Barta T, Schageman J, Cheng AG, Vlassov AV. Analysis of the RNA content of the exosomes derived from blood serum and urine and its potential as biomarkers. Philos Trans R Soc B-Biol Sci. 2014;369(1652):20130502.
Wang K, Jiang Z, Webster KA, Chen JH, Hu HX, Zhou Y, et al. Enhanced cardioprotection by human endometrium mesenchymal stem cells driven by exosomal MicroRNA-21. Stem Cells Transl Med. 2017;6(1):209–22.
Rana S, Yue SJ, Stadel D, Zoller M. Toward tailored exosomes: the exosomal tetraspanin web contributes to target cell selection. Int J Biochem Cell Biol. 2012;44(9):1574–84.
Nazarenko I, Rana S, Baumann A, McAlear J, Hellwig A, Trendelenburg M, et al. Cell surface tetraspanin Tspan8 contributes to molecular pathways of exosome-induced endothelial cell activation. Can Res. 2010;70(4):1668–78.
Williams C, Rodriguez-Barrueco R, Silva JM, Zhang WJ, Hearn S, Elemento O, et al. Double-stranded DNA in exosomes: a novel biomarker in cancer detection. Cell Res. 2014;24(6):766–9.
Liang GF, Zhu YL, Ali DJ, Tian T, Xu HT, Si K, et al. Engineered exosomes for targeted co-delivery of miR-21 inhibitor and chemotherapeutics to reverse drug resistance in colon cancer. J Nanobiotechnol. 2020. https://doi.org/10.1186/s12951-019-0563-2.
Yang TZ, Fogarty B, LaForge B, Aziz S, Pham T, Lai LN, et al. Delivery of small interfering Rna to inhibit vascular endothelial growth factor in Zebrafish using natural brain endothelia cell-secreted exosome nanovesicles for the treatment of brain cancer. Aaps J. 2017;19(2):475–86.
Raposo G, Nijman HW, Stoorvogel W, Leijendekker R, Harding CV, Melief CJM, et al. B lymphocytes secrete antigen-presenting vesicles. J Exp Med. 1996;183(3):1161–72.
Lee C, Mitsialis SA, Aslam M, Vitali SH, Vergadi E, Konstantinou G, et al. Exosomes mediate the cytoprotective action of mesenchymal stromal cells on hypoxia-induced pulmonary hypertension. Circulation. 2012;126(22):2601.
Reis LA, Borges FT, Simões MJ, Borges AA, Sinigaglia-Coimbra R, Schor N. Bone marrow-derived mesenchymal stem cells repaired but did not prevent gentamicin-induced acute kidney injury through paracrine effects in rats. PLoS ONE. 2012;7(9):e44092.
Li TF, Yan YM, Wang BY, Qian H, Zhang X, Shen L, et al. Exosomes derived from human umbilical cord mesenchymal stem cells alleviate liver fibrosis. Stem Cells Dev. 2013;22(6):845–54.
Lai RC, Arslan F, Lee MM, Sze NSK, Choo A, Chen TS, et al. Exosome secreted by MSC reduces myocardial ischemia/reperfusion injury. Stem Cell Res. 2010;4(3):214–22.
Heidari M, Pouya S, Baghaei K, Aghdaei HA, Namaki S, Zali MR, et al. The immunomodulatory effects of adipose-derived mesenchymal stem cells and mesenchymal stem cells-conditioned medium in chronic colitis. J Cell Physiol. 2018;233(11):8754–66.
Jie Z, Wang YH, Li ZG, Wang Y, Li BY, Kang HY, et al. Immunosuppressive effect of exosomes from mesenchymal stromal cells in defined medium on experimental colitis. Int J Stem Cells. 2019;12(3):440–8.
Shao LB, Zhang Y, Lan BB, Wang JJ, Zhang ZW, Zhang LL, et al. MiRNA-sequence indicates that mesenchymal stem cells and exosomes have similar mechanism to enhance cardiac repair. Biomed Res Int. 2017. https://doi.org/10.1155/2017/4150705.
Pelizzo G, Avanzini MA, Cornaglia AI, De Silvestri A, Mantelli M, Travaglino P, et al. Extracellular vesicles derived from mesenchymal cells: perspective treatment for cutaneous wound healing in pediatrics. Regen Med. 2018;13(4):385–94.
Gatti S, Bruno S, Deregibus MC, Sordi A, Cantaluppi V, Tetta C, et al. Microvesicles derived from human adult mesenchymal stem cells protect against ischaemia-reperfusion-induced acute and chronic kidney injury. Nephrol Dial Transplant. 2011;26(5):1474–83.
Clark K, Zhang S, Barthe S, Kumar P, Pivetti C, Kreutzberg N, et al. Placental mesenchymal stem cell-derived extracellular vesicles promote myelin regeneration in an animal model of multiple sclerosis. Cells. 2019;8(12):1497.
Reza-Zaldivar EE, Hernandez-Sapiens MA, Gutierrez-Mercado YK, Sandoval-Avila S, Gomez-Pinedo U, Marquez-Aguirre AL, et al. Mesenchymal stem cell-derived exosomes promote neurogenesis and cognitive function recovery in a mouse model of Alzheimer’s disease. Neural Regen Res. 2019;14(9):1626–34.
Vakhshiteh F, Atyabi F, Ostad SN. Mesenchymal stem cell exosomes: a two-edged sword in cancer therapy. Int J Nanomed. 2019;14:2847–59.
Yeo RWY, Lai RC, Zhang B, Tan SS, Yin YJ, Teh BJ, et al. Mesenchymal stem cell: an efficient mass producer of exosomes for drug delivery. Adv Drug Deliv Rev. 2013;65(3):336–41.
Ban JJ, Lee M, Im W, Kim M. Low pH increases the yield of exosome isolation. Biochem Biophys Res Commun. 2015;461(1):76–9.
Zhang HC, Liu XB, Huang S, Bi XY, Wang HX, Xie LX, et al. Microvesicles derived from human umbilical cord mesenchymal stem cells stimulated by hypoxia promote angiogenesis both in vitro and in vivo. Stem Cells Dev. 2012;21(18):3289–97.
Lee SC, Jeong HJ, Lee SK, Kim SJ. Hypoxic conditioned medium from human adipose-derived stem cells promotes mouse liver regeneration through JAK/STAT3 signaling. Stem Cells Transl Med. 2016;5(6):816–25.
Domenis R, Cifu A, Quaglia S, Pistis C, Moretti M, Vicario A, et al. Pro inflammatory stimuli enhance the immunosuppressive functions of adipose mesenchymal stem cells derived exosomes. Sci Rep. 2018. https://doi.org/10.1038/s41598-018-31707-9.
Zhang Q, Fu L, Liang Y, Guo Z, Wang L, Ma C, et al. Exosomes originating from MSCs stimulated with TGF-β and IFN-γ promote Treg differentiation. J Cell Physiol. 2018;233(9):6832–40.
Lee SC, Jeong HJ, Lee SK, Kim SJ. Lipopolysaccharide preconditioning of adipose-derived stem cells improves liver-regenerating activity of the secretome. Stem Cell Res Therapy. 2015. https://doi.org/10.1186/s13287-015-0072-7.
Kim YE, Sung SI, Chang YS, Ahn SY, Sung DK, Park WS. Thrombin preconditioning enhances therapeutic efficacy of human wharton’s jelly-derived mesenchymal stem cells in severe neonatal hypoxic ischemic encephalopathy. Int J Mol Sci. 2019;20(10):2477.
Du W, Zhang KY, Zhang SQ, Wang R, Nie Y, Tao HY, et al. Enhanced proangiogenic potential of mesenchymal stem cell-derived exosomes stimulated by a nitric oxide releasing polymer. Biomaterials. 2017;133:70–81.
Bai Y, Han YD, Yan XL, Ren J, Zeng Q, Li XD, et al. Adipose mesenchymal stem cell-derived exosomes stimulated by hydrogen peroxide enhanced skin flap recovery in ischemia-reperfusion injury. Biochem Biophys Res Commun. 2018;500(2):310–7.
Sokolova V, Ludwig AK, Hornung S, Rotan O, Horn PA, Epple M, et al. Characterisation of exosomes derived from human cells by nanoparticle tracking analysis and scanning electron microscopy. Colloids and Surfaces B-Biointerfaces. 2011;87(1):146–50.
Rong X, Liu J, Yao X, Jiang T, Wang Y, Xie F. Human bone marrow mesenchymal stem cells-derived exosomes alleviate liver fibrosis through the Wnt/β-catenin pathway. Stem Cell Res Therapy. 2019;10(1):98.
Damania A, Jaiman D, Teotia AK, Kumar A. Mesenchymal stromal cell-derived exosome-rich fractionated secretome confers a hepatoprotective effect in liver injury. Stem Cell Res Therapy. 2018. https://doi.org/10.1186/s13287-017-0752-6.
Chen F, Li XL, Zhao JX, Geng J, Xie J, Xu B. Bone marrow mesenchymal stem cell-derived exosomes attenuate cardiac hypertrophy and fibrosis in pressure overload induced remodeling. Vitro Cellular Dev Biol Animal. 2020;56(7):567–76.
Teng XM, Chen L, Chen WQ, Yang JJ, Yang ZY, Shen ZY. Mesenchymal stem cell-derived exosomes improve the microenvironment of infarcted myocardium contributing to angiogenesis and anti-inflammation. Cell Physiol Biochem. 2015;37(6):2415–24.
Zou LY, Ma XK, Lin S, Wu BY, Chen Y, Peng CQ. Bone marrow mesenchymal stem cell-derived exosomes protect against myocardial infarction by promoting autophagy. Exp Ther Med. 2019;18(4):2574–82.
Xin HQ, Li Y, Cui YS, Yang JJ, Zhang ZG, Chopp M. Systemic administration of exosomes released from mesenchymal stromal cells promote functional recovery and neurovascular plasticity after stroke in rats. J Cereb Blood Flow Metab. 2013;33(11):1711–5.
Zhang YL, Chopp M, Zhang ZG, Katakowski M, Xin HQ, Qu CS, et al. Systemic administration of cell-free exosomes generated by human bone marrow derived mesenchymal stem cells cultured under 2D and 3D conditions improves functional recovery in rats after traumatic brain injury. Neurochem Int. 2017;111:69–81.
Cui GH, Wu J, Mou FF, Xie WH, Wang FB, Wang QL, et al. Exosomes derived from hypoxia-preconditioned mesenchymal stromal cells ameliorate cognitive decline by rescuing synaptic dysfunction and regulating inflammatory responses in APP/PS1 mice. FASEB J. 2018;32(2):654–68.
Casado JG, Blazquez R, Vela FJ, Alvarez V, Tarazona R, Sanchez-Margallo FM. Mesenchymal stem cell-derived exosomes: immunomodulatory evaluation in an antigen-induced synovitis porcine model. Front Veterinary Sci. 2017;4:39.
Ma XX, Xu SQ. TNF inhibitor therapy for rheumatoid arthritis (Review). Biomedical Rep. 2013;1(2):177–84.
Martins M, Ribeiro D, Martins A, Reis RL, Neves NM. Extracellular vesicles derived from osteogenically induced human bone marrow mesenchymal stem cells can modulate lineage commitment. Stem Cell Rep. 2016;6(3):284–91.
Qin Y, Wang L, Gao Z, Chen G, Zhang C. Bone marrow stromal/stem cell-derived extracellular vesicles regulate osteoblast activity and differentiation in vitro and promote bone regeneration in vivo. Sci Rep. 2016;6(1):21961.
Narayanan R, Huang C-C, Ravindran S. Hijacking the cellular mail: exosome mediated differentiation of mesenchymal stem cells. Stem Cells Int. 2016;2016:3808674.
Shimbo K, Miyaki S, Ishitobi H, Kato Y, Kubo T, Shimose S, et al. Exosome-formed synthetic microRNA-143 is transferred to osteosarcoma cells and inhibits their migration. Biochem Biophys Res Commun. 2014;445(2):381–7.
Shao M, Xu Q, Wu Z, Chen Y, Shu Y, Cao X, et al. Exosomes derived from human umbilical cord mesenchymal stem cells ameliorate IL-6-induced acute liver injury through miR-455-3p. Stem Cell Res Ther. 2020;11(1):37.
Zhou Y, Xu HT, Xu WR, Wang BY, Wu HY, Tao Y, et al. Exosomes released by human umbilical cord mesenchymal stem cells protect against cisplatin-induced renal oxidative stress and apoptosis in vivo and in vitro. Stem Cell Res Therapy. 2013;4:34.
Zou XY, Gu D, Xing XY, Cheng ZL, Gong DL, Zhang GY, et al. Human mesenchymal stromal cell-derived extracellular vesicles alleviate renal ischemic reperfusion injury and enhance angiogenesis in rats. Am J Transl Res. 2016;8(10):4289–99.
Zhang B, Wu X, Zhang X, Sun Y, Yan Y, Shi H, et al. Human umbilical cord mesenchymal stem cell exosomes enhance angiogenesis through the Wnt4/β-catenin pathway. Stem Cells Transl Med. 2015;4(5):513–22.
Zhou J, Liu HX, Li SH, Gong YS, Zhou MW, Zhang JH, et al. Effects of human umbilical cord mesenchymal stem cells-derived exosomes on fracture healing in rats through the Wnt signaling pathway. Eur Rev Med Pharmacol Sci. 2019;23(11):4954–60.
Fang S, Xu C, Zhang Y, Xue C, Yang C, Bi H, et al. Umbilical cord-derived mesenchymal stem cell-derived exosomal micrornas suppress myofibroblast differentiation by inhibiting the transforming growth Factor-β/SMAD2 pathway during wound healing. Stem Cells Transl Med. 2016;5(10):1425–39.
Mao F, Wu YB, Tang XD, Kang JJ, Zhang B, Yan YM, et al. Exosomes derived from human umbilical cord mesenchymal stem cells relieve inflammatory bowel disease in mice. Biomed Res Int. 2017;2017:1.
Korolkova OY, Myers JN, Pellom ST, Wang L, M’Koma AE. Characterization of serum cytokine profile in predominantly colonic inflammatory bowel disease to delineate ulcerative and crohn’s colitides. Clin Med Insights Gastroenterol. 2015;8:29–44.
Eirin A, Zhu XY, Puranik AS, Tang H, McGurren KA, van Wijnen AJ, et al. Mesenchymal stem cell-derived extracellular vesicles attenuate kidney inflammation. Kidney Int. 2017;92(1):114–24.
Cui X, He Z, Liang Z, Chen Z, Wang H, Zhang J. Exosomes from adipose-derived mesenchymal stem cells protect the myocardium against ischemia/reperfusion injury through Wnt/β-catenin signaling pathway. J Cardiovasc Pharmacol. 2017;70(4):225–31.
Liu Z, Xu YD, Wan YG, Gao J, Chu YY, Li J. Exosomes from adipose-derived mesenchymal stem cells prevent cardiomyocyte apoptosis induced by oxidative stress. Cell Death Discov. 2019. https://doi.org/10.1038/s41420-019-0159-5.
Katsuda T, Tsuchiya R, Kosaka N, Yoshioka Y, Takagaki K, Oki K, et al. Human adipose tissue-derived mesenchymal stem cells secrete functional neprilysin-bound exosomes. Sci Rep. 2013. https://doi.org/10.1038/srep01197.
Reza A, Choi YJ, Yasuda H, Kim JH. Human adipose mesenchymal stem cell-derived exosomal-miRNAs are critical factors for inducing anti-proliferation signalling to A2780 and SKOV-3 ovarian cancer cells. Sci Rep. 2016. https://doi.org/10.1038/srep38498.
Acknowledgements
Not applicable.
Funding
This publication is the result of the project implementation: “Open scientific community for modern interdisciplinary research in medicine (OPENMED)”, ITMS2014+ : 313011V455 supported by the Operational Programme Integrated Infrastructure, funded by the ERDF.
Author information
Authors and Affiliations
Contributions
JJ drafted, directed and finalized the manuscript. LS and DH participated in editing the manuscrift. TS gave suggestions during preparations, contributed to collecting information and editing manuscript. JR made critical revisions. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Janockova, J., Slovinska, L., Harvanova, D. et al. New therapeutic approaches of mesenchymal stem cells-derived exosomes. J Biomed Sci 28, 39 (2021). https://doi.org/10.1186/s12929-021-00736-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12929-021-00736-4