
Abstract
Background
Growing evidence suggests a role for gut bacteria and their metabolites in host-signaling responses along the gut-brain axis which may impact mental health. Meditation is increasingly utilized to combat stress, anxiety, and depression symptoms. However, its impact on the microbiome remains unclear. This study observes the effects of preparation and participation in an advanced meditation program (Samyama) implemented with a vegan diet including 50% raw foods, on gut microbiome and metabolites profiles.
Methods
There were 288 subjects for this study. Stool samples were collected at 3-time points for meditators and household controls. Meditators prepared for 2 months for the Samyama, incorporating daily yoga and meditation practices with a vegan diet including 50% raw foods. Subjects were requested to submit stool samples for 3 time points – 2 months before Samyama (T1), right before Samyama (T2), and 3 months following Samyama (T3). 16 s rRNA sequencing was used to study participants' microbiome. Alpha and beta diversities along with short-chain fatty acid (SCFA) were assessed. Metabolomics were performed on a mass spectrometer coupled to a UHLPC system and analyzed by El-MAVEN software.
Results
Alpha diversity showed no significant differences between meditators and controls, while beta diversity showed significant changes (padj = 0.001) after Samyama in meditators’ microbiota composition. After the preparation phase, changes in branched short-chain fatty acids, higher levels of iso-valerate (padj = 0.02) and iso-buytrate (padj = 0.019) were observed at T2 in meditators. Other metabolites were also observed to have changed in meditators at timepoint T2.
Conclusion
This study examined the impact of an advanced meditation program combined with a vegan diet on the gut microbiome. There was an increase in beneficial bacteria even three months after the completion of the Samyama program. Further study is warranted to validate current observations and investigate the significance and mechanisms of action related to diet, meditation, and microbial composition and function, on psychological processes, including mood.
Trial registration
Registration number: NCT04366544; Registered on 29/04/2020.
Similar content being viewed by others


Background
Understanding the influence of the gut microbiota on nervous system function is gaining increasing interest from the scientific community [1,2,3]. Ailments linked to the gut-brain axis include inflammatory bowel diseases, irritable bowel syndrome [4, 5], Parkinson’s diseases, anxiety, and depression [6,7,8]. Clarifying the impact of psychological interventions on the microbiota and vice versa could enhance the effectiveness and scope of therapeutic approaches for psychological, neurological, and digestive diseases.
The human gut microbiota influences emotional and psychological states in a bidirectional way. Recent evidence points to the microbiome as part of a neuro-immune-endocrine matrix [9, 10]. Bacteria native to the gut can activate neural pathways that participate in anxiety and depression [6, 11]. These include the hypothalamus–pituitary–adrenal (HPA) axis, a neuroendocrine system linked to stress response [12,13,14,15], which regulates the gut-brain signaling, and may influence stress related diseases such as anxiety and depression [8]. The reverse is also true as stress can alter the microbiota leading to increasing inflammation and lowering the anti-inflammatory and anti-tumor effects of a healthy microbiome [16].
Over the past decade, meditation has become an increasingly popular method to address symptoms related to stress, anxiety, and depression. Meditation effectively reduces symptoms associated with anxiety and depression [17,18,19]. Considering the potential impact of meditation on the stress response, meditation may additionally impact the gut microbiota [11, 20] and modulate the gut-brain axis [21]. Meditators demonstrate differences in the gut microbiota compared to non-meditators, which is characterized by enrichment of beneficial bacterial genera such as Bifidobacterium, Roseburia, and Subdoligranulum [11, 16]. We have previously reported that advanced meditation improves symptoms associated with anxiety and depression [22], results in positive psychological metrics [22, 23], reduces the expression of blood biomarker of inflammation [22], and increases the expression of endocannabinoids and brain-derived neurotrophic factor (BDNF) [23, 24]. Additionally, we have observed alterations following Samyama in functional brain connectivity between the salience and default mode networks in meditators [25], along with increased gene expression of immune signaling molecules that are relevant for diseases such as COVID-19 and multiple sclerosis [26]. We have also seen that there are significant changes in the lipidomics profile after Samyama [27]. In this study, we aimed to assess the impact of Samyama, an advanced meditation program that includes a preparatory phase of 60-day vegan diet requirement on the gut microbiome. We will measure outcomes for 1) microbiota composition and 2) metabolite composition.
Materials and methods
Ethics statement
This study was conducted in accordance with the Declaration of Helsinki [28]. It was reviewed and approved by the Indiana University School of Medicine Internal Review Board (IRB) (#1,801,728,792). Subjects provided electronic informed consent for this study after completing initial electronic surveys. This study adheres to CONSORT guidelines [29] (Fig. 1).

CONSORT diagram
Subject recruitment
The study was registered in ClinicalTrials.gov (reg #: NCT04366544) on 29/04/2020. Subjects for this non-randomized parallel controlled study were recruited from the same participant pool as a previously published study [22]. The Isha Institute of Inner Sciences (McMinnville, TN) provided a list of registrants for the April 2018 Samyama Program [30]. Invitation letters with study information were sent electronically to all registrants 2–3 months before the program. Study participants were recruited from the United States. Spouses or other household members were recruited as non-randomized control subjects. Subject recruitment included a cohort of meditators and cohort of spouses who were not enrolled in the program. There were 759 subjects enrolled in the studies, of which 288 (265 Samyama participants, 23 household controls) participated in this study. Study eligibility criteria included: Advanced meditation program participants at least 18 years of age. Exclusion criteria: Inability to read or comprehend the consent form; subjects with medical conditions in which a blood draw would be contraindicated (e.g., severe anemia); active use of marijuana, opioids, or related drugs; use of antibiotics or probiotic/prebiotic supplements within 60 days of enrollment; participants living outside of the United States. Spouses who actively participated in meditation were also excluded from the spousal control group. Group details can be found in Table 1.
Samyama program – preparatory process
Dietary requirements
As part of the Samyama preparatory process (60 days before the program), Samyama participants (meditators) followed a vegan diet with at least 50% raw foods consumed. Additionally, they were requested to avoid dietary intake of garlic, onion, chili, eggplant, asafoetida, coffee, and tea. Finally, alcohol, cigarettes, stimulants, and illicit drugs were discouraged. Controls were not required to follow any dietary requirements.
Meditation practice requirements
Samyama participants (meditators) were required to take several prerequisite meditation programs before enrolling in Samyama. These included Inner Engineering [31], Bhava Spandana [32], Shoonya [33], and Yogasanas [34]. They were asked to perform multiple practices (learned in prerequisite programs) daily for the 60-day preparation period. These include kriya yoga practices (Shakti Chalana Kriya and Shambhavi Mahamudra Kriya), hata yoga (Surya Kriya and Yogasanas), Shoonya meditation twice a day, Sukha Kriya and Arda Siddhasana for at least 1 h per day. Kriya yoga practices are combinations of posture, breath, and sound. Hata yoga practices consist of physical postures. Shoonya meditation is a process of conscious non-doing. Sukha Kriya consists of alternate nostril breathing, which leads to regulation of breath. Ardha Siddhasana is a posture in which one sits cross-legged with the heel of the left foot placed at the perineum. Controls did not have any pre-program practice requirements.
Samyama program – retreat
During the program, meditators were to remain silent for the entire 8-day duration of the program. They took part in all-day meditation sessions with intermittent breaks. The program hall was closed to external influences. No specific instructions or programs were given to the controls. Upon completing the Samyama program, there were no further restrictions on meditators. Meditators were able to return to their previous lifestyle.
Stool sampling
Subjects were requested to submit stool samples for 3 time points – 2 months before Samyama (T1), immediately before Samyama (T2), and 3 months following Samyama (T3). Stool samples were collected from meditators and control subjects in sterile fecal collection containers. Upon receiving samples from participants, they were packaged with cold packs and shipped overnight. The samples were then stored at -80 ℃ until samples were analyzed.
Metabolomics
Metabolomics were performed as described previously [35, 36]. Briefly, metabolomics runs were performed on a Q Exactive™ HF Hybrid Quadrupole-Orbitrap™ Mass Spectrometer (Thermo-Fisher) coupled to a Vanquish™ UHPLC System (Thermo-Fisher). Chromatographic separation was achieved on a Syncronis HILIC UHPLC column (2.1 mm x 100 mm × 1.7μm, Thermo-Fisher) using a binary solvent system at a flow rate of 600μL/min. Solvent A, 20 mM ammonium formate pH 3.0 in mass spectrometry grade H2O; Solvent B, mass spectrometry grade acetonitrile with 0.1% formic acid (%v/v). A sample injection volume of 2μL was used. The mass spectrometer was run in negative full scan mode at a resolution of 240,000 scanning from 50-750 m/z.
Metabolite data was analyzed by El-MAVEN software package and identified by matching observed m/z signals (± 10 ppm) and chromatographic retention times to those observed from commercial metabolite standards (Sigma-Aldrich) [37, 38]. Next, metabolites were quantified by comparison to an eight-point quantification curve of metabolite standards.
Microbiota composition and data analysis
We used 16 s rRNA sequencing to study the participants' microbiome over time. Sequencing data from MiSeq was de-multiplexed and converted to FASTQ format using Illumina's bcl2fastq (RRID:SCR_015058) software. CutAdapt was used for initial quality trimming [39]. DADA2 v1.10.1 pipeline was used to generate and ASV (amplicon sequence variant) table. Taxonomy was assigned to the representative sequences using the idTaxa classifier [40] from DECIPHER R package using SILVAv.132 database [41]. Downstream analysis was done in R using phyloseq 1.30.0 [42]. Alpha diversity, summarizing the structure of a community with respect to its richness and evenness in a sample [43,44,45], was measured by calculating the Shannon and Chao1 index. Beta diversity, used to assess changes in microbial community across different groups, was visualized using principal coordinate analysis (PCoA) on the Bray–Curtis dissimilarity matrix, and changes in the bacterial community were assessed statistically using PERMANOVA. The short-chain fatty acid (SCFA) data was converted to proportion by dividing each SCFA by total SCFA concentration in a sample. If the data was normally distributed, parametric one-way analysis of variance (ANOVA) with Tukey's post-hoc tests was used. All measurements were normalized using autoscaling for metabolomics data, and then t-test was performed with adjustment for multiple testing.
Results
Demographic data
There were 265 meditators enrolled in this study, including 139 females and 126 males with an average age of 40.7 ± 10.9 years, and 23 house-hold controls including 10 females and 13 males with an average age of 42 ± 1.41. Participants’ demographic data is summarized in Table 1.
Microbial diversity
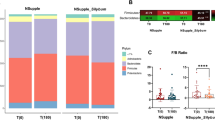
Figure 2 shows the phylum level relative abundance profile of all participants over time (Table 2). With these data we looked at alpha diversity to assess sample specific microbial composition and beta diversity to study the microbial community structure. Figure 3 shows two different alpha diversity measures, Shannon and Simpson, which resulted in no differences across or within meditator and control groups at all three time points. However, analysis of fecal microbiota beta-diversity demonstrated statistical differences between time points T2 and T3 within meditators (padj = 0.001) (Fig. 4).

Charts show phylum level relative abundance profile for meditators and controls at 3-time points

Alpha diversity plots A Boxplots of Shannon index for participants over time. B Boxplots of Simpson index for participants over time

Beta diversity plots. PCoA was used to visualize the Bray–Curtis similarity for all participants over time
Differential abundance analysis
To pinpoint the differences in the microbiota (genus level) we conducted differential abundance analysis over time in participants. We did not find any significantly different taxa in control samples over time (data not shown); hence we focused here on meditators only. Among meditators, Lachnospiraceae UCG-004 population increased (LFC = 0.91, padj = 0.0211) while Romboutsia (LFC = -2.2, padj = 5.1e-6) and Collinsella (LFC = -0.61, padj = 0.078) populations decreased at T2 from T1 (Fig. 5A). At timepoint T3 compared to T1, Lactobacillus (LFC = -3.4, padj = 0.02), Bifidobacterium (LFC = 0.82, padj = 0.003), Ruminococcaceae UCG-014 (LFC = 2.0, padj = 0.001) and Streptococcus (LF = 1.72, padj = 9.5e-6) populations increased while Bacteroides (LFC = -0.68,padj = 0.008) and Lachnospira (LFC = -0.92,padj = 0.01) populations decreased (Fig. 5B). The greatest changes were observed between T2 and T3 where Bacteroides (LFC = -0.9,padj = 5.1exp6), Lachnospiraceae UCG-004 (LFC = -1.48,padj = 4.6exp-5), Lachnospiraceae UCG-001 (LFC = -1.24,padj = 0.03), Lachnospira (LFC = 1.28,padj = 2.3exp-4) and [Eubacterium] eligens group (LFC = -1.48,padj = 9.2exp-5) populations decreased in T3, while Lactobacillus (LFC = 4.9,padj = 4.7exp-7), Ruminococcaceae UCG-014 (LFC = 2.03,padj = 0.017), Romboustia (LFC = 1.84,padj = 0.0018), Collinsella (LFC = 1.36,padj = 1.67exp-10) and Bifidobacterium (LFC = 1.25,padj3.67e-7 =) increased in T3 (Fig. 5C). Overall, by timepoint T3, Lactobacillus, Bifidobacterium, Ruminococcaceae, Streptococcus, and Collinsella populations were increased compared to both previous timepoints.

Taxa enrichment genus level results for meditators. A Baseline vs. Before Meditation (T1 vs. T2), B Baseline vs. After Meditation (T1 vs. T3), C Before vs. After Meditation (T2 vs. T3)
Short-chain fatty acid levels
We further investigated short chain fatty acid (SCFA) levels in participants over time. While we did not find differences over time in the proportions of SCFAs ascribed to carbohydrate metabolism (butyrate, acetate and propionate), we found that within meditators, branched SCFAs including iso-butyrate (Fig. 6A) and iso-valerate (Fig. 6B) levels were greater at T2 compared to T1 (padj = 0.019) and T3 (padj = 0.02). We also observed a lower proportion of valerate in meditators compared to controls at timepoint T3 (padj = 0.012) (Fig. 6C). No changes were seen within the controls.

SCFA showed a significant difference. A Iso Butyrate and B Iso-Valerate showed significant differences within meditators at T2 vs. T1 and T2 vs. T3. C Valerate showed significant differences in meditators vs. controls in T2. Significance is marked with an asterisk (*)
Other metabolites’ profiling
We also explored the changes in other microbial-derived metabolites over time in the participants (untargeted metabolomics analysis). While we did not find any metabolites to be significantly different in control participants over time, we found 46 metabolites to be significantly different in meditators overtime points. Most of these metabolites decreased in T2 and subsequently increased at T3 (Fig. 7).

Metabolomics intensity in log2 units. Heatmap of metabolites that have significantly changed across two timepoints. Most metabolites that show significant change are decreased in T2
Discussion
This non-randomized controlled study is among the first and largest to explore the gut microbiome and metabolome after an advanced meditation program delivered with a vegan diet. We demonstrated associations of beta diversity with time points T2 and T3 in the meditation group, suggesting compositional changes in the microbiota arising from either meditation or possibly returning to habitual diet from a vegan diet. However, measures of alpha diversity in the gut microbial communities were not different between groups at any time points. Additionally, we tested for sex differences in alpha and beta diversities (at each timepoint) but did not find any significant differences. This may be a consequence of a healthy, unselected participant group. We did not find any taxa to be differentially abundant between meditators and controls at any timepoint; however, an altered abundance of several genera was observed in meditators with changes of the greatest magnitude observed at T3 compared to T2. Furthermore, we demonstrated several changes in fecal metabolites over time.
Lachnospiraceae increased in abundance at T2 compared to T1, during the preparation phase. Lachnospiraceae has been linked to SCFAs production [46, 47], which was observed in our study by increased levels of iso-butyrate and iso-valerate in meditators during this time period. Lachnospiraceae can beneficially and adversely affect disease progression for a variety of ailments, including metabolic disorders, diabetes, inflammatory bowel disease, and depression [47]. Some taxa of Lachnospiraceae were shown to be positively correlated to a diet rich in unprocessed/raw foods, while others were correlated to higher intake of processed foods and meats [48]. We tested each SCFA for sex differences but except for iso-butyrate in T2 (pvalue = 0.02), we did not see any significant differences in the sexes. Another study showed an increase in Lachnospiraceae population after implementing a vegetarian diet [49]. This study validates previous reports that vegan diet increases Lachnospiraceae microbe in our study population of meditators practicing predominantly vegan diet.
We demonstrated increased abundance of Lactobacillus and Bifidobacterium at T3 compared to T1 and T2 (but not T1 to T2) in the meditation group, suggesting that the combination of vegan diet and meditation contributed to durable changes in bacterial composition. Bifidobacteria and Lactobacillus genera are considered probiotics [50], conferring positive health benefits to their host via their metabolic activities. Of interest, a double-blinded placebo-controlled trial conducted in medical students exposed to academic stress demonstrated that daily consumption of Lactobacillus casei improved sleep quality [51]. Gut microbial Lactobacillus counts were also associated with improved sleep quality in patients with bipolar disorder [52]. In addition, a negative correlation was identified between Bifidobacterium counts and cortisol levels in those patients, raising the possibility of a bacterial role in stress response [52].
A cross-sectional study [11] investigated the differences in gut microbial composition in 12 long-term meditators consuming a vegan diet and 12 omnivorous subjects who did not practice mediation. The abundance of 12 taxa in the samples of the meditation group was significantly higher than those in the control group, including Bifidobacterium and Collinsella.
Vegan diet and its effect on the gut microbiota has been well-studied during the last decade. Recently, a systematic review of the impact of vegan or vegetarian diet on gut microbial composition compared to an omnivorous diet identified no consistent association between a vegan or vegetarian diet and that of omnivores; in fact, some discordant results were noted [53]. At the genus level, significantly lower levels of Bacteroides in vegans were observed in some studies, although the majority of the included studies could not find any differences. Similarly, Bifidobacterium and Lactobacillus tended to be of lower abundance in vegans, but only in a small number of studies, with the majority not identifying differences between groups [53]. In contrast, a narrative review [54] concluded that compared with omnivores, vegetarians have higher gut microbiome diversity, and higher abundance of Prevotella, Clostridium, Lactobacillus, Ruminococcus, E. rectale, and F. prausnitzii, but lower abundance of Bacteroides Bifidobacterium.
Our findings relating to Bifidobacterium and Lactobacillus are similar to those identified in the above mentioned systematic review. Bifidobacterium increased at T3 compared to T1 and T2, but not at T2 compared to T1 that represents the period of vegan diet intervention. Higher Bifidobacterium abundance at T3 would suggest the possibility for habitual diet (rather than a vegan diet) to explain this finding, however this remains speculative as dietary data are not available to explore this hypothesis. An alternative hypothesis to explain higher bifidobacterium abundance at T3 could include effects of meditation. In the absence of stool collection immediately post meditation, this hypothesis remains speculative as well.
Although vegan diet quality may be variable, dietary fiber intake increases compared with vegetarians and omnivores [55]. Resistant starch is a form of dietary fiber that escapes small intestinal digestion and is fermented by the gut microbiota. Resistant starch sub-types have been associated with increased abundance of Bifidobacterium, Collinsella, and Romboutsia [56]. While unexpectedly, Collinesella and Romboutsia decreased in abundance during the strict vegan intervention (T1-T2), an increased abundance of both these genera at T3 compared to T2 and T1 in the intervention group was observed. These genera produce SCFAs which have an important role in intestinal immune cell regulation and maintenance of immune homeostasis, demonstrating an immunomodulatory potential [57]. In our study, no changes in major SCFAs were seen throughout the trial duration. However, changes in overall metabolite profiles and higher proportions of branched SCFA (BCFA) that may serve as markers of protein fermentation [58] were observed among meditators at T2. Although the broader effects of BCFA in human health are not yet well described, a recent report suggested the possibility that iso-butyrate may improve 5-hydroxytryptophan levels through upregulation of Tph1 expression to yield anti-depressant-like effects [59].
Increasing evidence is mounting for the role of gut bacteria and their metabolites in host-signaling responses along the gut-brain axis. Microbial components may influence brain functions via neuroendocrine and neuroimmune mechanisms [60]. A lower abundance of Bifidobacterium and Lactobacillus was previously observed in patients with major depressive disorders compared to healthy controls [61]. Lactobacillus plantarum DR7 intervention alleviated stress and anxiety in adults, and improved memory, attention, and emotional cognition. In addition, it was previously reported a positive correlation between Lactobacillus plantarum DR7 administration, gene expression of dopamine B-hydrolase, and abundance of genus Romboutsia suggesting a potential microbial impact on neurotransmitter function [60]. We recently demonstrated that in 632 adults who completed the Samyama program (including vegan diet and meditation), had improved scores of depression, anxiety, vitality, resilience, and joyfulness compared to baseline [22].
The body mass index (BMI) is lower in meditators compared to controls, and this may be a result of frequent yoga practices and greater adherence to vegetarian or vegan diets which frequently are observed in tandem. Microbial dysbiosis, specifically the Bacteroidetes to Firmicutes ratio is negatively correlated with BMI, which may be related to increased energy harvest, resulting in weight gain over time [62]. Additionally, a low-fat vegan diet was shown in a previous study to induce significant changes in gut microbiota which were related to changes in weight and body composition in otherwise healthy participants [63]. Therefore, BMI may have contributed mechanistically to our gut microbiome findings.
In this study, we demonstrated differences in several fecal metabolites in meditators over time, with the greatest differences observed at T2 (vegan diet period) compared to the other two timepoints. This suggests that a vegan diet leads to functional changes in the fecal microbiome. Although our current findings are preliminary and further study will be required to identify the affected microbial pathways that may drive altered metabolite profiles, others have demonstrated that gut bacterial metabolic activity differs significantly between those consuming vegan and omnivorous diets [64]. Although between group differences were not significant, the microbial compositional and functional changes observed in meditators but not the controls at T2 are consistent with vegan diet intervention, and findings at T3 could either be related to the effects of Samyama or a return to the usual diet. The authors cannot conclude the basis behind this change based on the data in this study as we do not have adequate dietary data. The strengths of this study include a large meditator sample size, longer follow-ups, and objective data collection that include blood samples, fMRI, and stool sample collection. As described in the introduction, several objective findings that include a sustained reduction in anxiety, depression, improved brain connectivity have been previously published from this study data [22, 25].
Study limitations include unselected study participants at baseline (underlying medical diagnoses are unknown therefore disease related microbial dysbiosis may have influenced study findings), lack of both baseline dietary assessment and validated measure of dietary compliance, and absence of stool collection immediately post intervention to inform the combined effects of vegan diet and meditation (before returning to habitual diet). Furthermore, controls did not have the same dietary restrictions as meditators. This includes restrictions for garlic, onions, peppers, eggplant, asafoetida, coffee, and tea. Additionally, while the T2 microbial composition and functional changes at T2 are likely reflective of vegan diet in the intervention group, impact of the pre-requisite yoga practices on these measures is unknown. Future studies may build from this body of work to rigorously detangle the relationship between diet and yoga practices. It is also important to mention that BMI is significantly lower in the meditator pool and this may have impacted findings. Finally, it was not determined if the controls conducted yoga/meditation practices or what practices were observed by meditators after completing the Samyama program, and if meditators returned to their habitual diet post Samyama.
Conclusion
This non-randomized controlled longitudinal prospective study is among the first and largest to explore the gut microbiome and metabolome profile after an advanced Samyama meditation program delivered with a vegan diet. First, we observed changes in meditator beta diversity after Samyama. Additionally, after the preparatory phase, there was an increase in branched short-chain fatty acids and changes to other metabolite composition. While the implications of these results are not fully understood, these findings do pave the way for further exploration of the impact of meditation and diet on the gut-brain axis. Further study is warranted to validate current observations, investigate significance and mechanisms of action related to diet, meditation, and microbial composition and function, on psychological processes, including mood.
Availability of data and materials
The data is available in from the United States National Library of Medicine and the National Center for Biotechnology Information. The BioProject is 904395 (https://www.ncbi.nlm.nih.gov/bioproject/904395).
Abbreviations
- ANOVA:
-
Analysis of variance
- ASV:
-
Amplicon sequence variant
- BCFA:
-
Branched chain fatty acid
- BDNF:
-
Brain-derived neurotropic factor
- HPA:
-
Hypothalamus-pituitary-adrenal
- PCoA:
-
Principal coordinate analysis
- SCFA:
-
Short-chain fatty acid
- BMI:
-
Body mass index
References
Wang HX, Wang YP. Gut microbiota-brain axis. Chin Med J (Engl). 2016;129(19):2373–80. https://doi.org/10.4103/0366-6999.190667. Epub 2016/09/21
Osadchiy V, Martin CR, Mayer EA. The gut-brain axis and the microbiome: mechanisms and clinical implications. Clin Gastroenterol Hepatol. 2019;17(2):322–32. https://doi.org/10.1016/j.cgh.2018.10.002. Epub 2018/10/08
Cryan JF, O'Riordan KJ, Cowan CSM, Sandhu KV, Bastiaanssen TFS, Boehme M, et al. The microbiota-gut-brain axis. Physiol Rev. 2019;99(4):1877–2013. https://doi.org/10.1152/physrev.00018.2018. Epub 2019/08/29
Moser G, Fournier C, Peter J. Intestinal microbiome-gut-brain axis and irritable bowel syndrome. Wien Med Wochenschr. 2018;168(3-4):62–6. https://doi.org/10.1007/s10354-017-0592-0. Epub 2017/09/10
Quigley EMM. The gut-brain axis and the microbiome: clues to pathophysiology and opportunities for novel management strategies in irritable bowel syndrome (Ibs). J Clin Med. 2018;7(1) https://doi.org/10.3390/jcm7010006. Epub 2018/01/06
Foster JA, McVey Neufeld KA. Gut-brain axis: how the microbiome influences anxiety and depression. Trends Neurosci. 2013;36(5):305–12. https://doi.org/10.1016/j.tins.2013.01.005. Epub 2013/02/07
Molina-Torres G, Rodriguez-Arrastia M, Roman P, Sanchez-Labraca N, Cardona D. Stress and the gut microbiota-brain axis. Behav Pharmacol. 2019;30(2 and 3-Spec Issue):187–200. https://doi.org/10.1097/FBP.0000000000000478. Epub 2019/03/08
Lach G, Schellekens H, Dinan TG, Cryan JF. Anxiety, depression, and the microbiome: a role for gut peptides. Neurotherapeutics. 2018;15(1):36–59. https://doi.org/10.1007/s13311-017-0585-0. Epub 2017/11/15
Cruz-Pereira JS, Rea K, Nolan YM, O’Leary OF, Dinan TG, Cryan JF. Depression’s unholy trinity: dysregulated stress, immunity, and the microbiome. Annu Rev Psychol. 2020;71:49–78. https://doi.org/10.1146/annurev-psych-122216-011613. Epub 2019/10/01
Peirce JM, Alvina K. The role of inflammation and the gut microbiome in depression and anxiety. J Neurosci Res. 2019;97(10):1223–41. https://doi.org/10.1002/jnr.24476. Epub 2019/05/31
Jia W, Zhen J, Liu A, Yuan J, Wu X, Zhao P, et al. Long-term vegan meditation improved human gut microbiota. Evid Based Complement Alternat Med. 2020;2020:9517897. https://doi.org/10.1155/2020/9517897. Epub 2020/07/28
Farzi A, Frohlich EE, Holzer P. Gut microbiota and the neuroendocrine system. Neurotherapeutics. 2018;15(1):5–22. https://doi.org/10.1007/s13311-017-0600-5. Epub 2018/01/31
Frankiensztajn LM, Elliott E, Koren O. The Microbiota and the Hypothalamus-Pituitary-Adrenocortical (Hpa) Axis, Implications for Anxiety and Stress Disorders. Curr Opin Neurobiol. 2020;62:76–82. https://doi.org/10.1016/j.conb.2019.12.003. Epub 2020/01/24
Du Y, Gao XR, Peng L, Ge JF. Crosstalk between the microbiota-gut-brain axis and depression. Heliyon. 2020;6(6):e04097. https://doi.org/10.1016/j.heliyon.2020.e04097. Epub 2020/06/13
Petra AI, Panagiotidou S, Hatziagelaki E, Stewart JM, Conti P, Theoharides TC. Gut-microbiota-brain axis and its effect on neuropsychiatric disorders with suspected immune dysregulation. Clin Ther. 2015;37(5):984–95. https://doi.org/10.1016/j.clinthera.2015.04.002. Epub 2015/06/06
Househam AM, Peterson CT, Mills PJ, Chopra D. The effects of stress and meditation on the immune system, human microbiota, and epigenetics. Adv Mind Body Med. 2017;31(4):10–25. Epub 2018/01/08
Saeed SA, Cunningham K, Bloch RM. Depression and anxiety disorders: benefits of exercise, yoga, and meditation. Am Fam Physician. 2019;99(10):620–7. Epub 2019/05/16
Saeed SA, Antonacci DJ, Bloch RM. Exercise, yoga, and meditation for depressive and anxiety disorders. Am Fam Physician. 2010;81(8):981–6. Epub 2010/04/15
Gonzalez-Valero G, Zurita-Ortega F, Ubago-Jimenez JL, Puertas-Molero P. Use of meditation and cognitive behavioral therapies for the treatment of stress, depression and anxiety in students. A systematic review and meta-analysis. Int J Environ Res Public Health. 2019;16(22) https://doi.org/10.3390/ijerph16224394. Epub 2019/11/14
Schnorr SL, Bachner HA. Integrative therapies in anxiety treatment with special emphasis on the gut microbiome. Yale J Biol Med. 2016;89(3):397–422. Epub 2016/10/05
Ningthoujam DS, Singh N, Mukherjee S. Possible roles of cyclic meditation in regulation of the gut-brain axis. Front Psychol. 2021;12:768031. https://doi.org/10.3389/fpsyg.2021.768031. Epub 2022/01/11
Sadhasivam S, Alankar S, Maturi R, Williams A, Vishnubhotla RV, Hariri S, et al. Isha Yoga practices and participation in samyama program are associated with reduced Hba1c and systemic inflammation, improved lipid profile, and short-term and sustained improvement in mental health: a prospective observational study of meditators. Front Psychol. 2021;12(1771) https://doi.org/10.3389/fpsyg.2021.659667.
Sadhasivam S, Alankar S, Maturi R, Vishnubhotla RV, Mudigonda M, Pawale D, et al. Inner engineering practices and advanced 4-day isha yoga retreat are associated with cannabimimetic effects with increased endocannabinoids and short-term and sustained improvement in mental health: a prospective observational study of meditators. Evid Based Complement Alternat Med. 2020;2020:8438272. https://doi.org/10.1155/2020/8438272. Epub 2020/07/01
Cahn BR, Goodman MS, Peterson CT, Maturi R, Mills PJ. Yoga, meditation and mind-body health: increased Bdnf, cortisol awakening response, and altered inflammatory marker expression after a 3-month yoga and meditation retreat. Front Hum Neurosci. 2017;11:315. https://doi.org/10.3389/fnhum.2017.00315. Epub 2017/07/12
Vishnubhotla RV, Radhakrishnan R, Kveraga K, Deardorff R, Ram C, Pawale D, et al. Advanced meditation alters resting-state brain network connectivity correlating with improved mindfulness. Front Psychol. 2021;12:745344. https://doi.org/10.3389/fpsyg.2021.745344. Epub 2021/12/07
Chandran V, Bermudez ML, Koka M, Chandran B, Pawale D, Vishnubhotla R, et al. Large-scale genomic study reveals robust activation of the immune system following advanced inner engineering meditation retreat. Proc Natl Acad Sci U S A. 2021;118(51) https://doi.org/10.1073/pnas.2110455118. Epub 2021/12/16
Vishnubhotla RV, Wood PL, Verma A, Cebak JE, Hariri S, Mudigonda M, et al. Advanced meditation and vegan diet increased acylglycines and reduced lipids associated with improved health: a prospective longitudinal study. J Integr Complement Med. 2022; https://doi.org/10.1089/jicm.2022.0480. Epub 2022/05/10
World Medical A. World Medical Association Declaration of Helsinki: ethical principles for medical research involving human subjects. JAMA. 2013;310(20):2191–4. https://doi.org/10.1001/jama.2013.281053. Epub 2013/10/22
Schulz KF, Altman DG, Moher D, Group C. Consort 2010 statement: updated guidelines for reporting parallel group randomised trials. PLoS Med. 2010;7(3):e1000251. https://doi.org/10.1371/journal.pmed.1000251. Epub 2010/03/31
Isha Foundation. Samyama (2019). Available from: https://isha.sadhguru.org/us/en/yoga-meditation/yoga-program-advanced/samyama. [cited 17 Feb 2022].
Isha Foundation. Sadhguru Offers Inner Engineering (2019). Available from: https://www.innerengineering.com/ [cited 17 Feb 2022].
Isha Foundation. Bhava Spandana Program (2019). Available from: https://isha.sadhguru.org/us/en/yoga-meditation/yoga-program-advanced/bhava-spandana [cited 02 Feb 2022].
Isha Foundation. Shoonya Meditation (2019). Available from: https://isha.sadhguru.org/us/en/yoga-meditation/yoga-program-advanced/shoonya-meditation [cited 17 Feb 2022].
Isha Foundation. Yogasanas (2019). Available from: https://isha.sadhguru.org/us/en/yoga-meditation/yoga-program-for-beginners/hatha-yoga/yoga-asanas [cited 17 Feb 2022].
Mager LF, Burkhard R, Pett N, Cooke NCA, Brown K, Ramay H, et al. Microbiome-derived inosine modulates response to checkpoint inhibitor immunotherapy. Science. 2020;369(6510):1481–9. https://doi.org/10.1126/science.abc3421. Epub 2020/08/15
Dong X, Rattray JE, Campbell DC, Webb J, Chakraborty A, Adebayo O, et al. Thermogenic hydrocarbon biodegradation by diverse depth-stratified microbial populations at a scotian basin cold seep. Nat Commun. 2020;11(1):5825. https://doi.org/10.1038/s41467-020-19648-2. Epub 2020/11/19
Clasquin MF, Melamud E, Rabinowitz JD. Lc-Ms data processing with maven: a metabolomic analysis and visualization engine. Curr Protoc Bioinformatics. 2012;Chapter 14:Unit14 1. https://doi.org/10.1002/0471250953.bi1411s37. Epub 2012/03/06
Melamud E, Vastag L, Rabinowitz JD. Metabolomic Analysis and Visualization Engine for Lc-Ms Data. Anal Chem. 2010;82(23):9818–26. Epub 2010/11/06. https://doi.org/10.1021/ac1021166.
Martin M. Cutadapt removes adapter sequences from high-throughput sequencing reads. 2011;17(1):3. https://doi.org/10.14806/ej.17.1.200. Epub 2011-08-02
Murali A, Bhargava A, Wright ES. Idtaxa: a novel approach for accurate taxonomic classification of microbiome sequences. Microbiome. 2018;6(1):140. https://doi.org/10.1186/s40168-018-0521-5. Epub 2018/08/11
Quast C, Pruesse E, Yilmaz P, Gerken J, Schweer T, Yarza P, et al. The silva ribosomal rna gene database project: improved data processing and web-based tools. Nucleic Acids Res. 2013;41(Database issue):D590–6. https://doi.org/10.1093/nar/gks1219. Epub 2012/11/30
McMurdie PJ, Holmes S. Phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PLoS One. 2013;8(4):e61217. https://doi.org/10.1371/journal.pone.0061217. Epub 2013/05/01
Whittaker RH. Evolution and measurement of species diversity. Taxon. 1972;21:213–51.
Whittaker RH. Vegetation of the Siskiyou Mountains, Oregon and California. Ecol Monographs. 1960;30(3):279–338. https://doi.org/10.2307/1943563.
Willis AD. Rarefaction, alpha diversity, and statistics. Front Microbiol. 2019;10:2407. https://doi.org/10.3389/fmicb.2019.02407. Epub 2019/11/12
Sorbara MT, Littmann ER, Fontana E, Moody TU, Kohout CE, Gjonbalaj M, et al. Functional and genomic variation between human-derived isolates of lachnospiraceae reveals inter- and intra-species diversity. Cell Host Microbe. 2020;28(1):134–46 e4. https://doi.org/10.1016/j.chom.2020.05.005. Epub 2020/06/04
Vacca M, Celano G, Calabrese FM, Portincasa P, Gobbetti M, De Angelis M. The controversial role of human gut lachnospiraceae. Microorganisms. 2020;8(4) https://doi.org/10.3390/microorganisms8040573. Epub 2020/04/25
Matsuyama M, Morrison M, Cao KL, Pruilh S, Davies PSW, Wall C, et al. Dietary Intake Influences gut microbiota development of healthy Australian children from the age of one to two years. Sci Rep. 2019;9(1):12476. https://doi.org/10.1038/s41598-019-48658-4. Epub 2019/08/30
Barrett HL, Gomez-Arango LF, Wilkinson SA, McIntyre HD, Callaway LK, Morrison M, et al. A vegetarian diet is a major determinant of gut microbiota composition in early pregnancy. Nutrients. 2018;10(7) https://doi.org/10.3390/nu10070890. Epub 2018/07/14
Hill C, Guarner F, Reid G, Gibson GR, Merenstein DJ, Pot B, et al. Expert Consensus Document. The International Scientific Association for Probiotics and Prebiotics Consensus Statement on the Scope and Appropriate Use of the Term Probiotic. Nat Rev Gastroenterol Hepatol. 2014;11(8):506–14. https://doi.org/10.1038/nrgastro.2014.66. Epub 2014/06/11
Takada M, Nishida K, Gondo Y, Kikuchi-Hayakawa H, Ishikawa H, Suda K, et al. Beneficial effects of lactobacillus casei strain shirota on academic stress-induced sleep disturbance in healthy adults: a double-blind, randomised, placebo-controlled trial. Benef Microbes. 2017;8(2):153–62. https://doi.org/10.3920/BM2016.0150. Epub 2017/04/27
Aizawa E, Tsuji H, Asahara T, Takahashi T, Teraishi T, Yoshida S, et al. Bifidobacterium and lactobacillus counts in the gut microbiota of patients with bipolar disorder and healthy controls. Front Psychiatry. 2018;9:730. https://doi.org/10.3389/fpsyt.2018.00730. Epub 2019/02/05
Trefflich I, Jabakhanji A, Menzel J, Blaut M, Michalsen A, Lampen A, et al. Is a vegan or a vegetarian diet associated with the microbiota composition in the Gut? Results of a new cross-sectional study and systematic review. Crit Rev Food Sci Nutr. 2020;60(17):2990–3004. https://doi.org/10.1080/10408398.2019.1676697. Epub 2019/10/22
Xiao W, Zhang Q, Yu L, Tian F, Chen W, Zhai Q. Effects of vegetarian diet-associated nutrients on gut microbiota and intestinal physiology. Food Sci Hum Wellness. 2022;11(2):208–17. https://doi.org/10.1016/j.fshw.2021.11.002.
Clarys P, Deliens T, Huybrechts I, Deriemaeker P, Vanaelst B, De Keyzer W, et al. Comparison of nutritional quality of the vegan, vegetarian, semi-vegetarian, pesco-vegetarian and omnivorous diet. Nutrients. 2014;6(3):1318–32. https://doi.org/10.3390/nu6031318. Epub 2014/03/29
Qin R, Wang J, Chao C, Yu J, Copeland L, Wang S, et al. Rs5 produced more butyric acid through regulating the microbial community of human gut microbiota. J Agric Food Chem. 2021;69(10):3209–18. https://doi.org/10.1021/acs.jafc.0c08187. Epub 2021/02/26
Ratajczak W, Mizerski A, Ryl A, Slojewski M, Sipak O, Piasecka M, et al. Alterations in Fecal Short Chain Fatty Acids (Scfas) and Branched Short-Chain Fatty Acids (Bcfas) in Men with Benign Prostatic Hyperplasia (Bph) and Metabolic Syndrome (Mets). Aging (Albany NY). 2021;13(8):10934–54. https://doi.org/10.18632/aging.202968. Epub 2021/04/14
Macfarlane GT, Gibson GR, Beatty E, Cummings JH. Estimation of short-chain fatty acid production from protein by human intestinal bacteria based on branched-chain fatty acid measurements. FEMS Microbiol Lett. 1992;101(2):81–8. https://doi.org/10.1016/0378-1097(92)90829-D.
Tian P, O'Riordan KJ, Lee YK, Wang G, Zhao J, Zhang H, et al. Towards a psychobiotic therapy for depression: bifidobacterium breve Ccfm1025 reverses chronic stress-induced depressive symptoms and gut microbial abnormalities in mice. Neurobiol Stress. 2020;12:100216. https://doi.org/10.1016/j.ynstr.2020.100216. Epub 2020/04/08
Liu G, Chong HX, Chung FY, Li Y, Liong MT. Lactobacillus plantarum Dr7 modulated bowel movement and gut microbiota associated with dopamine and serotonin pathways in stressed adults. Int J Mol Sci. 2020;21(13) https://doi.org/10.3390/ijms21134608. Epub 2020/07/03
Aizawa E, Tsuji H, Asahara T, Takahashi T, Teraishi T, Yoshida S, et al. Possible association of bifidobacterium and lactobacillus in the gut microbiota of patients with major depressive disorder. J Affect Disord. 2016;202:254–7. https://doi.org/10.1016/j.jad.2016.05.038. Epub 2016/06/12
Jumpertz R, Le DS, Turnbaugh PJ, Trinidad C, Bogardus C, Gordon JI, et al. Energy-balance studies reveal associations between gut microbes, caloric load, and nutrient absorption in humans. Am J Clin Nutr. 2011;94(1):58–65. https://doi.org/10.3945/ajcn.110.010132. Epub 2011/05/06
Kahleova H, Rembert E, Alwarith J, Yonas WN, Tura A, Holubkov R, et al. Effects of a low-fat vegan diet on gut microbiota in overweight individuals and relationships with body weight, body composition, and insulin sensitivity. A randomized clinical trial. Nutrients. 2020;12(10) https://doi.org/10.3390/nu12102917. Epub 2020/09/30
Prochazkova M, Budinska E, Kuzma M, Pelantova H, Hradecky J, Heczkova M, et al. Vegan diet is associated with favorable effects on the metabolic performance of intestinal microbiota: a cross-sectional multi-omics study. Front Nutr. 2021;8:783302. https://doi.org/10.3389/fnut.2021.783302. Epub 2022/01/25
Acknowledgements
The authors appreciate support provided by Isha Institute of Inner Sciences, McMinnville, TN and study volunteers for this prospective research on Samyama participants.
Funding
This research was funded by Indiana University, Indianapolis, IN.
Author information
Authors and Affiliations
Contributions
MR contributed to manuscript preparation, analysis, and experimental design. RV contributed to manuscript preparation and experimental design. HR contributed to analysis and manuscript preparation. MG and AS contributed to manuscript preparation. DP contributed to design and recruitment. BS and SS contributed to manuscript preparation, experimental design, and overall supervision. The author(s) read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This study was conducted in accordance with the Declaration of Helsinki (28). It was reviewed and approved by the Indiana University School of Medicine Internal Review Board (IRB) (#1801728792). Subjects provided electronic informed consent for this study after completing initial electronic surveys.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Raman, M., Vishnubhotla, R., Ramay, H.R. et al. Isha yoga practices, vegan diet, and participation in Samyama meditation retreat: impact on the gut microbiome & metabolome – a non-randomized trial. BMC Complement Med Ther 23, 107 (2023). https://doi.org/10.1186/s12906-023-03935-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12906-023-03935-8