
Abstract
Background
This study investigates the intricate role of Heme Oxygenase 1 (HMOX1) in ovarian cancer, emphasizing its prognostic significance, influence on immune cell infiltration, and impact on the malignant characteristics of primary ovarian cancer cells.
Materials and methods
Our research began with an analysis of HMOX1 expression and its prognostic implications using data from The Cancer Genome Atlas (TCGA) dataset, supported by immunohistochemical staining. Further analyses encompassed co-expression studies, Gene Ontology (GO) annotations, and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment. We utilized the TIMER and TISIDB platforms to evaluate the immunotherapeutic potential of HMOX1. Additionally, in vitro studies that involved modulating HMOX1 levels in primary ovarian cancer cells were conducted to confirm its biological functions.
Results
Our findings indicate a significant overexpression of HMOX1 in ovarian cancer, which correlates with increased tumor malignancy and poorer prognosis. HMOX1 was shown to significantly modulate the infiltration of immune cells, particularly neutrophils and macrophages. Single-cell RNA sequencing (scRNA-seq) analysis revealed that HMOX1 is predominantly expressed in tumor-associated macrophages (TAMs), with a positive correlation to chemokines and their receptors. An increase in HMOX1 levels was associated with heightened levels of immunoinhibitors, immunostimulators, and MHC molecules. Functional assays demonstrated that HMOX1 knockdown promotes apoptosis, attenuating cell proliferation and invasion, while its overexpression yields opposing effects.
Conclusion
HMOX1 emerges as a critical therapeutic target, intricately involved in immunomodulation, prognosis, and the malignant behavior of ovarian cancer. This highlights HMOX1 as a potential biomarker and therapeutic target in the fight against ovarian cancer.
Similar content being viewed by others


Introduction
Ovarian cancer poses a significant health concern for women, ranking third in gynecologic cancer incidence and second in cancer-related mortality worldwide [1]. It is categorized into two primary types: primary ovarian cancer, which accounts for 90–95% of cases, and metastatic cancer, comprising the remaining 5–10% [2]. The majority of ovarian cancer cases (over 90%) are classified as epithelial ovarian cancers (EOCs), which are further divided into five subgroups: high-grade serous cancer (HGSC), low-grade serous cancer (LGS), endometrioid cancer (EMC), clear cell cancer (CCC), and mucinous cancer (MC) [3]. Unfortunately, most patients are diagnosed at an advanced stage due to the absence of specific symptoms in the early phases of the disease. The current treatment strategy for ovarian cancer involves a combination of surgery and chemotherapy, which has evolved into personalized precision therapy [4]. Additionally, immunotherapies, including immune checkpoint inhibitors, have garnered attention in ovarian cancer research. However, their clinical effectiveness is often compromised by issues such as drug resistance, cancer recurrence, and immune escape. Consequently, there is an urgent need for the identification of new molecular targets to better assess the progression of ovarian cancer and enhance treatment options.
Ferroptosis, a newly identified form of programmed cell death, is characterized by excessive iron-dependent lipid peroxidation [5]. It has been shown to play a crucial role in the development and progression of various diseases, including tumors, neurological disorders, acute kidney injuries, and ischemia/reperfusion syndromes [6]. However, there are fewer than 100 articles that specifically focus on ferroptosis in ovarian cancer. These studies examine various aspects such as intracellular iron levels, the Hippo pathway, the transsulfuration pathway, and relevant genes like p53, SCD1, and FZD7 [7, 8]. Additionally, some publications investigate the potential of harnessing ferroptosis to overcome drug resistance to cisplatin and paclitaxel in ovarian cancer [9]. Looking ahead, it is anticipated that novel ferroptosis-related drugs and targets will be utilized in the diagnosis, prognosis, recurrence, and treatment of ovarian cancer. Consequently, our study aims to explore the role of ferroptosis regulators in ovarian cancer, with the objective of identifying new therapeutic strategies.
The heme oxygenase-1 (HMOX1, HO-1) enzyme plays a critical role in heme metabolism by degrading heme into carbon monoxide, ferrous ions, and biliverdin, which is subsequently converted to bilirubin [10]. HMOX1 is expressed at low levels in various tissues; however, it is overexpressed in cancers such as stomach, pancreatic, and colon cancer [11]. This overexpression is associated with poor prognosis, low survival rates, and reduced responses to therapy. Additionally, HMOX1 plays a significant role in ferroptosis. In fibrosarcoma cells (HT-1080), HMOX1 levels induced by erastin increase in a dose-dependent and time-dependent manner. Evidence from a Hmox1 knockout mouse model and the use of the HMOX1 inhibitor Zinc protoporphyrin IX indicate that HMOX1 promotes erastin-induced ferroptosis, which correlates well correlated with iron bioavailability [12]. Furthermore, HMOX1 expression levels are found to be higher in ovarian cancer tissues compared to normal tissues, and these levels are significantly correlated with FIGO staging and lymphatic metastases. HMOX1 also promotes epithelial-mesenchymal transition (EMT) in ovarian cancer cells, facilitating invasion and migration [13]. Following treatment of ovarian cancer tissues with the HMOX1 inducer heme for up to 24 h, there was an upregulation of Vimentin, Zeb1 and BCL-2, while keratin and the pro-apoptotic protein Bax were downregulated. These results were reversed 24 h after treatment with the HO-1 inhibitor Znpp [14]. However, the role of HMOX1 in regulating the immunosuppressive microenvironment and the infiltration of immune cells in ovarian cancer remains unclear. In this study, we conducted a bioinformatics analysis to systematically explore the biological function and immunomodulatory effect of HMOX1, which we further validated through experiments in ovarian cancer.
Materials and methods
Collection of the original data
Bulk RNA-seq data and clinical information from 371 patients diagnosed with high-grade ovarian serous carcinoma were sourced from the TCGA database. The mRNA expression data underwent normalization using the formula log2 (TPM + 1). Additionally, gene expression data were obtained from the GEO Dataset (http://ncbi.nlm.nih.gov/geo/), specifically from GSE40595 (Platform: GPL570), GSE172016 (Platform: GPL16791), and GSE180687 (Platform: GPL1261) were utilized. The GEO2R tool was employed to identify differentially expressed genes, selecting those that met the criteria of |log2(fold change)| > 1 and a false discovery rate (FDR) < 0.05 for further analysis. Furthermore, HMOX1 mRNA expression data in ovarian cancer cell lines were retrieved from the Cancer Cell Line Encyclopedia (CCLE, http://portals.btoadinstitute.org/CCLE/home) database [15].
Prognosis survival analysis
To evaluate the relationship between HMOX1 mRNA expression and survival outcomes in ovarian cancer patients, Kaplan-Meier survival curves and log-rank tests were performed using the Kaplan Meier Plotter (http://kmplot.com/). This analysis incorporated clinical survival data, including overall survival (OS), post-progression survival (PPS), and progression-free survival (PFS) from the GEO, EGA, and TCGA databases.
Functional analysis
TCGA ovarian cancer patients were categorized into two groups based on the median HMOX1 mRNA expression using the Home for Researchers platform (https://www.home-for-researchers.com/). Differentially expressed genes between the two groups were analyzed using the Limma package (version 3.40.2) in R software, applying the threshold criteria of adjusted P-values < 0.05 and |log2(fold change)| >1. To further elucidate the role of HMOX1 in ovarian cancer, the ClusterProfiler package (version 3.18.0) in R was employed to analyze the Gene Ontology (GO) function of the differentially expressed genes and to enrich the KEGG pathways.
Correlation analysis of ferroptosis
Genes associated with ferroptosis were obtained from the FerrDb database (http://www.zhounan.org/ferrdb/operations/download.html). The correlation between ferroptosis-related genes and HMOX1 was evaluated using the Pearson correlation coefficient. A correlation was deemed significant if the p-value was less than 0.05 and the absolute value of the correlation coefficient exceeded 0.3 (|R| > 0.3).
Immune infiltration and tumor microenvironment analysis
The influence of HMOX1 on tumor purity and various immune cell types—including neutrophils, macrophages, dendritic cells, B cells, CD8 + T cells, and CD4 + T cells—in ovarian cancer (OC) was assessed using the TIMER database. Additionally, TISIDB, a web portal that integrates multiple heterogeneous data types to predict immunotherapy responses and identify novel immunotherapy targets [16], was employed to investigate the relationship between the abundance of tumor-infiltrating lymphocytes (TILs), immunomodulators, chemokines (or receptors), and HMOX1 expression. Furthermore, we examined the distribution of HMOX1 expression across different immune and molecular subtypes, as well as the effects of drugs targeting HMOX1.
Sample collection, immunohistochemistry and isolation of primary ovarian cancer cell
In this study, we conducted immunohistochemical analysis using 10 paraffin-embedded ovarian tumor tissues alongside 10 control ovarian tissues to assess the expression of HMOX1 (abcam, 1:100). All participating patients provided written informed consent, and the study received approval from the Ethics Committee of Shunde Hospital of Southern Medical University.
Following previously established protocols [17], primary human ovarian cancer cells were isolated from the tumor tissues of a patient admitted to Shunde Hospital of Southern Medical University, employing an enzyme dispersion method. The isolated tumor cells were cultured and expanded, with cells at passages P1 to P4, achieving 75% confluence, selected for experimentation.
Cell treatment
Primary cancer cells were treated with 10 µM ZnPPIX (Selleck Chemicals) and 10 µM Hemin (Selleck Chemicals) for a duration of 72 h, in accordance with the manufacturer’s instructions. This treatment was designed to inhibit and activate HMOX1.
Apoptosis detection
The Annexin V apoptosis detection kit (Beyotime) was employed to quantify cell apoptosis. Following treatment with ZnPPIX/Hemin, the cells were washed with phosphate-buffered saline (PBS) and resuspended. Propidium iodide (PI) and annexin V-FITC were subsequently added, and the mixture was incubated in the dark for 30 min. Apoptosis in primary ovarian cancer cells was analyzed using flow cytometry (BD Biosciences, San Jose, California, USA).
EdU incorporation assay
The EdU incorporation assay was conducted to evaluate cell proliferation. Cells were seeded into 96-well plates and treated with ZnPPIX/Hemin. The following day, the cells were cultured with EdU reagent (diluted 1:1000, RiboBio, China) for 2 h. Subsequently, the cells were fixed with 4% paraformaldehyde and stained with fluorescent dyes and Hoechst. The percentage of EdU positive cells was then analyzed using Image J software.
Transwell assay
Primary ovarian cancer cells, characterized by uniform growth and normal morphology, were digested with trypsin and resuspended in serum-free medium for subsequent experimentation. Matrigel along with the cell suspension, was introduced into the upper chamber of the Transwell apparatus, while A medium containing 20% serum was added to the lower chamber. Following a 24-hour culture period, the invading cells were fixed, stained with crystal violet, and subsequently imaged and quantified using ImageJ.
Statistical analysis
Statistical differences between the two groups were assessed using a t-test. For comparisons involving more than two groups, one-way ANOVA or two-way ANOVA were employed. Kaplan-Meier survival curves and log-rank tests, as well as Spearman correlation analyses, were conducted to evaluate relationships between two variables. Detailed statistical analysis methods for online databases can be accessed on the corresponding website. A statistically significant difference was defined as p < 0.05.
Results
The expression of HMOX1 in ovarian cancer
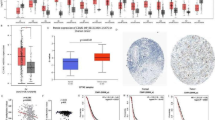
As illustrated in Fig. 1A, the pan-cancer analysis indicates that HMOX1 expression levels are significantly elevated in bladder cancer (BLCA), breast cancer (BRCA), kidney renal clear cell carcinoma (KIRC), kidney renal papillary cell carcinoma (KIRP), thyroid carcinoma (THCA), and uterine corpus endometrial carcinoma (UCEC) compared to their matched adjacent tissues. In contrast, a notable decrease in HMOX1 expression was observed in several human cancers, including colorectal adenocarcinoma (COAD), esophageal carcinoma (ESCA), liver hepatocellular carcinoma (LIHC), lung adenocarcinoma (LUAD), lung squamous cell carcinoma (LUSC), rectum adenocarcinoma (READ), and skin cutaneous melanoma (SKCM). To further investigate the expression profile of HMOX1, we analyzed the GSE40595 dataset, which revealed that HMOX1 expression in ovarian cancer stroma was higher than that in normal ovarian stroma (Fig. 1B). However, no significant difference in HMOX1 expression were detected across all stages of ovarian cancer (Fig. 1C). The prognosis of ovarian cancer patients is significantly influenced by chemotherapy resistance. We conducted an analysis of HMOX1 expression in both drug-resistant and sensitive tumor cells. In the GSE180687 cohort of ovarian cancer patients treated with anti-VEGF therapy, our findings demonstrated that drug-resistant endothelial cells exhibited elevated HMOX1 expression compared to their sensitive counterparts (Fig. 1D). Similarly, within the GSE172016 cohort, paclitaxel-resistant TOV21G cells displayed higher levels of HMOX1 expression than sensitive cells (Fig. 1E). Furthermore, HMOX1 was associated with the immunoreactive and mesenchymal subtypes, but not with the C1 subtype (Fig. 1F). To ascertain the specific cellular localization of the HMOX1 protein, we employed subcellular localization techniques, which revealed that HMOX1 is predominantly expressed on the plasma membrane, cell membrane, and Golgi apparatus (Fig. 1G).

The gene expression of HMOX1. (A) The expression levels of HMOX1 across various pan-carcinoma datasets. The datasets GSE40595 (B), GSE180687 (D), andGSE172016 (E) were used to investigate the expression of HMOX1 in ovarian cancer tissues and drug-resistant cells. (C) The correlation between HMOX1 expression and the pathological stages of ovarian cancer. (F-G) Expression of HMOX1 across different molecular and immune subtypes. (H) Subcellular localization of HMOX1 in the COMPARTMENTS database. “*” p < 0.05, “**” p < 0.01 and “***” p < 0.001
Survival analysis
The Kaplan-Meier curve illustrated that higher expression levels of HMOX1, determined by the optimal cut-off value, were associated with poorer overall survival outcomes in both the entire cohort of ovarian cancer patients and specifically in those with serous ovarian cancer (Fig. 2A-B). Furthermore, we evaluated the expression of HMOX1 in relation to disease-specific survival. The Kaplan-Meier analysis indicated a significant correlation between elevated HMOX1 levels and reduced progression-free survival (PFS) as well as post-progression survival (PPS) in patients diagnosed with serous ovarian cancer (Fig. 2C-D).

The survival analysis of HMOX1 in ovarian cancer. A-B The prognostic potential of HMOX1 expression on the overall survival (OS) rates of all ovarian cancer patients and specifically those with serous ovarian cancer. C-D The correlation between HMOX1 expression and both progression-free survival (PFS) and post-progression survival (PPS) in patients diagnosed with serous ovarian cancer
Co-expression analysis of ferroptosis related genes and HMOX1
A total of 428 ferroptosis-related genes were retrieved from the ferrDb database, encompassing drivers, inhibitors, and markers. The co-expression between HMOX1 and ferroptosis-related genes was evaluated using Pearson’s correlation coefficient. Among the genes analyzed, 70 exhibited a correlation (|Cor| > 0.3, P < 0.05), with the top 20 genes displayed in Figure S1A as a heatmap. TLR4 (R = 0.80, p = 0), CYBB (R = 0.79, p = 0), and TGFB1 (R = 0.67, p = 0) were identified as the most correlated genes. Several ferroptosis-related genes have been implicated in the progression of ovarian cancer (OC) progression. To further explore this association, we examined the expression of the top 20 genes in OC. The results indicated that 13 genes were upregulated while 7 genes were downregulated in OC (Figure S1B).
HMOX1 is implicated in the inflammatory and immune regulatory biological processes associated with ovarian cancer
Patients with ovarian cancer were categorized into two groups based on HMOX1 expression, followed by an analysis of differentially expressed genes (DEGs). The results indicated the presence of 306 upregulated genes and 5 downregulated genes (Fig. 3A-B). Subsequent analyses were conducted to investigate the functions of these DEGs. Gene Ontology (GO) analysis demonstrated that HMOX1 is involved in various biological processes, including the activation and proliferation of immune cells, cell adhesion, and signaling pathways modulated by the extracellular matrix (Fig. 3C). Furthermore, KEGG analysis corroborated that these genes are primarily associated with the inflammatory response and immune regulation (Fig. 3D). These findings suggest that HMOX1 may play a significant role in the immunoregulation of ovarian cancer and contribute to the abnormalities and heterogeneity observed in immune cell infiltration.

A functional enrichment analysis associated with HMOX1. A-B volcano plot and heatmap illustrating differentially expressed genes (DEGs) in two HMOX1 subgroups. C A bubble plot of the top 20 KEGG pathways enriched for DEGs. D The top 20 biological process gene ontology (GO) terms derived from DEGs
HMOX1 contributes to the infiltration of tumor-suppressive immune cells in OC
To investigate the relationship between HMOX1 mRNA expression and immune cell infiltration, a dataset from The Cancer Genome Atlas (TCGA) focused on OC was utilized. The analysis revealed a negative correlation between HMOX1 and tumor purity (Rho = -0.561; p = 2.11E-41) (Fig. 4A). In contrast, a strong positive correlation was observed between HMOX1 expression and the infiltration of five immune cell populations: neutrophils (Rho = 0.407, p = 1.49E-20), dendritic cells (Rho = 0.371, p = 4.4E-17), macrophages (Rho = 0.36, p = 4.32E-16), CD8 + T cells (Rho = 0.256, p = 1.21E-08), and CD4 + T cells (Rho = 0.239, p = 1.17E-07), with correlation coefficients indicating a range from large to small. Furthermore, the TISIDB database identified significant correlations between HMOX1 and all 22 immune subtypes (Figure S2). The four subtypes exhibiting the strongest correlations were T regulatory cells (Treg) (Rho = 0.779, p < 2.2E-16), macrophages (Rho = 0.768, p < 2.2E-16), myeloid-derived suppressor cells (MDSC) (Rho = 0.75, p < 2.2E-16), and follicular helper T cells (Tfh) (Rho = 0.72, p < 2.2E-16) (Fig. 4B-E). These results suggest that elevated HMOX1 expression may promote the infiltration of suppressive immune cells in OC.

The correlation between HMOX1 expression and immune cell infiltration. A The relationship between HMOX1 levels and both immune infiltration level and tumor purity as analyzed through the TIMER database. B-E The correlations between HMOX1 levels and the infiltration of T regulatory cells (Tregs), macrophages, myeloid-derived suppressor cells (MDSCs), and T follicular helper cells (Tfh) as assessed in the TISCH2 database
HMOX1 expression across various cell types was evaluated using single-cell RNA sequencing (scRNA-seq) data from the Tumor Immune Single-Cell Hub 2 (TISCH2). In the datasets EMTAB8107, GSE115007, GSE118828, GSE130000, GSE147082, GSE151214, GSE154600, GSE154763, and GSE158722, it was found that macrophages exhibited the highest levels of HMOX1 expression, followed by dendritic cells (Figure S3).
The expression of HMOX1 correlates with immune checkpoints in ovarian cancerWhile immune checkpoint therapies have demonstrated efficacy across various tumor types, they have not been effective for ovarian cancers. Consequently, we conducted an analysis to evaluate the correlation between HMOX1 expression and 24 immunoinhibitors, 46 immunostimulators, and 21 MHC molecules. Our findings indicate a strong association between elevated levels of HMOX1 and 19 immunoinhibitors (Fig. 5A). The top four immunoinhibitors identified are HAVCR2 (Rho = 0.814, p < 2.2E-16), CSF1R (Rho = 0.768, p < 2.2E-16), PDCD1LG2 (Rho = 0.644, p < 2.2E-16), and CD244 (Rho = 0.628, p < 2.2E-16) (Fig. 5B-E).

The association between HMOX1 expression and various immunoinhibitors. A Distribution of Spearman correlation coefficients between HMOX1 expression (Y-axis) and immunoinhibitors across different human cancers (X-axis). The correlation between HMOX1 expression and (B)HAVCR2, (C)CSF1R, (D)PDCD1LG2, (E)CD244 in ovarian cancer
Notably, high levels of HMOX1 are strongly correlated with 34 immunostimulators (Figure S4A). The top four immunostimulators identified are CD86 (Rho = 0.79, p < 2.2E-16), CD48 (Rho = 0.729, p < 2.2E-16), TNFSF13B (Rho = 0.685, p < 2.2E-16), and IL2RA (Rho = 0.634, p < 2.2E-16) (Figure S4B-E).
High levels of HMOX1 expression are strongly correlated with 19 major histocompatibility complex (MHC) molecules (Fig. 6A). The top four MHC molecules identified are HLA-DPB1 (Rho = 0.57, p < 2.2E-16), HLA-DPA1 (Rho = 0.563, p < 2.2E-16), HLA-DRA (Rho = 0.551, p < 2.2E-16), and HLA-DQA1 (Rho = 0.52, p < 2.2E-16) (Fig. 6B-E).
HMOX1 contributes inflammation regulation in ovarian cancer
To investigate the inflammatory regulation role of HMOX1 in ovarian cancer, we conducted correlation analysis on 41 chemokines and 18 receptors. Our analysis revealed that 29 chemokines and 12 receptors showed a strong correlation with HMOX1 expression (Figure S6A-B). The top four chemokines that exhibited the highest correlation were CCL4 (Rho = 0.629, p < 2.2E-16), CCL3 (Rho = 0.591, p < 2.2E-16), CCL5 (Rho = 0.576, p < 2.2E-16), and CCL2 (Rho = 0.544, p < 2.2E-16) (Fig. 7A-D). Furthermore, among the chemokine-chemokine receptors, CCR1 (Rho = 0.781, p < 2.2E-16), CCR5 (Rho = 0.682, p < 2.2E-16), CCR2 (Rho = 0.58, p < 2.2E-16), and CXCR6 (Rho = 0.56, p < 2.2E-16) exhibited the strongest association (Fig. 7E-H).

The analysis of chemokines and receptors associated with HMOX1 expression. The correlation between HMOX1 expression and various chemokines: (A) CCL4, (B) CCL3, (C) CCL5, and (D) CCL2 in ovarian cancer. The relationship between HMOX1 expression and chemokine receptors (E) CCR1, (F) CCR2, (G) CCR5, and (H) CXCR6
HMOX1 knockdown enhances apoptosis while inhibiting cell proliferation and invasion
Immunohistochemical staining was performed on ovarian cancer samples and healthy controls (n = 10 for both groups) to evaluate HMOX1 expression. The results indicated that HMOX1 expression was significantly higher in ovarian cancer samples compared to healthy controls (Fig. 6A). Following this, we conducted experiments to explore the impact of HMOX1 on ovarian cancer cells. Initially, primary ovarian cancer cells treated with ZnPPIX/Hemin underwent an EdU incorporation assay (Fig. 6B, C). The results demonstrated that the inhibition of HMOX1 (ZnPPIX group) significantly reduced the proliferation of ovarian cancer cells compared to both the control and overexpression groups (Hemin group). Furthermore, flow cytometry analysis revealed that inhibiting HMOX1 markedly increased overall apoptosis and necrosis of ovarian cancer cells (Fig. 6D, E). Additionally, in the transwell assay, we observed a significant reduction in the invasion capacity of ovarian cancer cells upon HMOX1 inhibition (ZnPPIX group) compared to the control and overexpression groups (Hemin group) (Fig. 6F, G). These findings underscore the substantial influence of HMOX1 the proliferation, apoptosis, and invasion of ovarian cancer cells.

The effect of HMOX1 on the proliferation, apoptosis and invasion of ovarian cancer cells. (A) The expression of HMOX1 in both ovarian cancer and normal tissues was assessed using immunohistochemistry (IHC). (B, C) The proliferation of ovarian cancer cells was evaluated through the EdU incorporation assay. (D, E) Apoptosis was measured using flow cytometry. (F, G) The Transwell invasion assay was employed to assess the invasion capability of ovarian cancer cells. Statistical significance is indicated as *P < 0.05; **P < 0.01; ***P < 0.001
Prediction of Small Molecular Drugs Targeting HMOX1
In addition, we predicted small molecule drugs targeting HMOX1 using the DrugBank database. A total of nine small molecule drugs were identified, including NADH, Formic Acid, Biliverdine IX Alpha, 12-Phenylheme, 2-Phenylheme, Verdoheme, Stannsoporfin, 1-({2-[2-(4-CHLOROPHENYL)ETHYL]-1,3-DIOXOLAN-2-YL}METHYL)-1 H-IMIDAZOLE and 1-(adamantan-1-yl)-2-(1 H-imidazol-1-yl)ethanone. Among these, NADH, Formic Acid, and Biliverdine IX Alpha exhibit multiple targets, whereas the remaining compounds specifically target HMOX1 (Table 1).
Discussion
HMOX1, an enzyme known to induce stress, has been found to be highly expressed in various gynecological cancers including ovarian, cervical, and endometrial cancer. Our study demonstrated that HMOX1 expression in ovarian cancer interstitial tissue is higher compared to normal tissue. Furthermore, elevated HMOX1 levels were observed in the immune response and mesenchymal subtypes of ovarian cancer, as well as in ovarian cancer cells exhibiting anti-angiogenesis and taxol resistance phenotypes. These findings suggest that increased HMOX1 expression contributes to the aggressiveness of ovarian cancer and reduces sensitivity to chemotherapy, potentially impacting prognosis.
In a separate study, HMOX1 was identified as an independent prognostic factor for recurrence and progression in non-muscle invasive bladder cancers [18]. Our research also confirmed that high HMOX1 expression in ovarian cancer patients is associated with poor prognosis. Specifically, we found that HMOX1 adversely affects overall survival, progression-free survival, and disease-free survival in patients with serous cancer, which accounts for more than half of ovarian carcinomas. Notably, HMOX1 shows promise as a prognostic factor for both TP53 mutant and wild-type patients, as TP53 mutation is the most common genomic alteration in the development of human ovarian carcinoma.
Recent advances in ovarian cancer (OC) research indicate that targeting ferroptosis may serve as an effective therapeutic strategy [19]. However, there is remains a paucity of relevant studies concerning the regulation of ferroptosis in OC by HMOX1, a crucial regulator of this process. Our study demonstrates that HMOX1 can promote cancer development by interacting with various ferroptosis suppressors and drivers, including TLR4, CYBB, and TGFB1. For instance, Tat San Lau et al. revealed that paclitaxel induces the death of immunogenic cells in ovarian cancer through a TLR4/IKK2/SNARE-dependent exocytosis mechanism [20]. De Almeida Chuffa Luiz Gustavo found that P-MAPA-mediated immunotherapy enhances the efficacy of cisplatin on serous ovarian cancer by targeting the TLR4 signaling pathway [21]. Lecker LSM discovered a significant correlation between TGFB1 expression and alternately activated macrophages in high-grade serous ovarian cancer, as evidenced by transcriptomics data. In vitro studies show that fibroblasts in the metastatic foci of high-grade serous ovarian cancer (HGSOC) express TGFB1 and macrophages to produce additional TGFB1. Treatment of mouse HGSOC tumors in situ with an anti-TGFB1 antibody can reduce the size of peritoneal tumors, increase tumor-associated monocytes, and activate unconventional T cells expressing TGFβ3 [22]. Furthermore, LY2157299, an inhibitor of the TGF-β1 signaling pathway, has demonstrated significant inhibition of ovarian cancer cell proliferation, metastasis, and epithelial-mesenchymal transition (EMT) induced by KDM6B [23]. These findings provide valuable insights into the potential mechanisms by which HMOX1 regulates ferroptosis in ovarian cancer.
This study highlights the multifaceted role of HMOX1 in modulating immune and inflammatory responses within the tumor microenvironment (TME), which is critical for cancer progression. HMOX1, encoding heme oxygenase-1 (HO-1), influences the TME by enhancing the infiltration of immunosuppressive cells such as macrophages and myeloid-derived suppressor cells (MDSCs) [24, 25]. These cells contribute to a suppressive immune milieu that diminishes T cell responses, thereby facilitating tumor growth and disease progression. The expression of HO-1 in tumor-associated macrophages (TAMs) may promote the formation of M2-polarized TAMs, which further support tumor proliferation and metastasis through mechanisms such as iron release and angiogenesis. Under oxidative stress, HO-1 induction mitigates reactive oxygen species (ROS) production, potentially modulating immune cell activity and TME infiltration [26, 27]. While initial HO-1 upregulation might protect normal cells from ROS-induced damage, preventing cellular transformation, increased HO-1 expression in established tumors could aid malignant cells in countering ROS toxicity, thus promoting tumor growth and dissemination [28]. Additionally, HO-1’s antioxidant properties may confer resistance to cancer therapies, suggesting that HO-1 inhibition could sensitize cancer cells to treatments such as chemotherapy and radiation [29]. The complex regulatory network of HO-1 in cancer, including its impact on immune cell infiltration, underscores its potential as a therapeutic target, necessitating strategies that could exploit this network for cancer treatment.
The therapeutic application of HMOX1 in ovarian cancer treatment necessitates a profound understanding of its mechanisms in both inflammatory and non-inflammatory tumor microenvironments. Recognized as a key modulator of inflammatory responses across various diseases, HMOX1’s expression correlates with multiple cytokines and receptor signaling pathways involved in inflammation regulation, particularly within the ovarian cancer microenvironment [30]. This correlation underscores the significance of HMOX1 in modulating inflammation in inflammatory ovarian cancer. In non-inflammatory ovarian cancer, HMOX1 may exert protective effects by reducing oxidative damage and promoting cellular repair, thereby inhibiting tumor progression [30]. Furthermore, the significant association of HMOX1 with immune modulators and MHC molecules suggests that its upregulation could enhance the expression of immune checkpoints, potentially improving the response to immunotherapy in patients with high HMOX1 expression levels. Future research should elucidate the expression and function of HMOX1 across different ovarian cancer subtypes, as well as its interaction with inflammation and immune responses, with clinical trials to validate HMOX1 as a biomarker and its predictive value in immune checkpoint inhibitor therapy. This multifaceted research approach will aid in developing novel treatment strategies that leverage the regulatory role of HMOX1 effectively, regardless of the presence or absence of inflammation in ovarian cancer patients.
Amidst the intricate biology of cancer, and notably in ovarian cancer, an array of pivotal mutational signatures such as BRCA1/2, the HRR gene cluster (comprising ATM, BARD1, and others), TP53, and PIK3CA not only propel disease onset and progression but also profoundly influence therapeutic responsiveness and clinical prognosis [31]. BRCA1/2 mutations elevate ovarian cancer risk and alter chemotherapy sensitivity, while HRR mutations induce homologous recombination deficiencies, enhancing sensitivity to platinum agents and PARP inhibitors [28]. Despite the absence of direct targeted therapies, the prevalent TP53 mutations remain a crucial biomarker for assessing tumour progression. PIK3CA, on the other hand, activates the PI3K/AKT/mTOR pathway, fostering aggressive tumour behaviour, with accelerated efforts underway to develop targeted agents. Moreover, inhibitors targeting the RAS-RAF-MEK-ERK pathway have demonstrated remarkable efficacy, particularly when BRAF and MEK inhibitors are combined, exhibiting synergistic effects that promise to delay the emergence of resistance [32]. Concurrently, the overexpression of HMOX1 in ovarian cancer emerges as a pivotal factor, intricately linked to tumour malignancy, prognosis, and the modulation of the immune microenvironment. By influencing immune cell infiltration, particularly the function of tumour-associated macrophages (TAMs), and regulating the levels of immunosuppressors, immunostimulators, and MHC molecules, HMOX1 exerts a substantial impact on tumour progression. Functional studies of HMOX1 underscore its pivotal role in directly modulating cellular apoptosis, proliferation, and invasion capabilities, with its expression fluctuations dictating these cellular processes. Collectively, these discoveries underscore the paramount importance of individualised therapeutic strategies tailored to tumour genomic profiles. Future research endeavors must delve deeper into the understanding of these mutational landscapes and the multifaceted role of HMOX1, expediting the development of novel therapeutics, particularly for patients who exhibit treatment resistance. The relentless pursuit of innovative strategies to overcome drug resistance holds the key to optimising treatment outcomes and ultimately improving the prognosis of ovarian cancer patients.
Finally, we investigated the potential of small molecule drugs targeting HMOX1 for the treatment of ovarian cancer. These drugs provide additional therapeutic options; however, further foundational experiments are necessary to validate their efficacy.
The study’s strengths lie in its comprehensive approach to understanding HMOX1’s role in ovarian cancer, robust use of TCGA data, validation through immunohistochemistry, and the application of advanced bioinformatics tools. In vitro functional studies provide direct evidence of HMOX1’s impact on cancer cell behavior. However, limitations include a potential lack of clinical correlation, limited generalizability to other ovarian cancer subtypes, unaddressed confounding factors, absence of in vivo studies, and a superficial exploration of HMOX1’s immunotherapeutic potential. The study provides valuable insights but requires further research to fully elucidate HMOX1’s clinical implications and therapeutic potential.
Conclusion
Our study establishes HMOX1 as a pivotal biomarker and therapeutic target in ovarian cancer, underscored by its profound influence on tumor malignancy, immune cell dynamics, and patient prognosis, offering a promising avenue for advancing ovarian cancer treatment strategies.
Data availability
All the data are available from online databases, link to the website are presented in the manuscript.
References
Kotrbova A, Ovesna P, Gybel’ T, Radaszkiewicz T, Bednarikova M, Hausnerova J, Jandakova E, Minar L, Crha I, Weinberger V, et al. WNT signaling inducing activity in ascites predicts poor outcome in ovarian cancer. THERANOSTICS. 2020;10(2):537–52.
Ji M, Zhao Z, Li Y, Xu P, Shi J, Li Z, Wang K, Huang X, Ji J, Liu W, et al. FBXO16-mediated hnRNPL ubiquitination and degradation plays a tumor suppressor role in ovarian cancer. CELL DEATH DIS. 2021;12(8):758.
Masiello T, Dhall A, Hemachandra L, Tokranova N, Melendez JA, Castracane J. A dynamic culture method to produce ovarian Cancer spheroids under physiologically-relevant Shear stress. CELLS-BASEL. 2018;7(12).
Cheng S, Xu C, Jin Y, Li Y, Zhong C, Ma J, Yang J, Zhang N, Li Y, Wang C, et al. Artificial Mini dendritic cells boost T cell-based immunotherapy for ovarian Cancer. ADV SCI. 2020;7(7):1903301.
Verma N, Vinik Y, Saroha A, Nair NU, Ruppin E, Mills G, Karn T, Dubey V, Khera L, Raj H et al. Synthetic lethal combination targeting BET uncovered intrinsic susceptibility of TNBC to ferroptosis. SCI ADV. 2020;6(34).
Yu Y, Yan Y, Niu F, Wang Y, Chen X, Su G, Liu Y, Zhao X, Qian L, Liu P, et al. Ferroptosis: a cell death connecting oxidative stress, inflammation and cardiovascular diseases. CELL DEATH DISCOV. 2021;7(1):193.
Ascenzi F, De Vitis C, Maugeri-Sacca M, Napoli C, Ciliberto G, Mancini R. SCD1, autophagy and cancer: implications for therapy. J EXP CLIN CANC RES. 2021;40(1):265.
Fang K, Du S, Shen D, Xiong Z, Jiang K, Liang D, Wang J, Xu H, Hu L, Zhai X, et al. SUFU suppresses ferroptosis sensitivity in breast cancer cells via Hippo/YAP pathway. ISCIENCE. 2022;25(7):104618.
Ji P, Wang X, Yin J, Yao Y, Du W. Amplification of ferroptosis with a liposomal nanoreactor cooperates with low-toxicity doxorubicin apoptosis for enhanced tumor chemotherapy. BIOMATER SCI-UK. 2022;10(6):1544–53.
Fernandez-Fierro A, Funes SC, Rios M, Covian C, Gonzalez J, Kalergis AM. Immune Modulation by inhibitors of the HO system. INT J MOL SCI. 2020;22(1).
Krukowska K, Magierowski M. Carbon monoxide (CO)/heme oxygenase (HO)-1 in gastrointestinal tumors pathophysiology and pharmacology - possible anti- and pro-cancer activities. BIOCHEM PHARMACOL. 2022;201:115058.
Li S, Qiu B, Lu H, Lai Y, Liu J, Luo J, Zhu F, Hu Z, Zhou M, Tian J, et al. Hyperhomocysteinemia accelerates acute kidney Injury to chronic kidney Disease Progression by Downregulating Heme Oxygenase-1 expression. ANTIOXID REDOX SIGN. 2019;30(13):1635–50.
Huang Y, Yang Y, Xu Y, Ma Q, Guo F, Zhao Y, Tao Y, Li M, Guo J. Nrf2/HO-1 Axis regulates the angiogenesis of gastric Cancer via Targeting VEGF. CANCER MANAG RES. 2021;13:3155–69.
Liu ZM, Chen GG, Ng EK, Leung WK, Sung JJ, Chung SC. Upregulation of heme oxygenase-1 and p21 confers resistance to apoptosis in human gastric cancer cells. Oncogene. 2004;23(2):503–13.
Yang S, Zhang D, Shen N, Wang G, Tang Z, Chen X. Dihydroartemisinin increases gemcitabine therapeutic efficacy in ovarian cancer by inducing reactive oxygen species. J CELL BIOCHEM. 2019;120(1):634–44.
Lanczky A, Gyorffy B. Web-based Survival Analysis Tool tailored for Medical Research (KMplot): development and implementation. J MED INTERNET RES. 2021;23(7):e27633.
Zhao E, Maj T, Kryczek I, Li W, Wu K, Zhao L, Wei S, Crespo J, Wan S, Vatan L, et al. Cancer mediates effector T cell dysfunction by targeting microRNAs and EZH2 via glycolysis restriction. NAT IMMUNOL. 2016;17(1):95–103.
Yim MS, Ha YS, Kim IY, Yun SJ, Choi YH, Kim WJ. HMOX1 is an important prognostic indicator of nonmuscle invasive bladder cancer recurrence and progression. J Urol. 2011;185(2):701–5.
Tan Z, Huang H, Sun W, Li Y, Jia Y. Current progress of ferroptosis study in ovarian cancer. FRONT MOL BIOSCI. 2022;9:966007.
Lau TS, Chan L, Man G, Wong CH, Lee J, Yim SF, Cheung TH, McNeish IA, Kwong J. Paclitaxel induces immunogenic cell death in Ovarian Cancer via TLR4/IKK2/SNARE-Dependent exocytosis. CANCER IMMUNOL RES. 2020;8(8):1099–111.
de Almeida CL, de Moura FG, Lupi LA, Da SNI, Favaro WJ. P-MAPA immunotherapy potentiates the effect of cisplatin on serous ovarian carcinoma through targeting TLR4 signaling. J OVARIAN RES. 2018;11(1):8.
Lecker L, Berlato C, Maniati E, Delaine-Smith R, Pearce O, Heath O, Nichols SJ, Trevisan C, Novak M, McDermott J, et al. TGFBI Production by macrophages contributes to an immunosuppressive microenvironment in Ovarian Cancer. CANCER RES. 2021;81(22):5706–19.
Liang S, Yao Q, Wei D, Liu M, Geng F, Wang Q, Wang YS. KDM6B promotes ovarian cancer cell migration and invasion by induced transforming growth factor-beta1 expression. J CELL BIOCHEM. 2019;120(1):493–506.
Li Y, Zhang Q, Wu M, Zhang P, Huang L, Ai X, Yang Z, Shen Q, Wang Y, Wang P, et al. Suppressing MDSC infiltration in Tumor Microenvironment serves as an option for treating Ovarian Cancer Metastasis. INT J BIOL SCI. 2022;18(9):3697–713.
Pi R, Yang Y, Hu X, Li H, Shi H, Liu Y, Wang X, Tong A, Lu T, Wei Y, et al. Dual mTORC1/2 inhibitor AZD2014 diminishes myeloid-derived suppressor cells accumulation in ovarian cancer and delays tumor growth. CANCER LETT. 2021;523:72–81.
Cuitino L, Obreque J, Gajardo-Meneses P, Villarroel A, Crisostomo N, San FI, Valenzuela RA, Mendez GP, Llanos C. Heme-Oxygenase-1 is decreased in circulating monocytes and is Associated with impaired phagocytosis and ROS production in Lupus Nephritis. FRONT IMMUNOL. 2019;10:2868.
Chiang SK, Chen SE, Chang LC. The role of HO-1 and its crosstalk with oxidative stress in Cancer Cell Survival. CELLS-BASEL. 2021;10(9).
Consoli V, Sorrenti V, Grosso S, Vanella L. Heme Oxygenase-1 Signaling and Redox Homeostasis in Physiopathological Conditions. BIOMOLECULES. 2021;11(4).
Glynn SA, Boersma BJ, Howe TM, Edvardsen H, Geisler SB, Goodman JE, Ridnour LA, Lonning PE, Borresen-Dale AL, Naume B, et al. A mitochondrial target sequence polymorphism in manganese superoxide dismutase predicts inferior survival in breast cancer patients treated with cyclophosphamide. CLIN CANCER RES. 2009;15(12):4165–73.
Campbell NK, Fitzgerald HK, Dunne A. Regulation of inflammation by the antioxidant haem oxygenase 1. NAT REV IMMUNOL. 2021;21(7):411–25.
Wang ZC, Birkbak NJ, Culhane AC, Drapkin R, Fatima A, Tian R, Schwede M, Alsop K, Daniels KE, Piao H, et al. Profiles of genomic instability in high-grade serous ovarian cancer predict treatment outcome. CLIN CANCER RES. 2012;18(20):5806–15.
Perrone C, Angioli R, Luvero D, Giannini A, Di Donato V, Cuccu I, Muzii L, Raspagliesi F, Bogani G. Targeting BRAF pathway in low-grade serous ovarian cancer. J GYNECOL ONCOL. 2024;35(4):e104.
Acknowledgements
Not applicable.
Funding
This work was supported by Shunde Hospital, Southern Medical University (SRSP2021004)and Health Commission of Guangdong Province (B2023368).
Author information
Authors and Affiliations
Contributions
Ruiwan Tan conceptualized and supervised the study, reviewed & edited the manuscript, Jinfa Huang performed the methodology, software analysis and validation, as well as original draft preparation.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The conduct of this study adhered strictly to the principles outlined in the Declaration of Helsinki. Prior to commencing the study, ethical approval was granted by the Ethics Committee of Shunde Hospital, Southern Medical University. Furthermore, informed consent was obtained from all participants, ensuring their voluntary participation and understanding of the study’s objectives and procedures.
Consent for publication
Not Applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplementary Material 1: Figure. S1.
Co-expression analysis of HOMX1 in OC. A Heatmap of correlation coefficient of top 20 ferroptosis related genes with HMOX1. The color depth represented the correlation degree. Red was positively correlated and blue was negatively correlated. B Expression of the top 20 correlated genes with HMOX1 in OC and normal tissues
Supplementary Material 2: Figure. S2.
Correlation between HMOX1 expression and infiltration abundance of 22 immune cell subtypes in different cancers. The abscissa is different cancer types, and the ordinate is different immune cell subtypes.
Supplementary Material 3: Figure. S3.
Expression of HMOX1 in different ovarian cancer cells in single cell sequencing data of GEO database. X-axis: cell types; Y-axis: datasets
Supplementary Material 4: Figure. S4.
The association between HMOX1 expression and various immunostimulators. A The distribution of Spearman correlations between HMOX1 expression (Y-axis) and immunostimulators across human cancers (X-axis). The correlation between HMOX1 expression and (B) CD86, (C) CD48, (D) TNFSF13B, (E) IL2RA in ovarian cancer
Supplementary Material 5: Figure. S5.
The association of HMOX1 expression with MHC molecules. A Distribution of Spearman correlation between HMOX1 expression and MHC molecules (Y axis) across human cancer (X axis). The correlation between HMOX1 expression and (B)HLA-DPB1,(C)HLA-DPA1, (D)HLA-DRA, (E)HLA-DQA1 in ovarian cancer.
Supplementary Material 6: Figure. S6.
Correlation between HMOX1 expression and chemokines and receptors in different cancers. A Chemokines; B Chemokine receptors.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Cite this article
Huang, J., Tan, R. HMOX1: A pivotal regulator of prognosis and immune dynamics in ovarian cancer. BMC Women's Health 24, 476 (2024). https://doi.org/10.1186/s12905-024-03309-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12905-024-03309-3