
Abstract
Background
Adjuvant therapies such as radiation therapy, chemotherapy, and immunotherapy are usually given after cancer surgery to improve the survival of cancer patients. However, despite advances in several adjuvant therapies, they are still limited in the prevention of recurrences.
Methods
We evaluated the immunological effects of RNA-based adjuvants in a murine melanoma model. Single-stranded RNA (ssRNA) were constructed based on the cricket paralysis virus (CrPV) internal ribosome entry site (IRES). Populations of immune cells in bone marrow cells and lymph node cells following immunization with CrPVIRES-ssRNA were determined using flow cytometry. Activated cytokine levels were measured using ELISA and ELISpot. The tumor protection efficacy of CrPVIRES-ssRNA was analyzed based on any reduction in tumor size or weight, and overall survival.
Results
CrPVIRES-ssRNA treatment stimulated antigen-presenting cells in the drain lymph nodes associated with activated antigen-specific dendritic cells. Next, we evaluated the expression of CD40, CD86, and XCR1, showing that immunization with CrPVIRES-ssRNA enhanced antigen presentation by CD8a+ conventional dendritic cell 1 (cDC1), as well as activated antigen-specific CD8 T cells. In addition, CrPVIRES-ssRNA treatment markedly increased the frequency of antigen-specific CD8 T cells and interferon-gamma (IFN-γ) producing cells, which promoted immune responses and reduced tumor burden in melanoma-bearing mice.
Conclusions
This study provides evidence that the CrPVIRES-ssRNA adjuvant has potential for use in therapeutic cancer vaccines. Moreover, CrPVIRES-ssRNA possesses protective effects on various cancer cell models.
Similar content being viewed by others

Introduction
The ideal vaccine protects against infection by inducing a long-term immunological memory response to the target antigen [1]. However, some vaccines can fail to elicit a sufficient immune response and, therefore, cannot protect against infection or disease recurrence. Various immune stimulants, namely adjuvants, have been developed that increase both innate and humoral responses [2]. Immune stimulation can generally be achieved through three signaling mechanisms [3, 4]: 1) Increased antigen presentation and antigen uptake [5,6,7] by allowing the interaction of pattern of recognition receptors (PRRs) such as Toll-like receptors (TLRs), retinoic acid-inducible gene I (RIG-I)-like receptors (RIG-Is) & NOD-like receptors, with pattern associated molecular pathogens [8,9,10,11]; 2) Activation of the co-stimulatory molecules of antigen-presenting cells (APCs), such as CD40, CD80, and CD86 cells [11, 12]; 3) Stimulation of naïve T cells to differentiate into Th1/Th2 cells and the modulation of antigen-specific antibody responses via cytokine activation and PRR stimulation [13,14,15,16].
Aluminum (alum)-containing supplements act as exogenous damage-associated molecular patterns (DAMPs), triggering an immune response and causing tissue damage [17]. Aluminum exposure has been shown to induce antibody-mediated responses and the proliferation of CD4 T cells [18]. Alum combined with antigenic proteins enhanced the differentiation of Th2 cells and increased antigen-specific follicular T cells [14]. On the other hand, nucleic acid-based adjuvants generally induce an immune response through the recognition of foreign agents and non-self-nucleic acids through TLRs located on the surface of APCs (TLR1, 2, 4, 5, and 6) and endosomal membranes (TLR3, 7, 8, and 9) [19]. RNA-based adjuvants have been reported to recognize a variety of TLRs, such as TLR3 for dsRNA and TLR7/8 for ssRNA [20, 21]. Poly(I:C) consists of double-stranded RNA, which generally induces antiviral effects by stimulating TLR3 signaling [22]. It also enhances type I interferon and Th1-related cytokines [13, 23], and subsequently activates co-stimulatory molecules on conventional dendritic cells (cDCs), which affects CD8 T cell activation [24,25,26]. However, poly(I:C) has significant drawbacks, including severe side effects, an undefined structure, and a lack of homogeneity [27]. In contrast to poly(I:C), synthetic or natural ssRNA is recognized by TLR7/8 in plasmacytoid dendritic cells (pDCs) which in turn produce type I interferon and proinflammatory cytokines [24, 25]. Activated TLR7/8 may also promote the activation of CD8 and effector T cells [26, 28].
Recently, an ssRNA-based adjuvant, RNA adjuvant®, has been developed. It consists of an ssRNA, a non-coding site, and non-capped RNA sequence containing with poly (U) repeats, with added cationic peptides for stabilization. RNA adjuvant® therapy has been shown to enhance antigen-specific antibody responses and balanced Th1/Th2 cell responses [29]. It also induced immune responses via TLRs, RIG signaling, and MyD88-/ MAVS-dependent mechanisms [29, 30]. The induction of these signaling pathways contributes to the activation of human CD141+ DCs, pDCs, monocytes, and CD8 T cells [31,32,33]. RNA adjuvant® has been incorporated into various types of vaccine delivery systems, including peptide, subunit, and virus-like particle vaccines to induce immune responses similar to poly(I:C) adjuvant [29, 34]. Our group has previously reported that ssRNA-based adjuvant treatment effectively increased the efficacy of various prophylactic vaccines, including inactivated, subunit, virus-like particle, and even live-attenuated vaccines by inducing balanced Th1/Th2 cell responses [35,36,37].
An adjuvant can be used to enhance the immune response of a therapeutic vaccine. The RNA-based adjuvant increased the antibody response, which is an indicator of efficacy for the preventive vaccine, and also increased the antigen-specific T cell anti-tumor immune response [34]. In this study, the cricket paralysis virus internal ribosome entry site (CrPVIRES)-ssRNA was established and administered to mice challenged with murine melanoma cells. The National Center for Health Statistics reported that approximately 90,000 cases of melanoma occur annually in the USA [38]. Melanoma is the most common type of skin cancer, which occurs in melanocytes that produce the melanin responsible for variations in skin color [39]. Melanin is most abundant in the epidermis but is also found in the irises, inside the nose or throat, and throughout the internal organs [40]. Although the etiology of all types of melanomas is not currently elucidated, it is known that sun-induced ultraviolet (UV) radiation promotes the growth of melanoma [41]. Most early-stage melanomas can be treated successfully with surgery; however, this is often combined with chemotherapy or targeted therapy for metastatic melanoma [39]. Unfortunately, the survival rate of patients is still low due to recurrence after treatment, and thus, there is an urgent need for better strategies to treat melanoma. Here, we evaluate the effects of CrPVIRES-ssRNA as an adjuvant therapy that can induce antigen-specific T cell responses. In addition, we identify the immunological mechanisms of CrPVIRES-ssRNA activity and investigate the efficacy of CrPVIRES-ssRNA as an adjuvant in melanoma treatment.
Material and methods
Mice and immunization
Six-week-old female C57BL/6 mice were purchased from Dae-Han Bio-Link (Eumseong, Korea). Mice were maintained under specific pathogen-free conditions with a 12 h light/dark cycle. According to the National Institutes of Health (NIH) guidelines, euthanasia of rodents was performed using carbon dioxide at a fill rate of 30–70% of the chamber volume per minute. All animal studies were conducted according to the guidelines of the Institutional Animal Care at the Catholic University of Korea (CUK-IACUC-2021–025-01).
To evaluate adjuvant therapy APC response in relation to total lymph node cells, mice were immunized intramuscularly with either Alum antigen in a 1:1 ratio (Thermo Fisher Scientific, Waltham, MA, USA), 20 µg poly(I:C) (Sigma Aldrich, St. Louis, MO, USA), or 20 µg ssRNA (n = 3) (Fig. 1). In the APCs activation test, mice were immunized intramuscularly with either saline, 20 µg poly(I:C), 20 µg ssRNA, or 100 µg ssRNA (Fig. 2). To test antigen-specific responses to Ova protein, mice were immunized intramuscularly twice with an interval of two weeks (day 0 and on day 14) with 50 µg Ova protein (Sigma Aldrich) in combination with either 20 µg poly(I:C) or 20 µg ssRNA. Saline injection was used as a control (Fig. 3). In the T-cell activation test, mice were immunized intramuscularly twice with an interval of two weeks (day 0 and on day 14) with either 50 µg Ova protein in isolation, 50 µg Ova protein with Alum antigen in a 1:1 ratio or 50 µg Ova protein with 20 µg ssRNA (Fig. 4).

CrPVIRES-ssRNA increases the number of antigen-presenting cells. C57BL/6 mice were intramuscularly injected with CrPVIRES-ssRNA, alum, and poly(I:C). Immune cells were analyzed in drain inguinal lymph nodes for 24 h. A Total number of cells in draining inguinal lymph nodes according to adjuvant (n = 3). B Total cell percentages of CD11c+ DCs (left) and B220+ B cells (right). C, D Efficacy of antigen uptake using FITC-conjugated Ova 12 h after immunization (n = 6). Quantitative plots (C) and graph (D) of flow cytometry. Data were represented as mean ± SD. Statistical significance was indicated by *p < 0.05, **p < 0.01, and ***p < 0.005. Ova: Ovalbumin

CrPVIRES-ssRNA activates dendritic cells in drain inguinal lymph nodes. A Mouse bone marrow-derived dendritic cells (BMDCs) were stimulated with CrPVIRES-ssRNA or poly(I:C) for 24 h. Cell activation was measured by flow cytometry using antibodies specific to CD40, CD80, and CD86. B Mice were intramuscularly immunized once with poly(I:C) (20 µg) and CrPVIRES-ssRNA (20 µg and 100 µg) for 24 h (n = 5). C Percentages of co-stimulatory molecules (CD40, CD80, and CD86) of CD11c+ DCs in lymphocytes isolated from drain inguinal lymph nodes. D, E Percentages of activation markers of DCs (cDC1, cDC2, pDC) in CD40 + cells (D) and CD80 + cells (E). Dendritic subsets were analyzed by flow cytometry using CD8a+, CD8−, and B220.+. Data were represented as mean ± SD. Statistical significance was indicated by *p < 0.05, **p < 0.01, and ***p < 0.005

Ova protein enhances CrPVIRES-ssRNA-mediated conventional DC 1 activation. A The mice were primed and boosted with Ova protein (50 µg) formulated with CrPVIRES-ssRNA (20 µg) or poly(I:C) (20 µg) (n = 5). Splenocytes and drain inguinal lymph nodes were harvested 1 day after boosting. B Frequency of co-stimulatory CD40, CD80, and CD86 of CD8+ cDC1 cells. C, D XCR1 expression of CD8.+ cDC1 was measured in splenocytes (C) and lymphocytes (D) isolated from drain inguinal lymph nodes. Data were represented as mean ± SD. Statistical significance was indicated by *p < 0.05, **p < 0.01, and ***p < 0.005

CrPVIRES-ssRNA conjugated with Ova protein induces antigen-specific CD8 T cell activation. A Mice (C57BL/6) were intramuscularly immunized twice at an interval of 2 weeks with Ova protein (50 µg) mixed alum (antigen ratio 1:1) or CrPVIRES-ssRNA (20 µg) (n = 5). Splenocytes and drain inguinal lymph nodes were harvested 7 days after boosting. B, C Ova-specific CD8 T cells were assessed using Ova 257–264 tetramer staining in splenocytes. Quantitative plots (B) and graph (C) of flow cytometry. D Frequency of effector T cells and Ki-67 were measured in inguinal lymph nodes. E ELISpot assay for IFN-γ secretion. Splenocytes were stimulated for 2 days with/without Ova-specific CD4 or CD8 T cell peptide. IFN-γ-producing cells were measured using ELISpot assay. Data were represented as mean ± SD. Statistical significance was indicated by *p < 0.05, **p < 0.01, and ***p < 0.005. ELISpot: enzyme-linked immunospot, IFN: interferon
In vitro transcription and RNA purification
The DNA platform was constructed based on the IRES intergenic region and the SV40 late-polyadenylation signal sequences. DNA templates were linearized using Not I (Enzynomics, Daejeon, Korea). For ssRNA production, in vitro transcription was assessed using the EZ T7 High Yield In Vitro Transcription Kit (Enzynomics), according to the manufacturer’s instructions.
Flow cytometry and antibodies
To confirm the presence of the OVA tetramer, splenocytes and the inguinal lymph node lymphocytes of the mice were harvested and blocked using CD16/CD32 (eBioscience, Waltham, MA, USA; Thermo Fisher Scientific Inc.) for 20 min at 4 °C, and then stained with the following antibodies for 20 min at 4 °C: H-2 Kb Ova tetramer with the Ova 257–264 epitope SIINFEKL. This was provided by the National Institutes of Health (NIH), and we followed the manufacturers’ protocol.
For surface staining, splenocytes and isolated immune cells from muscle and the inguinal lymph node were stained with the following antibodies for 15 min; CD8a (clone KT15, MBL), Fixable Viability Dye eFluor520 (eBioscience), CD8 (Clone 53–6.7, Invitrogen Waltham, MA, USA), CD44 (Clone IM7, Invitrogen), and CD62L (Clone MEL 14, BD Biosciences, Waltham, MA, USA). Prior to staining for the transcription factor Ki-67 (clone SolA15; eBioscience), the cells were permeabilized using the Foxp3/Transcription Factor Staining Buffer Set (eBioscience) for 30 min at 4 °C.
To determine the activation of macrophages, dendritic cells, and B and T cells, the isolated splenocytes and lymphocytes were blocked using CD16/CD32 (eBioscience; Thermo Fisher Scientific Inc.) for 20 min at 4 °C, and were stained using Fixable Viability Dye eFluor520 (eBioscience), F4/80 (clone BM8, eBioscience), CD11c (clone N418, eBioscience), CD11b (clone M1/70, eBioscience), Ova-FITC (Thermo Fisher Scientific), CD40 (clone 1C10, eBioscience), CD80 (clone 16-10A1, Biolegend, San Diego, CA, USA), CD86 (clone GL1, Biolegend), XCR1 (clone ZET, Biolegend), CD8a (clone 53–6.7, eBioscience), CD19 (clone 6D5, Biolegend), B220 (clone RA3-6B2, BD Biosciences), CD4 (clone GK1.5, Invitrogen), and CD8a (clone 53–6.7, Invitrogen) for 20 min at 4 °C without exposure to light. Stained cells were fixed with 1% paraformaldehyde. Cells were analyzed using the FACS Aurora (Cytek Biosciences, Fremont, CA, USA), CytoFLEX Flow Cytometer (Beckman Coulter, CA, USA), and the data were analyzed using Spectroflo (Cytek Biosciences), FlowJo (Tree Star, Inc., Ashland, OR, USA), and CytExpert (Beckman Coulter).
Splenocyte cell culture and Enzyme-Linked ImmunoSpot (ELISpot)
Splenocytes were stimulated with 4 µg/well Ova 323–339 CD4-specific T cell (ISQAVHAAHAEINEAGR) and Ova 257–264 CD8-specific T cell epitope peptide (SIINFEKL) for 48 h at 37 °C. The OVA peptides were synthesized using Peptron (Daejeon, Korea). The ELISPOT assays for the detection of IFN-γ-secreting T cells were performed using mouse IFN-γ (Mabtech, Stockholm, Sweden), according to the manufacturer’s instructions.
Preparation of bone marrow-derived dendritic cells (BMDCs)
The cells were prepared as previously described [42]. Bone marrow nucleated cells (1 × 106 cells/ml) were seeded in a modified 5 ml RPMI 1640 (Gibco; Thermo Fisher Scientific Inc.), Waltham, MA, USA) medium containing 10% FBS (Gibco), in 6-well plates. Subsequently, 100 ng/ml mouse recombinant GM-CSF (Granulocyte–Macrophage Colony-Stimulating Factor) (Peprotech, New Jersey, United States) and 50 ng/ml mouse recombinant IL-4 (Peprotech) were added to the medium to stimulate BMDCs.
Mouse cancer challenge
B16-Ova cells (5 × 105 cells) were subcutaneously injected into 6-week-old female C57BL/6 mice on day 0. On day 7 after tumor inoculation, tumor size was measured and the mice were intramuscularly immunized with one of the following: Ova protein (50 µg) in isolation, Ova protein (50 µg) in combination with poly(I:C) (20 µg), or Ova protein (50 µg) in combination with ssRNA (20 µg), at an interval of 2 days (days 9, 11, 13, and 15) to total 5 immunization treatments (n = 5). The mice were sacrificed when the size of tumor exceeded 1 cm3 according to animal ethical standards. On day 31 after the tumor challenge, the weight of the tumors from the sacrificed mice was measured.
Statistical analyses
All data are expressed as the mean ± SD. One-way ANOVA was used to assess the significance of differences among treatment groups; if significant deviations from variance homogeneity were detected using the Levene test, then the nonparametric Kruskal–Wallis H test was performed. The student’s t-test was used to assess significant differences between the two groups; if significant deviations from variance homogeneity were detected using the Levene test, then the nonparametric Mann–Whitney U test was performed. Statistical analyses were conducted using SPSS for Windows (release 14.0 K, IBM, Armonk, NY). Differences were considered significant at values of *p < 0.05, **p < 0.01, and ***p < 0.005.
Results
CrPVIRES-ssRNA recruits immune cells from the inguinal lymph node
The immune response induced by CrPVIRES-ssRNA was assessed using poly(I:C) and alum as positive controls. The immune cells in the drain inguinal lymph nodes were counted at an earlier time point (Fig. 1A, B). The CrPVIRES-ssRNA treated group demonstrated an increase in the number of cells isolated from drain inguinal lymph nodes 24 h after immunization (Fig. 1A, B; Supplementary Fig. S1A). The Poly(I:C) treatment group showed a similar increase in cells isolated from the drain inguinal lymph nodes, while no increase in immune cells was observed in the alum treated group (Fig. 1B). However, the proportion of CD4+/CD8+ T cells in drain lymph nodes did not change significantly in any of the groups (Supplementary Fig. S1B, C). Subsequently, the antigen uptake ability of activated APCs was evaluated using the fluorescein isothiocyanate-conjugated Ova (Ova-FITC). The antigen uptake ability was significantly higher in the CrPVIRES-ssRNA and poly(I:C) treated group than in the alum treated group (Fig. 1C, D). This indicates that CrPVIRES-ssRNA can act as an adjuvant by effectively recruiting antigen-presenting cells and enhancing antigen uptake.
CrPVIRES-ssRNA activates dendritic cells in drain inguinal lymph nodes
To identify the types of APCs recruited by CrPVIRES-ssRNA treatment, activation markers of dendritic cells and macrophages, such as CD40+, CD80+, and CD86+, were measured by flow cytometry. Bone marrow-derived dendritic cells (BMDCs) were treated with CrPVIRES-ssRNA and poly(I:C). The CrPVIRES-ssRNA treatment increased levels of the activation markers in a dose-dependent manner (Fig. 2A; Supplementary Fig. S2A). After we injected poly(I:C) and CrPVIRES-ssRNA intramuscularly without the antigen, inguinal lymph nodes were collected on day 1 (Fig. 2B). In the CrPVIRES-ssRNA-treated group, the expression of the co-stimulatory molecules, CD40+, CD80+, and CD86+, increased in CD11c-positive cells related to dendritic cells regardless of the concentration (Fig. 2C). However, the expression of macrophage activation markers in poly(I:C)- and CrPVIRES-ssRNA-treated groups was not significantly different from that in the control group (Supplementary Fig. S2B). Next, we investigated the dendritic cell types of drain lymph nodes affected by uptake of CrPVIRES-ssRNA treatment. The activated conventional DC1/2 (cDC1 and cDC2) and pDC in drain inguinal lymph nodes were identified using flow cytometry. The CrPVIRES-ssRNA treatment enhanced the activity of the CD8a+ cDC1 activation maker, while differences in the CD8a− cDC2 and pDC markers were insignificant (Fig. 2D, E). The poly(I:C)-treated group also showed a similar pattern (Fig. 2D-E). These results revealed that CrPVIRES-ssRNA primarily increases the activity of CD8a+ cDC1 cells among antigen-presenting cells.
Ovalbumin (Ova) protein formulated with CrPVIRES-ssRNA activates CD8a.+ cDC1
In immunology, Ova is widely used in vaccines as it is a key protein that stimulates immune responses, including allergic reactions [43]. To investigate the detailed mechanism of cDC1 activation by CrPVIRES-ssRNA treatment, we formulated a combination of CrPVIRES-ssRNA or poly(I:C) with Ova protein. After the second immunization (day 0 and day 14) with these constructs in mice, splenocytes and inguinal lymph nodes lymphocytes were harvested after one day (day 14 + 1) (Fig. 3A). Treatment of CrPVIRES-ssRNA with Ova protein resulted in enhanced expression of the co-stimulatory molecules, CD40+, CD80+, and CD86+, on CD8a+DC1 cell population in spleen and inguinal lymph nodes (Fig. 3B, C). Additionally, an increased expression of the chemokine receptor, XCR1+, which is also an activation marker of CD8a+ DC1 cells, was observed in the spleen and inguinal lymph node cells (Fig. 3D). This suggested that CrPVIRES-ssRNA with Ova protein could act as an adjuvant therapy by activating CD8a+ cDC1 cells.
CrPVIRES-ssRNA formulated with Ova protein elicits production and activation of T cells
To evaluate CrPVIRES-ssRNA-mediated T-cell activation, splenocytes, and inguinal lymph nodes lymphocytes were isolated on day 7 after the second immunization (day 14 + 7) of mice with saline (Group 1; G1), Ova protein (Group 2; G2), Ova with alum (Group 3; G3), and CrPVIRES-ssRNA formulated with Ova (Group 4; G4) (Fig. 4A). The population of Ova-specific CD8+ T cells in the spleen significantly increased in the CrPVIRES-ssRNA-Ova-treated group (G4) (Fig. 4B, C). Furthermore, G4 showed statistically significant difference increased population of Ova-specific CD8 effector T cells and Ki-67+, a proliferation marker, compared to G2 in the drain inguinal lymph nodes (Fig. 4D). IFN-γ is an inflammatory cytokine with anti-tumor function that is related to innate and adaptive immune responses [44]. In the CrPVIRES-ssRNA-Ova-treated group, the frequency of IFN-γ-secreting cells after stimulation with CD4 and CD8 specific peptides was significantly higher than that of G2 and G3 (Fig. 4E). These data suggest that CrPVIRES-ssRNA treatment enhances dendritic cell uptake and activates CD8a+ cDC1, especially when administered alone or in combination with antigen, leading to activated antigen-specific CD8 T cells.
CrPVIRES-ssRNA reduces tumor burden in a murine melanoma model
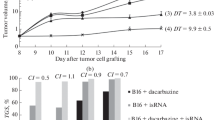
To test the in-vivo efficacy of CrPVIRES-ssRNA, mice were challenged with B16-Ova cells. Tumor-bearing mice (ten in each group) were inoculated intramuscularly with Ova protein, Ova protein-poly(I:C), and Ova protein-CrPVIRES-ssRNA. In the poly(I:C)- and CrPVIRES-ssRNA-treated groups, there was a decrease in cancer growth and weight compared to the control (Ova protein-treated) group (Fig. 5A-C). Mouse weight was not significantly different between groups (Fig. 5D). In addition, CrPVIRES-ssRNA and poly(I:C) treatment increased the survival rate. (Supplementary Fig. S3A). These results suggest that immune cells augmented by CrPVIRES-ssRNA may exhibit potential anti-tumor effects by protecting against melanoma growth (Fig. 6).

CrPVIRES-ssRNA reduces tumor burden in mouse melanoma. Mice (C57BL/6) were challenged with a subcutaneous injection of B16-Ova cells (5 × 10.5 cells) on day 0. Then, the mice were immunized intramuscularly with Ova protein (50 μg), poly(I:C) (20 μg) and CrPVIRES-ssRNA (20 μg) every 2 days until day 7. A Representative tumor images harvested on day 31. B Tumor size was measured every 2 days in the indicated groups. C Weight of tumors harvested on day 31. D Mice weights were measured every 2 days. Data were represented as mean ± SD. Statistical significance was indicated by *p < 0.05 and **p < 0.01

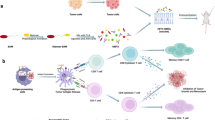
CrPVIRES-ssRNA activates immune signaling pathways associated with reduced mouse melanoma
Discussion
Cancer immunotherapy has been actively studied as a promising strategy in cancer treatment. However, there are many limitations in treating cancer such as immune tolerance and insufficient immune response. Adjuvant therapies that increase the immune response may play an important role in overcoming these limitations. For instance, cancer vaccines that introduce tumor antigens to APCs can enhance anti-tumor immune responses by attenuating immune resistance and generating sufficient immune responses in the cancer environment [45]. Dendritic cells, a type of APC activated by cancer vaccines, can initiate immune cell migration and activation [46], leading to the activation of antigen-specific CD4/CD8 T cells that can directly attack cancer.
Previous studies have reported that double-stranded RNA may be an important substance in cancer vaccines and may itself increase immune responses via TLR3 and RIG-Is [47]. Previous findings have confirmed that the CrPVIRES-ssRNA increased the expression of CXCR1, an inflammatory dendritic cell marker, around the injection site [35, 48]. In this study, we investigated the mechanism of the novel viral IRES-derived CrPVIRES-ssRNA in inducing immune responses [35]. CrPVIRES-ssRNA gradually increased the total cell population such as DCs, macrophages, and B cells in the drain inguinal lymph nodes. Moreover, it enhanced the activation of APCs (Fig. 1). In general, activated APCs have been reported to enhance antigen presentation to major histocompatibility (MHC) molecules, leading to adaptive and innate immunity [12, 49, 50]. Expression analysis of co-inflammatory signature proteins revealed that CrPVIRES-ssRNA increased antigen uptake by dendritic cells. Notably, CrPVIRES-ssRNA treatment significantly increased the co-stimulatory molecules of CD8+cDC1, such as CD40+, CD80+, CD86+, and XCR1+ (Figs. 2 and 3), suggesting that CrPVIRES-ssRNA is strongly associated with specific types of DCs. It is known that CD8+ cDC1 cells contribute to the effective cross-presentation of antigens [51,52,53] and the induction of Th1 responses [54, 55], as well as enhancing the production of chemokines associated with effector and memory CTLs [56, 57]. Functionally, expression of XCR1+ on CD8+ cDC1 leads to an increase in the proliferation and activation of antigen-specific CD8+ T cells and secretion of IFN-γ [58, 59]. These results indicated that CrPVIRES-ssRNA can induce a general immune response by stimulating APCs, especially cDC1 cells, to induce activation of effector T cells.
In addition, this study investigated whether CrPVIRES-ssRNA induces antigen-specific T-cell responses. CrPVIRES-ssRNA significantly increased the total population and proliferative capacity of antigen-specific effector CD8 T cells in drain inguinal lymph nodes (Fig. 4D), suggesting that CrPVIRES-ssRNA activates antigen-specific CD8 T cells by stimulating the maturation of cDC1 cells. It has been reported that cDC 1 cells induce the expansion of CD8 T cells primarily by presenting specific antigens expressed on MHC class I molecules [60]. The activation of CD8 T cells was involved in anti-tumor immune responses [61, 62] and anti-viral responses [63]. Interestingly, CrPVIRES-ssRNA increased the frequency of IFN-γ-secreting cells (Fig. 4E). In addition, although not statistically significant in the spleen of challenged mice, a slight increase the population of IFN-γ-secreting cells can be seen than in the negative control group (G1) (Supplementary Fig. S3B). It has been observed that antigen-specific CD4 and CD8 T cells inhibited the growth of melanoma cells, mainly by inducing IFN-γ cytokines that increase tumor-infiltrating lymphocytes and activating APCs to kill melanoma cells [64]. Indeed, CrPVIRES-ssRNA delayed the growth of cancer cells in a murine melanoma model (Fig. 5).
Conclusions
In conclusion, a novel cricket paralysis virus IRES-based single-stranded RNA may play an essential role as an adjuvant in inducing cDC1-CD8 T cell-mediated immune responses in cancer immunotherapy. Based on the immunological mechanism induced by the ssRNA adjuvant (Fig. 6), it can be applied to various cancer vaccines and adjuvant cancer treatment regimes.
Availability of data and materials
All data generated or analyzed during this study are included in this published article and its supplementary information files.
Abbreviations
- ssRNA:
-
Single-stranded RNA
- CrPV:
-
Cricket paralysis virus
- IRES:
-
Internal ribosome entry site
- cDC1:
-
Conventional dendritic cell 1
- PRRs:
-
Pattern recognition receptors
- TLRs:
-
Toll-like receptors
- RIG-I:
-
Retinoic acid-inducible gene I
- RIG-Is:
-
RIG-I-like receptors
- APCs:
-
Antigen-presenting cells
- cDCs:
-
Conventional dendritic cells
- pDCs:
-
Plasmacytoid dendritic cells
- UV:
-
Ultraviolet
- Ova-FITC:
-
Fluorescein isothiocyanate-conjugated Ova
- cDC1,cDC2:
-
Conventional DC1/2
References
Petrovsky N, Aguilar JC. Vaccine adjuvants: current state and future trends. Immunol Cell Biol. 2004;82(5):488–96.
Guerrero Manriquez GG, Tuero I. Adjuvants: friends in vaccine formulations against infectious diseases. Hum Vaccin Immunother. 2021;17(10):3539–50.
Pasquale AD, Preiss S, Silva FTD, Garçon N. Vaccine Adjuvants: from 1920 to 2015 and Beyond. Vaccines. 2015;3(2):320–43.
Pulendran B, Ahmed R. Immunological mechanisms of vaccination. Nat Immunol. 2011;12(6):509–17.
Desmet CJ, Ishii KJ. Nucleic acid sensing at the interface between innate and adaptive immunity in vaccination. Nat Rev Immunol. 2012;12(7):479–91.
Schijns VEJC. Immunological concepts of vaccine adjuvant activity: Commentary. Curr Opin Immunol. 2000;12(4):456–63.
Kubelkova K, Macela A. Innate Immune Recognition: An Issue More Complex Than Expected. Front Cell Infect Microbiol. 2019;9:241.
Gavin AL, Hoebe K, Duong B, Ota T, Martin C, Beutler B, Nemazee D. Adjuvant-enhanced antibody responses in the absence of toll-like receptor signaling. Science. 2006;314(5807):1936–8.
Apostólico JdS. Lunardelli VAS, Coirada FC, Boscardin SB, Rosa DS: Adjuvants: Classification, Modus Operandi, and Licensing. JImmunol Res. 2016;2016:1459394.
Franchi L, Park JH, Shaw MH, Marina-Garcia N, Chen G, Kim YG, Núñez G. Intracellular NOD-like receptors in innate immunity, infection and disease. Cell Microbiol. 2008;10(1):1–8.
Reed SG, Bertholet S, Coler RN, Friede M. New horizons in adjuvants for vaccine development. Trends Immunol. 2009;30(1):23–32.
Ho NI, Huis in 't Veld LGM, Raaijmakers TK, Adema GJ. Adjuvants enhancing cross-presentation by dendritic cells: the key to more effective vaccines? Front Immunol. 2018;9:2874.
Longhi MP, Trumpfheller C, Idoyaga J, Caskey M, Matos I, Kluger C, Salazar AM, Colonna M, Steinman RM. Dendritic cells require a systemic type I interferon response to mature and induce CD4+ Th1 immunity with poly IC as adjuvant. J Exp Med. 2009;206(7):1589–602.
Bauer DL, Kurtz JR, Grant SB, Immethun VE, McLachlan JB: Adjuvant combinations activate dendritic cells and contribute to antigen-specific CD4 T cell expansion. J Immunol 2020, 204(1 Supplement):166.115–166.115.
Sugai T, Mori M, Nakazawa M, Ichino M, Naruto T, Kobayashi N, Kobayashi Y, Minami M, Yokota S. A CpG-containing oligodeoxynucleotide as an efficient adjuvant counterbalancing the Th1/Th2 immune response in diphtheria–tetanus–pertussis vaccine. Vaccine. 2005;23(46):5450–6.
Stils HF Jr. Adjuvants and antibody production: dispelling the myths associated with freund’s complete and other adjuvants. ILAR J. 2005;46(3):280–93.
Comberlato A, Paloja K, Bastings MMC. Nucleic acids presenting polymer nanomaterials as vaccine adjuvants. J Mater Chem B. 2019;7(41):6321–46.
Hogenesch H. Mechanism of immunopotentiation and safety of aluminum adjuvants. Front Immunol. 2013;3:406–406.
Duthie MS, Windish HP, Fox CB, Reed SG. Use of defined TLR ligands as adjuvants within human vaccines. Immunol Rev. 2011;239(1):178–96.
Diebold SS, Kaisho T, Hemmi H, Akira S. Reis e Sousa C: Innate antiviral responses by means of TLR7-mediated recognition of single-stranded RNA. Science. 2004;303(5663):1529–31.
Alexopoulou L, Holt AC, Medzhitov R, Flavell RA. Recognition of double-stranded RNA and activation of NF-kappaB by Toll-like receptor 3. Nature. 2001;413(6857):732–8.
Martins KA, Bavari S, Salazar AM. Vaccine adjuvant uses of poly-IC and derivatives. Expert Rev Vaccines. 2015;14(3):447–59.
Stahl-Hennig C, Eisenblätter M, Jasny E, Rzehak T, Tenner-Racz K, Trumpfheller C, Salazar AM, Uberla K, Nieto K, Kleinschmidt J, et al. Synthetic double-stranded RNAs are adjuvants for the induction of T helper 1 and humoral immune responses to human papillomavirus in rhesus macaques. PLoS Pathog. 2009;5(4):e1000373.
Temizoz B, Kuroda E, Ishii KJ. Combination and inducible adjuvants targeting nucleic acid sensors. Curr Opin Pharmacol. 2018;41:104–13.
Kawai T, Akira S. The role of pattern-recognition receptors in innate immunity: update on Toll-like receptors. Nat Immunol. 2010;11(5):373–84.
Li Q, Yan Y, Liu J, Huang X, Zhang X, Kirschning C, Xu HC, Lang PA, Dittmer U, Zhang E, et al. Toll-Like Receptor 7 Activation Enhances CD8+ T Cell Effector Functions by Promoting Cellular Glycolysis. Front Immunol. 2019;10:2191–2191.
Komal A, Noreen M, El-Kott AF. TLR3 agonists: RGC100, ARNAX, and poly-IC: a comparative review. Immunol Res. 2021;69(4):312–22.
Oh JZ, Kurche JS, Burchill MA, Kedl RM. TLR7 enables cross-presentation by multiple dendritic cell subsets through a type I IFN-dependent pathway. Blood. 2011;118(11):3028–38.
Ziegler A, Soldner C, Lienenklaus S, Spanier J, Trittel S, Riese P, Kramps T, Weiss S, Heidenreich R, Jasny E, et al. A New RNA-based adjuvant enhances virus-specific vaccine responses by locally Triggering TLR- and RLH-Dependent Effects. J Immunol. 2017;198(4):1595–605.
Negishi H, Yanai H, Nakajima A, Koshiba R, Atarashi K, Matsuda A, Matsuki K, Miki S, Doi T, Aderem A et al: Cross-interference of RLR and TLR signaling pathways modulates antibacterial T cell responses. Nat Immunol 2012, 13(7):659–666.
Hochheiser K, Klein M, Gottschalk C, Hoss F, Scheu S, Coch C, Hartmann G, Kurts C. Cutting Edge: The RIG-I Ligand 3pRNA Potently Improves CTL Cross-Priming and Facilitates Antiviral Vaccination. J Immunol. 2016;196(6):2439–43.
Bourquin C, Schmidt L, Lanz AL, Storch B, Wurzenberger C, Anz D, Sandholzer N, Mocikat R, Berger M, Poeck H, et al. Immunostimulatory RNA oligonucleotides induce an effective antitumoral NK cell response through the TLR7. J Immunol. 2009;183(10):6078–86.
Jongbloed SL, Kassianos AJ, McDonald KJ, Clark GJ, Ju X, Angel CE, Chen C-JJ, Dunbar PR, Wadley RB, Jeet V et al: Human CD141+ (BDCA-3)+ dendritic cells (DCs) represent a unique myeloid DC subset that cross-presents necrotic cell antigens. The Journal of experimental medicine 2010, 207(6):1247–1260.
Heidenreich R, Jasny E, Kowalczyk A, Lutz J, Probst J, Baumhof P, Scheel B, Voss S, Kallen KJ, Fotin-Mleczek M. A novel RNA-based adjuvant combines strong immunostimulatory capacities with a favorable safety profile. Int J Cancer. 2015;137(2):372–84.
Kwak HW, Park H-J, Ko HL, Park H, Cha MH, Lee S-M, Kang KW, Kim R-H, Ryu SR, Kim H-J, et al. Cricket paralysis virus internal ribosome entry site-derived RNA promotes conventional vaccine efficacy by enhancing a balanced Th1/Th2 response. Vaccine. 2019;37(36):5191–202.
Kim YH, Bang YJ, Park HJ, Li Ko H, Park SI, Hwang KA, Kim H, Nam JH. Inactivated influenza vaccine formulated with single-stranded RNA-based adjuvant confers mucosal immunity and cross-protection against influenza virus infection. Vaccine. 2020;38(39):6141–52.
Lee SJ, Park H-J, Ko HL, Lee JE, Lee HJ, Kim H, Nam J-H. Evaluation of glycoprotein E subunit and live attenuated varicella-zoster virus vaccines formulated with a single-strand RNA-based adjuvant. Immun Inflamm Dis. 2020;8(2):216–27.
Rezaei T, Davoudian E, Khalili S, Amini M, Hejazi M, de la Guardia M, Mokhtarzadeh A. Strategies in DNA vaccine for melanoma cancer. Pigment Cell Melanoma Res. 2021;34(5):869–91.
Kuryk L, Bertinato L, Staniszewska M, Pancer K, Wieczorek M, Salmaso S, Caliceti P, Garofalo M: From Conventional Therapies to Immunotherapy: Melanoma Treatment in Review. Cancers (Basel) 2020, 12(10).
Lakshmi Narendra B, Eshvendar Reddy K, Shantikumar S, Ramakrishna S. Immune system: a double-edged sword in cancer. Inflamm Res. 2013;62(9):823–34.
Mitra D, Luo X, Morgan A, Wang J, Hoang MP, Lo J, Guerrero CR, Lennerz JK, Mihm MC, Wargo JA, et al. An ultraviolet-radiation-independent pathway to melanoma carcinogenesis in the red hair/fair skin background. Nature. 2012;491(7424):449–53.
Zhang W, Du J-Y, Jiang Z, Okimura T, Oda T, Yu Q, Jin J-O. Ascophyllan purified from Ascophyllum nodosum induces Th1 and Tc1 immune responses by promoting dendritic cell maturation. Mar Drugs. 2014;12(7):4148–64.
Honma K, Kohno Y, Saito K, Shimojo N, Horiuchi T, Hayashi H, Suzuki N, Hosoya T, Tsunoo H, Niimi H. Allergenic epitopes of ovalbumin (OVA) in patients with hen’s egg allergy: inhibition of basophil histamine release by haptenic ovalbumin peptide. Clin Exp Immunol. 1996;103(3):446–53.
Mendoza JL, Escalante NK, Jude KM, Sotolongo Bellon J, Su L, Horton TM, Tsutsumi N, Berardinelli SJ, Haltiwanger RS, Piehler J, et al. Structure of the IFNγ receptor complex guides design of biased agonists. Nature. 2019;567(7746):56–60.
Cuzzubbo S, Mangsbo S, Nagarajan D, Habra K, Pockley AG, McArdle SEB. Cancer Vaccines: Adjuvant Potency, Importance of Age, Lifestyle, and Treatments. Front Immunol. 2020;11:615240.
McKee AS, Munks MW, Marrack P. How Do Adjuvants Work? Important Considerations for New Generation Adjuvants. Immunity. 2007;27(5):687–90.
Jin B, Sun T, Yu XH, Liu CQ, Yang YX, Lu P, Fu SF, Qiu HB, Yeo AE. Immunomodulatory effects of dsRNA and its potential as vaccine adjuvant. J Biomed Biotechnol. 2010;2010:690438.
Łyszkiewicz M, Witzlau K, Pommerencke J, Krueger A. Chemokine receptor CX3CR1 promotes dendritic cell development under steady-state conditions. Eur J Immunol. 2011;41(5):1256–65.
Awate S, Babiuk LA, Mutwiri G. Mechanisms of action of adjuvants. Front Immunol. 2013;4:114–114.
Guéry JC, Ria F, Adorini L. Dendritic cells but not B cells present antigenic complexes to class II-restricted T cells after administration of protein in adjuvant. J Exp Med. 1996;183(3):751–7.
Pooley JL, Heath WR, Shortman K. Cutting edge: intravenous soluble antigen is presented to CD4 T cells by CD8- dendritic cells, but cross-presented to CD8 T cells by CD8+ dendritic cells. J Immunol. 2001;166(9):5327–30.
Dudziak D, Kamphorst AO, Heidkamp GF, Buchholz VR, Trumpfheller C, Yamazaki S, Cheong C, Liu K, Lee HW, Park CG, et al. Differential antigen processing by dendritic cell subsets in vivo. Science. 2007;315(5808):107–11.
Nierkens S, Tel J, Janssen E, Adema GJ. Antigen cross-presentation by dendritic cell subsets: one general or all sergeants? Trends Immunol. 2013;34(8):361–70.
Martínez-López M, Iborra S, Conde-Garrosa R, Sancho D. Batf3-dependent CD103+ dendritic cells are major producers of IL-12 that drive local Th1 immunity against Leishmania major infection in mice. Eur J Immunol. 2015;45(1):119–29.
Hor JL, Whitney PG, Zaid A, Brooks AG, Heath WR, Mueller SN. Spatiotemporally Distinct Interactions with Dendritic Cell Subsets Facilitates CD4+ and CD8+ T Cell Activation to Localized Viral Infection. Immunity. 2015;43(3):554–65.
Alexandre YO, Ghilas S, Sanchez C, Le Bon A, Crozat K, Dalod M. XCR1+ dendritic cells promote memory CD8+ T cell recall upon secondary infections with Listeria monocytogenes or certain viruses. J Exp Med. 2016;213(1):75–92.
Spranger S, Dai D, Horton B, Gajewski TF. Tumor-Residing Batf3 Dendritic Cells Are Required for Effector T Cell Trafficking and Adoptive T Cell Therapy. Cancer Cell. 2017;31(5):711-723.e714.
Dorner BG, Dorner MB, Zhou X, Opitz C, Mora A, Güttler S, Hutloff A, Mages HW, Ranke K, Schaefer M, et al. Selective expression of the chemokine receptor XCR1 on cross-presenting dendritic cells determines cooperation with CD8+ T cells. Immunity. 2009;31(5):823–33.
Crozat K, Guiton R, Contreras V, Feuillet V, Dutertre CA, Ventre E, Vu Manh TP, Baranek T, Storset AK, Marvel J, et al. The XC chemokine receptor 1 is a conserved selective marker of mammalian cells homologous to mouse CD8alpha+ dendritic cells. J Exp Med. 2010;207(6):1283–92.
Murphy TL, Murphy KM. Dendritic cells in cancer immunology. Cell Mol Immunol. 2022;19(1):3–13.
Raskov H, Orhan A, Christensen JP, Gogenur I. Cytotoxic CD8(+) T cells in cancer and cancer immunotherapy. Br J Cancer. 2021;124(2):359–67.
Kim CW, Kim KD, Lee HK. The role of dendritic cells in tumor microenvironments and their uses as therapeutic targets. BMB Rep. 2021;54(1):31–43.
Theisen D, Murphy K: The role of cDC1s in vivo: CD8 T cell priming through cross-presentation. F1000Res 2017, 6:98.
Karachaliou N, Gonzalez-Cao M, Crespo G, Drozdowskyj A, Aldeguer E, Gimenez-Capitan A, Teixido C, Molina-Vila MA, Viteri S, De Los Llanos Gil M et al: Interferon gamma, an important marker of response to immune checkpoint blockade in non-small cell lung cancer and melanoma patients. Ther Adv Med Oncol 2018, 10:1758834017749748.
Acknowledgements
Not applicable
Funding
J.-H.N. was supported by the Research Unit of Korea Center for Disease Control and Prevention grant (grant number 2020-ER5303-02), the National Research Foundation of Korea (NRF) grant (grant number NRF-2021M3E5E3080588), the Ministry of Food and Drug Safety grant (grant number 22213MFDS421) and partially supported by the Brain Korea 21 Plus Program, H.-J.P. is supported by the grant from the Ministry of Food and Drug Safety in 2022 (grant number 22203MFDS404).
Author information
Authors and Affiliations
Contributions
H.W. K. and J.-H. N. conceived and supervised the research and designed the experiments. H.W. K and Y.-J. B. confirmed the efficacy of adjuvant in ex vivo. H.W.K., J.-Y. K., Y.-S. L., and S.-H. B. performed the vaccinations and sacrificed in mice. H.W.K. and H.-J. P. flow cytometry analysis and interpreted the data. S.-H. H., H.-J. P., H. Y., and J.-H.N. assisted in the adjustments of directions and interpretation of the mechanistic aspects of the results. H.W.K., H. Y., and J.-H.N. contributed to writing the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
All animal studies were conducted according to the guidelines of and with the ethical approval of the Institutional Animal Care at the Catholic University of Korea (CUK-IACUC-2021–025-01). All methods were carried out in accordance with ARRIVE guidelines and regulations (https://arriveguidelines.org). All methods were reported in accordance with ARRIVE guidelines for the reporting of animal experiments.
Consent for publication
Not applicable.
Competing interests
The author(s) declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Addition al file 1: Supplementary Fig. S1.
CrPVIRES-ssRNA recruits immune cells into drain lymph node. C57BL/6 mice were intramuscularly injected with alum, CrPVIRES-ssRNA, and poly(I:C) for 48 h. Macrophage and T cells were analyzed using flow cytometry in drain inguinal lymph nodes. (A) Cell percentage of macrophages in the indicated groups (n = 3). (B, C) Cell percentage of CD4-positive T cells (B) and CD8-positive T cells (C) in drain inguinal lymph nodes (n = 3). Data were represented as mean ± SD. Statistical significance was indicated by *p < 0.05.
Additional file 2: Supplementary Fig. S2.
CrPVIRES-ssRNA activates co-stimulatory molecules, CD40, CD80, and CD86. (A) BMDCs were stimulated with CrPVIRES-ssRNA or poly(I:C) according to the table for 6 h. Cell activation was measured by flow cytometry using antibodies to CD40, CD80, and CD86. (B) The inguinal lymph nodes were isolated 1 day after immunization. Plots show percentages of co-stimulatory molecules CD40, CD80, and CD86 of macrophages in lymphocytes isolated from drain inguinal lymph nodes. Data were represented as mean ± SD. Statistical significance was indicated by *p < 0.05, **p < 0.01, and ***p < 0.005.
Additional file 3: Supplementary Fig. S3.
CrPVIRES-ssRNA is associated with melanoma antigen specific IFN- γ. After challenge with a subcutaneous injection of B16-Ova cells (5×105cells), the mice (C57BL/6) were immunized intramuscularly with Ova protein (50 μg), poly(I:C) (20 μg), and CrPVIRES-ssRNA (20 μg) every 2 days until day 7. (A) According to animal ethics standards, mice were sacrificed when the size of the cancer was greater than 1 cm3.The survival % was measured for mice in all groups over time to day 27 (n = 10). (B) Splenocytes were stimulated for 2 days with/without Ova-specific CD4 or CD8 T cell peptide. The IFN-γ-producing cells were measured using ELISpot in the indicated groups.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Kwak, H.W., Hong, SH., Park, HJ. et al. Adjuvant effect of IRES-based single-stranded RNA on melanoma immunotherapy. BMC Cancer 22, 1041 (2022). https://doi.org/10.1186/s12885-022-10140-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12885-022-10140-2