
Abstract
Malaria is a significant global health concern, with a majority of cases in Sub-Saharan African nations. Numerous antimalarial drugs have been developed to counter the rampant prevalence of Plasmodium falciparum malaria. Artemisinin-based Combination Therapy (ACT) has served as the primary treatment of uncomplicated malaria in Ghana since 2005. However, a growing concern has emerged due to the escalating reports of ACT resistance, particularly in Southeast Asia, and its encroachment into Africa. Specifically, mutations in the Kelch propeller domain on chromosome 13 (Pfk13) have been linked to ACT resistance. Yet, our understanding of mutation prevalence in Africa remains largely uncharted. In this study, we compared Pfk13 sequences obtained from 172 P. falciparum samples across three ecological and transmission zones in Ghana. We identified 27 non-synonymous mutations among these sequences, of which two of the mutations, C580Y (found in two samples from the central region) and Y493H (found in one sample from the north), had previously been validated for their association with artemisinin resistance, a phenomenon widespread in Southeast Asia. The Pfk13 gene diversity was most pronounced in the northern savannah than the central forest and south coastal regions, where transmission rates are lower. The observed mutations were not significantly associated with geographical regions, suggesting a frequent spread of mutations across the country. The ongoing global surveillance of artemisinin resistance remains pivotal, and our findings provides insights into the potential spread of resistant parasites in West Africa. Furthermore, the identification of novel codon mutations in this study raises their potential association to ACT resistance, warranting further investigation through in vitro assays to ascertain their functional significance.
Similar content being viewed by others


Introduction
Malaria is a significant global public health concern, causing around 247 million cases and 619,000 deaths worldwide [1]. More than 90% of global malaria cases are from countries in Sub-Saharan Africa where malaria is endemic [2]. Malaria transmission is determined by various factors such as environment, climate, abundance of vector mosquitoes, human migration and immune responses, and the level of malaria control measures. For example, in the northern savannah and central forest regions of Ghana, irrigation and gold mining activities create breeding grounds and/or nesting sites for Anopheles mosquitoes, contributing to high transmission rates. By contrast, in the southern coastal region with more urban setting and fewer mosquito habitats, transmission rate is relatively low [3, 4]. In addition, socioeconomic factors such as inadequate healthcare access in the rural remote areas also increase malaria incidence [4, 5]. Prior studies suggested that transmission intensity may not be necessarily associated with the prevalence of antimalarial drug resistance [6,7,8,9]. Instead, the level of host immunity [7], population size [8], and/or the number of parasite clones co-infecting the same host [9] are factors more likely affecting drug resistance.
The antimalarial drug treatment regime has undergone frequent changes over the past decades due to the rise of drug resistance. During the 1950s, Chloroquine (CQ) was the primary antimalarial drug against P. falciparum [10]. In 2012, WHO recommended chemoprevention with a monthly course of amodiaquine and sulfadoxine-pyrethamine (SP) to combat malaria during high transmission seasons [11]. Resistance of P. falciparum to CQ and SP became widespread in the 1950s and 1960s, with the first reported CQ resistance in Ghana in 1987 [12]. In response to the increased malaria related morbidity and mortality, artemisinin-based combination therapy (ACT) was recommended as the first line treatment [13]. Artemisinin has been shown to be effective against CQ sensitive and resistant P. falciparum malaria [14]. However, artemisinin has a short half-life and is ineffective against latent forms of primary and hepatic malaria [14]. Because artemisinin monotherapies (ART) alone did not yield unsatisfactory results, artemisinin has been combined with other antimalarial drugs that target various development stages of the parasite, particularly the early stages [15]. Selective pressure has driven to resistance against artemisinin in South America and Southeast Asia in the late 1990’s [16], and is especially prevalent in high transmission areas of Thailand and Cambodia [17]. Studies over the last decade have shown that ART resistance in P. falciparum is attributed to selection for mutations in genes that govern the physiological responses of the parasites and determine parasite clearance. Specifically, mutations within the highly conserved kelch propeller domain on chromosome 13 (pfk13) inactivate proteins needed for endocytosis of hemoglobin. Given ART is activated by the products from hemoglobin degradation, the lack of these products prevents ART activation and results in parasitic resistance [18]. Pfk13 mutations were shown to disrupt various aspects of the parasite’s developmental program by affecting cell-cycle timing, protein response, degradation, trafficking, and mitochondrial function. Mitochondrial processes can contribute to pfk13 mutants defense against artemisinin via temporary quiescence and rapid growth recovery, a process reversed by the mitochondrial electron transport chain inhibitor atovaquone [19]. Several codon mutations in pfk13 gene associated with artemisinin resistance such as C580Y, Y493H, R539T, I543T, and N458Y have been identified in the field isolates from Southeast Asia and parts of Africa (Supplementary Table 1 [20, 21].
In Ghana, artemisinin combination therapy have been implemented since 2005 [3]. Though malaria transmission occurs all year round, different regions vary in transmission intensity due to climatic and landscape variations, population density, and urbanization [22]. Our previous study has shown that mutations in Pfcrt, Pfmdr1, Pfdhfr, and Pfdhps associated with CQ and SP resistance were not significantly different across the three ecological zones of Ghana, and that the majority of the isolates had the wildtype codons, with no sign of selection pressure [5]. Several non-synonymous as well as few synonymous pfk13 mutations have been reported in the central forest region [23,24,25]. However, it remains unclear if the mutation frequency of Pfk13 differs among ecological zones. This study determined and compared Pfk13 non-synonymous and synonymous mutations among the three ecological regions of Ghana and examined the phylogenetic relatedness between the mutant and wildtype isolates, with the goals to understand the evolution and improve the surveillance of artemisinin resistance in West Africa.
Materials and methods
Ethics statement
Scientific and ethical approval was given by the Institutional Scientific and Ethical Review boards of the Noguchi Memorial Institute of Medical Research, University of Ghana, Ghana and the University of North Carolina at Charlotte, USA (IRB00001276). Written informed consent/assent for study participation was obtained from all consenting parents/guardians (for minors under the age of 18), and everyone who was willing to participate in the study. All methods were reviewed and approved by the institutional review board (IRB) and performed in accordance with the relevant guidelines and regulations stated in the IRB protocols.
Study sites and sample collection
Samples were collected from five sites including Pagaza (PZ) in Tamale Municipality and Kpalsogou (KG) in Kumbungu district in the northern savannah region; Duase (KD) in Konongo in the central forest region; and Ada (AD) and Dodowa (DO) in the southern coastal region of Ghana during June–July of 2018 with a malaria prevalence of 21.4%, 24% and 14% respectively [5]. In the northern savannah zone, malaria transmission is highly seasonal and intense during/after the rainy season (June–October), whereas in the south coastal (low elevation) and central forest areas, malaria transmission is perennial [26] (Fig. 1). Finger-prick blood samples were collected from 172 asymptomatic children, aged between 3 and 13 years, who were not experiencing fever, and who attended primary schools at the designated research locations, were selected for participation. All children within this specified age range, irrespective of gender and socioeconomic background, were qualified for inclusion. However, only those whose parent or guardian granted written parental consent and obtained the child’s agreement were enrolled in the study. Thus, these individuals did not receive any antimalarial treatment prior to blood sample collection. Thick and thin blood smears were prepared for microscopic examination. Blood samples (30–50 μL) were blotted on Whatman 3MM filter papers. The filter papers were air-dried and stored in sealed plastic bags with silica gel absorbent at room temperature until DNA extraction.

Map of Ghana showing topography and sampling. In the northern area, there were a total of N = 72 samples, with 64% (46) representing the wild type (WT) and 36% (25) corresponding to mutants (MT). Among the mutants, 68% were classified as synonymous, while 32% were categorized as non-synonymous. In the central region, there were N = 59 samples, of which 75% (44) were WT and 25% (15) were MT. Among the mutations observed, 75% were non-synonymous, whereas 25% were synonymous. Lastly, in the southern region, there were N = 41 samples, with 49% (20) being WT and 51% (21) being MT. Regarding the mutations in this area, 67% were synonymous, and 33% were non-synonymous. Samples with no mutations are represented in blue, samples with synonymous mutations in Grey and non-synonymous mutations in orange. The difference in size of the pie charts is proportional to the sample size
Parasitic DNA was extracted from the dried blood spots using a modified version of the Chelex®-saponin method [27]. Each filter paper punch was incubated overnight at 4 °C in 1 mL of 0.5% saponin in phosphate buffered saline (PBS). The punches were washed for 30 minutes in PBS at 4 °C, transferred into new tubes containing 300 μL of 20% Chelex® and vortexed for 30 seconds. Then the tubes were heated at 95 °C for 12 minutes to elute the DNA, vortexed, and centrifuged at 10,000×g for 8 minutes. The supernatants (± 200 μL) were transferred into new tubes. The DNA extract was kept at 4 °C for use within a few hours or at − 20 °C for long time storage.
The SYBR Green quantitative real-time PCR (qPCR) assay of the 18S rRNA was conducted to screen for P. falciparum using the published protocols [28]. For the P. falciparum-confirmed samples, a 849-bp amplicon of the Pfk13 gene was amplified using the published primers (Forward: GTAAAGTGAAGCCTTGTTG-3′; Reverse: 5′-TTCATTTGTATCTGGTGAAAAG − 3′) spanning nucleotide positions 1139–1979 [29]. PCR was conducted in a 20ul reaction mixture containing 2ul of genomic DNA, 10ul of 2xDreamTaq Green PCR Master Mix (Thermo Fisher) and 0.5uM primer. Reaction was performed with an initial denaturation at 94 °C for 2 min, followed by 30 cycles at 94 °C for 30 sec, 55 °C for 30 sec, and 72 °C for 60 sec, with a final 5 min extension at 72 °C. PCR products were purified prior to Sanger sequencing.
Pfk13 sequence analyses
We assessed the quality of Sanger sequencing chromatograms, focusing on well-resolved peaks and minimal noise using Chromas (version 2.6.6) [30] . To validate the detected mutations, we employed bioinformatics software, specifically using Clustal_W (Clustal W 2.1) [31] and BioEdit (version 7.2.5) [32], for precise sequence data analysis. This involves aligning the obtained sequences with the PF3D7 reference sequence (PF3D7_1343700) to identify the k13 mutation accurately. Confirmation is achieved through replicate sequencing reactions, and the results were rigorously evaluated and only samples that exhibited consistency in the identification of mutations were included, providing confidence in the detected genetic changes. We specifically focused on previously identified Pfk13 mutations that have been validated for artemisinin resistance in vitro [23], in addition to other synonymous and non-synonymous mutations (Supplementary Table 2). To determine and compare sequence polymorphisms among regions, haplotype diversity (Hd; the probability that two randomly sampled alleles are different), nucleotide diversity (Pi; the average number of nucleotide differences per site in pairwise comparisons), and Tajima’s D value were estimated using DNASP [33]. The Tajima’s D and Fu’s FS tests were conducted to distinguish between evolving neutrally and one evolving under a non-random process, including directional or balancing selection. To infer the genetic relatedness among the Pfk13 sequences from different transmission zones, we employed the maximum likelihood method using the Randomized Accelerated Maximum Likelihood (RAxML) for phylogenetic inferences with 500 iterations of bootstrapping for assessing confidence [34]. The resulted tree was visualized in FigTree v1.4.2 [35]. The sequence data generated in this study are available in the GenBank repository (accession number OQ102653-OQ102768).
Results
Pfk13 mutations
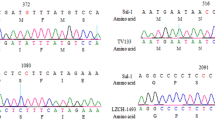
A total of 172 Pfk13 sequences (north = 72; central = 59; south = 41) were analysed. In the north (PZ and KG), 11 out of the 72 samples (15%) were detected with non-synonymous mutations at 11 codon positions including I418M, S423G, R471C, P475L, S477F, Y493H, V494P, N499Y, Y500F, D501N (Fig. 2). Each of these mutations was found in a single sample except D501N found in two samples. Among all codons found in the north, only Y493H has been validated in vitro that confers to ART resistance [36] (Table 1).

A Chart of the number of samples with non-synonymous mutations. In the northern region, non-synonymous mutations were identified in 15% of the samples (11 out of 72 samples) at 11 distinct codon positions. Each of these mutations was present in a single sample, accounting for 1.4% of the northern samples, except for D501N, which was observed in two samples, constituting 2.8% of the samples. In the central region, non-synonymous mutations were detected in 17% of the samples (10 out of 59 samples) at six codon positions. Mutations G449A and C580Y were each found in two samples, making up 3.4% of the samples in the central region. Mutation I543L was identified in three samples, accounting for 5% of the samples. The remaining mutations were each present in a single sample, representing 1.7% of the samples. In the southern region, non-synonymous mutations were discovered in 17% of the samples (7 out of 41 samples) at six codon positions. Mutation D501G was found in 2.4% of the samples. Furthermore, P475L was identified in two samples, constituting 5% of the samples in the south. B Structure of the kelch13 propeller domain, showing the position of mutations found in this study
In the central region (KD), 10 out of the 59 samples (17%) were detected with non-synonymous mutations at six codon positions including G449A, N458K, A481T, I543L, N548I, and C580Y. Mutations G449A and C580Y were each found in two different samples; and mutation I543L was detected in three (Fig. 2). C580Y is the most prevalent non-synonymous mutation in Southeast Asia that mediates ART resistance both in vitro and in vivo [20, 36]. Other mutations including A481T, N458K, N548I were found only in a single sample, respectively. In the south (AD and DO), 7 of the 41 samples (17%) were detected with non-synonymous mutations at six codon positions including N408D, M460T, M472T, Y482F, P475L, and D501G (Fig. 2). None of these mutations has been shown to be linked to ART resistance based on prior studies (Table 1). Each of these mutations was found in a single sample, except for P475L that was also detected in a sample from the north.
Apart from non-synonymous mutations, several synonymous mutations were also found in the Pfk13 gene among samples. In the north, there were 46 non-wildtype samples, of which 68% of the mutations were synonymous and 32% were non-synonymous. Similar proportion was observed in the south where 67% of the mutations were synonymous and 33% were non-synonymous among 21 non-wildtype samples (Fig. 1). By contrast, in the central region, 75% of the mutations were non-synonymous and 25% were synonymous among the 15 non-wildtype samples (Fig. 1).
All mutations previously reported in Ghana are compiled in Supplementary Table 3.
Phylogenetic and diversity analyses
There was no clear distinction between samples from the north, central, or south as they were found in all well-supported clades (Fig. 3). A total of 28 different pfk13 haplotypes were detected among the samples. Samples containing the 580Y and 493H mutations showed only one amino acid change (Supplementary Table 4). Both haplotype and nucleotide diversity were higher in the north (Hd = 0.8; Pi = 0.003) and central regions (Hd = 0.9; Pi = 0.004) than the southern region (Hd = 0.2; Pi = 0.0012; Table 2). Negative Tajima’s D values were observed in the three regions, suggesting that the Pfk13 gene is under selection pressure. The results of Fu’s FS test statistics were negative for all regions, indicating that the presence of a higher number of rare/unique Pfk13 haplotypes than what would be expected under neutrality (p-value = 0.005; Table 2).

Phylogenetic tree of the 172 sequences from north central and south of Ghana. There was no clear distinction between samples from the north, central, or south as they were found in all clades. Samples containing the C580Y and Y493H both validated ART resistance mutation are highlighted. Asterisks denote clades with bootstrap values over 95%
Discussion
Considering the trends of growing ART resistance in Southeast Asia and evidence of ART resistance in Sub-Saharan Africa [37], monitoring the spread of P. falciparum malaria and identification of Pfk13 mutations is crucial to controlling the disease. This study highlighted a relatively high occurrence of Pfk13 mutations in the northern and central regions of Ghana where malaria is most prevalent, compared to the southern region where transmission is low. The lack of statistical significance could likely be attributed to limited sample size. There are multiple biomarkers of ART resistance in Southeast Asia and Africa. In Southeast Asia, isolates with mutations C580Y, Y493H, R539T, I543T, and N458Y were showed with delayed clearance [20]. Of the 27 non-synonymous mutations identified in the Ghanaian samples, C580Y and Y493H has been previously validated for ART resistance and are widespread across Southeast Asia. A recent study, with a significantly larger sample size, identified 78 novel mutations that were exclusively observed in Ghana [38]. Out of these mutations, our study found only one, N458, in common. Other studies conducted between 2007 and 2016, indetified persisting mutations such as N599Y, K607E, and V637G, highlighting the ongoing need for vigilance in monitoring and responding to drug resistance [23]. However, none of these mutations have been validated for ART resistance.
The higher prevalence of pfk13 mutations in the north of Ghana could be attributed to a combination of factors, including the region’s poverty rate, limited access to healthcare, and a predominantly young population composed of children under 14. These factors can contribute to high malaria transmission, leading to increased genetic recombination, mutations, and spread of ART-resistant strains. The presence of shared mutations across different ecological zones suggests the role of human migrations and parasite gene flow across the country, emphasizing the intricate interplay among transmission dynamics, human movement, and parasite evolution in the context of artemisinin resistance [39]. In the north, malaria transmission is seasonal and significantly higher during and after the rainy season. Consistent to other studies, parasites from the southern region have low prevalence of Pfk13 mutations [26]. Despite the humid and warmer weather near the coast climate that is optimal for mosquito development, the southern region is densely populated, highly urbanized, higher socio-economic status, and greater access to healthcare services [40]. These factors likely contribute to low incidence of malaria, as people living in this area are more likely to be diagnosed and treated promptly for malaria.
The central forest region serves as a transitional zone between the north savannah and urban south; and reflects how the parasitic prevalence and variability may change as urbanization occurs. The central region of Ghana has the country’s major mineral reserves. Large-scale mining activity leads to landscape damage and pockets of stagnant water favor mosquito development [41]. In this region, we found relatively low frequency of Pfk13 mutations. One of the mutations G449A was previously identified as a candidate mutation for ART surveillance in Southeast Asia [20]. The type of amino acid substitution is also important when inferring the impact on resistance. For example, while I543L has not been yet confirmed as a mutation resulting in ART resistance, I543T was validated as a key marker for ART resistance [20]. Further analysis by CCF53_62 matrices that determines the effect of amino acid changes to protein function could offer deeper insights into the significance of the type of substitutions [42]. Other codon mutations associated with delayed ART clearance among African P. falciparum parasites include Q613H reported in Senegal [43], P574L in Rwanda [44] and Henan Province of China [45]), A675V in Rwanda [44] and Uganda [46], P553L in Kenya [47], and K189T in Senegal [43] and Uganda [48]. Previous study has shown that the mutation 580Y was observed in Chinese migrant workers returning from Ghana [49]. A study conducted between 2007 and 2016 in Ghana have reported a high number of mutations of which 77% was nonsynonymous [23], though their functional significance still remains unclear and warrants further investigations. Though not validated in function, A578S is the most commonly reported non-synonymous mutation in different parts of Africa including Ghana [50, 51] Equatorial Guinea [52], Mali [53], Kenya [54], Togo [55], but this mutation was not found in this study.
Based on neutrality tests, the Pfk13 gene was detected with significance selection pressure, particularly in the central region where there is a greater ratio of nonsynonymous to synonymous substitutions. It is plausible that easy access to antimalarial drugs in this region accelerates the development of selection pressure to ART resistance. With more than 10,000 licensed chemical seller (LCS) shops in Ghana, these shops are often the first point of care for febrile illnesses such as malaria, particularly in hard-to-reach areas [56]. Due to the increased accessibility of antimalarials, patients tend to opt for self-treatment, which can result in inadequate and inappropriate usage of non-prescribed drugs [56, 57]. As there are more LCS shops relative to health facilities in rural areas, artemisinin combination therapy could become more available in the northern savannah region and increase the Pfk13 mutation frequency. Many mutant Pfk13 codons in Ghana were found to be zone specific, implying that intra-country gene flow could be limited in spreading parasites through human migration [23]. While Sanger sequencing remains a valuable tool, its sensitivity constraints often hinder the accurate detection of minor mutants. Amplicon deep sequencing holds promises for unveiling a more comprehensive and nuanced mutational landscape, particularly in complex scenarios like polyclonal and mixed infections. Another limitation of this study is the lack of clinical efficacy data. Therefore, novel candidate mutations should be examined for both in vivo and in vitro resistance to artemisinin to clarify the significance of the mutations. While the Pfk13 gene is a valuable marker for understanding artemisinin resistance and genetic diversity, it might not reveal a full picture of the resistance mechanisms. Whole-genome sequencing and analysis of additional genetic markers may offer more comprehensive insights into the overall genetic diversity, evolution, and potential resistance mechanisms of P. falciparum.
Conclusion
The global surveillance of artemisinin resistance is and will continue to be crucial to understanding how antimalarial drug resistance evolves and spreads. This study highlighted a relatively high occurrence of Pfk13 mutations in the northern and central regions of Ghana where malaria is most prevalent, compared to the southern region where transmission is low. Our sequence analyses revealed 27 non-synonymous mutations and most of them have not been validated for ART resistance nor previously reported elsewhere in Africa. Our findings emphasize the need for further clinical and/or in vitro testing of the functional significance of novel Pfk13 codon mutations. Future studies should investigate the association of transmission intensity with pfk13 mutation prevalence at a broader geographical scale. Understanding how factors such as intra- and inter-country migration patterns and urbanization affect parasitic diversity and antimalarial resistance will have great implications on controlling malaria spread in Africa.
Availability of data and materials
The datasets generated and/or analyzed during the current study are available in the GenBank repository, [GenBank accession number BankIt2656762: OQ102653 - OQ102768.]
References
Monroe A, et al. Reflections on the 2021 world malaria report and the future of malaria control. Malar J. 2022;21(1):154.
Noor AM, Kinyoki DK, Mundia CW, et al. The changing risk of Plasmodium falciparum malaria infection in Africa: 2000-10: a spatial and temporal analysis of transmission intensity. Lancet. 2014;383(9930):1739-47.
Duah NO, et al. Mutations in plasmodium falciparum chloroquine resistance transporter and multidrug resistance genes, and treatment outcomes in Ghanaian children with uncomplicated malaria. J Trop Pediatr. 2007;53(1):27–31.
Dixon J, Tenkorang EY, Luginaah I. Ghana's National Health Insurance Scheme: A national level investigation of members' perceptions of service provision. BMC Int Health Hum Rights. 2013;13:35.
Dieng CC, et al. Contrasting asymptomatic and drug resistance gene prevalence of plasmodium falciparum in Ghana: Implications on seasonal malaria chemoprevention. Genes. 2019;10(7)
Artzy-Randrup Y, Alonso D, Pascual M. Transmission intensity and drug resistance in malaria population dynamics: Implications for climate change. PLoS One. 2010;5(10):e13588.
Klein EY, et al. Clinically immune hosts as a refuge for drug-sensitive malaria parasites. Malar J. 2008;7(1):1–9.
Pongtavornpinyo W, et al. Spread of anti-malarial drug resistance: Mathematical model with implications for ACT drug policies. Malar J. 2008;7(1):1–12.
Hastings I, d’Alessandro U. Modelling a predictable disaster:: the rise and spread of drug-resistant malaria. Parasitol Today. 2000;16(8):340–7.
D'Alessandro U, Buttiëns H. History and importance of antimalarial drug resistance. Tropical Med Int Health. 2001;6(11):845–8.
Organization, W.H. WHO policy recommendation: seasonal malaria chemoprevention (SMC) for plasmodium falciparum malaria control in highly seasonal transmission areas of the Sahel sub-region in Africa. World Health Organization; 2012.
Neequaye J, et al. Falciparum malaria not sensitive to chloroquine emerges in Accra in 1987. 1988.
Organization, W.H. Monitoring antimalarial drug resistance: report of a WHO consultation, Geneva, Switzerland, 3-5 December 2001. World Health Organization; 2002.
Tilley L, et al. Artemisinin action and resistance in plasmodium falciparum. Trends Parasitol. 2016;32(9):682–96.
Álvarez G, et al. Dynamics of plasmodium falciparum parasitemia regarding combined treatment regimens for acute uncomplicated malaria, Antioquia, Colombia. Am J Trop Med Hyg. 2010;83(1):90.
Organization, W.H. The world health report: 1999: making a difference. World Health Organization; 1999.
Ashley EA, et al. Spread of artemisinin resistance in plasmodium falciparum malaria. N Engl J Med. 2014;371(5):411–23.
Birnbaum J, et al. A Kelch13-defined endocytosis pathway mediates artemisinin resistance in malaria parasites. Science. 2020;367(6473):51–9.
Mok S, et al. Artemisinin-resistant K13 mutations rewire plasmodium falciparum's intra-erythrocytic metabolic program to enhance survival. Nat Commun. 2021;12(1):530.
Ariey F, et al. A molecular marker of artemisinin-resistant plasmodium falciparum malaria. Nature. 2014;505(7481):50–5.
Balikagala B, et al. Evidence of Artemisinin-resistant malaria in Africa. N Engl J Med. 2021;385(13):1163–71.
Frank C, et al. Spatial heterogeneity of malaria in Ghana: A cross-sectional study on the association between urbanicity and the acquisition of immunity. Malar J. 2016;15:84.
Matrevi SA, et al. Plasmodium falciparum kelch propeller polymorphisms in clinical isolates from Ghana from 2007 to 2016. Antimicrob Agents Chemother. 2019;63(11):e00802–19.
Mensah BA, et al. Antimalarial drug resistance profiling of plasmodium falciparum infections in Ghana using molecular inversion probes and next-generation sequencing. Antimicrob Agents Chemother. 2020;64(4)
Tornyigah B, et al. Effect of drug pressure on promoting the emergence of antimalarial-resistant parasites among pregnant women in Ghana. Antimicrob Agents Chemother. 2020;64(6)
Ayanful-Torgby R, et al. Seasonal variations in plasmodium falciparum parasite prevalence assessed by varying diagnostic tests in asymptomatic children in southern Ghana. PLoS One. 2018;13(6):e0199172.
Miguel RB, et al. Evaluation of three different DNA extraction methods from blood samples collected in dried filter paper in plasmodium subpatent infections from the Amazon region in Brazil. Rev Inst Med Trop Sao Paulo. 2013;55(3)
Lo E, et al. Low parasitemia in submicroscopic infections significantly impacts malaria diagnostic sensitivity in the highlands of Western Kenya. PLoS One. 2015;10(3):e0121763.
Mohon AN, et al. Mutations in plasmodium falciparum K13 propeller gene from Bangladesh (2009–2013). Malar J. 2014;13(1):1–6.
Treves DS. Review of three DNA analysis applications for use in the microbiology or genetics classroom. J Microbiol Biol Educ. 2010;11(2):186–7.
Thompson JD, et al. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997;25(24):4876–82.
Hall TA. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. In: Nucleic acids symposium series. London: Information Retrieval Ltd.; 1999. p. c1979–2000.
Librado P, Rozas J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics. 2009;25(11):1451–2.
Stamatakis A. Using RAxML to infer phylogenies. Curr Protoc Bioinformatics. 2015;51:6.14.1–6.14.14.
Rambaut A. FigTree v1. 4.2, a graphical viewer of phylogenetic trees. Available from left angle bracket, http://tree.bio.ed.ac.uk/software/figtree/rightanglebracket. 2014.
Straimer J, et al. K13-propeller mutations confer artemisinin resistance in plasmodium falciparum clinical isolates. Science. 2015;347(6220):428–31.
Ocan M, et al. K13-propeller gene polymorphisms in plasmodium falciparum parasite population in malaria affected countries: A systematic review of prevalence and risk factors. Malar J. 2019;18(1):1–17.
Matrevi SA, et al. Novel pfk13 polymorphisms in plasmodium falciparum population in Ghana. Sci Rep. 2022;12(1):7797.
Kafai NM, Odom John AR. Malaria in children. Infect Dis Clin N Am. 2018;32(1):189–200.
Stow DA, et al. Inter-regional pattern of urbanization in southern Ghana in the first decade of the new millennium. Appl Geogr. 2016;71:32–43.
Appiah JO, Ohene J, Afram A. Effects of mining activities on access to potable water: Households’ perception and practices at Konongo and Odumasi communities in Ghana. Civil Environ Res. 2013;12:2224–5790.
Brick K, Pizzi E. A novel series of compositionally biased substitution matrices for comparing plasmodium proteins. BMC Bioinform. 2008;9(1):1–15.
Boussaroque A, et al. Emergence of mutations in the K13 propeller gene of plasmodium falciparum isolates from Dakar, Senegal, in 2013-2014. Antimicrob Agents Chemother. 2016;60(1):624–7.
Tacoli C, et al. Artemisinin resistance–associated K13 polymorphisms of plasmodium falciparum in southern Rwanda, 2010–2015. Am J Trop Med Hyg. 2016;95(5):1090.
Yang C, et al. Polymorphisms of plasmodium falciparum k13-propeller gene among migrant workers returning to Henan Province, China from Africa. BMC Infect Dis. 2017;17(1):1–8.
Ikeda M, et al. Artemisinin-resistant plasmodium falciparum with high survival rates, Uganda, 2014–2016. Emerg Infect Dis. 2018;24(4):718.
Taylor SM, et al. Absence of putative artemisinin resistance mutations among plasmodium falciparum in sub-Saharan Africa: A molecular epidemiologic study. J Infect Dis. 2015;211(5):680–8.
Conrad MD, et al. Polymorphisms in K13 and falcipain-2 associated with artemisinin resistance are not prevalent in plasmodium falciparum isolated from Ugandan children. PLoS One. 2014;9(8):e105690.
Feng J, et al. Evaluation of antimalarial resistance marker polymorphism in returned migrant workers in China. Antimicrob Agents Chemother. 2015;59(1):326–30.
Project, M.P.f.C. Genomic epidemiology of artemisinin resistant malaria. elife. 2016;5:e08714.
Kamau E, et al. K13-propeller polymorphisms in plasmodium falciparum parasites from sub-Saharan Africa. J Infect Dis. 2015;211(8):1352–5.
Li J, et al. Limited artemisinin resistance-associated polymorphisms in plasmodium falciparum K13-propeller and PfATPase6 gene isolated from Bioko Island, Equatorial Guinea. Int J Parasitol Drugs Drug Resist. 2016;6(1):54–9.
Ouattara A, et al. Polymorphisms in the K13-propeller gene in artemisinin-susceptible plasmodium falciparum parasites from Bougoula-Hameau and Bandiagara, Mali. Am J Trop Med Hyg. 2015;92(6):1202.
Muwanguzi J, et al. Lack of K13 mutations in plasmodium falciparum persisting after artemisinin combination therapy treatment of Kenyan children. Malar J. 2016;15(1):1–6.
Dorkenoo AM, et al. Therapeutic efficacy trial of artemisinin-based combination therapy for the treatment of uncomplicated malaria and investigation of mutations in k13 propeller domain in Togo, 2012–2013. Malar J. 2016;15(1):1–9.
Adjei AA, et al. Insights into the affordable medicines Facility-malaria in Ghana: The role of caregivers and licensed chemical sellers in four regions. Malar J. 2016;15(1):1–11.
Buabeng KO, et al. Self-reported use of anti-malarial drugs and health facility management of malaria in Ghana. Malar J. 2007;6(1):1–6.
Acknowledgments
We thank the field team from The University of Ghana for their technical assistance in sample collection; the communities and hospitals, for their support and willingness to participate in this research; and undergraduates at The University of North Carolina at Charlotte who involve in sample processing.
Funding
This research was funded by Faculty Research Grant at The University of North Carolina at Charlotte.
Author information
Authors and Affiliations
Contributions
Conceptualization, CCD, LD, VM, LC and EL; resources, LA, YA, LC and EL; experiment and data analyses, CCD, LD, VM and EL; writing—original draft preparation, CCD and EL; writing—review and editing, CCD, LD, VM and EL, and EL; funding acquisition, EL. All authors have read and agreed to the published version of the manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Scientific and ethical clearance was given by the institutional scientific and ethical review boards of The University of Ghana and University of North Carolina at Charlotte (USA). Written informed consent/assent was obtained from all consenting heads of households, parents/guardians (for minors under 18 years old), and individuals who participated in this study. All methods were carried out in accordance with relevant guidelines and regulations.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Dieng, C.C., Morrison, V., Donu, D. et al. Distribution of Plasmodium falciparum K13 gene polymorphisms across transmission settings in Ghana. BMC Infect Dis 23, 801 (2023). https://doi.org/10.1186/s12879-023-08812-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12879-023-08812-w