
Abstract
Background
Studies have shown that the human T-lymphotropic virus 2 (HTLV-2) is endemic in several indigenous populations of the Brazilian Amazon and molecular analyses have shown the exclusive presence of HTLV-2 subtype 2c among the indigenous groups of this geographical region.
Methods
The present study characterizes the prevalence of HTLV-2 infection in three new villages of the Xikrin tribe, in the Kayapo group, according to their distribution by sex and age. The study included 263 samples from individuals from the Kateté, Djujeko and Oodjã villages. Plasma samples were tested for the presence of anti-HTLV-1/2 antibodies using enzyme-linked immunosorbent assays (ELISA). Seropositive samples were confirmed using real-time PCR, nested PCR and sequencing.
Results
The serological and molecular results confirmed the sole presence of HTLV-2 in 77 (29%) samples, with a prevalence of 38% among women and 18% among men. In these communities, it was found that the prevalence of HTLV-2 infection increased with age. Nucleotide sequences (642 bp, 5’LTR) from eight samples were subjected to phylogenetic analysis by the neighbor-joining method to determine the viral subtype, which confirmed the presence of HTLV-2c.
Conclusions
The results of the present study establish the presence of HTLV-2 infection in three new villages of the Xikrin tribe and confirm the high endemicity of the infection in the Kayapo indigenous group of the Brazilian Amazon.
Similar content being viewed by others
Backgrounds
The human T-lymphotropic viruses 1 and 2 (HTLV-1 and HTLV-2), members of the family Retroviridae, are retroviruses with tropism for T lymphocytes [1, 2]. HTLV-1 and HTLV-2 are believed to have been disseminated along human migratory pathways from Africa to Europe, Asia and the Americas, and they are currently distributed worldwide [3,4,5,6,7,8,9,10].
HTLV-2 is endemic in several African populations [8, 9] and in indigenous groups in North, Central and South America [6, 11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27]. Molecular studies have shown the existence of four molecular subtypes of HTLV-2: HTLV-2a, HTLV-2b, HTLV-2c and HTLV-2d based on the sequencing of the viral env gene and the 5′ long terminal repeat (LTR) region [5, 6, 15, 17, 28, 29].
HTLV-2 occurs with high prevalence among indigenous tribes of different linguistic families inhabiting the Brazilian Amazon, including in Kayapo villages, where subtype 2c was described for the first time [15, 17]. Later, its occurrence among the Kararaô, Gorotire, Tiriyo, Araral do laranjal and Zo’é indigenous populations was confirmed, and the same molecular subtype was found in urban populations of Brazil [6, 17, 30]. These results suggest that HTLV-2c is an endemic and autochthonous subtype in Brazil, particularly among indigenous populations of the Brazilian Amazon.
The geographical distribution of HTLV-2c in the Amazon region of Brazil ranges from the north (Tiriyo village) to the south (Kararaô and Gorotire villages), which suggests that its presence was derived during their settlement about 11,000 to 13,000 years ago during their migratory spread from the North to the South and the contact among different indigenous populations [6, 15]. In this context, the present study describes, for the first time, the occurrence of HTLV-2c and its dispersal in three new villages (Kateté, Djudjeko and Oodjã) of the Xikrin (Kayapo) ethnic group in the Brazilian Amazon region.
Methods
Study population and sample collection
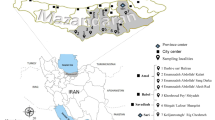
Blood samples (n = 263) were collected from individuals living in the Kateté (n = 121), Djujeko (n = 113) and Oodjã (n = 29) villages of the Xikrin tribe, Kayapo group, Jé linguistic family, who live in the Cateté Indigenous Territory, bordered by the Itacaiúnas and Cateté rivers, in the municipality of Parauapebas, state of Pará (Fig. 1). The study included individuals of both sexes aged 2–99 years old, from whom 10 mL of peripheral blood were collected in vacuum tubes containing ethylenediamine tetraacetic acid (EDTA) as anticoagulant for the separation of plasma and leukocytes. The samples were frozen at − 20 °C until subsequent analysis.

Map of the Brazilian Amazon region showing the geographic location of the Kateté, Djudjeko, Oodjã villages (Xikrin area) and Belem city, Capital of Para State (PA)
Ethical considerations
The project was approved by the National Committee for Research Ethics (CONEP, process n° 961.451/2015). Following the CONEP’s recommendations, the project received, through formal written permission, the approval and consent of the communities through their leaders on behalf of the participants (adults and Children’s parents) and the National Indian Foundation (FUNAI), exempting individual consent in writing in accordance with national regulations.
Serology
Plasma samples were tested for the presence of anti-HTLV-1/2 antibodies using an enzyme-linked immunosorbent assay (ELISA, Symbiosis Diagnostica LTDA, Leme, Brasil). The samples with positive or inconclusive results (presenting an optical density (OD) with values 20% below or above the cut-off value) were subjected to Real-Time PCR (Applied Biosystems, Foster City, CA, USA), for confirmation and differentiation between HTLV-1 and HTLV-2 infection.
Real-time PCR (qPCR)
DNA from the seropositive and inconclusive samples was extracted according to the manufacturer’s protocol of the AxyPrep™Blood Genomic DNA Miniprep extraction kit (Axygen Biosciences, CA, USA) to amplify the pol and 5′ LTR regions. The qPCR reactions were prepared using the TaqMan® Universal master mix according to the following protocol: 10 μL of MasterMix, 4.0 μL of water, 1 μL of Assay-by-Design (primer and probe set) and 5 μL of DNA in a final volume of 20 μL. The following cycling protocol was used: one cycle of 50 °C for 2 min, 95 °C for 10 min and 40 cycles of 95 °C for 15 s and 60 °C for 1 min. Two target sequences, an endogenous control (human albumin gene) and the non-homologous regions of the pol gene (186 bp) of HTLV-1 and HTLV-2, were used. The primer sequences used were: 5′-gaacgctctaatggcattcttaaaaccc-3′(HTLV-1F), 5′-gtggttgattgattgtccatagggctat-3′ (HTLV-1R), 5′-caaccccaccagctacagg-3′ (HTLV-2F), 5′-gggcatgacaggttttgcaatatta-3′ (HTLV-2R), 5′-gctcaactccctattgctattcaca-3′ (Albumin F), 5′-gggcatgagaggttttgcaatatta-3′; and the probe sequences used were FAM-5′-acaaacccgacctaccc-3′-NFQ (HTLV-1), FAM-5′-tcgagagaaccaatggtataat-3′-NFB (HTLV-2) and FAM-5′-ttgtgggtgtaatcat-NFQ (Albumin).
Nested PCR
Eight samples that were confirmed as positive for HTLV-2 were subjected to nested PCR reactions for amplification of the 5’LTR region for the later analysis of nucleotide sequencing and construction of phylogenetic trees. The 1st and 2nd round PCR reactions were run in a final volume of 50 μL containing 500 ng of extracted DNA, 10 μM of each dNTP, 20 pmol/μL of each primer, 50 μM MgCl2, 1 x buffer (50 mM KCl, 10 mM Tris-HCl pH 8.3) and 5 U of Taq DNA polymerase. In each amplification reaction, after initial denaturation at 94 °C for 5 min, 35 cycles were performed, with 40 s at 94 °C, followed by 30 s at 57 °C and 1 min at 72 °C. These 35 cycles were followed by a final extension for 10 min at 72 °C. The primers used were 5′-tcgcgatgacaatggcgactagcctc-3′ (F-IILTR) and 5′-gggggctttgggtattggagttggg-3′ (Long Gag) for the first round and 5′-gcctcccaagccagccac-3′ (Mo15) and 5′-gggaaagcccgtggatttgccccat-3′ (MSW-Gag) for the second round. The nested PCR products were visualized after electrophoresis (100 V/45 min) on 2% agarose gel in 1x TAE buffer (50x TAE stock solution - 1.6 M TrisBase, 0.8 M sodium acetate and 40 mM EDTA-Na2 in 1000 mL of deionized water) that contained 5 μL of ethidium bromide (10 mg/mL), using a transilluminator with an ultraviolet light source, and were subsequently purified using the manufacturer’s protocol of the QIAquick PCR Purification Kit (Qiagen, Inc., Valencia, CA, USA) for improving the nucleotide sequencing.
Analysis of nucleotide sequences
The analyses of the degree of genetic divergence between the nucleotide sequences and the phylogenetic profile of the strains were performed using the MEGA-6 - Molecular Evolutionary Genetics Analysis 6.0 software [31].
Phylogenetic analysis
The nucleotide sequences of the 5′ LTR region (642 nt) amplified in this study (DJU214 -MH194234, DJU218 - MH194235, OOD394 - MH194236, OOD396 - MH194237, KAT48 - MH194238, KAT147 - MH194239, KAT 116 - MH194240 and KAT46 - MH194241) were used to establish phylogenetic relationships with the HTLV-2 sequences that were previously described in the literature and are available in the GenBank. Sequence alignment was performed using the BioEdit software [32]. The degree of genetic diversity between the nucleotide sequences and the phylogenetic profile of the strains was analyzed using the MEGA-6 - Molecular Evolutionary Genetics Analysis 6.0 software [31]. The Neighbor-Joining method was used to construct the phylogenetic tree using the Kimura 2-parameters model. Statistical support of the phylogenetic tree was performed through bootstrap analysis, which generated 2000 replicates of the database.
Results
Serological findings
Antibodies to HTLV-1/2 were detected in 113 (43%) samples and 3 (1.1%) were undetermined (Table 1). The presence of HTLV-2 was confirmed by qPCR in 77 (29%) persons, with the following distribution: 46/121 (38%) from the Kateté village, 25/113 (22%) from the Djudjeko village and 6/29 (21%) from the Oodjã village. The indeterminate samples were negative on qPCR testing and no HTLV-1 infection was detected among samples tested.
The prevalence rate according to sex was 38% in women and 18% in men (Table 2). This difference was observed in two communities. In the Djudjeko village, the values found were highly significant (p = 0.0030), corresponding to an infection prevalence of 32% in women and 8.5% in men. In the Kateté village, the percentage was 45% in women and 28% in men (p = 0.0609). In Oodjã, the prevalence rates (33 and 15%, respectively) were not statistically significant (p = 0.3391).
A significant difference (p = 0.0005, Table 2) in HTLV-2 infection was observed when the cases were stratified by age. The prevalence was higher among individuals aged between 31 and 40 years (43%). No infection was observed in the 0- to 10-year-old age group. The difference according to age group was significant in the Kateté village (p = 0.0411).
A further association of infection with both age and sex was significant only in women (p = 0.0002, Table 3). The distribution of infection according to sex in each on age group showed significant differences between females and males in two groups aged 31–40 years (p = 0.017) and those over 70 years (p = 0.0019).
Molecular characterization
Eight samples were subjected to amplification of a 788 bp segment of the 5′ LTR region for sequencing and subsequent phylogenetic analysis. The sequencing and editing of nucleotide bases generated a 642-bp fragment from the 5 ‘LTR region.
The sequences included samples from the three villages and were aligned and compared to each other, as well as with the prototype strains of subtypes 2a (HTLV-2Mot.) [33] and 2b (HTLV-2NRA) [34]. The samples from the present study presented 99.9% similarity to each other, and when compared to prototypes 2a and 2b, the identity between the sequences was 96,7% and 94,6%, respectively. The generated tree showed that the eight samples (KAT46, KAT48, KAT116, KAT147, DJU214, DJU218, OOD394 and OOD396) were clustered with the molecular subtype HTLV-2c as part of a monophyletic clade (Fig. 2).

Rooted phylogenetic tree showing genetic relationships among the HTLV-2 strains of the samples identified in the present study (KAT46, KAT48, KAT116, KAT147, DJU214, DJU218, OOD394 and OOD396) and those available in the GenBank. STLV-2 strain (U90557-Afr) was used as outgroup. The tree was constructed using the Neighbor-Joining method after aligning the 642 nucleotides of the 5′ LTR region. The bootstrap test was applied using 2000 replicates
Discussion
In the present study the prevalence of 29% of HTLV-2 infection was found in three villages of the Xikrin tribe, Kayapo group: Kateté, Djujeko and Oodjã. The finding is consistent with previous results that demonstrated that the Kayapo group presents one of the highest endemic rates of HTLV-2 infection in the world [6, 11, 12, 14, 15].
Considering the results found for other indigenous peoples of Brazil, the prevalence rate found in the present study was higher. In the state of Paraná, in the Guarani group, the prevalence was previously reported as 5.8% [21]. More specifically, in the Brazilian Amazon region, lower rates were detected among the Kraho (12.2%), Mekranoiti (12.19%), Mundurukú (8.1%) and Macaw Laranjal Indians (11.4%) [11, 12, 15].
The prevalence found in the present study is similar to that observed in previous studies involving indigenous populations in the Americas. In Venezuela, the prevalence among the Guahibo [19] was estimated at 24.8%, and among the Seminole Indians in the United States, at 24% [24]. However, lower prevalence rates have been reported in the Yahgans in Chile (9.1%) [26], in the Wayuu in Colombia (4.8%) [35] and in the Wichi-Mataco in Argentina (above 3%) [25]. In two independent studies conducted in the Shipibo-Konibo group of the Peruvian Amazon [36], prevalence rates of 2.1 and 3.8% were found, but even lower prevalence rates were reported among the Nuu-Chah-Nulth of Canada (1.6%) and the Maya of Mexico (0.23%) [22, 27].
By contrast, the prevalence observed here was lower than those of several indigenous groups in the Americas. In Venezuela, the communities of Yaruro and Guahibo [20] had an estimated prevalence of 61%, and this rate is two times higher than that found in the present study. Higher prevalence rates were also found among the Chorote Indians in Argentina (35%), the Chulupi in Paraguay (34.0%) [18] and the Alcaf in Chile (34.8%) [26].
These differences in prevalence are expected and likely due to numerous factors, such as the: (i) differences in the methodologies adopted, which may have different sensitivities and specificities; (ii) differences in the sample sizes analyzed in each study; and (iii) differences in socio-cultural aspects that favor or limit the spread of the virus in the local population [6, 16]. Indigenous peoples, especially those in the Amazon, have undergone population fission and fusion events, which can contribute to increases or decreases in the incidence of the virus in a given community and often lead to the phenomenon known as the “founder effect”, where a small group of individuals (including virus carriers), due to internal conflicts over leadership, leave the tribe and start a new village. This situation, together with inbreeding, different women in the tribe breastfeeding children, polygamy and skin scarification rituals, may facilitate the interpersonal transmission of the virus and may consequently increase the prevalence of infection in the community [6]. These aspects may also explain the apparent increase in the prevalence of HTLV-2 infection in the Xikrin tribe when we compared our results with those of previous studies, which indicated a prevalence rate ranging from 13 to 15% [12, 13, 15].
It is generally agreed that HTLV infection is more easily transmitted from men to women, which would likely explain the higher prevalence of infection among women [18,19,20, 25, 37]. The first studies of HTLV-2 prevalence among the tribes of the Kayapo group also investigated the distribution of the infection according to gender [14, 15].
In the present study, there was a significant difference in infection prevalence between women and men, with the prevalence rate among women nearly twice the value in men. These results are in agreement with other findings in indigenous populations that found a predominance of infection among women. In the Peruvian Amazon, among the Shipibo-Konibo group, the reported seroprevalence was 3.1% in women and 2.5% in men [36]. In Venezuela, among the Yaruro and Guahibo, the prevalence of infection was 56% in men and 64% in women [20]. Among indigenous populations living in the Gran Chaco forest, the Chorote from Argentina and the Chulupi from Paraguay, there was a significant difference in infection between women (26.0%) and men (19.2%) [18]. Einsiedel et al. [38] reported, in central Australia, HTLV-1 infection associated with increasing age, male gender and sexual transmission infection in adults. The authors concluded that virus transmission from men to women was more efficient, which may also be suggested from the results of the present study.
However, the results also contrast with those of several studies conducted among indigenous groups in the Americas. Ishak et al. [15] did not observe differences in the prevalence rates between men (31.4%) and women (34.2%) when six tribes of the Kayapo indigenous groups were analyzed together. Similar results were observed in the Xikrin for women (11.4%) and men (11.7%) over the age of fourteen [14].
It is generally accepted that social and cultural characteristics are responsible to the differences of prevalence rates among native Indian communities, but these studies and the present results refer to three Kayapo Indians who share similar cultural behavior and common origin [14, 15]. For example, the hypothesized higher efficiency of male-to-female transmission commonly refers to the sexual route of transmission, and it is important to consider that this is not the only way by which the virus can be spread within a village.
Vertical transmission and breastfeeding are also common routes of spread among epidemiologically closed communities [6, 14, 15], which give support to the large dissemination of HTLV-2c among Kayapo children aged 0 to 9 years. It should be noted that in these villages, an infected lactating woman does not exclusively breastfeed her son or daughter, but she often feeds the offspring of other women from the tribe. In this sense, transmission via breastmilk could contribute to an equal distribution between the sexes. This was not described in the present study, which seems to suggest that a completely new behavior may have been introduced in the investigated communities. HTLV-2 is a persistent virus and the absence of infections among children 0 to 10 years (and in two groups up 20 years), may suggest that transmission of the virus is almost exclusively via sexual relations. The virus is apparently introduced in the community among young women who start to procreate in the second and third decades of life.
When the infection prevalence was analyzed by age group, a significant difference was observed between sexes. The highest prevalence was found in individuals between 31 and 40 years, and it was seven times higher than that observed in individuals aged 11 to 20 years and three times higher than that found in those aged 51–60 years. These findings are consistent with other studies conducted in other indigenous populations [15, 38]. Among the Toba, from Argentina, the prevalence found in individuals over forty years of age was 59%, which was four times higher than the infection rate in children (15.0%) younger than fifteen years [25]. In Amerindians living in the Gran Chaco forest in Argentina and Paraguay, the prevalence at 34 years was 26.3%, approximately twice as high as that found in children under the age of 13, whose prevalence rate was 14.2% [18]. In Venezuela, in the Guahibo group [19], there was an increase in infection prevalence with age, similar to that reported among the Kayapo [14, 15]. In the first study on the Kayapo, a prevalence of 60% was found in individuals over sixty years of age, approximately five times greater than that observed in children under fifteen years of age, which was estimated to be 12% [14]. In a second study among this group, conducted by Ishak et al. [15], the prevalence was 60% in individuals of both sexes older than 40 years and 21.6% in children under the age of nine. Thus, the results presented here, together with those of previous studies, reinforce the hypothesis that the sexual transmission of HTLV-2 is an important route of dissemination in the indigenous populations because its prevalence increases with age [14, 15, 18, 19].
The phylogenetic analysis is in agreement with previous studies that have demonstrated the endemic presence of HTLV-2, subtype c, in indigenous and urban populations of the Amazon [6, 15]. The samples formed a monophyletic clade with others that were previously identified as belonging to subtype 2c, thus confirming the isolated occurrence of this molecular subtype among the Kayapo.
A comparison of the nucleotide diversity of the 5’LTR region among the samples amplified in the present study identified a genetic similarity of 99.9% between them. When the analysis was performed considering the HTLV-2a Mot and HTLV-2b NRA prototype strains, the mean similarity was 96.7 and 94.6%, respectively. These results are similar to those observed among the Gorotire, Kararaô and Tiriyo indigenous communities and in the urban region of Belém [6]. Similarity of 99.4% was observed when comparing the nucleotide sequences of the LTR region between K96, isolated from the Kayapo Indians described as HTLV-2c and RP329 isolated from an inhabitant of the urban area in the state of São Paulo [37]. This subtype was also isolated in samples from HTLV-seropositive blood donors in the urban area of Porto Alegre, in the state of Rio Grande do Sul [30]. These results reveal the high degree of genetic similarity between the various HTLV-2c strains that have been isolated from Brazilian populations, which can be attributed to a unique, autochthonous origin and the miscegenation process that originated the neo-Brazilian population, as described recently by Vallinoto and Ishak [39].
Conclusions
The results of the present study reveal the incidence of HTLV-2 in three new villages of the Xikrin tribe, thus confirming the high endemic nature of the infection in the Kayapo indigenous group of the Brazilian Amazon.
Abbreviations
- CONEP:
-
National Committee for Ethics in Research
- EDTA:
-
Ethylenediamine tetraacetic acid
- ELISA:
-
Enzyme-linked immunosorbent assay
- FUNAI:
-
National Indian Foundation
- HTLV-1:
-
Human T-lymphotropic virus 1
- HTLV-2:
-
Human T-lymphotropic virus 2
- LTR:
-
Long terminal repeat
- OD:
-
Optical density
- qPCR:
-
Real-Time polymerase chain reaction
References
Yoshida M, Miyosh I, Hinuma Y. Isolation and charactezation of retrovirus from cells lines of human adult T-cell leukemia and its implication in the disease. Proct Natl AcadSCi. 1982;79(6):2031–5.
Gallo RC, Wong-Staal ASF. Origin of human T-cell leukaemia-lymphoma virus. Lancet. 1983;2(8353):962–3.
Hall WW, Zhu SW, Horal P, Furura Y, Zagaany G, Vahlne A. HTLV-II infection in Mongolia. AIDS Res Hum Retrovir. 1994;10:443.
Switzer WM, Pieniazek D, Swanson P, Samdal HH, Soriano V, Khabbaz RF, Kaplan JE, Lal B, Heneine W. Phylogenetic relationship and geographic distribution of multiple human T-cell lymphotropic virus type II subtypes. J Virol. 1995;69(2):621–32.
Salemi M, VAN Dooren S, Vandame A. Origin and evolution of human and simian T-cell lymphotropic viruses. AIDS Rev. 1999;1:131–9.
Vallinoto ACR, Ishak MOG, Azevedo V, Hall AC, Ishak R. Molecular epidemiology of human T- lymphotrpicvírus type II infection in Amerindian and urban populations of the Amazon Region of Brazil. Hum Biol. 2002;74(5):633–44.
Vallinoto ACR, Muto N, Pontes GS, Machado LFA, Azevedo VN, Santos SEB, Ribeiro-Dos-Santos AKC, Ishak MOG, Ishak R. Serological and molecular evidence of HTLV-I infection among Japanese immigrants living in the Amazon Region of Brazil. Jpn J Infect Dis. 2004;57(4):156–9.
Gessain A, Mauclere P, Fromenti A, Biglione M, Le Hesra NY, Tekaiat F, et al. Isolation and molecular characterization of a human T-cell lymphotropic virus type II (HTLV-II), subtype B, from a healthy pygmy living in a remote area of Cameroon: an ancient origin for HTLV-II in Africa. Proc Natl Acad Sci U S A. 1995;92(9):4041–5.
Mauclère P, Afonso PV, Meertens L, Plancoulaine S, Calattini S, Froment A, et al. HTLV-2B strains, similar to those found in several amerindian tribes, are endemic in central african Bakola pygmies. J Infect Dis. 2011;203(9):1316–23.
Gessain A, Cassar O. Epidemiological aspects and world distribution of HTLV-1. Front Microbiol. 2012;3:388.
Maloney EM, Biggar RJ, Neel JV, Taylor ME, Hahn BH, Shaw GM, Blattner WA. Endemic human T-cell lymphotropic vírus type II infection among isolated Brazilian Amerindians. J Infect Dis. 1992;166(1):100–7.
Nakauchi CM, Maruyama K, Kanzadi LI, Linhares AC, Azevedo VN, Fukushima T. Prevalence of HTLV-I antibody among two distinct ethnic groups inhabiting the Amazon region of Brazil. Rev Inst Med Trop Sao Paulo. 1992;34(4):323–8.
Gabbai AA, Bordin JO, Vieira-Filho JPB, Kuroda A, Oliveira ASB, Cruz MV, et al. Selectivity of human T Lymphotropic virus Type-1 (HTLV-1) and HTLV-2 infection among different population in Brazil. Am J Trop Med Hyg. 1993;49(6):664–71.
Black FL, Biggar RJ, Neel JV, Maloney EM, Watersd J. Endemic transmission of HTLV type II among Kayapo Indians of Brazil. AIDS Res Hum Retrovir. 1994;10(9):1165–71.
Ishak R, Harrington WJ Jr, Azevedo VN, Eiraku N, Ishak MOG, Guerreiro JF, et al. Identification of human T cell Lymphotropic virus type IIa infection in the Kayapo, an indigenous population of Brazil. AIDS Res Hum Retrovir. 1995;11(7):813–21.
Ishak R, Vallinoto ACR, Azevedo VN, Vicente A, Hall WW, Ishak MOG. Molecular evidence for infection by HTLV-2 among individuals with negative serological screening tests for HTLV antibodies. Epidemiol Infect. 2007;135(4):604–9.
Eiraku N, Novoa P, Ferreira MC, Monken C, Ishak R, Ferreira OC, et al. Identification and characterization of a new and distinct molecular subtype of human T-cell lymphotropic virus type 2. J Virol. 1996;70(3):1481–92.
Ferrer JF, Esteban E, Dube S, Basombrio MA, Segovia A, Peralta-Ramos M, et al. Endemic infection with human T cell leukemia/lymphoma virus type IIB in Argentinean and Paraguayan Indians: epidemiology and molecular characterization. J Infect Dis. 1996;174:944–53.
Leon-Ponte M, Oscar N, Bianco B, De Perez GE. Highly endemic human T-lymphotropic virus type II (HTLV-II) infection in a Venezuelan Guahibo Amerindian group. J Acquir Immune Defic Syndr Hum Retrovirol. 1996;13(3):281–6.
Leon-Ponte M, Perez GE, Bianco N, Hengst J, Dube S, Love J, et al. Endemic infection with HTLV-IIb in Venezuelan Indians: molecular characterization. J Acquir Immune Defic Syndr Hum Retrovirol. 1998;17(5):458–64.
Menna-Barreto M, Bender AL, Bonatto SL, Freitas LB, Salzano FM, Tsuneto LT, et al. Human T-cell lymphotropic virus type II in Guarani Indians, Southern Brazil. Cad Saúde Pública. 2005;21(6):1947–51.
Peters AA, Coulthart MB, Oger JF, Waters DJ, Crandall KA, Baumgartner AA, et al. HTLV type I/II in British Columbia Amerindians: a seroprevalence study and sequence characterization of an HTLV type IIa isolate. AIDS Res Hum Retrovir. 2000;16(9):883–92.
Hjelle B, Zhu SW, Igichi S, Takahashi H, HAL WW. Endemic HTLV-2 infection in southwestern U.S. Indians involves two prototype variants of vírus. J Infect Dis. 1993;168(3):737–40.
Lowis G, Sheremata WA, Wickman PR, Dube S, Dub DK, Poiesz BJ. HTLV-II risk factors in native Americans in Florida. Neuroepidemiology. 1999;18(1):37–47.
Biglione M, Vidan O, Mahieux R, Colombo M, Basualdo MLA, Bonnet M, et al. Seroepidemiological and molecular studies of human T cell lymphotropic virus type II, subtype b, in isolated groups of Mataco and Toba Indians of Northern Argentina. AIDS Res Hum Retrovir. 1999;15(5):407–17.
Fujiyoshi T, Li H-C, Lou H, Yashiki S, Karino S, Zaninovic V, et al. Characteristic distribution of HTLV type I and HTLV type II carriers among native ethnic groups in South America. AIDS Res Hum Retrovir. 1999;15(14):1235–9.
Gongora-Bianchi RA, Lal RB, Rudolph DL, Castro-Sansores C, Gonzalez-Martinez P, Pavia-Ruz N. Low prevalence of HTLV-II in Mayan indians in the Yucatan Peninsula, Mexico. Arch Med Res. 1997;28(4):555–8.
Hall WW, Takahashi H, Liu C, Kaplan MH, Scheewind O, Ijichi S, et al. Multiple isolates and characteristics of human T-cell leukemia virus type II. J Virol. 1992;66(4):2456–63.
Vandamme A-M, Salemi M, Brussel MV, Liu H-F, Laethem KV, Ranst MV, et al. African origin of Uman T-lymphotropic virus type 2 (HTLV-2) supported by a potential new HTLV-2d subtype in congolese Bambuti Efe pygmies. J Virol. 1998;72(5):4327–40.
Renner JDP, Laurino JP, Menna-Barreto M, Schmitt VM. Molecular evidence of HTLV-II subtype B among an urban population living in South Brazil. AIDS Res Hum Retrovir. 2006;22(4):301–6.
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol. 2013;30(12):2725–9.
Hall TA. BioEdit: a user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucl Acids Symp Ser. 1999;41:95–8.
Shimotohno K, Takahashi Y, Shimizu N, Gojobori T, Golde DW, Chen ISY, et al. Complete nucleotide sequence of an infectious clone of human T-cell leukemia virus type II: an open reading frame for the protease gene. Proc Natl Acad Sci U S A. 1985;82(10):3101–5.
Lee H, Idler KB, Swanson P, Aparicio JJ, Chin KK, Lax JP, et al. Complete nucleotide sequence variation of HTLV-II isolates from U.S. blood donors and Italian intravenous drug users. Virology. 1993;196(1):57–69.
Dueñas-Barajas E, Bernal JE, Vaught DR, Briceño I, Clemencia Duran C, Yanagihara R, et al. Coexistence of human T-lymphotropic virus types I and II among the Wayuu Indians from the Guajira region of Colombia. AIDS Res Hum Retrovir. 1992;8(11):1851–5.
Alva IE, Orellana ER, Blas MM, Bernabe-Ortiz A, Cotrina A, Chiappe M, et al. HTLV-1 and -2 infections among 10 indigenous groups in the Peruvian Amazon. Am J Trop Med and Hyg. 2012;87(5):954–6.
Covas DT, Kashima S. Complete nucleotide sequences of the genomes of two Brazilian specimens of human T lymphotropic virus type 2 (HTLV-2). AIDS Res Hum Retrovir. 2003;19(8):689–97.
Einsiedel L, Woodman RJ, Flynn M, Wilson K, Cassar O, Gessain A. Human T-Lymphotropic virus type 1 infection in an indigenous Australian population: epidemiological insights from a hospital-based cohort study. BMC Public Health. 2016;16:787–97.
Vallinoto ACR, Ishak R. HTLV-2: an ancient infection among Indian people of Brazilian Amazon Region. Rev Pan-Amaz Saúde. 2017;8(2):9–11.
Acknowledgements
We express our thanks to all participants of the Xikrin tribe.
Funding
The present study was fully supported by grants from Conselho Nacional de Desenvolvimento Científico e Tecnológico – CNPQ (Grant #302582/2013–4) and Universidade Federal do Pará (PAPQ/2018). The funding source(s) had no involvement in the conduct of the research and/or preparation of the article.
Availability of data and materials
All data generated during this study are included in this published article. The sequences are available at GenBank (https://www.ncbi.nlm.nih.gov/nuccore/) under the accession codes (DJU214 -MH194234, DJU218 - MH194235, OOD394 - MH194236, OOD396 - MH194237, KAT48 - MH194238, KAT147 - MH194239, KAT 116 - MH194240, KAT46 -MH194241).
Author information
Authors and Affiliations
Contributions
ACRV, MOGI, JFG and RI designed the study. JFG was the general coordinator of the project. ILJB, KSGS, MW, and MAFQ provided technical assistance and executed the experiments. ILJB, SSL and MAFQ analyzed all data. ILJB, ANRS, ACRV and IMVC wrote the manuscript with input from all authors. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The project received the approval and consent of the communities through their leaders, with formal collective authorization and from the National Indian Foundation (FUNAI). It was approved by the National Committee for Ethics in Research (CONEP), process 961.451/2015.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Braço, I.L.J., de Sá, K.S.G., Waqasi, M. et al. High prevalence of human T-lymphotropic virus 2 (HTLV-2) infection in villages of the Xikrin tribe (Kayapo), Brazilian Amazon region. BMC Infect Dis 19, 459 (2019). https://doi.org/10.1186/s12879-019-4041-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12879-019-4041-0