
Abstract
Background
The interaction among plants and their pollinators has been a major factor which enriched floral traits known as pollination syndromes and promoted the diversification of flowering plants. One of the bee-pollination syndromes in Faboideae with keel blossoms is the formation of a landing platform by wing and keel petals. However, the molecular mechanisms of elaborating a keel blossom remain unclear.
Results
By performing large scale mutagenesis, we isolated and characterized a mutant in Vigna radiata, love on wings (low), which shows developmental defects in petal asymmetry and vasculature, leading to a failure in landing platform formation. We cloned the locus through map-based cloning together with RNA-sequencing (RNA-seq) analysis. We found that LOW encoded a nucleus-localized Dof-like protein and was expressed in the flower provascular and vascular tissues. A single copy of LOW was detected in legumes, in contrast with other taxa where there seems to be at least 2 copies. Thirty one Dof proteins have been identified from the V. radiata’s genome, which can be further divided into four Major Cluster of Orthologous Groups (MCOGs). We also showed that ectopic expression of LOW in Arabidopsis driven by its native promoter caused changes in petal vasculature pattern.
Conclusions
To summarize, our study isolated a legume Dof-like factor LOW from V. radiata, which affects vasculature development in this species and this change can, in turn, impact petal development and overall morphology of keel blossom.
Similar content being viewed by others

Background
The majority of flowering plants have different strategies in order to attract pollinators, such as alterations in floral color, size, scent, nectar as well as shape. These changes are, in turn, under selection by different pollinators, resulting in a collection of floral traits known as pollination syndromes [1]. It has been proposed that Faboideae species with keel blossoms show adaptation towards bee pollination [2,3,4,5]. Different petals on a keel blossom play different roles in terms of pollination: the dorsal petal (or vexillum or standard or flag) acts as a billboard to attract pollinators; the ventral petals (or keel or carina) provide a space which protects the sporophyll column; and the lateral petals (or wing or alae) together with the ventral petals form a wing-keel complex, serving as a landing platform for the insects [4, 5]. Although we already know that CYCLOIDEA-like (CYC-like) genes are involved in the differentiation of petals along dorsal-ventral axis, it is still unclear how the elaborate petal shape is formed and how it leads to the genesis of a landing platform [6,7,8].
Organ shape and vasculature are closely linked during the evolution of flowering plants [9, 10]. The analyses of mutants with abnormal shape and vasculature in assorted lateral organs have provided new insights on the relationship between them [11, 12]. During leaf organogenesis, the final leaf form temporally coordinates with the formation of major veins, whereas the pattern of the minor veins does not completely reflect the final leaf shape [9]. Further studies unveiled complex mechanisms and genetic networks in the control of vascular tissue development, coordinated by different phytohormones, several signal peptides and multiple transcription factors [13,14,15,16]. Nevertheless, most of the conclusions are drawn from limited model species. Hence, the scenarios in other plants are still obscure, especially when referring to the origin of novel developmental traits, such as keel blossoms.
The Dof genes encode plant-specific transcription factors, which have a highly conserved DNA-binding Dof domain [17,18,19,20]. Dof genes are ubiquitous in angiosperms, gymnosperms and other early diverged lineages such as moss and algae. However, the number of Dof genes is highly variable among green plants and tends to be proportional with morphological complexity of plant species [17, 20]. Many Dof genes (20 out of total 36 in Arabidopsis thaliana) are expressed in the vascular system, suggesting their roles during the development and function of vascular tissues [21, 22]. In Arabidopsis, different sub-clades of Dof2.4 and Dof5.8 are expressed in distinct early stages of leaf vasculature: Dof2.4 is highly expressed in the primary vein of leaf primordia, while Dof5.8 shows high expression in both primary and secondary veins, as well as petal vasculature, stamens and carpels [23, 24]. No apparent phenotype was observed in the single mutant of dof5.8, but it enhanced the cotyledon vascular defects of a weak allele of auxin response factor 5–2, indicating that Dof5.8 functions in an auxin-dependent regulation [25]. Another close paralog Dof3.4, or DOF TF OBF BINDING PROTEIN 1, which shows similar expression to Dof5.8, may act redundantly in the control of leaf vascular development [26]. Dof5.6 or HIGH CAMBIAL ACTIVITY2, another sub-clade of Dof transcription factors, predominantly exists in the vascular tissues of assorted organs and its gain-of-function mutant shows pleiotropic morphological changes including increased cambial activity [27]. A recent study found that cytokinin promotes the expression of a group of Dof genes designated as PHLOEM EARLY DOF in the procambial tissue, including Dof1.1, Dof2.4, Dof3.2, Dof5.1, Dof5.3 and Dof5.6 [28]. Multiple loss-of-function Arabidopsis Dof mutants exhibit variably reduced radial growth around early protophloem-sieve-element cells, causing further reduction of cell number in root vasculatures [28].
In this study, we evaluated a legume crop Vigna radiata, also known as mung bean, which is of great economic importance in Asia. Unlike classic Faboideae species with zygomorphic flowers, a part of Vigna spp. including V. radiata, have a left-handed asymmetrical flower with the left wing-keel complex generating a landing platform [29]. By large scale mutagenesis, we isolated and characterized a floral mutant love on wings (low), whose left-wing petal attaches to the ventral petal and thus, leads to a failure in landing platform formation. We observed abnormality in the petal vasculature accompanied with changes in petal shape and asymmetry. We further cloned the LOW locus, which encodes a plant specific Dof-like transcription factor localized to the nucleus and expressed in the flower vascular tissues. A single copy of LOW was detected in legumes in contrast with other taxa, and we found that ectopic expression of LOW in Arabidopsis disrupted the petal vasculature. Altogether, we infer that LOW plays an essential role in floral vascular development of keel blossoms.
Results
V. radiata has a left-handed keel blossom
The wild-type (WT) V. radiata flower exhibits a left-handed keel blossom (Additional file 1: Figure S1). The right lateral petal encloses the right ventral petal, whilst the left lateral petal is inflexed over the spur developed on left ventral petal, and together they form the landing platform on the left side of the flower (Additional file 1: Figure S1A). We observed that the honeybee alighted on the left landing platform and forced its head towards the base of the dorsal petal, where there is a narrow gap for the insect to insert its proboscis to the nectary (Additional file 1: Figure S1B-C).
Characterization of the love on wings (low) mutant
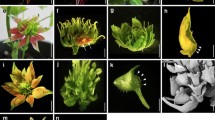
Using large-scale gamma ray mutagenesis, we characterized one mutant, which showed defects in the landing platform formation (Fig. 1). In the mutant, unlike the WT flower, lateral petals “hugged” ventral petals tightly, thus we named this mutant love on wings (low). There were basically two different types of flowers on the low mutant: the mild type (51 out of 100), exhibited right lateral petal development similar to the WT, but the left lateral petal enclosed the left ventral petal, hindering the formation of the left wing-keel landing platform (Fig. 1b); the other type (49 out of 100) showed severe developmental defects, in which the petal arrangement was so defected since the ventral petal enclosed the lateral petals (Fig. 1c). The two floral morphologies ratio was approximately 1:1.
Then, we dissected the newly opened V. radiata flower and examined the morphologies of different floral organs. In both types of the mutant flowers, the shapes of lateral and ventral petals had changed (Fig. 1). In the WT flower, two bulged structures grew outwards the base of lateral petals; however, there were three bulged structures in the mutant’s lateral petals (Fig. 1). Both the right and left lateral petals in the mutant became more curved with more symmetrical petal shapes, compared with the WT flower (Fig. 1b-c). Moreover, in the WT flower, two ventral petals formed a keel structure, while in the mutant, a single ventral petal developed into a keel-like shape, similar to the phenotype of a pea mutant, symmetric petals 1 [7]. We did not find any obvious morphological abnormality in other floral organs.
Since the plant organ shape is closely associated with organ vasculature, we then examined the petal vascular pattern in WT and mutant flowers. We dissected the 2 mm and 5 mm flower buds together with 12 mm mature flowers (Fig. 2). We found that in all the developmental stages we examined, lateral petal shape of the WT flower was more asymmetric compared with the mutant lateral petal (Fig. 2). This phenotype is consistent with changes in petal vascular pattern, especially in the main veins (Fig. 2). Petal internal asymmetry and the asymmetric vasculature were further enhanced along with the developmental processes (Fig. 2).
As mentioned before, the single ventral petal in the mutant developed into a keel-like structure. This is also evident when we observed the ventral petals from 2 mm or 5 mm flower buds. Additional tissue of ventral petal developed in the 2 mm mutant flower (Fig. 2b). In 5 mm stage, the WT ventral petal exhibited a kidney-like shape and the spur on the left petal has not emerged yet (Fig. 2a). However, additional tissues were further grown on the opposite side of the mutant, forming a keel (Fig. 2b). We also noticed that vascular pattern on the ventral petal was also abnormal compared with the WT (Fig. 2). These results favor the hypothesis that changes in petal shape are linked to the defects in petal vasculature.

Flowers of in wild-type (WT) and the love on wings (low) mutant. a A WT Vigna radiata flower. b-c Two types of mutant flowers. DP, dorsal petal; LP, lateral petal; VP, ventral petal. The white arrows mark the bulged structures in the base of right later petals. Bars = 10 mm

Petal vasculature in wild-type (WT) and mutant. a Petals from WT flowers. b Petals from mutant flowers. 5 LP, 5 mm lateral petal; 5 VP, 5 mm ventral petal; 12 LP, 12 mm lateral petal. Bars = 2 mm
Cloning and phylogenetic analysis of LOW
To generate the M2 mapping population, we crossed the original mutant with another cultivar, AL127. Mutation Mapping Analysis Pipeline for Pooled RNA-seq method based on 40 individuals with mutant phenotype suggested that a large region on chromosome 7 would be the possible site where the LOW is located (Fig. 3a). LOW locus was further mapped and located between two markers, M9 and M10 (Fig. 3b). There are 54 putative genes between them and we found one candidate gene (Vr07g10060/LOC106767037) significantly down regulated in the mutant (Fig. 3b). Vr07g10060/LOC106767037 encodes a Dof-like transcription factor, and we detected that in the low mutant, there was a 2 base-pairs substitution followed by 11 base-pairs deletion in the Dof domain of Vr07g10060/LOC106767037, leading to a frame-shift and precocious termination of transcription (Fig. 3c and Additional file 2: Figure S2). Subcellular localization assay using Arabidopsis protoplasts demonstrated that green fluorescent protein fused LOW protein was co-localized with a nucleus marker, indicating its function possibly as a transcription factor (Additional file 3: Figure S3).
We further analyzed its orthologous proteins in different eudicots lineages (Fig. 3d). In the basal eudicot Aquilegia coerulea, only one copy was detected named AcDof1. At least one independent duplication event occurred within the diversification of rosids Salicaceae, Brassicaceae and asterids Solanaceae (Fig. 3d). However, in rosids Fabaceae, except for Glycine max, in which ancient whole genome duplication once occurred, only one ortholog of LOW exists in each legume’s genome (Fig. 3d).
To identify the DOF proteins from the mung bean genome, the consensus amino acid sequence of Dof domain was used to BLAST (Basic Local Alignment Search Tool) against its genome database on Legume Information System (https://legumeinfo.org/). Thirty one Dof proteins have been identified and all of them contain a typical Dof DNA binding domain (Additional file 4: Figure S4). To evaluate the evolutionary history among the 31 mung bean Dof proteins, we performed a phylogenetic analysis using their full length protein sequences. Phylogeny tree of these proteins indicated that Dof family have undergone multiple times of duplication (Fig. 4). Based on a previous study [30], the mung bean Dof proteins were divided into four Major Cluster of Orthologous Groups (MCOGs), which could be further divided into multiple subgroups supported by high bootstrap values and motif analysis (Fig. 4). We noticed that although LOW belongs to the MCOG Dd group, its sequence is quite different from other MCOG Dd members, indicating early divergence of this Dof protein (Fig. 4).

Cloning and phylogeny analysis of LOW. a Association analysis based on Mutation Mapping Analysis Pipeline for Pooled RNA-seq method. The red region on chromosome 7 indicates strongest association and the red arrow marks the chromosome. b Physical map of the large region in Chromosome 7 of Vigna radiata, where LOW is located. Marker information (M) and recombination frequency (Rec.) are shown. In the lower lane, relative expression heat map of candidate genes between M9 and M10 is shown; the red arrow marks the Vr07g10060/LOC10676703. c The gene structure of LOW, nucleotide numbers, start and terminal codons are shown; the red arrow indicates the mutation. d Maximum-likelihood tree of LOW-like Dof genes from Aquilegia coerulea (Ac), Arabidopsis thaliana (At), Brassica rapa (Br), Capsella rubella (Cr), Glycine max (Gm), Medicago truncutula (Mt), Populus trichocarpa (Pt), Phaseolus vulgaris (Pv), Solanum lycopersicum (Sl), Solanum tuberosum (St) and Vigna radiata (Vr). 1000 times of bootstrap (value in percentage) is marked at each node and the accession number is presented in the parentheses of each sequence; the red branches highlight LOW and its homologs within legume species; the red triangle marks the LOW and blue circles indicate two paralogues from Arabidopsis. DOF5.6 was chosen as an outgroup

Neighbor-joining tree (left) and motif analysis (right) of 31 Dof proteins from Vigna radiata genome. Bootstrapping value is located in each node as percentages (when > 50%) along the branches. Four Major Clusters of Orthologous Genes (MCOG) are shown in different colors. The motif diagrams were generated in MEME, and different colors representing different motifs are shown below
The spatial-temporal expression pattern of LOW
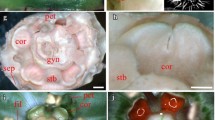
We extracted RNA from various plant tissues, and through qRT-PCR (Quantitative Reverse Transcription Polymerase Chain Reaction), found that LOW was highly expressed in the inflorescence with up to 2 mm flower buds (Additional file 5: Figure S5). The expression of LOW was rapidly decreased in later flower buds indicating that LOW may function in early flower developmental stages (Additional file 5: Figure S5). We further examined the spatial-temporal expression pattern of LOW by RNA in-situ hybridization (Fig. 5a-j). The mRNA of LOW accumulated specifically in the central veins of flower organ primordia, including petals, stamens and carpels of early developmental stages (Fig. 5a-f). The longitudinal section of a late stage flower bud showed that LOW was expressed in the petals with discontinuously dot-like signals, indicating its expression in secondary petal veins (Fig. 5g). In the transverse sections of a late stage flower bud, the signals of LOW were accumulated in defined narrow regions within the petals, which were parallel with the whole flower plane (Fig. 5h-i). Moreover, the mRNA of LOW was detected in the anther tapetum, central ovary and ovules within a late flower bud (Fig. 5h-i).
A 2 kb DNA fragment of the LOW promoter region was fused to a GUS (β-glucuronidase) reporter gene (designated as LOWp:GUS) and then transformed into Arabidopsis. We detected strong GUS activity in the floral vasculature including pedicels, sepals, petals, filaments, styles and carpels (Additional file 6: Figure S6). The expression pattern of LOW in Arabidopsis system is similar to its native expression in V. radiata, suggesting that functional analysis of LOW in A. thaliana may help to understand its roles in V. radiata.

The spatial-temporal expression pattern of LOW. a Flower organogenesis observed under a scanning electronic microscopy, the dot lines and the black arrow represent the places and direction, where five consecutive longitudinal sections (b-f) were made; dp, dorsal petal primordium; lp, lateral petal primordium; vp, ventral petal primordium; st, stamen primordium; c, carpel primordium. b-j The spatial-temporal expression pattern of LOW in wild-type (WT) Vigna radiata detected by RNA in-situ hybridization. b-j are longitudinal sections of an early flower bud; g shows a longitudinal section of an late flower bud; (h-i) show transverse sections of a late flower bud; red, green and blue arrows in (i) mark the tapetum, ovary and ovule, respectively. Dark brown regions in (b-i) represent signals detected by LOW antisense probe; (J) is a longitudinal section of an early flower bud detected by sense probe of LOW as a negative control. Bars = 100 μm
Floral phenotypes of transgenic Arabidopsis
Since the 2 kb LOW promoter showed specific expression in the floral vasculature of Arabidopsis, we further ectopically expressed LOW (designated as pLOW::LOW) driven by its own 2 kb promoter. Fifteen independent transgenic lines were obtained, and we carefully examined the floral morphology of each line. Petal shape in the transgenic lines was similar to the wild type plants (Fig. 6a-d). However, when comparing the petal vasculature, we found that in the WT, vascular strands usually formed four enclosed vascular loops emanating from the midvein, while in the pLOW::LOW lines, vascular strands failed to form loops (Fig. 6a-d). These results corroborate that LOW functions in floral vasculature pattern.

Floral morphology of Col-0 (a) and three independent transgenic Arabidopsis thaliana lines of pLOW::LOW (b-d). For each plant, the upper panel exhibits an intact flower and the lower panel shows the petal vasculature. Bars =2 mm
Discussion
Co-evolution between plants and their pollinators involves changes of multiple genes among species. Although shift from one pollination syndrome to another requires complex genetic changes, it indeed occurred frequently beyond our expectations. In snapdragon, the ventral petal supported by the lateral petals is incurved at the region named hinge between the petal tube and lobe, forming a landing platform for the insects [31]. A MIXTA-like gene AmMYBML1 reinforces the specialization of ventral petal hinge and thus the formation of the landing platform [31]. In another Lamiales species, Torenia fournieri, an ALOG family homolog TfALOG3 is essential for the development of the corolla neck, which may protect their nectar reward to pollinators [32]. In this study, we identified another class of factors from V. radiata involved in petal elaboration and keel blossom pattern.
Organ asymmetry has been thought to be evolved independently multiple times [33]. In terms of petal, internal asymmetry can be observed either, in dorsal and lateral petals (i.e. snapdragon and wishbone flower), or in lateral and ventral petals (i.e. many keel blossoms). The first factor related to organ asymmetry was characterized in snapdragon. A CYC-like gene, DICHOTOMA, is expressed in the dorsal half of the dorsal petal primordia [33]. The cyc dich double mutant possesses five symmetric ventralized petals, favoring that the CYC-like factors couple floral dorsiventral asymmetry and petal internal asymmetry in Antirrhinum [33]. Unlike snapdragon, the ventral petal of a typical keel blossom is asymmetric, thus the ventralized petal should also be asymmetric. This is evident in the Lotus japonicus CYC triple mutants, where all petals become asymmetric, indicating that the floral organ internal asymmetry of a keel blossom is also related to CYC-like factors [34]. In pea, we previously isolated several mutants with defects in petal asymmetry, symmetric petal 1, symmetric petal 5, elephant ear-like leaf 1 and bigger organs. In syp1–1, the petals are bilaterally symmetric and increased organs are developed among approximately 1/3 of the flowers, with abnormal primordia initiation found during early developmental stages [7]. Similar to symmetric petal 1, mutations in ELEPHANT EAR-LIKE LEAF 1 and BIGGER ORGANS also exhibit several defects in petal asymmetry; these two proteins physically interact with each other and may act in the same genetic pathway [35]. In symmetric petal 5 and a weaker allele of bigger organs, later petals in these mutants become more symmetric compared with the WT’s, and genetic analysis suggests that these two factors act in an addictive manner [35]. Nevertheless, unlike the low mutant, these mutants display other pleiotropic phenotypic defects [7, 35, 36].
In the low mutant, we only observed the morphological abnormalities in flower perianth, where organ asymmetry in lateral and ventral petals were abolished (Fig. 1). We also found that changes in the asymmetry of vasculature can impact, somehow, the shape of asymmetric petals (Fig. 2), suggesting that petal vasculature development and floral dorsiventral asymmetry may interact with each other, possibly through a direct or indirect regulation of CYC genes or other genes involved in floral asymmetry. The transgenic Arabidopsis lines that bare the LOW’s promoter and its coding sequence, do not show any obvious changes in petal symmetry (Fig. 6). This could be due to the fact that the LOW construct was introduced into a heterologous system (A. thaliana) where endogenous CYC genes are likely differentially expressed and regulated, compared with what happens in V. radiata and other zygomorphic Fabaceae flowers.
LOW encodes a plant-specific Dof-like transcription factor. Various numbers of Dof genes have been found in different plant genomes with different expression patterns [17, 24]. Dof transcription factors play completely distinct roles in plant-specific processes, including light responsiveness, circadian rhythm, seed development, cell cycle regulation, phenylpropanoid metabolism, branching and vascular development [17, 18]. LOW was predominantly expressed in the floral vasculature (Fig. 5), which is similar but more specific comparing with the expressions of its orthologs Dof3.4 and Dof5.8 in Arabidopsis [23, 26]. According to the phylogenetic tree of Vigna Dof proteins, only the MCOG Dd clade that LOW belongs to has strong support, the other clades needs more phylogenetic analyses (Fig. 4). An interesting question is why we observed so specific floral phenotypes in the low mutant. Phylogenetic analysis of LOW’s orthologs suggested that this sub-clade of genes has undergone extensively duplication among many other plant lineages including Brassicaceae (Fig. 3d), which might explain the non-redundant function of LOW in mung bean. Since the expression of LOW is more specific and pLOW::LOW transgenic Arabidopsis only shows abnormal vascular pattern rather than shape change, we assume that the role of LOW in vascular patterning is ancient, while its role in petal morphology may be an evolutionary novelty.
Dof-like transcription factors work as either transcriptional activators or repressors by binding to the sequences containing the core AAAG motif [18, 37,38,39,40,41]. In A. thaliana, a Dof-like transcription factor DOF4.2 negatively affects flavonoid biosynthesis by repressing expression of genes such as FLAVONOL-3-HYDROXYLASE, DIHYDROFLAVONOL REDUCTASE and LEUCOANTHOCYANIDIN DIOXYGENASE, while positively influences the accumulation of hydroxycinnamic acids by promoting the expression of genes including PHENYLALANINE AMMONIA LYASE, CINNAMATE-4-HYDROXYLASE and 4-COUMAROYL-COA LIGASE 5 [37]. In Pinus pinaster, the PpDof5 transcription factor may regulate the expression of glutamine synthetase (GS) genes by activating the transcription of the GS1b, or in contrast, by repressing the expression of GS1a [38]. In the moss Physcomitrella patens, two Dof-like transcription factors, PpDof1 and PpDof2, show transcriptional repressor activities in protoplast transient assays [40]. In the fruit banana Musa acuminata, a Dof transcription factor MaDof23 works as a repressor, acting antagonistically in regulating ripening-related genes associated with cell wall degradation and aroma formation [41].
Conclusions
To summarize, we have characterized a legume Dof gene, LOW, which is involved in the differentiation of keel blossom by regulating floral vasculature pattern and petal internal asymmetry of mung bean. In the future, it is of interest to study how LOW regulates the petal vasculature and organ asymmetry at a molecular, genetic and developmental level.
Methods
Plant materials and map-based cloning
Two cultivars of V. radiata, Sulu and AL127, have been purified by selfing for three generations in a greenhouse at 28 ± 2 °C with a 16 h-light/8 h-dark photoperiod at 200 μmol m− 2 s− 1. A. thaliana Col-0 were grown at 20 ± 2 °C with a 16 h-light/8 h-dark photoperiod at 150 μmol m− 2 s− 1. The seeds of Sulu, AL127 and A. thaliana Col-0 were obtained from the germplasm bank in our lab.
The gamma ray mutagenesis was performed as we previously described [42]. low mutant was isolated from the M2 population of the mutagenized cultivar Sulu background. A 576 F2 mapping population was produced by crossing low (from the sulu background) to AL127. RNA-seq libraries based on the published genomic data from 40 individuals with mutant phenotype were generated using Mutation Mapping Analysis Pipeline for Pooled RNA-seq method [43, 44]. This result suggested that a large region on chromosome 7 would be the possible site where the LOW mutation is mapped. Low was further mapped with the F2 population based on the marker information that we published previously [45]. The primer sequences used in mapping are listed in the Supporting Information (Additional file 7: Table S1).
Microscopy
Inflorescences or different flower buds were fixed in FAA (3.7% formaldehyde, 50% ethanol, 5% acetic acid) fixative solution prior to clearing in 95% ethanol. Floral organs from buds in a series of developmental stages were dissected and observed under a light or florescence microscope. Petal vasculatures of 5 mm buds and mature flowers became visible under a light microscope after fixation and clearing, while petals from 2 mm buds were observed under the ultra violet laser. For scanning electron microscopy, fixed samples were treated and observed under the Jeol JSM 6360LV (Jeol, Tokyo, Japan) scanning electron microscope as previously reported [46]. Adobe PHOTOSHOP CS6 (Adobe, San Jose, CA, USA) was used to adjust the contrast of the images.
Phylogeny analysis, motif-based sequence analysis and subcellular localization
For phylogeny analysis of Dof-like family, protein sequences were obtained from the genomic database of Medicago truncatula (Mt4.0) and Vigna radiata (Vr1.0) in the Legume Information System (https://legumeinfo.org/home), or the Arabidopsis Information Resource (https://www.arabidopsis.org/). Amino acid sequences were aligned using CLUSTALW or MUSCLE followed by the generation of a Neighbor-joining tree with 1000 bootstrap replicates in MEGA6 [47]. Further analysis of LOW sub-clade Dof-like factors, nucleotide sequences from Aquilegia coerulea, Arabidopsis thaliana, Brassica rapa, Capsella rubella, Glycine max, Medicago truncatula, Populus trichocarpa, Phaseolus vulgaris, Solanum lycopersicum, Solanum tuberosum and Vigna radiata were obtained from the Phytozome 12 (https://phytozome.jgi.doe.gov/pz/portal.html#). Maximum-likelihood trees of these genes were also generated with 1000 times of bootstrap in MEGA6 [47]. Dof protein sequences were submitted to motif-based sequence analysis website (MEME; http://meme-suite.org/tools/meme) for motif mining under the parameters: -time 18,000, −mod zoops, −nmotifs 50, −minw 6, −maxw 50, −objfun classic, −markov_order 0.
For subcellular localization, healthy leaves from 2 to 3 week-old plants of A. thaliana were collected for the preparation of protoplasts. The in-frame LOW coding sequence was fused with a green fluorescent protein in the C-terminal region under a constitutive expression promoter POLYUBIQUITIN 10 and was co-transformed into the leaf mesophyll protoplasts with a nucleus marker, ARF19IV-mCherry, by PEG-induced transformation as previously used [48, 49]. The fluorescent signals were observed using a confocal laser scanning microscopy Zeiss7 DUO NLO (Zeiss, Oberkochen, Germany).
qRT-PCR and RNA in situ hybridization
Plant genomic DNA and total RNA were extracted from different tissues as described [46]. For qRT-PCR, 1 μg total RNA from different tissues was reverse transcribed using PrimeScript RT reagent Kit with gDNA Eraser (Takara, Beijing, China) following the manufacturer’s instructions. The PCR assays were performed under the manual of LightCycler 480 Real-Time PCR System (Roche, Shanghai, China). Briefly, the target temperature was set to 58 °C and 45 cycles were used for amplification. All the data were normalized against the expression of constitutively expressed reference gene VrTUB (Vradi05g13910) as reported [50]. The gene expression level was calculated from three biological replicates and three technical replicates. Graphs were produced by GraphPad Prism (GraphPad Software). The primer sequences used in qRT-PCR were listed in the Supporting Information (Additional file 7: Table S1).
For RNA in situ hybridization, flowers at different stages of development were fixed and treated as previously reported [51]. DNA Fragment for producing the sense and antisense probes was cloned and ligated to pTA2 plasmid (TOYOBO, Shanghai, China). Probes were then labeled with digoxigenin-UTP (Roche, Shanghai, China). The non-radioactive in situ hybridization processes were carried out as described [52]. The primer sequences used in RNA in situ hybridization were listed in the Supporting Information (Additional file 2: Table S1).
Arabidopsis transformation and GUS staining
For GUS assay, a 2 kb DNA fragment corresponded to the 5′ promoter and untranslated region of LOW was fused to a GUS gene on pCXGUS-P vector as described [53]. For functional analysis, the full-length coding sequences of LOW was cloned and inserted into pFGC-RCS vector driven by the native 2 kb LOW promoter as described [53]. The plasmids were transformed into the EHA105 Agrobacterium tumefaciens strains and plant transformation was performed under the instruction of floral dipping method as described [54]. The seeds of transgenic plants were selected on Murashige and Skoog (MS) culture media containing proper antibiotics. Histochemical GUS staining assay was performed as described [55]. The stained tissues were examined, dissected and photographed under a stereomicroscope.
Availability of data and materials
The datasets supporting the conclusions of this article are included within the article.
Abbreviations
- BLAST:
-
Basic Local Alignment Search Tool
- CYC:
-
CYCLOIDEA
- GS:
-
Glutamine synthetase
- GUS:
-
β-glucuronidase
- LOW:
-
LOVE ON WINGS
- MCOGs:
-
Major Cluster of Orthologous Groups
- qRT-PCR:
-
Quantitative Reverse Transcription Polymerase Chain Reaction
- RNA-seq:
-
RNA-sequencing
- WT:
-
Wild-type
References
Fenster CB, Armbruster WS, Wilson P, Dudash MR, Thomson JD. Pollination syndromes and floral specialization. Annu Rev Ecol Evol S. 2004;35:375–403.
Aronne G, Giovanetti M, De Micco V. Morphofunctional traits and pollination mechanisms of Coronilla emerus L. flowers (Fabaceae). Sci World J. 2012;2012:1–8.
Toon A, Cook LG, Crisp MD. Evolutionary consequences of shifts to bird-pollination in the Australian pea-flowered legumes (Mirbelieae and Bossiaeeae). BMC Evol Biol. 2014;14(1):43.
Lopez J, Rodriguez-Riano T, Ortega-Olivencia A, Devesa JA, Ruiz T. Pollination mechanisms and pollen-ovule ratios in some Genisteae (Fabaceae) from southwestern Europe. Plant Syst Evol. 1999;216(1–2):23–47.
Westerkamp C. Keel blossoms: bee flowers with adaptations against bees. Flora. 1997;192(2):125–32.
Feng X, Zhao Z, Tian Z, Xu S, Luo Y, Cai Z, et al. Control of petal shape and floral zygomorphy in Lotus japonicus. Proc Natl Acad Sci U S A. 2006;103(13):4970–5.
Wang Z, Luo Y, Li X, Wang L, Xu S, Yang J, et al. Genetic control of floral zygomorphy in pea (Pisum sativum L.). Proc Natl Acad Sci U S A. 2008;105(30):10414–9.
Jiao K, Li X, Guo W, Su S, Luo D. High-throughput RNA-Seq data analysis of the Single Nucleotide Polymorphisms (SNPs) and Zygomorphic Flower Development in Pea (Pisum sativum L.). Int J Mol Sci. 2017;18(12):2710.
Dengler N, Kang J. Vascular patterning and leaf shape. Curr Opin Plant Biol. 2001;4(1):50–6.
Nelson T, Dengler N. Leaf vascular pattern formation. Plant Cell. 1997;9(7):1121–35.
Alonso-Peral MM. The HVE/CAND1 gene is required for the early patterning of leaf venation in Arabidopsis. Development. 2006;133(19):3755–66.
Franks RG, Liu Z, Fischer RL. SEUSS and LEUNIG regulate cell proliferation, vascular development and organ polarity in Arabidopsis petals. Planta. 2006;224(4):801–11.
Ruonala R, Ko D, Helariutta Y. Genetic networks in plant vascular development. Annu Rev Genet. 2017;51:335–59.
De Rybel B, Adibi M, Breda AS, Wendrich JR, Smit ME, Novak O, et al. Integration of growth and patterning during vascular tissue formation in Arabidopsis. Science. 2014;345(6197):1255215.
Mahonen AP, Bishopp A, Higuchi M, Nieminen KM, Kinoshita K, Tormakangas K, et al. Cytokinin signaling and its inhibitor AHP6 regulate cell fate during vascular development. Science. 2006;311(5757):94–8.
Galweiler L, Guan C, Muller A, Wisman E, Mendgen K, Yephremov A, et al. Regulation of polar auxin transport by AtPIN1 in Arabidopsis vascular tissue. Science. 1998;282(5397):2226–30.
Noguero M, Atif RM, Ochatt S, Thompson RD. The role of the DNA-binding one zinc finger (DOF) transcription factor family in plants. Plant Sci. 2013;209:32–45.
Yanagisawa S. The Dof family of plant transcription factors. Trends Plant Sci. 2002;7(12):555–60.
Umemura Y, Ishiduka T, Yamamoto R, Esaka M. The Dof domain, a zinc finger DNA-binding domain conserved only in higher plants, truly functions as a Cys2/Cys2 Zn finger domain. Plant J. 2004;37(5):741–9.
Moreno-Risueno MA, Martinez M, Vicente-Carbajosa J, Carbonero P. The family of DOF transcription factors: from green unicellular algae to vascular plants. Mol Gen Genomics. 2007;277(4):379–90.
Le Hir R, Bellini C. The plant-specific dof transcription factors family: new players involved in vascular system development and functioning in Arabidopsis. Front Plant Sci. 2013;4:164.
Gardiner J, Sherr I, Scarpella E. Expression of DOF genes identifies early stages of vascular development in Arabidopsis leaves. Int J Dev Biol. 2010;54(8–9):1389–96.
Konishi M, Yanagisawa S. Sequential activation of two Dof transcription factor gene promoters during vascular development in Arabidopsis thaliana. Plant Physiol Bioch. 2007;45(8):623–9.
Shigyo M, Tabei N, Yoneyama T, Yanagisawa S. Evolutionary processes during the formation of the plant-specific Dof transcription factor family. Plant Cell Physiol. 2007;48(1):179–85.
Konishi M, Donner TJ, Scarpella E, Yanagisawa S. MONOPTEROS directly activates the auxin-inducible promoter of the Dof5.8 transcription factor gene in Arabidopsis thaliana leaf provascular cells. J Exp Bot. 2015;66(1):283–91.
Skirycz A, Radziejwoski A, Busch W, Hannah MA, Czeszejko J, Kwaśniewski M, et al. The DOF transcription factor OBP1 is involved in cell cycle regulation in Arabidopsis thaliana. Plant J. 2008;56(5):779–92.
Guo Y, Qin G, Gu H, Qu LJ. Dof5.6/HCA2, a Dof transcription factor gene, regulates Interfascicular cambium formation and vascular tissue development in Arabidopsis. Plant Cell. 2009;21(11):3518–34.
Miyashima S, Roszak P, Sevilem I, Toyokura K, Blob B, Heo JO, et al. Mobile PEAR transcription factors integrate positional cues to prime cambial growth. Nature. 2019;565(7740):490–4.
Hedström I, Thulin M. Pollination by a hugging mechanism in Vigna vexillata (Leguminosae-Papilionoideae). PI Syst Evol. 1986;154(3/4):275–83.
Lijavetzky D, Carbonero P, Vicente-Carbajosa J. Genome-wide comparative phylogenetic analysis of the rice and Arabidopsis Dof gene families. BMC Evol Biol. 2003;3:17.
Perez-Rodriguez M, Jaffe FW, Butelli E, Glover BJ, Martin C. Development of three different cell types is associated with the activity of a specific MYB transcription factor in the ventral petal of Antirrhinum majus flowers. Development. 2005;132(2):359–70.
Xiao W, Su S, Higashiyama T, Luo D. A homolog of the ALOG family controls corolla tube differentiation in Torenia fournieri. Development. 2019;146(16):dev177410.
Luo D, Carpenter R, Copsey L, Vincent C, Clark J, Coen E. Control of organ asymmetry in flowers of antirrhinum. Cell. 1999;99(4):367–76.
Wang J, Wang Y, Luo D. LjCYC genes constitute floral dorsoventral asymmetry in Lotus japonicus. J Integr Plant Biol. 2010;52(11):959–70.
Li X, Liu W, Zhuang L, Zhu Y, Wang F, Chen T, Luo D. BIGGER ORGANS and ELEPHANT EAR-LIKE LEAF1 control organ size and floral organ internal asymmetry in pea. J Exp Bot. 2018;70(1):179-91.
Li X, Zhuang LL, Ambrose M, Rameau C, Hu XH, Yang J, et al. Genetic analysis of ele mutants and comparative mapping of ele1 locus in the control of organ internal asymmetry in garden pea. J Integr Plant Biol. 2010;52(6):528–35.
Skirycz A, Jozefczuk S, Stobiecki M, Muth D, Zanor MI, Witt I, et al. Transcription factor AtDOF4;2 affects phenylpropanoid metabolism in Arabidopsis thaliana. New Phytol. 2007;175(3):425–38.
Rueda-Lopez M, Crespillo R, Canovas FM, Avila C. Differential regulation of two glutamine synthetase genes by a single Dof transcription factor. Plant J. 2008;56(1):73–85.
Santopolo S, Boccaccini A, Lorrai R, Ruta V, Capauto D, Minutello E, et al. DOF affecting germination 2 is a positive regulator of light-mediated seed germination and is repressed by DOF affecting germination 1. BMC Plant Biol. 2015;15:72.
Sugiyama T, Ishida T, Tabei N, Shigyo M, Konishi M, Yoneyama T, et al. Involvement of PpDof1 transcriptional repressor in the nutrient condition-dependent growth control of protonemal filaments in Physcomitrella patens. J Exp Bot. 2012;63(8):3185–97.
Feng BH, Han YC, Xiao YY, Kuang JF, Fan ZQ, Chen JY, et al. The banana fruit Dof transcription factor MaDof23 acts as a repressor and interacts with MaERF9 in regulating ripening-related genes. J Exp Bot. 2016;67(8):2263–75.
Jiao K, Li X, Su S, Guo W, Guo Y, Guan Y, et al. Genetic control of compound leaf development in the mungbean (Vigna radiata L.). Hortic Res. 2019;6:23.
Hill JT, Demarest BL, Bisgrove BW, Gorsi B, Su YC, Yost HJ. MMAPPR: mutation mapping analysis pipeline for pooled RNA-seq. Genome Res. 2013;23(4):687–97.
Kang YJ, Kim SK, Kim MY, Lestari P, Kim KH, Ha BK, et al. Genome sequence of mungbean and insights into evolution within Vigna species. Nat Commun. 2014;5:5443.
Jiao K, Li X, Guo W, Yuan X, Cui X, Chen X. Genome re-sequencing of two accessions and fine mapping the locus of lobed leaflet margins in mungbean. Mol Breeding. 2016;36(9):128.
Su S, Xiao W, Guo W, Yao X, Xiao J, Ye Z, et al. The CYCLOIDEA-RADIALIS module regulates petal shape and pigmentation, leading to bilateral corolla symmetry in Torenia fournieri (Linderniaceae). New Phytol. 2017;215(4):1582–93.
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol. 2013;30(12):2725–9.
Zhai C, Zhang Y, Yao N, Lin F, Liu Z, Dong Z, et al. Function and interaction of the coupled genes responsible for Pik-h encoded rice blast resistance. PLoS One. 2014;9(6):e98067.
Yoo SD, Cho YH, Sheen J. Arabidopsis mesophyll protoplasts: a versatile cell system for transient gene expression analysis. Nat Protoc. 2007;2(7):1565–72.
Sairam RK, Dharmar K, Chinnusamy V, Meena RC. Waterlogging-induced increase in sugar mobilization, fermentation, and related gene expression in the roots of mung bean (Vigna radiata). J Plant Physiol. 2009;166(6):602–16.
Su S, Shao X, Zhu C, Xu J, Lu H, Tang Y, et al. Transcriptome-wide analysis reveals the origin of Peloria in Chinese cymbidium (Cymbidium sinense). Plant Cell Physiol. 2018;59(10):2064–74.
Coen ES, Romero JM, Doyle S, Elliott R, Murphy G, Carpenter R. Floricaula: a homeotic gene required for flower development in antirrhinum majus. Cell. 1990;63(6):1311–22.
Qi H, Xia FN, Xie LJ, Yu LJ, Chen QF, Zhuang XH, et al. TRAF family proteins regulate autophagy dynamics by modulating autophagy protein6 stability in Arabidopsis. Plant Cell. 2017;29(4):890–911.
Clough SJ, Bent AF. Floral dip: a simplified method for agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998;16(6):735–43.
Konishi M, Sugiyama M. A novel plant-specific family gene, root primordium defective 1, is required for the maintenance of active cell proliferation. Plant Physiol. 2006;140(2):591–602.
Acknowledgements
We thank Miss Maria João Ferreira (University of Porto, Portugal) for her constructive suggestions during manuscript revision. We are also grateful to the anonymous reviewers for their encouraging and valuable comments.
Funding
This work was supported by the National Natural Science Foundation of China (Grant No. 31700186) and the Chang Hungta Science Foundation of Sun Yat-sen University. S.S. is supported by an overseas postdoctoral fellowship from Japan Society for the Promotion of Science. The funding agencies had no role in the experimental design, data collection and analysis or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
WG, DL and SS designed the research; WG, DL, KJ, XZ, QP and SS performed the research; WG, KJ and SS analyzed the data; WG and SS wrote the paper; WG, DL and SS revised the manuscript. All authors read and approved the manuscript
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Additional file 1: Figure S1.
Left keel-wing complex in Vigna radiata functions as a landing platform for the bee pollinators. (A) Diagram of a V. radiata flower, the left keel and left wing are marked in pink and blue, respectively. (B) Front view of a bee visitation. (C) Side view of a bee visitation.
Additional file 2: Figure S2.
Alignment of Vr07g10060/LOC106767037 coding sequences in wild type and mutant. Box indicates mutated region in the mutant and the blue line marks the Dof domain.
Additional file 3: Figure S3.
Subcellular localization of LOW-GFP fused protein. Signals from GFP, mCherry, chloroplast and merged channels are shown; nuclear marker ARF19IV-mCherry plasmid was co-transformed with LOW-GFP construct; Bars = 10 μm.
Additional file 4: Figure S4.
Dof domain sequence alignment of the 31 mung bean proteins. Identical and similar (> 80%) amino acids are highlighted in black and grey, respectively.
Additional file 5: Figure S5.
Quantitative RT-PCR analysis of LOW in different tissues, error bars of gene expression are ±1 SD from three replicates.
Additional file 6: Figure S6.
GUS-stained inflorescence and floral organs of transgenic Arabidopsis thaliana lines of LOWp:GUS. Bar = 2 mm.
Additional file 7: Table S1.
Primers used in this study.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Guo, W., Zhang, X., Peng, Q. et al. LOVE ON WINGS, a Dof family protein regulates floral vasculature in Vigna radiata. BMC Plant Biol 19, 495 (2019). https://doi.org/10.1186/s12870-019-2099-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-019-2099-x