
Abstract
Background
The recently identified Puccinia graminis f. sp. tritici (Pgt) race TTKSK (Ug99) poses a severe threat to global wheat production because of its broad virulence on several widely deployed resistance genes. Additional virulences have been detected in the Ug99 group of races, and the spread of this race group has been documented across wheat growing regions in Africa, the Middle East (Yemen), and West Asia (Iran). Other broadly virulent Pgt races, such as TRTTF and TKTTF, present further difficulties in maintaining abundant genetic resistance for their effective use in wheat breeding against this destructive fungal disease of wheat. In an effort to identify loci conferring resistance to these races, a genome-wide association study was carried out on a panel of 250 spring wheat breeding lines from the International Maize and Wheat Improvement Center (CIMMYT), six wheat breeding programs in the United States and three wheat breeding programs in Canada.
Results
The lines included in this study were grouped into two major clusters, based on the results of principal component analysis using 23,976 SNP markers. Upon screening for adult plant resistance (APR) to Ug99 during 2013 and 2014 in artificial stem rust screening nurseries at Njoro, Kenya and at Debre Zeit, Ethiopia, several wheat lines were found to exhibit APR. The lines were also screened for resistance at the seedling stage against races TTKSK, TRTTF, and TKTTF at USDA-ARS Cereal Disease Laboratory in St. Paul, Minnesota; and only 9 of the 250 lines displayed seedling resistance to all the races. Using a mixed linear model, 27 SNP markers associated with APR against Ug99 were detected, including markers linked with the known APR gene Sr2. Using the same model, 23, 86, and 111 SNP markers associated with seedling resistance against races TTKSK, TRTTF, and TKTTF were identified, respectively. These included markers linked to the genes Sr8a and Sr11 providing seedling resistance to races TRTTF and TKTTF, respectively. We also identified putatively novel Sr resistance genes on chromosomes 3B, 4D, 5A, 5B, 6A, 7A, and 7B.
Conclusion
Our results demonstrate that the North American wheat breeding lines have several resistance loci that provide APR and seedling resistance to highly virulent Pgt races. Using the resistant lines and the SNP markers identified in this study, marker-assisted resistance breeding can assist in development of varieties with elevated levels of resistance to virulent stem rust races including TTKSK.
Similar content being viewed by others

Background
Stem rust of wheat, caused by the fungal pathogen Puccinia graminis Pers. f. sp. tritici (Pgt), is considered potentially the most damaging disease of wheat. Historical crop losses caused by stem rust epidemics have been recorded in all wheat growing regions of the world [1, 2]. This disease has been primarily controlled via genetic resistance with resistance genes discovered in bread wheat and its relative species [3–5]. However, the emergence of the highly virulent stem rust race TTKSK (also known as Ug99) and its variants has rendered many of the resistance genes ineffective to the pathogen [5, 6], and threatens global wheat production and supply.
The race TTKSK [7], first observed in Uganda in 1998 was found to be virulent to Sr31 [8], a widely deployed and important stem rust resistance gene. Within a few years, virulence to three other resistance genes was documented in the Ug99 lineage: Sr24 was defeated by race TTKST [7], Sr36 was defeated by race TTTSK [7, 9], and Sr9h was defeated by race TTKSF+ [10, 11]. The urediniospores of the stem rust fungus can travel long distances with the flow of wind [1, 12]. Consequently, Ug99 race groups have traveled from hotspot areas in East Africa to South Africa and Iran in the North [13, 14]. The Ug99 race group is projected to spread further to other wheat growing areas in the world [5, 14, 15].
Realizing the threat posed by the Ug99 race group, over 200,000 wheat lines including accessions from germplasm collections to breeding materials from wheat breeding programs throughout the world were screened for resistance to Ug99 in Kenya and Ethiopia [5]. The results showed that 85-95 % of wheat lines grown globally are susceptible to Ug99. The results obtained from screening global germplasm highlights the risk looming over worldwide wheat production due to the susceptibility of current varieties. It is therefore essential that resistance genes are identified and used in breeding programs in these areas, including North America, to prepare for the possible arrival of the Ug99 race group and other highly virulent races. One of such highly virulent races is the newly identified race TKTTF, which has a different genetic lineage from the Ug99 race group [16]. This race caused yield losses close to 100 % on the most widely grown wheat cultivar, ’Digalu’, in southeastern regions of Ethiopia in 2013–2014 [16]. Hence, there is urgent need to identify and characterize new genes for resistance to such races, and their rapid incorporation in the breeding pipeline to develop varieties with improved level of resistance.
Two main types of resistance strategies are used in wheat breeding against stem rust: 1) all-stage resistance (ASR) or seedling resistance, and 2) adult plant resistance (APR). ASR is usually characterized by a hypersensitive reaction upon fungal attack, and usually confers a high level of resistance that is effective in all stages of plant development [17]. On the contrary, APR is generally expressed during the adult growth stages of the plant, usually beginning at booting stage. The major drawback of ASR is the high likelihood of the gene being defeated by new pathogen races when resistance genes are deployed singly, in so called “boom and bust cycles” [18]. While APR genes are considered more durable to wheat rusts than ASR genes, they may not provide adequate levels of resistance to high disease pressure [19]. Therefore, a gene-pyramiding strategy that utilizes a few seedling genes or 4–5 APR genes or genes of both resistance types would be highly desirable. Discovery and development of reliable markers for effective marker-assisted gene introgression and selection is vital to routinely combine resistance genes. Such genes and linked markers can be identified in primary, secondary, and tertiary gene pools of wheat and its related species. However, to minimize linkage drag during resistance gene introgression from wild relatives and increase breeding efficiency, discovery of resistant material in existing breeding programs is preferable.
Association mapping (AM), or linkage disequilibrium (LD) mapping, is a powerful technique used to identify marker-trait associations, and has been used successfully in several crop species such as wheat, barley, soybean, and maize [20–22]. The AM strategy exploits historical recombination events existing in the lines being studied, which can range from natural collections to breeding populations. Since existing populations can be used for mapping loci associated with traits of interest, as opposed to specifically designed populations, this approach is widely used to study the genetic makeup controlling trait variation in wider germplasm pools [22, 23]. As allelic variation and marker polymorphisms are observed at a higher frequency in a genome-wide association study (GWAS) panel compared to a biparental population [24, 25], useful and novel alleles associated with traits of interest may be identified when pairing with high-throughput marker technologies. One drawback of AM is that the underlying population stratification due to breeding history, selection, genetic drift, or founder effects can lead to false associations [26, 27]. This issue, however, can be reduced by accounting for population structure using the relationship matrix or distance matrix among the lines [28].
The objective of this study was to evaluate spring wheat lines from nine breeding programs in the United States and Canada, in addition to CIMMYT lines, for their field-based resistance to the Ug99 race group and conduct a GWAS to identify loci associated with resistance. We also made an attempt to distinguish between loci associated with APR and seedling resistance to exotic virulent Pgt races TTKSK, TRTTF, and TKTTF.
Methods
Plant material
Two-hundred and fifty spring wheat lines from wheat breeding programs in North America were assembled as part of the Triticeae Coordinated Agricultural Project (www.triticeaecap.org). Elite lines were representative of the following wheat breeding programs in the United States: Montana State University (MSU), South Dakota State University (SDSU), University of California-Davis (UCD), University of Idaho (UI), University of Minnesota (UMN), and Washington State University (WSU); in Canada: Agriculture and Agri-Food Canada (Ag-Canada) Manitoba, Ag-Canada Saskatchewan, and Ag-Canada Alberta; and in Mexico: the International Maize and Wheat Improvement Center (CIMMYT). This germplasm panel was previously used to assess root morphology traits [29].
Field stem rust evaluation
A panel of 250 lines was evaluated for field response to stem rust in four disease environments: at Njoro, Kenya during the off-season (January to April 2013), and the main-season (June to October 2013), and at Debre Zeit, Ethiopia during the off-season (January to June 2013, and 2014). These environments are referred in the text as KenOff13, KenMain13, EthOff13, and EthOff14, respectively. In all environments, phenotypic data was collected from plant heading stage up to grain maturing stage (i.e. between Zadoks 50 and 90) [30], when the susceptible checks reached maximum severity (usually ~80 % severity).
In Njoro, plots were arranged in an augmented design with the lines represented once and the susceptible check line ‘Red Bobs’ planted after every fifty entries. The lines were sown as 70 cm long twin rows, 20 cm apart, flat bed. Spreader rows were sown perpendicular to the twin rows, surrounding the field to initiate disease development and maintain uniform disease pressure in the nursery. The spreader rows comprised of a mixture of lines susceptible to race TTKST (Ug99 + Sr24 virulence): ‘CCK’ (Canadian Cunningham Kennedy), ‘PBW343’, ‘Morocco’ and few susceptible CIMMYT lines. The disease was initiated by inoculating the spreader rows using a bulk inoculum of Pgt urediniospores collected at the Njoro field site. Wheat stem rust differential lines with known stem rust resistance genes indicated that the predominant, if not only, race present in the nursery since 2008 was race TTKST; [31]). The urediniospores were suspended in water and injected into spreader plants at 1 m distance prior to booting (growth stage Z35-Z37; [30]). The spreader plants were then sprayed with urediniospores suspended in light mineral oil Soltrol 170 (Chevron Phillips Chemical Company, The Woodlands, TX).
The nursery in Debre Zeit was set up similar to the Njoro nursery. Lines were planted in 1 m long twin rows, flanked by spreader rows comprised of a mixture of susceptible wheat varieties ‘PBW343’, ‘Morocco’, and ‘Local Red’. The spreader rows were artificially inoculated with a bulk of fresh urediniospores collected from PBW343 (PBW343 has Sr31 and several races in the Ug99 race group are virulent to Sr31) and also collected from local fields. Inoculation was carried out in Debre Zeit as it was in the Njoro nursery.
Disease severity on a 0-100 % modified Cobb scale [32] and infection response [17] were recorded for each line. Severity and infection response notes were recorded 2–3 times during the season; the terminal data, which exhibited better disease segregation among the lines in the panel, was used in the analysis. The infection response to disease was assigned constant values as recommended by Stubbs et al. [33] with the response types ‘resistant – moderately resistant’ and ‘moderately susceptible – susceptible’ coded as 0.3 and 0.9, respectively. The stem rust severity values were multiplied by the infection response values to obtain coefficient of infection values [33], which were used in subsequent analyses.
Seedling stem rust evaluation
Seedling assays were conducted at the United States Department of Agriculture, Agricultural Research Service (USDA-ARS) Cereal Disease Laboratory during the winter months between December and February starting December 2012 through February 2014. The 250 spring wheat lines were evaluated with three virulent races of Pgt race TTKSK (isolate 04KEN156/04, Ug99), race TRTTF (isolate 06YEM34-1), and race TKTTF (isolate 13ETH18-1). Race TRTTF was detected in Yemen and Ethiopia and characterized as broadly virulent to wheat stem rust resistance genes including Sr13 and Sr1RS Amigo [34]. Seedling assays were performed as described previously by Rouse et al. [35] for evaluating wheat germplasm for reaction to Pgt races. Two biological replicates of the seedling assays were performed for each Pgt race. Infection types (ITs) were recorded on a 0 to 4 scale according to Stakman et al. [36]. ITs less than or equal to 2+ are considered low infection types whereas ITs greater than or equal to 3- are considered high infection types [36]. In order to use the Stakman ITs in the GWAS, the 0–4 scale was converted to a 0–9 linear scale as proposed by Zhang et al. [37] (Additional file 1). The average linear scale score across the two replications was used in the AM analyses.
Statistical analysis
Data sets from each field environment were fitted into a mixed model with environment as a fixed effect and wheat lines as a random effect to correct for data distortion due to trial effects. Using this model, best linear unbiased predictors (BLUPs) for each line were predicted from the combined analysis model using SAS 9.1, from which final corrected trait values were obtained. After the mean values were normalized for each environment, trait values for each line in all environments were averaged, and used for genome-wide mapping. Heritability on an entry mean basis was calculated based on the method described by Holland et al. [38].
SNP genotyping and analysis of molecular data
The AM panel was genotyped at the USDA-ARS Cereal Crops Research Unit, Fargo, ND as part of the TCAP project for 90,000 gene-based SNPs using a custom Infinium iSelect bead chip assay following the manufacturer’s instructions (Illumina Inc., Hayward, CA) [39]. Allele calls were performed using the computer program GenomeStudio v2011.1 (Illumina Inc.). As genotyping of these lines was carried out as a part of the TCAP, the genotype data was obtained from The Triticeae Toolbox. During data download, SNP markers with minor allele frequency less than 0.05 and no calls for more than 10 % of the lines were removed from the dataset. Lines that were genotyped at less than 20 % loci were also removed from the dataset. These filtering steps yielded 23,976 SNP markers and 241 lines that were used for genome-wide association analysis.
Principal component analysis (PCA) on the marker data of the lines was performed by using the Genomic Association and Prediction Integrated Tool (GAPIT) R package [40]. For the purpose of displaying the PCA results, the R package ‘princomp’ was used to reconstruct the covariance matrix. Principal component 1 scores were plotted against principal component 2 scores for each line in the mapping panel. In order to confirm the results of PCA analysis, the lines were also clustered by their genetic distances using JMP Pro 11.0.0 (SAS Institute Inc.) with Ward’s hierarchical clustering method [41].
Linkage disequilibrium between the markers was estimated as squared allele-frequency correlations (r2). The package ‘LDheatmap’ in R was used to calculate r2 in each of the A, B, and D genomes of common wheat. The LD decay between the marker pairs for each genome was estimated using the least squares regression function, and is represented by the exponential curve. Map positions of the SNPs were obtained from Wang et al. [39].
Linkage disequilibrium and association analysis
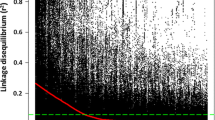
A total of 18,302 mapped SNP markers were used to estimate pairwise LD. LD between markers were calculated for the A, B, and D genomes, and plotted against the genetic distance in centimorgans (cM). The extent of LD between marker pairs was visualized by fitting locally weighted polynomial regression (LOESS) curves into the scatter plot.
Genome-wide association analysis investigating the marker-trait association was performed using the R package GAPIT [40], with the growth stages of genotypes included as covariate for Pgt resistance. The population parameters previously determined (P3D) model [42] was used to conduct association analyses with all trait data. Based on the model selection using the Bayesian information content criterion (BIC), a kinship-mixed linear model (K-MLM) approach that accounts for Type I error rate due to relatedness was used for all traits.
Results
Phenotypic data
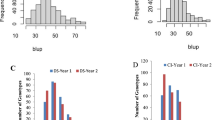
Adequate disease pressure was observed in each environment to discriminate among the entries, as indicated by highly significant (p < 0.01) F-values from ANOVA results of the field data (results not shown). Distribution of rust severities in all environments indicated a quantitative mode of disease distribution with the Njoro main season (KenMain13) recording both higher severity and wider distribution of scores (Fig. 1a). On average however, disease severity was relatively lower in Njoro environments with mean values of 33 % during the 2013 offseason and 35 % during the 2013 main season, compared to Debre Zeit, Ethiopia with 52 % severity during the 2013 offseason and 50 % during the 2014 offseason. Moderate correlations (r ranging from 0.44 to 0.57) between the field data from the four seasons was observed (Table 1). The estimated broad sense heritability across the four environments was 0.72.

Frequency distribution of stem rust severity scores for the North American Spring Wheat Breeding Germplasm (250 lines) during (a) four seasons in East Africa; (b) screening for resistance at the seedling stage with three stem rust races. Seedling infection type (IT) scores have been converted to the linear scale, as discussed in ‘Methods’
The complete panel of 250 lines was inoculated with Pgt races TTKSK, TRTTF, and TKTTF at seedlings to uncover genetic factors contributing resistance to these virulent stem rust races. Replications of the seedling tests were highly correlated with r-value ranging from 0.84-0.99 (p < 0.001). However, the pair-wise correlations among the three races were low and not significant (data not shown). Most of the lines screened for seedling resistance against race TTKSK were susceptible, and only 15 (6 %) showed resistant reactions (IT 22+ or lower) (Fig. 1b). Seedling resistance was more common to races TRTTF, with 44 %, and TKTTF, with 63 % of lines with low infection types. Of the seedling resistant lines, nine lines showed resistance to all three races; three lines showed resistance only to races TTKSK and TRTTF; and five lines showed resistance only to races TTKSK and TKTTF (Additional file 1). The University of Minnesota cultivar ‘Thatcher’ was heterogeneous for resistance to race TTKSK (IT 0; / X in the first replication and IT 0; / 3+ in the second replication) yet susceptible to both TRTTF and TKTTF (IT 33+ to both races in both replications). The average adult plant disease severity of lines with seedling resistance to race TTKSK was lower in all four environments than lines susceptible to this race (t-test p-value of 0.02 at α = 0.05; Additional file 1).
Population structure
To investigate the population structure of the germplasm panel, the genotypes were analyzed for clustering based on principal component (PC) values. The groupings of lines belonging to each breeding program based on PC values are presented in Fig. 2. The first and second PC values explained 8.9 % and 4.3 % genetic variation in the panel, respectively. Although the genetic relationship among representative lines of different breeding programs was not very distinct, a general statement can be made that two main clusters are observed based on genetic differentiation. The cluster on the left of Fig. 1 (hereafter referred to as Cluster 1) comprises lines from breeding programs in the Upper-Midwest of the United States: MSU, SDSU, and UMN; and in Canada. The cluster on the right (hereafter referred to as Cluster 2) comprises lines from the Western United States: UCD, UI, and WSU; and in Mexico: CIMMYT. The CIMMYT lines included in this analysis were selected because of their potential heat and drought stress resistance and may not be representative of the entire CIMMYT breeding germplasm. Despite the noticeable amount of relatedness among breeding programs, these two main clusters may be divided further into six sub-clusters: Cluster 1 into two sub-clusters with lines from MSU clustering separately while lines from Alberta, Manitoba, Saskatchewan, SDSU, and UMN form a single sub-cluster; and Cluster 2 into four sub-clusters with lines from CIMMYT, UCD, UI, and WSU. The population stratification and germplasm sharing among the lines revealed by the PC was also corroborated by the results from hierarchical clustering using Ward’s method in JMP (Additional file 1).

A scatter plot of principal component 1 (PC1) plotted against principal component 2 (PC2). All 241 lines in the AM panel are represented by a symbol, with lines of each breeding program labeled by different symbol and distinct color. Proportion of the variance explained by the principal component values are indicated in parenthesis
Linkage disequilibrium
In the A-genome, LD declined to 50 % of its original value at about 8 cM (Fig. 3a), whereas these values for the B-genome and the D-genome were about 7 cM and 6 cM, respectively (Figs. 3b and c, respectively). These values are similar to the LD values reported by Chao et al. [43] in their detailed LD characterization of wheat varieties having different growth habits from several breeding programs.

A scatter plot showing squared allele-frequency correlations (r2) distribution plotted against genetic distance (cM) for each subgenome of wheat: Figs. 3a, b, and c represent the A, B, and D subgenomes of wheat, respectively. The decline of linkage disequilibrium (LD) is shown by fitting a locally weighed polynomial regression (LOESS) curve into the plot. The inset shows a zoomed-out view of the subgenome whereas the main picture shows a zoomed-in view of the distribution within 50 cM distance
Association analysis
APR mapping
Initial association analysis was conducted on all 241 lines, without removing any lines that showed resistance to race TTKSK during seedling screening of the lines as described later. This approach detected 24 SNP markers on seven different chromosomes (2A, 2B, 3B, 4A, 6A, 6B, and 7A) that were significantly associated (p-value <0.001) with field resistance to Ug99 (Table 2). In addition, five significant markers with unknown map positions were detected. The phenotypic variance explained by these 29 SNP markers ranged from 0.2 % to 4.6 %.
As the germplasm in the panel was known to possess ASR genes that are effective to the Ug99 race group, it was assumed that these ASR genes could lead to the masking of potential APR genes. Therefore, in an attempt to detect loci conferring APR, lines that were resistant to race TTKSK during seedling screening were removed. Lines that were resistant (IT ranging from 0 to 2+) in either of the two replications of seedling screening, and with a complex score with low and high ITs (for example 2 + 3-) were also removed. This filtering step yielded a subset of 219 lines for the APR-specific genome-wide association analysis.
The APR-specific AM approach identified 26 SNP markers providing APR to Ug99 race group (Table 2). Of the 26 significant SNPs, 23 SNPs were distributed across nine chromosomes (1B, 2A, 2B, 2D, 3B, 4A, 5A, 7A, 7B), whereas the remaining three SNPs were unmapped. Six of the 23 mapped SNPs were also detected in the initial analysis on the whole set of 241 lines. The R2 values for the significant SNPs ranged from 0.1 % to 0.6 % (Table 2).
Significant SNP markers detected on the complete panel and by APR-specific mapping were cross-checked with the GWAS results for each of the four environments. While no significant SNPs were detected in the Kenya 2013 offseason environment, three, six, and nine SNPs were confirmed in the Ethiopia offseason 2013, Ethiopia offseason 2014, and Kenya main-season 2014, respectively (Table 2). Two SNPs – IWA3120 (mapped to 1B) and IWB35697 (mapped to 6B) were common between the Ethiopia 2013 and 2014 environments. No SNP markers were common in all four environments.
Mapping of seedling resistance
The genome-wide scan for SNPs linked with seedling resistance to race TTKSK detected 16 significant SNP markers on chromosomes 1D, 3B, 4A, and 5B, and 7 additional significant SNP markers with no mapped locations (Table 3). These SNPs explained 2 to 7 % of variation observed in race TTKSK seedling resistance. The results also revealed that the loci conferring seedling resistance to TTKSK are different than those involved in APR to Ug99 (Table 3, Table 2).
Similarly, GWAS was conducted for resistance to race TRTTF, a virulent race of Yemeni origin. This resulted in detection of 77 significant SNPs on six chromosomes (1B, 1D, 5D, 6A, 6B, 7A), and 9 SNPs that were unmapped (Table 4). Additionally, mapping of resistance was carried out for a new virulent race TKTTF of Ethiopian origin. A genome-wide scan for resistance loci resulted in 109 SNPs distributed on five chromosomes (1A, 4A, 5A, 6B, 7A), and 2 unmapped SNPs (Table 5).
Discussion
Wheat stem rust disease has been primarily controlled by the use of resistant genes discovered in hexaploid wheat and its related species. However, the Ug99 race group has defeated many of the widely deployed resistance genes, and thus poses a threat to wheat production globally. Moreover, several of the previously identified genes discovered in wild progenitors or landraces are not desirable for their use in resistance breeding because of linkage drag [3, 44]. Therefore, discovery of loci contributing resistance to Ug99 and other virulent races in elite breeding germplasm is a clear advantage. The resistance uncovered in this study, composed of elite germplasm from North American breeding programs, can provide a great resource for the fight against Ug99 and stem rust in general. As no SNP markers were significant across all four field environments, differences among the disease environments with regard to races present, temperature, and other environmental factors as well as locus by environment interaction are likely involved in this lack of consistency. Lack of strong correlations among the environments also corroborates this assumption (Table 1).
Comparison of significant APR Loci with published studies
The map locations of significant SNP markers in our study, obtained from Wang et al. [39], were compared to positions of markers and genes/quantitative trait loci (QTL) reported in previous mapping studies conducted to uncover loci associated with stem rust resistance. In this section, we have used from the integrated genetic map consisting of different marker types generated by Maccaferri et al. [45] to obtain the relative distances between previously reported markers and the significant markers in our study.
Five significant SNP markers (IWA3120, IWB21176, IWB31027, IWB56771, IWB59663) were detected at position 90 cM on chromosome 1B, of which all except IWA3120 were also detected in Ethiopia 2013. The marker cfd48 reported by Pozniak et al. [46] in a durum wheat (Triticum durum Desf.) GWAS study is located 4 cM from the SNP markers we detected, and could represent the same locus. Bhavani et al. [47] and Njau et al. [48] both reported the marker wPt-1560 on 1BL to be associated with Ug99 resistance in separate spring wheat RIL populations. This marker, as well as Sr58, an APR gene for stem rust of wheat [49, 50], are located at a distance of >50 cM from these five SNP markers. QTL on chromosome 2A providing APR to Ug99 have also been mainly reported in durum wheat mapping populations. Letta et al. [51] detected gwm1045 to be significantly associated with Ug99 resistance in a durum wheat AM panel; and Haile et al. [52] reported a QTL linked to the marker gwm1198 on 2A that confers resistance to Ug99 in the durum wheat population Kristal/Sebatel. Neither of these markers was in proximity to the SNP markers detected in our study. We detected one significant marker, IWB8481, located at 9 cM on chromosome 2D. The only reported QTL on 2D that provides APR to Ug99 and its derivative races is in the CIMMYT biparental population PBW343/Kiritati [47]. Two Sr genes – Sr32, and Sr46 have been mapped to the short arm of 2D [49, 53], and both provide resistance to Ug99 [44]. It should be noted that Sr32 has also been introgressed to 2A and 2B [54], but is not expected to be present in the 250 lines analyzed in this study. We used the Sr32 markers developed by Mago et al. [53] to screen our panel but found the markers to be not predictive of the gene (Additional file 1). As no reliable marker for Sr46 has been developed, we are unable to distinguish between these two genes and the marker we found on 2D.
The marker IWA4275 detected on chromosome 2B (position 197 cM) in our study is very close (distance of 2.7 cM) to the marker wPt-8460, known to be linked to Sr9h in 1956 Rockefeller Foundation cultivar Gabo 56 (CI 14035) [11]. The same marker was also reported by Yu et al. [55] in their association mapping study constituting of CIMMYT spring wheat germplasm. Sr9h, previously temporarily designated as SrWeb, is derived from the Canadian wheat cultivar ‘Webster’ (RL6201) and confers ASR gene effective to TTKSK [56]. Markers developed by Rouse et al. [11] showed that Sr9h is present in 13 lines (5 %) in our panel (Additional file 1), implying that IWA4275 could represent the Sr9h locus in our panel. The gene Sr9a is also located on 2BL [55, 57], but is ineffective to Ug99 [44].
On 2B, we also detected five SNP markers: IWA8534, IWB23660, IWB25868, IWB69631, and IWB25869 located at 126 cM. Based on the consensus map published by Maccaferri et al. [45], these markers are located at a distance of 4 cM from wmc332, which is linked to Sr28 [58]. We used two markers: wmc332 [58] and a newly developed SNP marker (Michael Pumphrey, personal communication) to investigate if Sr28 was present in our panel. While both markers have only been tested on a limited panel of lines and are not confirmed as diagnostic, we detected that up to 20 lines (8 %) could possess Sr28 (Additional file 1). Thus, our marker-trait association may be detecting Sr28 on this 2B region.
Nine SNP markers were detected on the short arm of chromosome 3B with 8 SNPs located at positional range of 11.5 – 14.1 cM and one additional SNP at 32.2 cM. These 8 SNPs in the range 11.5 – 14.1 cM may be proximal to Sr2, a highly important APR gene for stem rust of wheat [3, 59]. Upon marker screening, it was found that 22 lines (9 %) in the panel contain Sr2 (Additional file 1). This gene is used extensively in the CIMMYT spring wheat breeding program, and is shared by some US breeding programs that also incorporated this gene in their germplasm for broad-spectrum resistance. It is possible that the SNP at 32.2 cM is associated with Ug99 resistance that has been observed near the Sr12 locus [60]. Another stem rust APR gene, Sr57 [61], is located on chromosome 7D. Screening of the panel using the sequence-tagged site marker developed by Lagudah et al. [62] showed that 97 lines (39 %) could contain Sr57. For other two stem rust APR genes: Sr55, located on 4D [63] and Sr56, located on 5B [64], no diagnostic markers are available. As no SNP markers were detected on chromosomes 4D and 5B during the analysis, we believe these genes are not present in our mapping panel.
Several QTL located on chromosome 4A that provide resistance to Ug99 have been reported in association mapping studies [55, 65, 66], and in biparental studies [47] in CIMMYT germplasm. These sources of stem rust resistance are not located in the vicinity of the SNPs IWB46973, IWB56556, and IWB67877 detected also on 4A in our study. Similarly, QTL on chromosome 5A providing resistance against Ug99 have been reported in biparental and association mapping studies [46, 47]. However, chromosome positions of the QTL and significant loci reported in these studies differ from those detected in our study.
We detected only one significant SNP (IWA233) on 6AS. Mapped at 66 cM, this SNP is located away (>100 cM) from the marker gwm617 reported by Pozniak et al. [46], and from the marker Sr26#43 linked to Sr26, which provides resistance to the Ug99 and its derivative races [55, 67]. Marker screening confirmed that Sr26 is absent in the panel under study (Additional file 1). Several QTL effective to Ug99 and its derivative races have also been discovered on chromosome 6B [4, 49]. Of the reported QTL, the DArT marker wPt-6116 in the AM study conducted by Yu et al. [65] is located very close to the significant markers detected in this study: 1.1 cM from IWB24757 and 2.2 cM from IWB45581. The gene Sr11 is located on 6BL, but is ineffective to Ug99 and its derivative races [3, 5]. Likewise, several QTL have been reported on 7A that provide field resistance to Ug99 [46, 47, 51, 52, 68]. However, none of the reported QTL or positions of significant marker effects coincide with the significant markers detected in this study. Two 7B SNP markers – IWB47548 and IWA4175 – were significantly associated with resistance to Ug99. Letta et al. [51] have reported loci associated with resistance to Ug99 in durum wheat germplasm, however they are located at a large distance (>50 cM) from both markers in our study.
The significant SNP markers associated with APR to Ug99 reported in this study provide several resistance loci to fight the disease, of which some are likely novel. Validation of the significant markers in all chromosomes is essential to confirm the identity of the associated resistance loci as well as to test their usefulness in marker assisted resistance breeding in breeding programs.
Comparison of significant seedling-resistance Loci with existing resistance genes
The results of the GWAS for seedling-resistance in this study were compared with previous findings for ASR to stem rust of wheat. As the discovered SNPs are suspected to be linked primarily with existing or putatively novel resistance genes, a search for similarities in chromosomal location with known resistance genes was emphasized.
We detected 23 SNPs in our germplasm panel that were significantly associated with race TTKSK resistance at the seedling stage. The SNP marker IWA642 mapped at 67.7 cM on 1D is relatively close to Sr50, a gene that provides resistance to the Ug99 group of races [28]. The seedling resistance genes SrCad and SrTmp are considered to be present in the panel used in this study. SrCad is a stem rust resistance gene derived from the Canadian wheat lines ‘Peace’ and ‘AC Cadillac’, and is effective to Ug99 and its derivative races [69]. Located on chromosome 6D, this gene confers a highly resistant reaction (IT of 1 to 12) to TTKSK in seedling stages, and is moderately resistant to Ug99 in field nurseries. SrCad has not been shown to be different than Sr42 in either map position or resistance specificity [70]. SrTmp is another gene resistant to TTKSK yet no SNPs were detected on 4B where the SrTmp gene is thought to be located [3]. Additional data suggest that SrTmp may be located on 6DS at a similar location to Sr42/SrCad [71]. We used two markers: SSR marker cfd49 [72] and a SNP marker (Gao et al., unpublished) to screen the panel for presence/absence of Sr42. Results indicated that at least 71 or more lines in the panel could carry this gene, yet the markers did not support each other (Additional file 1). Neither marker results also corroborate our TTKSK seedling screening results. We are not aware of any study carried out on broad germplasm to determine if these two markers are diagnostic or even predictive. From our results, it appears that they are neither diagnostic nor predictive of Sr42/SrCad. Similarly, Sr9h has a resistant reaction to race TTKSK at the seedling stages (1 to 2 infection type) [56]. The presence of Sr9h was confirmed by marker screening, as discussed above. Except for the likely presence of Sr9h, the position of the loci conferring Ug99 resistance in this study suggest that different genes than the ones discussed above could be present in our panel. We suspect that association mapping is limited by the low frequency of resistance loci (only 15 (6 %) of 250 lines resistant to TTKSK), leading to lack of detection of SNPs significantly associated with SrCad or SrTmp. Since no ASR genes effective to Ug99 are known to exist on chromosomes 3B and 5B, our findings indicate that the North American breeding germplasm might contain previously undiscovered important sources of resistance to the disease.
The genes Sr24 and Sr36 are resistant to the race TTKSK, yet no SNPs associated with resistance to this race were detected in the chromosomes containing these genes. Sr24, located on 3DL, is widely used in Mexico and the USA; Sr36, located on 2BS, is known to be present in wheat lines in the USA [3]. Lack of detection of these genes can be attributed to either 1) representative germplasm with these genes are not present in our GWAS panel; or 2) if present, the allele frequency is very low which does not pass our stringent analysis filters. Upon marker screening (Additional file 1), we discovered that Sr36 is not present in our panel; and only 7 lines (3 %) contain Sr24, confirming our assumptions.
Of the 77 mapped SNPs significantly associated with resistance to TRTTF, 57 SNPs were located on the short arm of chromosome 6A (position range 2 cM – 26 cM). These markers are most likely linked to the gene Sr8a which is located on 6AS and is effective to the race TRTTF [34, 73]. Similarly, the SNP marker IWB48466 located on the long arm of 7A (217 cM) is in the same region as the stem rust resistance gene Sr22. This gene was introgressed into 7AL of hexaploid wheat from its diploid relative Triticum boeoticum [74], and is effective against TRTTF [75]. Marker screening of the GWAS panel with a robust sequence tagged site (STS) marker developed by Periyannan et al. [76] confirmed that Sr22 is not present in the panel. Sr31, while ineffective against TTKSK, is effective against TRTTF, and is located on 1BL [75, 77]. Our GWAS results detected 13 significant SNPs, all on the short arm of chromosome 1B (position range 44 cM – 65 cM). Given the presence of CIMMYT lines in our panel and the widespread use of Sr31 in breeding programs, screening of lines with Sr31 with these markers is needed to determine if the markers are linked to Sr31, or if a novel source of resistance to TRTTF is located on 1BS.
Chromosome 1DS is known to harbor multiple Sr genes [49], and could be represented by the two SNPs that were detected on 1DS in our analysis. We also discovered markers on 5DL and 6BS associated with resistance to TRTTF. As no ASR genes effective against TRTTF are known to exist on 5DL and 6BS, the North American elite breeding germplasm likely possesses novel genes for resistance to the Yemeni stem rust race TRTTF.
One-hundred and nine SNPs associated with seedling resistance to the newly detected Ethiopian stem rust race TKTTF were detected on five chromosomes: 1AS, 4AL, 5AL, 6BL, and 7AS. The 52 6BL SNPs distributed in the positional range of 109 cM – 123 cM likely represent the gene Sr11 which is effective to this race. Fifty-one significant SNPs were located on 4AL (142 cM – 164 cM) possibly indicative of resistance gene Sr7a (TKTTF is virulent to Sr7b). No ASR genes are known to be located on chromosome 5AL, and therefore the germplasm under study may possess a new source of resistance to the race TKTTF in this region. APR QTL providing resistance to the Ug99 and its derivative races have been detected in the 1AS region [49, 55]. Additionally, the gene Sr1RS Amigo is located on the 1RS.1AL rye chromosome arm translocation. Chromosome 7AS does not possess any known ASR genes, yet APR QTL effective to Ug99 and its derivative races have been detected in the region [49, 68].
None of the SNPs associated with seedling resistance for the three races were common, suggesting that none of the genes in this material are broadly effective. Further studies involving development of populations for fine mapping and allelism tests are required to elaborate and confirm the nature of the genetic mechanisms controlling the resistance to these three rust races.
Using breeding lines in GWAS
One of the main advantages of conducting association mapping on a panel consisting of breeding germplasm is to explore the genetic composition of the lines, and estimate the effects of significantly associated loci with the trait(s) of interest. The discovery of significant SNPs can allow for tagging of lines that are enriched for alleles associated with the trait, and their use in gene introgression for resistance breeding. More importantly, as the lines used in this AM study are elite, they possess the desired agronomic traits, and are adapted to the desired regions. This helps in avoiding the problems that could otherwise arise from linkage drag, when more diverse germplasm is used to introgress alleles of interest. Singh et al. [5] have reported that up to 95 % of germplasm from global seed collections and breeding programs are susceptible to Ug99. As Ug99 and its derivative races have not yet been observed in North America, it is prudent to prepare for their possible arrival by developing resistant varieties. Discovery of resistant sources in existing breeding programs can speed up the process of gene introgression into elite lines, gene pyramiding for elevated resistance to the disease, and possible identification of diagnostic markers that can be used in marker assisted resistance breeding. Germplasm sharing among the breeding programs for this purpose, at least within the US, is plausible given the genetic similarity among the lines, as observed in Fig. 2. The availability of SNP alleles associated with reduced disease severity (as well as increased severity) in both adult plant and seedling stages (Additional file 1) should be useful for breeders to make decisions about selection of lines to be used as parents in their breeding programs. Breeders may also use the significant SNP markers we have provided to design assays for possible marker assisted selection or screening of resistant materials in their own breeding programs. Additionally, Table 6 has been populated with a list of lines that exhibited high levels of APR to Ug99 (Table 6), and seedling resistance to TTKSK (Table 7). The complete genotypic and phenotypic data presented in this study have also been made available on The Triticeae Toolbox (T3 webportal) with the goal of facilitating line selection based on Sr marker associations. We are confident that the North American wheat breeding programs can fortify the stem rust resistance in their germplasm by capitalizing on the information provided in this study.
Conclusions
In this study, we report the frequency and variability in seedling resistance and APR present in North American spring wheat breeding germplasm to virulent exotic Pgt races. Several loci were found to be significant, which is an indication that despite the relatively narrow goals for germplasm development, enough genetic variation lies within the current North American breeding germplasm that can be utilized to breed for resistance against the virulent stem rust races, including the Ug99 race group. While only a small portion (6 %) of the germplasm showed seedling resistance, APR to Ug99 revealed several likely-novel genomic regions associated with resistance to Ug99. The lines that performed well at either or both growth stages (seedling and adult) could be used immediately to make crosses with elite lines to generate lines with improved rust resistance. Specific crosses could also be made to create mapping populations to fine map the regions of interest in an effort to identify diagnostic markers linked with the resistance loci. The discovery of such diagnostic markers will add great value to recurrent selection breeding programs as well as in identification of lines that carry the resistance loci. As such, further characterization and validation of the detected loci is necessary for effective utilization of these results. The availability of marker and trait data on the T3 webportal that were generated for this GWAS panel should enable interested groups to pursue these studies.
Availability of supporting data
The genotypic data generated on this GWAS panel and used in this article are available under the genotyping experiment ‘TCAP90K_SpringAM_panel’ in The Triticeae Toolbox repository, https://triticeaetoolbox.org/wheat/display_genotype.php?trial_code=TCAP90K_SpringAM_panel ). The phenotypic data collected on this GWAS panel and used in this article are available under the experiment set ‘USSpring_GWAS’ in The Triticeae Toolbox repository, https://triticeaetoolbox.org/wheat/view.php?table=experiment_set&uid=48 ). The data sets supporting the results of this article are included within the article and its additional files.
Abbreviations
- Pgt :
-
Puccinia graminis f. sp. tritici
- ASR:
-
All-stage resistance
- APR:
-
Adult plant resistance
- QTL:
-
Quantitative trait locus
- AM:
-
Association mapping
- LD:
-
Linkage disequilibrium
- GWAS:
-
Genome-wide association study
- PCA:
-
Principal component analysis.
References
Roelfs AP. Epidemiology in North America. In 'The Cereal Rusts'. Vol. II. Orlando, Florida, USA: Academic; 1985.
McIntosh RA, Brown GN. Anticipatory breeding for resistance to rust diseases in wheat. Annu Rev Phytopathol. 1997;35(1):311–26.
McIntosh RA, Wellings CR, Park RF. Wheat rusts: an atlas of resistance genes. Victoria, Australia: CSIRO Publications; 1995.
Haile JK, Röder MS. Status of genetic research for resistance to Ug99 race of Puccinia graminis f. sp. tritici: A review of current research and implications. Afr J Agr Res. 2013;8:6670–80.
Mago R, Brown-Guedira G, Dreisigacker S, Breen J, Jin Y, Singh R, et al. An accurate DNA marker assay for stem rust resistance gene Sr2 in wheat. Theor Appl Genet. 2011;122(4):735–44.
Pretorius ZA, Singh RP, Wagoire WW, Payne TS. Detection of virulence to wheat stem rust resistance gene Sr31 in Puccinia graminis f. sp. tritici in Uganda. Plant Dis. 2000;84:203.
Jin Y, Szabo LJ, Pretorius ZA, Singh RP, Ward R, Fetch T. Detection of virulence to resistance gene Sr24 within race TTKS of Puccinia graminis f. sp. tritici. Plant Dis. 2008;92(6):923–6.
Pretorius ZA, Singh RP, Wagoire WW, Payne TS. Detection of virulence to wheat stem rust resistance gene Sr31 in Puccinia graminis f. sp. tritici in Uganda. Plant Dis. 2000;84:203.
Jin YS, Rouse MN, Pretorius ZA, Wanyera R, Njau P. Detection of virulence to resistance gene Sr36 within the TTKS race lineage of Puccinia graminis f. sp. tritici. Plant Dis. 2009;93:367–70.
Pretorius ZA, Szabo LJ, Boshoff WHP, Herselman L, Visser B. First Report of a new TTKSF race of wheat stem rust (Puccinia graminis f. sp. tritici) in South Africa and Zimbabwe. Plant Dis. 2012;96(4):590.
Rouse MN, Nirmala J, Jin Y, Chao S, Fetch Jr TG, Pretorius ZA, et al. Characterization of Sr9h, a wheat stem rust resistance allele effective to Ug99. Theor Appl Genet. 2014;127:1681–8.
Hirst JM, Hurst GW. Long-distance spore transport. In "Airborne Microbes". London and New York: Cambridge University Press; 1967.
Singh RP, Hodson DP, Huerta-Espino J, Jin Y, Njau P, Wanyera R, et al. Will stem rust destroy the world's wheat crop? Adv Agron. 2008;98:272–309.
Hodson DP, Nazari K, Park RF, Hansen J, Lassen P, Arista J, et al. Putting Ug99 on the map: an update on current and future monitoring. St. Paul, MN, USA: BGRI 2011 Technical Workshop; 2011. p. 3–13.
Park R, Fetch T, Hodson D, Jin Y, Nazari K, Prashar M, et al. International surveillance of wheat rust pathogens: Progress and challenges. Euphytica. 2011;179:109–17.
Olivera Firpo P, Newcomb M, Szabo L, Rouse MN, Johnson JL, Gale SW, et al.: Phenotypic and genotypic characterization of race TKTTF of Puccinia graminis f. sp. tritici that caused a wheat stem rust epidemic in southern Ethiopia in 2013/14. Phytopathology. 2015;105(7):917-28.
Roelfs AP, Singh RP, Saari EE. Rust diseases of wheat: Concepts and methods of disease management. CIMMYT: Mexico; 1992.
Parlevliet J. Durability of resistance against fungal, bacterial and viral pathogens; present situation. Euphytica. 2002;124(2):147–56.
Singh RP, Hodson DP, Jin Y, Huerta-Espino J, Kinyua MG, Wanyera R, et al. Current status, likely migration and strategies to mitigate the threat to wheat production from race Ug99 (TTKS) of stem rust pathogen. CAB Reviews: Perspectives in Agriculture, Veterinary Science, Nutrition and Natural Resources. 2006;1:1-13.
Rostoks N, Ramsay L, MacKenzie K, Cardle L, Bhat PR, Roose ML, et al. Recent history of artificial outcrossing facilitates whole-genome association mapping in elite inbred crop varieties. Proc Natl Acad Sci. 2006;103(49):18656–61.
Ersoz E, Yu J, Buckler E. Applications of linkage disequilibrium and association mapping in crop plants. In: Varshney R, Tuberosa R, editors. Genomics-Assisted Crop Improvement. Netherlands: Springer; 2007. p. 97–119.
Zhu C, Gore M, Buckler ES, Yu J. Status and prospects of association mapping in plants. Plant Genome. 2008;1(1):5–20.
Abdurakhmonov IY, Abdukarimov A: Application of association mapping to understanding the genetic diversity of plant germplasm resources. Int J Plant Genomics. 2008;2008:574927.
Jannink J-L, Bink MCAM, Jansen RC. Using complex plant pedigrees to map valuable genes. Trends Plant Sci. 2001;6(8):337–42.
Buckler ES, Thornsberry JM. Plant molecular diversity and applications to genomics. Curr Opin Plant Biol. 2002;5(2):107–11.
Lander ES, Schork NJ. Genetic dissection of complex traits. Science. 1994;265(5181):2037–48.
Slatkin M. Inbreeding coefficients and coalescence times. Genet Res. 1991;58(2):167–75.
Pritchard JK, Rosenberg NA. Use of unlinked genetic markers to detect population stratification in association studies. Am J Hum Genet. 1999;65(1):220–8.
Narayanan S, Vara Prasad PV. Characterization of a spring wheat association mapping panel for root traits. Agron J. 2014;106:1593–604.
Zadoks JC, Chang TT, Konzak CF. A decimal code for the growth stages of cereals. Weed Res. 1974;14(6):415–21.
Njau PN, Jin Y, Huerta-Espino J, Keller B, Singh RP. Identification and evaluation of sources of resistance to stem rust race Ug99 in wheat. Plant Dis. 2010;94(4):413–9.
Peterson RF, Campbell AB, Hannah AE. A diagramatic scale for estimating rust intensity of leaves and stem of cereals. Can J Res. 1948;26c(5):496–500.
Stubbs RW, Prescott JM, Saari EE, Dubin HJ. Cereal disease methodology manual. Mexico: CIMMYT; 1986.
Olivera PD, Jin Y, Rouse M, Badebo A, Fetch Jr T, Singh RP, et al. Races of Puccinia graminis f. sp. tritici with com- bined virulence to Sr13 and Sr9e in a field stem rust screening nursery in Ethiopia. Plant Dis. 2012;96:623–8.
Rouse MN, Wanyera R, Njau P, Jin Y. Sources of resistance to stem rust race Ug99 in spring wheat germplasm. Plant Dis. 2011;95:762–6.
Stakman EC, Stewart DM, Loegering WQ. Identification of physiologic races of Puccinia graminis var. tritici. United States Department of Agriculture, Agricultural Research Service. 1962; E-617.
Zhang D, Bowden R, Bai G, BGRI. A method to linearize Stakman infection type ratings for statistical analysis. In: BGRI 2011 Technical Workshop. St Paul, MN, USA: Borlaug Global Rust Initiative; 2011. p. 28.
Holland JB, Nyquist WE, Cervantes-Martínez CT. Estimating and interpreting heritability for plant breeding: an update. In: Plant Breeding Reviews. New Jersey, USA: John Wiley & Sons, Inc.; 2010: 9–112.
Wang S, Wong D, Forrest K, Allen A, Chao S, Huang BE, et al. Characterization of polyploid wheat genomic diversity using a high-density 90 000 single nucleotide polymorphism array. Plant Biotechnol J. 2014;12:787–96.
Lipka AE, Tian F, Wang Q, Peiffer J, Li M, Bradbury PJ, et al. GAPIT: Genome Association and Prediction Integrated Tool. Bioinformatics 2012;28:2397–99.
Ward JH. Hierarchical grouping to optimize an objective function. J Am Stat Assoc. 1963;58(301):236–44.
Zhang Z, Ersoz E, Lai C-Q, Todhunter RJ, Tiwari HK, Gore MA, et al. Mixed linear model approach adapted for genome-wide association studies. Nat Genet. 2010;42(4):355–60.
Chao S, Dubcovsky J, Dvorak J, Luo M-C, Baenziger S, Matnyazov R, et al. Population- and genome-specific patterns of linkage disequilibrium and SNP variation in spring and winter wheat (Triticum aestivum L.). BMC Genomics. 2010;11(1):727.
Singh RP, Hodson DP, Huerta-Espino J, Jin Y, Bhavani S, Njau P, et al. The emergence of Ug99 races of the stem rust fungus is a threat to world wheat production. Annu Rev Phytopathol. 2011;49(1):465–81.
Maccaferri M, Zhang J, Bulli P, Abate Z, Chao S, Cantu D, et al. A genome-wide association study of resistance to stripe rust (Puccinia striiformis f. sp. tritici) in a worldwide collection of hexaploid spring wheat (Triticum aestivum L.). G3: Genes|Genomes|Genetics. 2015;5(3):449–65.
Pozniak CJ, Reimer S, Fetch T, Clarke JM, Clarke FR, Somers DJ, et al. Association mapping of Ug99 resistance in diverse durum wheat population. Brisbane, Australia: Proceedings of the 11th International Wheat Genetics Symposium; 2008. p. 809–11.
Bhavani S, Singh RP, Argillier O, Huerta-Espino J, Singh S, Njau P, et al. Mapping durable adult plant stem rust resistance to the race Ug99 group in six CIMMYT wheats. In: BGRI 2011 Technical Workshop. St Paul, MN, USA: Borlaug Global Rust Initiative; 2011. p. 43–53.
Njau PN, Bhavani S, Huerta-Espino J, Keller B, Singh RP. Identification of QTL associated with durable adult plant resistance to stem rust race Ug99 in wheat cultivar ‘Pavon 76’. Euphytica. 2013;190(1):33–44.
Yu L-X, Barbier H, Rouse M, Singh S, Singh R, Bhavani S, et al. A consensus map for Ug99 stem rust resistance loci in wheat. Theor Appl Genet. 2014;127(7):1561–81.
Singh RP, Herrera-Foessel SA, Huerta-Espino J, Lan CX, Basnet BR, Bhavani S, et al. Pleiotropic gene Lr46/Yr29/Pm39/Ltn2 confers slow rusting, adult plant resistance to wheat stem rust fungus. New Delhi, India: Borlaug Global Rust Initiative 2013 Technical Workshop; 2013. p. 17.
Letta T, Maccaferri M, Badebo A, Ammar K, Ricci A, Crossa J, et al. Searching for novel sources of field resistance to Ug99 and Ethiopian stem rust races in durum wheat via association mapping. Theor Appl Genet. 2013;126(5):1237–56.
Haile J, Nachit M, Hammer K, Badebo A, Röder M. QTL mapping of resistance to race Ug99 of Puccinia graminis f. sp. tritici in durum wheat (Triticum durum Desf.). Mol Breeding. 2012;30(3):1479–93.
Mago R, Verlin D, Zhang P, Bansal U, Bariana H, Jin Y, et al. Development of wheat–Aegilops speltoides recombinants and simple PCR-based markers for Sr32 and a new stem rust resistance gene on the 2S#1 chromosome. Theor Appl Genet. 2013;126(12):2943–55.
McIntosh RA, Dubcovsky J, Rogers WJ, Morris CF, Appels R, Xia XC. Catalogue of gene symbols for wheat: 2011 supplement. Annual Wheat Newsletter. 2011;57:303–21.
Yu L-X, Lorenz A, Rutkoski J, Singh R, Bhavani S, Huerta-Espino J, et al. Association mapping and gene–gene interaction for stem rust resistance in CIMMYT spring wheat germplasm. Theor Appl Genet. 2011;123(8):1257–68.
Hiebert C, Fetch Jr T, Zegeye T. Genetics and mapping of stem rust resistance to Ug99 in the wheat cultivar Webster. Theor Appl Genet. 2010;121(1):65–9.
Tsilo TJ, Jin Y, Anderson JA. Microsatellite markers linked to stem rust resistance allele Sr9a in wheat. Crop Sci. 2007;47(5):2013–20.
Rouse M, Nava I, Chao S, Anderson J, Jin Y. Identification of markers linked to the race Ug99 effective stem rust resistance gene Sr28 in wheat (Triticum aestivum L.). Theor Appl Genet. 2012;125(5):877–85.
Spielmeyer W, Sharp PJ, Lagudah ES. Identification and validation of markers linked to broad-spectrum stem rust resistance gene Sr2 in wheat (Triticum aestivum L.). Crop Sci. 2003;43(1):333–6.
Rouse MN, Talbert LE, Singh D, Sherman JD. Complementary epistasis involving Sr12 explains adult plant resistance to stem rust in Thatcher wheat (Triticum aestivum L.). Theor Appl Genet. 2014;127(7):1549–59.
Lagudah ES, McFadden H, Singh RP, Huerta-Espino J, Bariana HS, Spielmeyer W. Molecular genetic characterization of the Lr34/Yr18 slow rusting resistance gene region in wheat. Theor Appl Genet. 2006;114(1):21–30.
Lagudah ES, Krattinger SG, Herrera-Foessel S, Singh RP, Huerta-Espino J, Spielmeyer W, et al. Gene-specific markers for the wheat gene Lr34/Yr18/Pm38 which confers resistance to multiple fungal pathogens. Theor Appl Genet. 2009;119(5):889–98.
Herrera-Foessel S, Singh R, Lillemo M, Huerta-Espino J, Bhavani S, Singh S, et al. Lr67/Yr46 confers adult plant resistance to stem rust and powdery mildew in wheat. Theor Appl Genet. 2014;127(4):781–9.
Bansal U, Bariana H, Wong D, Randhawa M, Wicker T, Hayden M, et al. Molecular mapping of an adult plant stem rust resistance gene Sr56 in winter wheat cultivar Arina. Theor Appl Genet. 2014;127(6):1441–8.
Yu L-X, Morgounov A, Wanyera R, Keser M, Singh S, Sorrells M. Identification of Ug99 stem rust resistance loci in winter wheat germplasm using genome-wide association analysis. Theor Appl Genet. 2012;125(4):749–58.
Crossa J, Burgueño J, Dreisigacker S, Vargas M, Herrera-Foessel SA, Lillemo M, et al. Association analysis of historical bread wheat germplasm using additive genetic covariance of relatives and population structure. Genetics. 2007;177(3):1889–913.
Mago R, Bariana HS, Dundas IS, Spielmeyer W, Lawrence GJ, Pryor AJ, et al. Development of PCR markers for the selection of wheat stem rust resistance genes Sr24 and Sr26 in diverse wheat germplasm. Theor Appl Genet. 2005;111(3):496–504.
Bajgain P, Rouse M, Bhavani S, Anderson J. QTL mapping of adult plant resistance to Ug99 stem rust in the spring wheat population RB07/MN06113-8. Mol Breeding. 2015;35(8):1–15.
Hiebert C, Fetch T, Zegeye T, Thomas J, Somers D, Humphreys DG, et al. Genetics and mapping of seedling resistance to Ug99 stem rust in Canadian wheat cultivars ‘Peace’ and ‘AC Cadillac’. Theor Appl Genet. 2011;122(1):143–9.
Ghazvini H, Hiebert CW, Zegeye T, Liu S, Dilawari M, Tsilo T, et al. Inheritance of resistance to Ug99 stem rust in wheat cultivar Norin 40 and genetic mapping of Sr42. Theor Appl Genet. 2012;125(4):817–24.
Lopez-Vera EE, Nelson S, Singh RP, Basnet BR, Haley SD, Bhavani S, et al. Resistance to stem rust Ug99 in six bread wheat cultivars maps to chromosome 6DS. TAG Theoretical and applied genetics Theoretische und angewandte Genetik. 2014;127(1):231–9.
Somers DJ, Isaac P, Edwards K. A high-density microsatellite consensus map for bread wheat (Triticum aestivum L.). Theor Appl Genet. 2004;109(6):1105–14.
Letta T, Olivera P, Maccaferri M, Jin Y, Ammar K, Badebo A, et al. Association mapping reveals novel stem rust resistance loci in durum wheat at the seedling stage. Plant Genome. 2014;7(1):1–13.
Gerechter-Amitai ZK, Wahl I, Vardi A, Zohary D. Transfer of stem rust seedling resistance from wild diploid einkorn to tetraploid durum wheat by means of a triploid hybrid bridge. Euphytica. 1971;20(2):281–5.
Rouse MN, Jin Y. Stem rust resistance in A-genome diploid relatives of wheat. Plant Dis. 2011;95:941–4.
Periyannan SK, Bansal UK, Bariana HS, Pumphrey M, Lagudah ES. A robust molecular marker for the detection of shortened introgressed segment carrying the stem rust resistance gene Sr22 in common wheat. Theor Appl Genet. 2011;122(1):1–7.
Mago R, Spielmeyer W, Lawrence G, Lagudah E, Ellis J, Pryor A. Identification and mapping of molecular markers linked to rust resistance genes located on chromosome 1RS of rye using wheat-rye translocation lines. Theor Appl Genet. 2002;104(8):1317–24.
Acknowledgements
Funding for this project was provided by the USDA-NIFA Triticeae Coordinated Agricultural Project (CAP Grant no. 2011-68002-30029), the Durable Rust Resistance in Wheat project administrated by Cornell University and funded by the Bill and Melinda Gates Foundation and the United Kingdom Department for International Development, the USDA-ARS National Plant Disease Recovery System, and USDA-ARS appropriated project 3640-21220-021-00. Lines for the AM panel were contributed by Curtis Pozniak (University of Saskatchewan), Dean Spaner (University of Alberta), Gavin Humphries (Agriculture and Agri-Food Canada), Jianli Chen (University of Idaho), Karl Glover (South Dakota State University), Jim Anderson (University of Minnesota), Jorge Dubcovsky (University of California-Davis), Luther Talbert (Montana State University), Mike Pumphrey (Washington State University), Pierre Hucl (University of Saskatchewan), Ron DePauw (Agriculture and Agri-Food Canada), Ron Knox (Agriculture and Agri-Food Canada), and the International Maize and Wheat Improvement Center (CIMMYT). We also thank our colleagues at the USDA-ARS Small Grains Genotyping Center, the Ethiopian Institute of Agricultural Research (EIAR), Kenya Agricultural and Livestock Research Organization (KALRO), International Maize and Wheat Improvement Center (CIMMYT), and University of Minnesota Supercomputing Institute for providing the necessary resources at various stages of this study.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
PB carried out the statistical analysis, interpretation of the results, and drafted the manuscript. MNR, SB, RW, PNN, and WL supervised planting of the GWAS panel, disease inoculation, and participated in agronomic supervision in the East African rust nurseries. MNR, SB, and TG recorded the phenotype. MNR designed the experiments and conducted seedling disease assays. MOP coordinated the collection of wheat lines used in this study. MOP and PBulli supervised genotyping of the plant materials. MNR, PBulli, MOP, and JAA supervised the project and assisted in preparing the manuscript. All authors contributed to and approved the final manuscript.
Authors’ information
Not applicable.
Availability of data and materials
Not applicable.
Additional file
Additional file 1:
Additional details on data analysis on the GWAS panel. The file should be opened in Microsoft Office Excel. (XLSX 433 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Bajgain, P., Rouse, M., Bulli, P. et al. Association mapping of North American spring wheat breeding germplasm reveals loci conferring resistance to Ug99 and other African stem rust races. BMC Plant Biol 15, 249 (2015). https://doi.org/10.1186/s12870-015-0628-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-015-0628-9