
Abstract
Background
Bacillus cereus is implicated in severe foodborne infection in humans. This study intended to assess the occurrence, groEL gene sequencing, biofilm production, and resistance profiles of emerged multidrug resistant (MDR) B. cereus in meat and meat product samples. Moreover, this work highlights the virulence and toxigenic genes (hblABCD complex, nheABC complex, cytK, ces, and pc-plc) and antimicrobial resistance genes (bla1, tetA, bla2, tetB, and ermA).
Methods
Consequently, 200 samples (sausage, minced meat, luncheon, beef meat, and liver; n = 40 for each) were indiscriminately collected from commercial supermarkets in Port Said Province, Egypt, from March to May 2021. Subsequently, food samples were bacteriologically examined. The obtained isolates were tested for groEL gene sequence analysis, antibiotic susceptibility, biofilm production, and PCR screening of toxigenic and resistance genes.
Results
The overall prevalence of B. cereus among the inspected food samples was 21%, where the highest predominance was detected in minced meat (42.5%), followed by beef meat (30%). The phylogenetic analysis of the groEL gene exposed that the examined B. cereus strain disclosed a notable genetic identity with other strains from the USA and China. Moreover, the obtained B. cereus strains revealed β-hemolytic activity, and 88.1% of the recovered strains tested positive for biofilm production. PCR evidenced that the obtained B. cereus strains usually inherited the nhe complex genes (nheA and nheC: 100%, and nheB: 83.3%), followed by cytK (76.2%), hbl complex (hblC and hblD: 59.5%, hblB: 16.6%, and hblA: 11.9%), ces (54.7%), and pc-plc (30.9%) virulence genes. Likewise, 42.9% of the examined B. cereus strains were MDR to six antimicrobial classes and encoded bla1, bla2, ermA, and tetA genes.
Conclusion
In summary, this study highlights the presence of MDR B. cereus in meat and meat products, posing a significant public health risk. The contamination by B. cereus is common in minced meat and beef meat. The molecular assay is a reliable fundamental tool for screening emerging MDR B. cereus strains in meat and meat products.
Similar content being viewed by others


Background
Communicable diarrheal diseases are incriminated in significant morbidity and mortalities all over the world [1]. Bacillus cereus (B. cereus) is one the utmost predominant food-borne pathogens, resulting in severe food poisoning. The disease is mainly associated with severe diarrhea, vomiting, liver failure, abdominal pain, and necrotic enteritis [2]. Moreover, B. cereus is reported frequently as the most prevailing pathogen found in several food products, including meat and meat products, milk products, and food of plant origin, which is considered a public health threat [3].
B. cereus is a Gram-positive, motile, and spore-forming rod. B. cereus, a ubiquitous pathogen, is widely distributed in food, soil, water, and plants [4]. The pathogenicity of B. cereus is endorsed mainly by various virulence factors and toxins encoded by the corresponding genes. The infection takes place as a result of the ingestion of contaminated food with B. cereus. The cells of B. cereus attached to the host intestinal mucosa then undergo colonization, followed by enterotoxin production [5, 6]. The most common virulence factors associated with B. cereus infection include the heat-labile enterotoxins group: 1-cytotoxin K (regulated by the cytK gene), 2-non-hemolytic enterotoxin (encoded by the nheABC gene complex), and 3-hemolysin BL (encoded by the hblABCD gene complex), the cereulide toxin (regulated by the ces gene), and phospholipase C (regulated by the pc-plc gene) [7,8,9]. Several food poisoning outbreaks triggered by B. cereus were reported globally [10]. The disease is associated with two clinical forms: diarrheal and emetic forms. Cytotoxin K, a potent enterotoxin, is considered the main virulence determinant incriminated in severe diarrhea. Besides, the heat-labile enterotoxins are responsible for the diarrheal syndrome. Moreover, the emetic syndrome is ascribed mainly to the cereulide toxin [6, 11, 12]. Furthermore, B. cereus is incriminated in severe infections in humans, including pneumonia [13], bacteremia in neonates [5], gas gangrene, bacterial meningitis, and eye infections [14].
The biofilm production is commonly associated with B. cereus to enhance its survival in adverse environmental circumstances. Biofilms produced by B. cereus are considered the principal source of device contamination during food processing [15]. Spore production by B. cereus makes biofilm difficult to eliminate due to the resistance of spores to radiation and heat [16].
Antimicrobial resistance is markedly raised all over the world, suggesting a public health hazard [17]. B. cereus inherited several antibiotic-resistance genes that enable the bacteria to resist several antimicrobial classes [6]. B. cereus is frequently resistant to antibiotics belonging to the β-lactam class [18, 19]. Moreover, B. cereus could display acquired resistance to some antibiotics such as streptomycin, tetracycline, and erythromycin [20]. A previous molecular investigation emphasized that B. cereus harbored resistance genes against macrolides, tetracycline, and β-lactam antibiotics [6]. The indiscriminate use of antimicrobial agents consequences the development of multidrug-resistant pathogens [21, 22].
Herein, we intended to determine the prevalence, groEL gene sequencing, biofilm production, and resistance profiles of emerged MDR B. cereus in meat and its products. Moreover, this work highlights the virulence and toxigenic genes (hblABCD complex, nheABC complex, cytK, ces, and pc-plc) and antimicrobial resistance genes (bla1, tetA, bla2, tetB, and ermA) inherited and acquired by the obtained isolates.
Materials and methods
Sampling
Two hundred samples (sausage, minced meat, luncheon, beef meat, and liver; n = 40 for each) were indiscriminately collected from commercial supermarkets in Port-Said Province, Egypt, from March to May 2021. Consequently, samples were transported as soon as possible to the laboratory.
Isolation, identification, and enumeration of B. cereus
About 25 g of each collected sample was homogenized in 225 mL of tryptic soy broth (BD Difco, USA) using a bag-mixer stomacher followed by serial dilution. Afterward, 0.1 ml of each diluted specimen was spread evenly with a spreader onto Mannitol Egg-Yolk Polymyxin (MYP) (BD Difco, USA) agar plates and left incubated at 37 °C for 24 h [23]. Typical B. cereus colonies were pink and bounded by a zone of precipitation, specifying the lecithinase activity. Then, colonies were enumerated with a colony counter. B. cereus was identified as consistent with culture and morphological characteristics, endospores, and biochemical tests (catalase, indole, H2S production, oxidase, methyl red, sugar fermentation, and Voges-Proskauer tests), according to Maturin and Peeler [24]. The identity of isolates was determined genetically using specific primers of groEL gene, as previously described [25].
groEL gene sequencing
Herein, the retrieved B. cereus isolates revealed similar phenotypic characteristics; therefore, a PCR product of one indiscriminately chosen isolate was used for groEL sequencing in both directions (QIAGEN Sciences Inc., Germantown, MD, USA). The Bigdye Terminator V3.1 cycle sequencing kit was used in gene sequencing (Thermo Fisher Scientific, Waltham, MA, USA). Besides, the phylogenetic tree was assembled in accordance with the neighbor-joining approach using the MEGA 11 software [26].
Hemolytic activity and biofilm production
The retrieved strains were streaked on 5% sheep blood agar (BD Difco, USA) and incubated at 24 °C for 48 h. The occurrence of β-hemolysis specifies the positive result, according to Wiwat and Thiramanas [27].
Using the microtitre plate method, a 200 µl suspension of B. cereus (108 CFU/ mL) in tryptone soy broth (Difco, USA) was added to a 96-well polystyrene microtiter plate. The bacterial cell suspension was incubated at 30 °C for 24 h. Subsequently, plates were decanted, washed, dried for 30 min, and stained with crystal violet for 15 min. After staining, the plates were decanted, washed, and dried for 15 min. For each well, 150 µL of ethanol (95%) was added, and plates were left for 30 min. The absorbance was estimated at 595 nm using a microplate reader (un-inoculated wells included as negative controls). Besides, B. cereus ATCC 10876 was involved as a positive control. The tested isolates were classified in consistent their OD value into strong (4 x ODc < ODs), moderate (2 x ODc < ODs ≤ 4 x ODc), weak (ODc < ODs ≤ 2 x ODc), and non (ODs ≤ ODc) biofilm producers [28].
Antimicrobial susceptibility testing
The antibiotic susceptibility of the obtained B. cereus strains was assessed on Mueller-Hinton agar (BD Dico, USA) using the disc diffusion test consistent with Park et al. [29]. Eleven antimicrobial discs were involved including erythromycin (E, 15 µg), ampicillin (AMP/ 10 µg), streptomycin (S/ 10 µg), amoxicillin (AMX/ 30 µg), sulfamethoxazole/trimethoprim (SXT/ 25 µg), cefepime (FEP/ 30 µg), ceftriaxone (CRO/ 30 µg), amoxicillin-clavulanic acid (AMC/ 30 µg), tetracycline (TE/ 30 µg), vancomycin (VA/ 30 µg), and levofloxacin (LEV/ 5 µg) (Oxoid, UK). The tested antimicrobial agents are commonly used in the veterinary and health sectors in Egypt. The interpretation of results was carried out as consistent with CLSI, 2017 [30]. S. aureus ATCC 29213 was involved as a control strain. Also, the obtained B. cereus strains were classified into MDR according to Magiorakos et al. [31]. Besides, the MAR (multiple antibiotic resistance) index was assessed consistent with Krumperman [32].
Detection of toxigenic and antibiotic resistance genes
Detection of virulence-related (hblABCD, nheABC, cytK, ces, and pc-plc) and antibiotic resistance genes (tetA, bla1, ermA, tetB, and bla2) was performed using PCR. The extraction of genomic DNA was performed by the QIAamp DNA Mini Kit (QIAGEN Sciences Inc., Germantown, USA). The bacterial pellet was mixed with 180 µL Buffer AL in a 1.5 ml microcentrifuge tube and incubated at 70 °C for 10 min. Subsequently, 200 µL of was added, mixed by pulse-vortex for 15 s, and centrifuged to remove drops from inside the lid of the collecting tube. The mixture was transferred to the spin column and centrifuged at 6500 x g for 1 min. The spin column was then moved to a 2 mL Eppendorf tube and washed multiple times with 500 µL of Buffer AW1 and AW2 at 6,000–20,000 x g for 1–3 min. The extracted DNA was collected in a new collecting tube by adding 200 µL of elution Buffer AE. Moreover, DNA-free reactions were used as negative controls, while reference B. cereus strains (The AHRI, Dokki, Egypt) were used as positive controls. Table 1 illustrates the used primer sequences.
Statistical analyses
G*Power (3.1.9.7) was used to evaluate sample size using Effect size f = 0.25 and power (1-β err prob = 0.80). Categorical variables were presented as frequencies and percentages. Associations between categorical variables were determined using the chi-square test (SAS software, version 9.4, SAS Institute, Cary, NC, USA) (p-value < 0.05 points to a significant variance). The “cor” function in the R-software (version 4.0.2; https://www.r-project.org/) and the “corrplot” package were used to create a correlation matrix for resistance profiles and resistance genes, as well as biofilm production and antimicrobial resistance. The heatmap of antimicrobial resistance patterns was performed using the GraphPad Software (version 8.0.1).
Results
Phenotypic features and the frequency of B. cereus isolates
Herein, the prevalence of B. cereus in the inspected food samples was 21% (42/200). The highest prevalence was noticed in minced meat (42.5%; 17/40), followed by beef meat (30%; 12/40), sausage (12.5%; 5/40), luncheon (10%; 4/40), and liver (10%; 4/40), as demonstrated in Table 2. Statistically, there is a significant variance in the frequency of B. cereus in various inspected food samples (p < 0.05).
The obtained isolates exhibited the distinctive morphological features of B. cereus, where all the isolates were Gram-positive, motile, short bacilli with non-bulging endospores. On MYP media, colonies were pink with a characteristic precipitation zone (lecithinase positive). Moreover, the obtained isolates tested positive for Voges-Proskauer, catalase, citrate utilization, glucose fermentation, and nitrate reduction. Likewise, the isolates tested negative for oxidase, H2S production, methyl red, indole, and mannitol fermentation. Besides, all the obtained isolates inherited the groEL gene.
In this study, the mean of B. cereus count was 11.75 × 103, 3 × 103, 2.87 × 103, 2.56 × 103, and 2.1 × 103 CFU/g in the examined minced meat, beef meat, sausage, luncheon, and liver samples, respectively (Table 2).
Phylogenetic analysis of the groEL gene
The phylogenetic analysis confirmed that the examined B. cereus strain (Accession number: MZ424866) revealed a notable genetic similarity with other isolates from various geographical areas. For instance, B. cereus strain Q1 (100%) of China (Accession number: CP000227), strain AH187 (99.79%) of the USA (Accession number: CP001177), and B. cereus strain JEM-2 (99.36%) isolated from the USA (Accession number: CP018935), as illustrated in Fig. 1.

This figure clarifies the phylogenetic analyses of groEL sequencing. The tree illuminates the genetic similarity among the selected B. cereus strain in this study and other strains acquiesced in the GenBank database. The tree was generated through bootstrap analysis with 1000 replicates, and the results are depicted above the branches
Hemolysis and biofilm production ability
All the tested B. cereus strains (n = 42) disclosed β-hemolysis on sheep blood agar. Moreover, 88.1% (37/42) of the recovered strains tested positive for biofilm production. Among the positive B. cereus strains, four isolates (10.8%, 4/37) were weak biofilm producers, ten isolates (27%, 10/37) were moderate biofilm producers, and twenty-three isolates (62.2%, 23/37) were strong biofilm producers (Fig. 2).

The rate of biofilm production between the retrieved B. cereus strains
Antimicrobial susceptibility of the retrieved B. cereus strains
Resistance to antimicrobial agents among the B. cereus isolates was identified as follows: ampicillin and amoxicillin (100%, n = 42), tetracycline (85.7%, n = 36), cefepime and ceftriaxone (80.9%, n = 34), amoxicillin-clavulanic acid (78.6%, n = 33), trimethoprim-sulfamethoxazole (76.2%, n = 32), and erythromycin (42.9%, n = 18). Moreover, the recovered isolates were sensitive to vancomycin (100%, n = 42) and levofloxacin (95.2%, n = 40) (Table 3; Fig. 3). Statistically, the tested B. cereus strains revealed a significant variance in their susceptibility to various antimicrobials (p < 0.05).

The antibiogram of the retrieved B. cereus strains. The bar (0-100) indicates the percentage of susceptibility
Presence of toxigenic and antimicrobial resistance genes
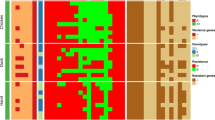
Virulence-related genes were identified in B. cereus isolates as follows: nhe complex genes (nheA and nheC: 100%, n = 42, and nheB: 83.3%, n = 35), followed by cytK (76.2%, n = 32), hbl complex (hblC and hblD: 59.5%, n = 25, hblB: 16.6%, n = 7, and hblA: 11.9%, n = 5), ces (54.7%, n = 23), and pc-plc (30.9%, n = 13) virulence genes. Also, PCR evidenced that the tested B. cereus isolates inherited or acquired the bla1, bla2, tetA, ermA, and tetB antibiotic resistance genes with a prevalence of 100% (n = 42), 80.9% (n = 34), 71.4% (n = 30), 42.9% (n = 18), and 14.3% (n = 6), respectively (Table 4; Fig. 4). Statistically, a significant variance (p < 0.05) was noticed in the frequency of toxigenic and resistance genes in the recovered strains.

The occurrence of toxigenic and antimicrobial resistance genes in the retrieved B. cereus from food samples
Phenotypic and genotypic resistance patterns of the obtained B. cereus strains
Herein, 42.9% (18/42) of the examined B. cereus strains were MDR to eight antimicrobial agents (six antimicrobial classes) and encoded bla1, bla2, ermA, and tetA genes, while 28.6% (12/42) of the isolated B. cereus strains were MDR to eight antimicrobial agents (six antimicrobial classes) and inherited tetA, bla1, and bla2 genes. Besides, 7.1% (3/42) of the recovered strains expressed multidrug resistance to five antimicrobial agents (three antimicrobial classes) and encoded bla1 and bla2 genes, as clarified in Table 5; Fig. 5. In this study, the multiple antibiotic resistance (MAR) index values ranged 0.27–0.73 signifying that the obtained B. cereus strains were developed from high-risk contamination. Moreover, the correlation coefficient (r) was estimated between the identified resistance genes and the involved antimicrobial agents. Positive correlations were detected among bla1 gene, AMX, and AMP; bla2 and FEP; bla1 and CRO; ermA and E (r = 1 for each); bla2 and CRO (r = 0.99); bla1 and FEP (r = 0.99); tetA and TE (r = 0.97); bla1 and AMC (r = 0.98); bla2 and AMC (r = 0.91) as revealed in Fig. 6.

The occurrence of MDR among the retrieved B. cereus isolates from food samples

The correlation coefficient between the detected resistance genes and the involved antimicrobial agents (0.2 ≤ r ≤ 0.39: Weak, 0.4 ≤ r ≤ 0.59: Moderate, and r ≥ 0.6: Strong positive correlation)
The correlation between biofilm production and antimicrobial resistance
In the present study, the strong biofilm producers were resistant to eight antimicrobial agents. Besides, the moderate biofilm producers were resistant to 5–8 antimicrobial agents. Moreover, the weak biofilm producers were resistant to 3–4 antimicrobial agents, while the non-biofilm producers were resistant to 3 antimicrobial agents, as illustrated in Table 6. Statistically, a positive correlation was noticed between biofilm production and antimicrobial resistance (Fig. 7).

Illustrates the correlation between biofilm production and antimicrobial resistance
Discussion
Bacillus cereus is one the utmost predominant food-borne pathogens, resulting in severe food poisoning in humans. Herein, we intended to investigate the occurrence, groEL gene sequencing, biofilm production, and resistance traits of emerged MDR B. cereus in meat and its products. Moreover, this work highlights the toxigenic and antimicrobial resistance genes.
In this study, all the obtained isolates revealed the characteristic phenotypic traits of B. cereus and disclosed coordination in their biochemical activities, as previously reported by Gdoura-Ben et al. in Tunisia [39]. Herein, the total prevalence of B. cereus in the inspected samples was 21%, where the highest predominance was detected in minced meat and beef meat. These findings signify that contaminated ready-to-eat food products could be a probable risk to human consumers. The higher prevalence of minced meat and beef meat, in comparison with luncheon, could be attributed to the heat treatment of luncheon during the processing stages, which reduces the potential B. cereus contamination [40]. A higher frequency of B. cereus (35%) was recorded in ready-to-eat foods in China; the prevalence was 34% in the cocked meat samples [41]. B. cereus is a ubiquitous pathogen, so open-air shops raise the potential for environmental pollution with B. cereus spores. Moreover, poor hygienic procedures during the processing and storage of various food products favor the contamination with B. cereus [42, 43].
Alarmingly, in this work, the mean B. cereus counts in the examined food samples exceeded the permissible limits reported by the Health Protection Agency [44] (should be less than 103 CFU/g or mL). Moreover, Stenfors et al. [3] confirmed that lower doses of B. cereus in foods could result in severe food poisoning outbreaks in human consumers. Besides, the food is not suitable for human consumers if the B. cereus count is > 104 CFU/g or mL [45]. The remarkably high counts of B. cereus in food products could be attributed to inadequate hygienic measures during food processing, contamination of equipment, inappropriate handling, bad storage conditions, and inadequate sterilization of equipment and machines [46].
The findings of this study showed that all the B. cereus isolates carried the groEL gene. Wei et al. [47] reported that groEL is a consistent diagnostic biomarker when compared with the gyrB gene to differentiate B. cereus from other pathogens in food products. The sequence analysis of the groEL gene emphasized that the examined B. cereus strain revealed a notable genetic identity as well as cross-lineage with other strains derived from various geographical areas, such as B. cereus strain Q1 in China [48], B. cereus strain B. cereus strains JEM-2 and AH187 in the USA [49].
Remarkably, all retrieved B. cereus strains exhibited β-hemolysis on sheep blood agar. These findings agreed with those confirmed by Hwang and Park in the Republic of Korea [50]. Potent hemolytic activity is usually associated with foodborne B. cereus strains due to inherited factors [51]. In the current study, 88.1% of the recovered B. cereus strains tested positive for biofilm production (out of them, 62.2% were strong biofilm producers), suggesting the recovered isolates are highly pathogenic. Our findings agree with Osman et al. [52], who reported biofilm production in 83.3% of isolates (out of them, 33.3% were strong biofilm producers) obtained from different meat samples in Egypt. Strong biofilm producers are highly pathogenic and frequently resistant to phagocytosis, antimicrobials, and antiseptics [16]. The biofilms perform a substantial role in the binding of pathogens to the biotic and non-biotic surfaces that specify the positive correlation between biofilm production and the occurrence of infection. Besides, it reveals the potential public health threat of B. cereus in causing food poisoning to human consumers [53].
In this study, vancomycin and levofloxacin exerted potent antibacterial activity toward the recovered B. cereus strains from various examined food samples. These findings agree with those confirmed by Ikeda et al. in Japan [54], who reported that all the obtained B. cereus strains were highly susceptible to vancomycin, and only 10.3% were resistant to levofloxacin. Moreover, the obtained B. cereus strains were resistant to ampicillin, amoxicillin, tetracycline, cefepime, ceftriaxone, amoxicillin-clavulanic acid, and trimethoprim-sulfamethoxazole. Our findings agree with those highlighted by Savić et al. [55]. Also, a previous study in Egypt reported that 40% of the retrieved B. cereus isolates were MDR to eight tested antibiotics [19]. Besides, Yu et al. [41] confirmed that the recovered B. cereus isolates from food products in China were MDR to penicillin, cephalothin, ampicillin, cefoxitin, and amoxicillin-clavulanic acid. These outcomes confirmed the occurrence of MDR B. cereus in various food products, suggesting that ready-to-eat foods could be a main source of transmission of foodborne MDR B. cereus to human consumers [56]. The haphazard use of antimicrobials in the agriculture and health sectors favors the development of MDR strains [19].
Concerning the dissemination of toxigenic genes, our findings agree with the results confirmed by Tewari et al. in India [43]. Moreover, the combination of nheA and nheC genes was noticed in all tested strains, which agrees with the results reported by Fraccalvieri et al. in Italy [57]. In this work, all the obtained B. cereus strains inherited two or more enterotoxigenic genes, highlighting their public health significance as a causative agent of severe food poisoning in man, consistent with Owusu-Kwarteng et al. [58]. Foodborne B. cereus strains frequently encode one or more hbl complex genes [33]. Food poisoning caused by B. cereus is concomitant mainly with these virulence determinants: non-hemolytic enterotoxin (regulated by nheABC complex genes), the hemolysin BL (regulated by hblABCD complex genes), the cytotoxin K (regulated by cytK), and the cereulide toxin (regulated by the ces gene) [9, 59].
In this study, the recovered B. cereus strains were MDR to eight tested antimicrobial agents (six different classes) and commonly carried bla1, ermA, tetA, and bla2 genes. Our findings are consistent with those confirmed by Fiedler et al. in Germany [6]. Moreover, the MARI values were 0.27–0.73 (> 0.2), worryingly highlighting that the retrieved strains resulted from high-risk contamination. The bla1 gene is mainly responsible for penicillinase enzymatic activity, whereas the bla2 gene is related to cephalosporins and penicillinase enzymatic activity [60]. Furthermore, the combination of bla1 and bla2 resistance genes is mainly responsible for the resistance of the β-Lactam-β-lactamase inhibitor combination [61]. Likewise, the resistance of B. cereus strains to tetracycline is mainly accredited to the tetA and or tetB gene, and their resistance to erythromycin is endorsed by the ermA gene. The existence of tetA and tetB genes in B. cereus emphasized the horizontal transfer of antimicrobial resistance genes from resistant pathogens to B. cereus [37].
Herein, a positive correlation was detected between the biofilm production and the antimicrobial resistance. Strong biofilm-producing B. cereus strains are highly virulent and usually resistant to disinfectants and antibiotics [16]. Biofilm-producing B. cereus is a highly adapted pathogen that could gain resistance to several antibiotics due to incessant disclosure to the antimicrobials, harboring or acquiring the resistance genes. Moreover, biofilm plays a vital role in drug resistance by delivering a proper environment for resistance determinant transfer [62].
In summary, this study highlights the presence of MDR B. cereus in meat and meat products, posing a significant public health risk. The contamination by B. cereus is common in minced meat and beef meat. MDR B. cereus isolates from food products often exhibit biofilm production and commonly harbor the nhe complex, cytK, hbl complex, ces, and pc-plc virulence genes, and bla1, tetA or tetB, bla2, and ermA antibiotic resistance genes. Vancomycin and levofloxacin demonstrate promising antibacterial activity toward the retrieved isolates. The molecular assay is a reliable fundamental tool for screening emerging MDR B. cereus strains in meat and meat products.
Data availability
No datasets were generated or analysed during the current study.
References
Shujatullah F, Shukla I, Gupta R, Khan PA, Khan F. Emergence of antimicrobial resistance in Vibrio cholerae of North India. Int Res J Microbiol. 2012;3:136–9.
Park K. Park’s textbook of preventive and social medicine. Jabalpur Banarasidas Bhanot 463. 2011.
Stenfors Arnesen LP, Fagerlund A, Granum PE. From soil to gut: Bacillus cereus and its food poisoning toxins. FEMS Microbiol Rev. 2008;32(4):579–606.
Ramarao N, Tran S-L, Marin M, Vidic J. Advanced methods for detection of Bacillus cereus and its pathogenic factors. Sensors. 2020;20(9):2667.
Hilliard NJ, Schelonka RL, Waites KB. Bacillus cereus bacteremia in a preterm neonate. J Clin Microbiol. 2003;41(7):3441–4.
Fiedler G, Schneider C, Igbinosa EO, Kabisch J, Brinks E, Becker B, et al. Antibiotics resistance and toxin profiles of Bacillus cereus-group isolates from fresh vegetables from German retail markets. BMC Microbiol. 2019;19(1):1–13.
Ehling-Schulz M, Fricker M, Scherer S. Identification of emetic toxin producing Bacillus cereus strains by a novel molecular assay. FEMS Microbiol Lett. 2004;232(2):189–95.
Ehling-Schulz M, Vukov N, Schulz A, Shaheen R, Andersson M, Märtlbauer E, et al. Identification and partial characterization of the nonribosomal peptide synthetase gene responsible for cereulide production in emetic Bacillus cereus. Appl Environ Microbiol. 2005;71(1):105–13.
Li F, Zuo S, Yu P, Zhou B, Wang L, Liu C, et al. Distribution and expression of the enterotoxin genes of Bacillus cereus in food products from Jiangxi Province, China. Food Control. 2016;67:155–62.
EFSA. Panel on Biological Hazards (BIOHAZ). Risks for public health related to the presence of Bacillus cereus and other Bacillus spp. including Bacillus thuringiensis in foodstuffs. EFSA J. 2016;14(7):e04524.
Agata N, Ohta M, Yokoyama K. Production of Bacillus cereus emetic toxin (cereulide) in various foods. Int J Food Microbiol. 2002;73(1):23–7.
EFSA. European Centre for Disease Prevention and Control (ECDC). The European Union one health 2018 zoonoses report. EFSA J. 2019;17(12):e05926.
Hoffmaster AR, Hill KK, Gee JE, Marston CK, De BK, Popovic T, et al. Characterization of Bacillus cereus isolates associated with fatal pneumonias: strains are closely related to Bacillus anthracis and harbor B. Anthracis virulence genes. J Clin Microbiol. 2006;44(9):3352–60.
Bottone EJ. Bacillus cereus, a volatile human pathogen. Clin Microbiol Rev. 2010;23(2):382–98.
Gurler N, Oksuz L, Muftuoglu M, Sargin F, Besisik S. Bacillus cereus catheter related bloodstream infection in a patient with acute lymphoblastic leukemia. Mediterranean J Hematol Infect Dis. 2012;4(1).
Auger S, Ramarao N, Faille C, Fouet A, Aymerich S, Gohar M. Biofilm formation and cell surface properties among pathogenic and nonpathogenic strains of the Bacillus cereus group. Appl Environ Microbiol. 2009;75(20):6616–8.
Algammal AM, Eidaroos NH, Alfifi KJ, Alatawy M, Al-Harbi AI, Alanazi YF et al. Opr L gene sequencing, resistance patterns, virulence genes, Quorum Sensing and Antibiotic Resistance genes of XDR Pseudomonas aeruginosa isolated from broiler chickens. Infect Drug Resist. 2023:853–67.
Citron DM, Appleman MD. In vitro activities of daptomycin, ciprofloxacin, and other antimicrobial agents against the cells and spores of clinical isolates of Bacillus species. J Clin Microbiol. 2006;44(10):3814–8.
Algammal AM, Alfifi KJ, Mabrok M, Alatawy M, Abdel-Moneam DA, Alghamdi S et al. Newly emerging MDR B. cereus in Mugil Seheli as the First Report Commonly Harbor nhe, hbl, cyt K, and pc-Plc virulence genes and bla 1, bla 2, tet A, and erm a resistance genes. Infect Drug Resist. 2022:2167–85.
Jensen LB, Baloda S, Boye M, Aarestrup FM. Antimicrobial resistance among Pseudomonas spp. and the Bacillus cereus group isolated from Danish agricultural soil. Environ Int. 2001;26(7–8):581–7.
Shafiq M, Zeng M, Permana B, Bilal H, Huang J, Yao F, et al. Coexistence of bla NDM–5 and tet (X4) in international high-risk Escherichia coli clone ST648 of human origin in China. Front Microbiol. 2022;13:1031688.
Algammal A, Hetta HF, Mabrok M, Behzadi P. Emerging multidrug-resistant bacterial pathogens superbugs: a rising public health threat. Front Microbiol. 2023;14:1135614.
Ministry of Health of the People’s Republic of China. GB 4789.14–2014. National food safety standard: Food microbiological examination: Bacillus cereus. China Standard Press, Beijing. 2014.
Maturin L, Peeler J. BAM: aerobic plate count. US Food and Drug Administration. USA: Silver Spring, MD; 2001.
Chang Y-H, Shangkuan Y-H, Lin H-C, Liu H-W. PCR assay of the groEL gene for detection and differentiation of Bacillus cereus group cells. Appl Environ Microbiol. 2003;69(8):4502–10.
Tamura K, Stecher G, Kumar S. MEGA11: molecular evolutionary genetics analysis version 11. Mol Biol Evol. 2021;38(7):3022–7.
Wiwat C, Thiramanas R. Detection of hemolysin BL gene of Bacillus cereus isolates. Mahidol Univ J Pharm Sci. 2014;41:22–30.
Stepanović S, Vukovic D, Dakic I, Savic B, Svabic-Vlahovic M. A modified microtiter-plate test for quantification of 426 staphylococcal biofilm formation. J Microbiol Methods. 2000;40:175–9. https://doi.org/10.1016/S0167-7012(00)00122-6.
Park Y-B, Kim J-B, Shin S-W, Kim J-C, Cho S-H, Lee B-K, et al. Prevalence, genetic diversity, and antibiotic susceptibility of Bacillus cereus strains isolated from rice and cereals collected in Korea. J Food Prot. 2009;72(3):612–7.
CLSI. Bacillus. In: Clinical and Laboratory Standards Institute: Performance Standards for Antimicrobial Susceptibility Testing: 27th Ed 424 Informational Supplement. CLSI. ; 2017. CLSI Doc. M100-S20 (2017). Wayne, PA, USA.
Magiorakos A-P, Srinivasan A, Carey RB, Carmeli Y, Falagas M, Giske C, et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin Microbiol Infect. 2012;18(3):268–81.
Krumperman PH. Multiple antibiotic resistance indexing of Escherichia coli to identify high-risk sources of fecal contamination of foods. Appl Environ Microbiol. 1983;46(1):165–70.
Guinebretière M-H, Vr B, Nguyen-The C. Enterotoxigenic profiles of food-poisoning and food-borne Bacillus cereus strains. J Clin Microbiol. 2002;40(8):3053–6.
Ehling-Schulz M, Guinebretiere M-H, Monthán A, Berge O, Fricker M, Svensson B. Toxin gene profiling of enterotoxic and emetic Bacillus cereus. FEMS Microbiol Lett. 2006;260(2):232–40.
Schraft H, Griffiths M. Specific oligonucleotide primers for detection of lecithinase-positive Bacillus spp. by PCR. Appl Environ Microbiol. 1995;61(1):98–102.
Chen Y, Tenover FC, Koehler TM. β-Lactamase gene expression in a penicillin-resistant Bacillus anthracis strain. Antimicrob Agents Chemother. 2004;48(12):4873–7.
Rather MA, Aulakh RS, Gill JPS, Mir AQ, Hassan MN. Detection and sequencing of plasmid encoded tetracycline resistance determinants (tetA and tetB) from food–borne Bacillus cereus isolates. Asian Pac J Trop Med. 2012;5(9):709–12.
Adimpong DB, Sørensen KI, Thorsen L, Stuer-Lauridsen B, Abdelgadir WS, Nielsen DS, et al. Antimicrobial susceptibility of Bacillus strains isolated from primary starters for African traditional bread production and characterization of the bacitracin operon and bacitracin biosynthesis. Appl Environ Microbiol. 2012;78(22):7903–14.
Gdoura-Ben Amor M, Siala M, Zayani M, Grosset N, Smaoui S, Messadi-Akrout F, et al. Isolation, identification, prevalence, and genetic diversity of Bacillus cereus group bacteria from different foodstuffs in Tunisia. Front Microbiol. 2018;9:447.
Shawish R, Tarabees R. Prevalence and antimicrobial resistance of Bacillus cereus isolated from beef products in Egypt. Open Veterinary Journal. 2017;7(4):337–41.
Yu S, Yu P, Wang J, Li C, Guo H, Liu C, et al. A study on prevalence and characterization of Bacillus cereus in ready-to-eat foods in China. Front Microbiol. 2020;10:3043.
Ng Y-F, Wong S-L, Cheng H-L, Yu PH-F, Chan S-W. The microbiological quality of ready-to-eat food in Siu Mei and Lo Mei shops in Hong Kong. Food Control. 2013;34(2):547–53.
Tewari A, Singh S, Singh R. Incidence and enterotoxigenic profile of Bacillus cereus in meat and meat products of Uttarakhand, India. J Food Sci Technol. 2015;52:1796–801.
Health Protection Agency. Guidelines for assessing the microbiological safety of ready-to‐eat foods placed on the market. In.: Health Protection Agency London; 2009.
Food Standards Australia New Zealand (FSANZ). Microbiological quality guide for ready-to-eat foods. A guide to interpreting microbiological results; 2001.
Kumari S, Sarkar PK. Bacillus cereus hazard and control in industrial dairy processing environment. Food Control. 2016;69:20–9.
Wei S, Chelliah R, Park B-J, Park J-H, Forghani F, Park Y-S, et al. Molecular discrimination of Bacillus cereus group species in foods (lettuce, spinach, and kimbap) using quantitative real-time PCR targeting groEL and gyrB. Microb Pathog. 2018;115:312–20.
Xiong Z, Jiang Y, Qi D, Lu H, Yang F, Yang J, et al. Complete genome sequence of the extremophilic Bacillus cereus strain Q1 with industrial applications. J Bacteriol. 2009;191(3):1120–1.
Stevens MJ, Tasara T, Klumpp J, Stephan R, Ehling-Schulz M, Johler S. Whole-genome-based phylogeny of Bacillus cytotoxicus reveals different clades within the species and provides clues on ecology and evolution. Sci Rep. 2019;9(1):1984.
Hwang J-Y, Park J-H. Characteristics of enterotoxin distribution, hemolysis, lecithinase, and starch hydrolysis of Bacillus cereus isolated from infant formulas and ready-to-eat foods. J Dairy Sci. 2015;98(3):1652–60.
Pirhonen T, Andersson M, Jääskeläinen EL, Salkinoja-Salonen M, Honkanen-Buzalski T, Johansson T-L. Biochemical and toxic diversity of Bacillus cereus in a pasta and meat dish associated with a food-poisoning case. Food Microbiol. 2005;22(1):87–91.
Osman KM, Kappell AD, Orabi A, Al-Maary KS, Mubarak AS, Dawoud TM, et al. Poultry and beef meat as potential seedbeds for antimicrobial resistant enterotoxigenic Bacillus species: a materializing epidemiological and potential severe health hazard. Sci Rep. 2018;8(1):11600.
Coleri Cihan A, Karaca B, Ozel BP, Kilic T. Determination of the biofilm production capacities and characteristics of members belonging to Bacillaceae family. World J Microbiol Biotechnol. 2017;33:1–13.
Ikeda M, Yagihara Y, Tatsuno K, Okazaki M, Okugawa S, Moriya K. Clinical characteristics and antimicrobial susceptibility of Bacillus cereus blood stream infections. Ann Clin Microbiol Antimicrob. 2015;14(1):1–7.
Savić D, Miljković-Selimović B, Lepšanović Z, Tambur Z, Konstantinović S, Stanković N, et al. Antimicrobial susceptibility and β-lactamase production in Bacillus cereus isolates from stool of patients, food and environment samples. Vojnosanit Pregl. 2016;73(10):904–9.
Bhunia AK. Foodborne microbial pathogens: mechanisms and pathogenesis. New York, NY. Springer; 2018. https://doi.org/10.1007/978-1-4939-7349-1.
Fraccalvieri R, Bianco A, Difato LM, Capozzi L, Del Sambro L, Simone D, et al. Toxigenic genes, pathogenic potential and antimicrobial resistance of Bacillus cereus group isolated from ice cream and characterized by whole genome sequencing. Foods. 2022;11(16):2480.
Owusu-Kwarteng J, Wuni A, Akabanda F, Tano-Debrah K, Jespersen L. Prevalence, virulence factor genes and antibiotic resistance of Bacillus cereus sensu lato isolated from dairy farms and traditional dairy products. BMC Microbiol. 2017;17(1):1–8.
Berthold-Pluta A, Pluta A, Garbowska M, Stefańska I. Prevalence and toxicity characterization of Bacillus cereus in food products from Poland. Foods. 2019;8(7):269.
Fenselau C, Havey C, Teerakulkittipong N, Swatkoski S, Laine O, Edwards N. Identification of β-lactamase in antibiotic-resistant Bacillus cereus spores. Appl Environ Microbiol. 2008;74(3):904–6.
Sornchuer P, Saninjuk K, Prathaphan P, Tiengtip R, Wattanaphansak S. Antimicrobial susceptibility profile and whole-genome analysis of a strong biofilm-forming Bacillus sp. B87 strain isolated from food. Microorganisms. 2022;10(2):252.
Michaelis C, Grohmann E. Horizontal Gene Transfer of Antibiotic Resistance Genes in Biofilms A. 2023; 12: 328. https://doi.org/10.3390/antibiotics12020328.
Acknowledgements
Not applicable.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
A.M.A. conception and study design. A.M.A, R.M.E, H.M.E, S.G.E, G.M.M, and H.F.H; conducted the experiments. A.M.A. drafted the manuscript. A.M.A, R.M.E, H.M.E, S.G.E, G.M.M, H.F.H, S.A, R.A, H.G, E.A.A, and T.M.A; acquisition of data, statistical analysis, and interpretation of data. AMA, S.A, R.A, E.A.A, T.M.A, and H.G; critically reviewing the article. All authors have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
All the experiments and protocols were approved by the Scientific Research Ethics Committee, Suez Canal University (Approval No.: SCU 2023069).
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Algammal, A.M., Eid, H.M., Alghamdi, S. et al. Meat and meat products as potential sources of emerging MDR Bacillus cereus: groEL gene sequencing, toxigenic and antimicrobial resistance. BMC Microbiol 24, 50 (2024). https://doi.org/10.1186/s12866-024-03204-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12866-024-03204-9