
Abstract
Background
Shiga toxin-producing Escherichia coli (STEC) is a zoonotic pathogen, that is transmitted from a variety of animals, especially cattle to humans via contaminated food, water, feaces or contact with infected environment or animals. The ability of STEC strains to cause gastrointestinal complications in human is due to the production of Shiga toxins (sxt). However, the transmission of multidrug-resistance STEC strains are linked with a severity of disease outcomes and horizontal spread of resistance genes in other pathogens. The result of this has emerged as a significant threat to public health, animal health, food safety, and the environment. Therefore, the purpose of this study is to investigate the antibiogram profile of enteric E. coli O157 isolated from food products and cattle faeces samples in Zagazig City, Al-Sharkia, Egypt, and to reveal the presence of Shiga toxin genes stx1 and stx2 as virulence factors in multidrug-resistant isolates. In addition to this, the partial 16S rRNA sequencing was used for the identification and genetic recoding of the obtained STEC isolates.
Results
There was a total of sixty-five samples collected from different geographical regions at Zagazig City, Al-Sharkia—Egypt, which were divided into: 15 chicken meat (C), 10 luncheon (L), 10 hamburgers (H), and 30 cattle faeces (CF). From the sixty-five samples, only 10 samples (one from H, and 9 from CF) were identified as suspicious E. coli O157 with colourless colonies on sorbitol MacConkey agar media with Cefixime- Telurite supplement at the last step of most probable number (MPN) technique. Eight isolates (all from CF) were identified as multidrug-resistant (MDR) as they showed resistance to three antibiotics with multiple antibiotic resistance (MAR) index ≥ 0.23, which were assessed by standard Kirby-Bauer disc diffusion method. These eight isolates demonstrated complete resistance (100%) against amoxicillin/clavulanic acid, and high frequencies of resistance (90%, 70%, 60%,60%, and 40%) against cefoxitin, polymixin, erythromycin, ceftazidime, and piperacillin, respectively. Those eight MDR E. coli O157 underwent serological assay to confirm their serotype. Only two isolates (CF8, and CF13), both from CF, were showed strong agglutination with antisera O157 and H7, as well as resistance against 8 out of 13 of the used antibiotics with the highest MAR index (0.62). The presence of virulence genes Shiga toxins (stx1 and stx2) was assessed by PCR technique. CF8 was confirmed for carrying stx2, while CF13 was carrying both genes stx1, and stx2. Both isolates were identified by partial molecular 16S rRNA sequencing and have an accession number (Acc. No.) of LC666912, and LC666913 on gene bank. Phylogenetic analysis showed that CF8, and CF13 were highly homologous (98%) to E. coli H7 strain, and (100%) to E. coli DH7, respectively.
Conclusion
The results of this study provides evidence for the occurrence of E. coli O157:H7 that carries Shiga toxins stx1 and/or stx2, with a high frequency of resistance to antibiotics commonly used in human and veterinary medicine, in Zagazig City, Al-Sharkia, Egypt. This has a high extent of public health risk posed by animal reservoirs and food products with respect to easy transmission causing outbreaks and transfer resistance genes to other pathogens in animal, human, and plants. Therefore, environmental, animal husbandry, and food product surveillance, as well as, clinical infection control, must be strengthened to avoid the extra spread of MDR pathogens, especially MDR STEC strains.
Similar content being viewed by others


Background
Shiga toxin-producing Escherichia coli (STEC) is considered as an important foodborne zoonotic pathogen worldwide that causes gastrointestinal complications in humans [1]. Healthy domestic ruminants such as cattle, sheep, goats, and pigs can harbor STEC and E. coli O157:H7 in their faeces and are thus, natural reservoirs of these pathogens. Other animals, such as, chicken, dogs, horses, etc. are considered as spillover hosts that are susceptible to STEC colonization and can transmit disease [2]. Humans are sporadically infected with STEC that emerged as foodborne pathogens through the ingestion of food, water, or vegetables contaminated with animal faeces, or by direct contact with infected animals [3]. Then, STEC infections are easily transmitted from person-to-person [4]. Diseases caused by STEC isolates, especially serotype O157:H7, ranges from mild diarrhoea to haemorrhagic colitis (HC) and haemolytic uraemic syndrome (HUS). These complications typically affect children, elderly, and immunocompromised patients [5]. The systemic illness of HC, which naturally follows gastrointestinal infections with STEC, typically begins with abdominal cramps and diarrhea, followed by bloody diarrhea. Additionally, HUS which is characterized by acute renal failure, noon-bloody diarrhea converted to bloody diarrhea after one to five days in almost 80% of both, children and adult cases [6]. Due to the severity and pathogenicity of STEC-associated diseases, hospitalization is frequently required and may lead to morbidity and mortality in more than 2–10% of cases [7, 8]. This is mainly attributed to Shiga toxin (Stx) production, as a main virulence factor in STEC [5]. Shiga toxins (Stx) have two major families Stx1 and Stx2, with 70 percent similarity at the amino acid level [9]. Both toxins produce diseases in people, Stx1 is produces less serious diseases linked to human illness, while Stx2 is more often linked with the development of HC and HUS [10]. STEC refers to E.coli pathotype capable of producing Stx1, and/or Stx2 [11]. The Shiga toxins mode of actions starts when the pentamer of matching B subunits from Stx permits the toxin to bind to globotriaosylceramide (Gb3) on the host cell [12]. Then, Shiga toxins A subunit inhibits protein synthesis by removing an adenosine residue from the 28S rRNA of the host cell 60S ribosome. Both epithelial and endothelial cells intoxicated with Stx may undergo an apoptotic cell death after intoxication [13, 14]. Moreover, Stx acts on the host cell signal transduction and immune modulation, causing infections [15].
Remarkably, different serotypes of E. coli strains are capable of producing Shiga toxins and are associated with diseases worldwide [16]. O157 and non‐O157 STEC strains have been isolated from cases of sporadic diarrhoea and HUS with varying frequencies [11]. Johnson et al. 1996 [17] reported that a quarter of HUS cases in USA were caused by non-O157 STEC. Both, O157 and non-O157 STEC can cause outbreaks attributed to the consumption of contaminated food products, in particular, those derived from cattle, or consumption of water, or vegetables contaminated with ruminant faeces, or direct contact with infected animals [18].
Antimicrobial resistance is a global public and animal health problem, with negative impacts on environmental, individual, social and economic levels [19]. The term multidrug-resistant (MDR) refers to strains that have shown resistance to more than two antibiotics from different antibiotic groups. This is an unfortunate phenomenon that leads to depletion of treatments [20]. Although antibiotic therapy is not a treatment option for human infections caused by STEC O157:H7, MDR strains of STEC O157:H7 or other serotypes (non-O157) should be of great concern. This is because they can easily transfer resistance genes horizontally to other pathogens in hosts and environment [21]. Retail animal food products act as an important vehicle for community wide dissemination of MDR STEC strains. Egypt is part of the Middle East, this geographical region has the maximum annual incidence rates of human STEC cases (122:249 cases/ 100,000 people/ year; 160 HUS cases) compared to other regions worldwide [22]. Thus, if MDR STEC strains are not detected early and addressed, this will definitively lead to serious health problem concerns due to the levitation in the cost of managing infections, and limitation of antibiotic options and emergence of superbugs (MDR strains) in the environment including the aquatic ecosystem. To the best of our knowledge, there is no existing data about the presence of MDR isolates of STEC O157:H7 and non-O157 isolated from both, retail animal food products and cattle faeces in Al-sharkia-Egypt. Therefore, the purpose of the present study was to determine the approximate percentage of E. coli O157:H7 and the other serotypes present in the selected food products and cattle’s faeces collected from different geographical regions at Zagazig City, Al-Sharkia-Egypt. Additionally, their susceptibility to widely used antibiotics in human and animal husbandry was evaluated, and it was determined if the highly resistant isolates carry the virulence factors of Shiga toxins genes. Finally, molecular identification for the highly MDR E. coli O157:H7 that carries Shiga toxin genes based on partial sequence of 16S rRNA, record the accession number with determination of phylogenetic tree was recorded.
Results
The most probable number (MPN) method is a highly used qualitative and quantitative technique for enumerating the fecal coliform bacteria (FCB) in different food, water, animal or human samples [23, 24]. The results of MPN values are illustrated in Table 1. The number of observed positive tubes with gas production or change in media colour indicated the FCB presence in these samples and called presumptive test, Table 2. Out of 65 total samples, in the presumptive test, FCB were isolated from 46%, 60%, 50%, and 100% of chicken meat, luncheon, hamburger and faeces samples, respectively. The prevalence of enteric E. coli in the same samples were decreased to 26.7%, 30%, 20%, and 50% after streaking on EMB agar (confirmatory test), as shown in Table 2. After that, only 10 isolates (one isolate from hamburger and 9 isolates from cattle’s faeces) were detected and identified as E. coli of serotype O157 by using selective differential sorbitol MacConkey agar media with Cefixime- Telurite supplement (CT-SMAC) in a complete test. Figure 1 (A & B) showed metallic sheen appearance and colourless colonies on EMB, and CT-SMAC, respectively.

Colonies of E. coli on different used selective differential media. A on eosin methylene blue (EMB) agar media with metallic sheen appearance, and B on sorbitol MacConkey agar media with Cefixime-Telurite supplement (CT-SMAC) with pale (colourless) appearance
Antibiotic susceptibility pattern for isolated E. coli O157
Ten suspected E. coli O157 isolates (1 from hamburger sample, and 9 from cattle’s feces samples) were selected and differentiated on CT-SMAC. Then, the isolates were screened for antibiotic susceptibility against the 13 chosen antibiotics, each with different modes of action, and widely used in human and veterinary medicine, Fig. 2 and Table 3. All isolates showed complete resistance (100%) against AMC, with high frequencies of resistance (90%, 70%, 60%,60%, and 40%) against FOX, PB, CAZ, E and PRL, respectively. On the other side, the same isolates showed complete sensitivity (100%) against IPM, AK, and CIP. This means that the higher potency of these antibiotics are able to overcome the pathogenicity of these isolates.

Percentage of resistance and sensitivity of the ten E. coli O157 isolates against the 13 chosen antibiotics, FOX: cefoxitin, FEP: cefepime, CAZ: ceftazidime, CRO: ceftriaxon, AMC: amoxicillin/clavulanic acid, PRL: piperacillin, IPM: imipenem, AK: amikacin, CN: gentamicin, SXT: trimethoprim/sulfamethoxazole, PB: polymixin, CIP: ciprofloxacin, and E: erythromycin.

Most of E. coli O157 isolates were resistant to the tested antibiotics. Multidruge resistance (defined as resistance to three antibiotic classes or more) was observed in 8 out of 9 faecal samples (89%) with the highest MAR index values (ranged between 0.23 to 0.62). Whereas, only one faecal sample (11%) expressed resistance to two antibiotics; FOX, and AMC, both antibiotics from β-lactam group. The same resistance pattern is observed in the only isolate recovered from hamburger (5H).
Out of 8 suspected E. coli O157 isolates, only two (CF8 and CF13) had clearly strong agglutinations with both antisera O157 and H7. This means that CF8 and CF13 were confirmed as STEC O157:H7. Noticeably, both isolates CF8 and CF13 were recovered from cattle faeces and showed the highest MAR index (0.62) amongst all isolates. Both isolates were resistant to 8 antibiotics (FOX, FEP, CAZ, CRO, AMC, PRL, PB, and E) belonging to 3 different groups (β-lactam, polypeptide, and macrolides).
PCR analysis was performed to confirm the presence of the main virulence factor in STEC O157:H7; Shiga toxins (stx1 (encodes variant 1 [Stx1]), and stx2 (encodes variant 2 [Stx2]).
Multiplex PCR for stx1, and stx2 genes
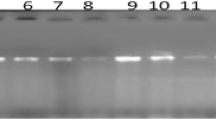
PCR analysis was performed to confirm the presence of main virulence factor in both STEC O157:H7 isolates; Shiga toxins genes (stx1 (encodes variant 1 [Stx1]), and stx2 (encodes variant 2 [Stx2])). Figure 3 showed that isolate CF8 contains stx2 gene while isolate CF13 had stx1 and stx2 genes.

Agarose gel electrophoresis of DNA fragments generated by multiplex PCR for virulence gene- shiga toxins of selected E. coli isolates, Lane M: DNA molecular weight marker (100 bp), Lane C+: Positive control, Lane C-: Negative control, Lane1: E.coli isolate of code no. CF8, Lane 2: E.coli isolate of code no. CF13
Molecular identification of multidrug- resistant bacterial isolates based on partial sequence of 16S rRNA gene
In Table 4, the identification of the two isolates of E.coli O157:H7 were confirmed using the 16S rRNA gene. The Gene-Bank nucleotide sequence accession numbers for partial sequences of 16S rRNA gene were generated in this study using DDBJ Center, and recorded as CF8 Escherichia coli LC666912 and Escherichia coli LC666913. Figure 4 showed the sequence of mono toxin E.coli O157:H7 isolate No (CF8) was similar 98% to strain CP024273,CP024269 and CP02422 that were enterotoxigenic Escherichia coli Strain E.coli O169:H41 [25, 26], and CP041429 which was Escherichia coli Stx2 producing strain serotype O100:H19 [27], as well as other strains of Escherichia coli. On the other hand, the sequence of di toxin E.coli O157:H7 isolate No (CF13) was similar 99% to Escherichia coli that cause diarrhea and gastrointestinal disease [28].

Phylogenetic tree analysis based on 16S rRNA nucleotide sequence alignment for our two E.coli O157:H7 isolates with other related members that possess the best similarity. The scale bar at the bottom (left) indicates similarity coefficient (%)
Discussion
Cattle are considered to be the principal reservoirs of pathogenic E. coli O157:H7 causing environmental hazards and human diseases, and transmission may occur through a variety of routes[29]. For example, the bovine faeces can contaminate crops, and drinking or recreational water intended for human consumption, animal contact, and person-to-person spread [30,31,32]. Additionally, consumption of contaminated raw/ undercooked meat or poultry products as hamburgers and luncheon is considered amongst the major routes for STEC O157:H7 and other serotypes transmission as a zoonotic foodborne pathogen [33, 34]. Our local results in Zagazig City, Al-Sharkia, Egypt indicated that, 10 isolates (1 from hamburger sample, and 9 from cattle’s faeces samples) out of 65 total samples (representing 15.4%) were highly suspected to be E. coli O157 by using EMB, and CT-SMAC media. However, E.coli ferment lactose and sucrose appear as metallic green sheen colony on EMB [35]. While, SMAC medium specific for isolation of non-fermented sorbitol E.coli O157 [36], and supplemented with cefixime and potassium tellurite to increase the selectivity of E.coliO157 isolation [37]. SMAC medium have high specificity for detection of E.coli O157.
Our results revealed high values of multiple antibiotic resistance (MAR) index for all suspected E. coli O157 isolates (ranged between 0.15 and 0.62). These values are higher than the accepted international standard for MAR index [38]. Whereas, the MAR index is calculated as the ratio between the number of antibiotics to which an organism is resistant / the total number of antibiotics to which the organism is exposed. For that, bacteria having MAR index ≥ 0.2 mainly originate from a high-risk source of contamination in which several antibiotics are used. Eighty % is the resistance percentage among our local suspected E.coli O157 isolates, that showed resistance against two or more antibiotics having different mode of actions, and can be classified as multidrug-resistant (MDR) isolates. This percentage of resistance among our local suspected E.coli O157 isolates were significantly higher than the isolates of the E. coli O157:H7 from cattle (53%) and beef (57%) reported by [39] from another geographic region (Spain, Europe). Among all the isolates, antibiotic resistance was 100%, 90%, 70%, 60%, and 60% against AMC, FOX, polymyxin B (PB), CAZ, and erythromycin (E), respectively. These results are consistent with previous study [40]. Several studies have reported complete resistance (100%) against AMC with 70% resistance against FOX [41, 42]. Meanwhile, other studies demonstrated a lower percentage of resistance (45.5%, and 36.4%) against AMC and PB, respectively, with complete resistance (100%) against E, in Malaysian and Korean E.coli O157 isolated from food and animal sources [43]. The same results were reported in poultry samples from Saudi Arabia [44] and from Bangladesh [45]. All antibiotics tested here are considered to be critically or highly important for human medicine by the World Health Organization, and commonly used in Egypt [46] Nevertheless, nearly all classes of antibiotics offered to humans have also been used in animal husbandry [47]. The same antibiotics used in this study were reported to be used to treat respiratory infections, diarrhoea, mastitis, and other infections in beef and dairy cattle [48]. In Egypt, several antibiotics have been recorded to be widely used in animal husbandry for several reasons, for example: to guarantee healthy livestock before slaughtering, avoid infections, and promote animal growth. Such indiscriminate use of antibiotics will lead to presence of resistance pathogens in faeces and in animal tissues, which may be easily widespread to other creatures [20]. This may strongly explain the high prevalence of MDR amongst our local suspected E.coli O157 collected from food products and cattle faeces. Katsuda et al., 2009 [49] reported that antimicrobial resistance has been increasingly identified in animal pathogens.
It is noteworthy in this study that the most frequent source for MDR E.coli O157 in Al-Sharkia, Egypt that may cause environment and public health risk, was the CF samples. Where, eight out of the nine (89%) CF samples that were suspect to have E.coli O157 isolates had the highest MAR index (ranged between 0.23- 0.62), and were considered as MDR. Our local results suggest that CF samples are the leading cause of pathogenic E. coli O157 contamination. Wang et al. [50] pointed out that the bovine faeces are a potential source for the spread of STEC strains to the human food chain, as well as to the environment. The effective control of STEC strains mandate correct handling and usage of bovine (cattle) faeces to decrease the risk of contamination of the environment and food by this pathogen. Even more so, by the presence of many reports for long-term survival of both E. coli O157 and non-O157 in faeces of dairy and beef cattle [50].
Two isolates out of our local suspected E. coli O157, were both obtained from CF samples (CF8, and CF13), and confirmed as E.coli O157:H7 by serological test, as they gave strong agglutination with O157 antigen and H7 antigen. The agglutination test appeared specific for the diagnosis of E. coli O157:H7, these results are in agreement with, [51, 52], who used latex agglutination test for serotyping of E. coli O157:H7. It is a rapid, reliable test that is easy to perform.
Our results revealed that, the prevalence of E.coli O157:H7 in Al-Sharkia, Egypt (two out of 30 samples) is equal to 6.7%. This ratio is similar to results that were obtained from sheep faeces in another Egypt governorate (Giza) [53], which is close to the prevalence of E. coli O157 in sheep faeces at Pastures in Scotland (6.5%) [54], and 7.1% in Northern Spain [55]. Nonetheless, our ratio for prevalence of E.coli O157 in Al-sharkia, Egypt is clearly higher than that recorded in South Yorkshire, UK (1–2%) and Spain (2.6%) by [23, 56], respectively, and also higher than in Great Britain, Spain, and Switzerland as recorded [57,58,59], respectively.
Pathogenicity of E.coli O157:H7 is associated with different virulence factors, production of Shiga toxins (stx1 and stx2) being the most potent one [60, 61] . Louise et al. [62] reported that stx1 and stx2 cause different grades and types of tissue damage, and stx2 is more toxic than stx1 to renal endothelial cells in mammals. Among our two tested MDR E.coli O157:H7 isolates, CF8 was found to carry stx2 gene, and CF13 carry stx1 and stx2 genes; this is according to molecular analysis by multiplex PCR.
Particularly, as E. coli O157 can survive in the environment for more than 10 months, as reported by [63], humans and animals could be at risk of infection long after an environment is initially contaminated. Very low concentrations of STEC O157:H7 can be a source of outbreaks or sporadic infection, as stated by [64]; whom have estimated that the infectious dose for this STEC pathogen to be under 100 colony-forming unit/ patient. Others, estimated the infectious dose to be just 10 bacteria [65]. Occurrence of STEC O157:H7 in local cattle faeces suggest higher direct, or indirect transmission of STEC O157 may lead to outbreaks, due to work of the large population et al.-Sharkia in agriculture and cattle breeding. Direct transmission of STEC O157:H7 via the fecal–oral route, may be from calves to children, manure exposure in the field, or dirty hands. While, indirect transmission could be through: slaughtering, the natural progression of the food chain, or from contaminated soil, water, or vegetables with cattle faeces.
Shiga toxin-producing E. coli (STEC) has been deeply involved in foodborne, waterborne and airborne outbreaks in several studies all over the world [66], and have demonstrated that the outbreak of E. coli O157:H7, caused the infection for 20 campers out of 337 at scout camp on May 2000, it was due to an environmental source. Investigations do not suggest that it was a food vehicle, as they did not find any E.coli isolates in food or drinking water. However, investigations supported that the transmission of E. coli O157:H7 was due to exposure to the contaminated environment by hands, either straightly from hand to mouth or via food. They found that the food in studied cases worked as an inert vehicle for E. coli O157:H7 transmission, not as a growth medium (source of infection) for the bacterial pathogen. The contamination of the environment may be due to the fact that the camp took place on an agricultural showground used for sheep grazing. Additionally, the location of the camp had been fully contaminated with sheep faeces.
The isolated toxigenic O157 E.coli strains carrying stx1 and stx2 genes mainly showed MDR towards numerous antibiotics that are medically important, as they commonly used in human and animal treatment. There is a high possibility that these resistant strains can endure the environment for a long time and, then, eventually transmit these resistance determinants to other environmental bacteria [67].
Such transfer of resistance determinants could fuel the spread of MDR bacteria, this could have grave implications on the health of humans and animals; thus, increasing the burden of disease in the community. Therefore, there is a great need for urgent policy formulations on the prudent use of antimicrobials in both human and veterinary medicine, because failure in this regard could spell doom in the near future [68].
There are high recommendations from different authorities worldwide for testing all ground meat/poultry samples for STEC O-serotypes, as these organisms are shed in faeces, and will lead to environmental contamination for an extended period of time as the USDA Food Safety and Inspection Service states.
Conclusions
The importance of recent study is to monitor the prevalence of suspected E.coli O157 in Zagazig City et al.-Sharkia, Egypt by 15.4% in grounding beef, as hamburger and cattle faeces samples are of epidemiological significance. Furthermore, there is a high prevalence of MDR (80%) determinants in suspected E.coli O157. Accordingly, this will lead to the emergence of superbugs of clinical relevance due to the ease of transfer of resistance genes to other pathogens. In addition, two isolates (6.7%) recovered from cattle faeces were carried the Shiga toxin genes stx1 and/or stx2, which suggests that the farm environment may serve as a channel for the dissemination of antimicrobial-resistant STEC strains to other niches, including the food chain. Risk assessment is needed to protect human, animal, and environmental health. Furthermore, animal faeces should be treated before discharge, continuous monitoring should be considered, and surveillance for environment, livestock, and food product should be strengthened.
Methods
Samples collection and preparation
A total of sixty-five samples were collected in the period between Jan. 2021 to March 2021. Out of this, 35 samples (15 chicken meat, 10 luncheon, and 10 hamburger) were collected from diverse markets and butcher’s shops in Zagazig City, AL-Sharkia Governorate- Egypt. While, the remaining 30 samples, were cattle’s faecal samples collected from different house farms, and the Veterinary Medicine in Zagazig University at Zagazig City, AL-Sharkia Governorate- Egypt.
Following the same procedure as [20], all samples were packaged in sterilized polyethylene bags and transferred directly within 1–2 h, in a cold box at 4 °C to the laboratory. Each bag was labeled with the source name and number.
At the laboratory, each sample underwent the following process. Twenty-five grams of the sample was transferred to a sterile flask using sterile spatula, and 225 ml of Tryptone Soya Broth (TSB) was added. Homogenization took place in a clean blender at 2000 r.p.m for 1–2 min. Then, the homogenized sample was transferred into a sterilized flask and incubated at 37 ± 2 °C for 18-24 h.
Enumeration of faecal coliform bacteria in samples, isolation, and purification of E. coli O157
To enumerate the low concentration of the total fecal coliform bacteria FCB (including undetectable E. coli) in each sample alone, the three-tubes most probable number (MPN) technique was performed as in the U.S. Food and Drug Administration Bacteriological Analytical Manual, which is well described by [69]. The MPN is a statical assay based on probability theory, and a multi-step assay consisting of three-steps; presumptive test, confirmed test, and completed test [70] based on lactose fermentation. Briefly, at the first screening test (presumptive test), serial dilutions (0.1,0.01,0.001) of each sample were prepared. Then, each dilution was inoculated into three test tubes containing 10 ml of MacConkey broth (MB) (Oxoid, UK), and Durham’s tube. After overnight incubation at 37 ± 2 °C, positive samples the presence of FCB produced gas or changed MB colour from red to yellow. The number of tubes giving a positive reaction (in each dilution) were compared to a standard chart, and the number of FCB present in each sample was determined by using the MPN index (just for the 35 food samples). The Confidence 95% intervals were used to represent the probability of the results that cover the actual concentration before inoculating the tubes, which was at least 95%. Loopful from each positive tube was re-streaked on MacConkey Agar medium (Oxoid, UK) to get pure colonies of FCB isolates.
Then, the confirmatory test was performed by picking up pure colonies of FCB isolates and streaked on eosin methylene blue (EMB) agar medium (oxoid CM69, UK), in order to isolate and differentiate E. coli colonies from other FCB isolates. In this test, a loopful from each positive tube was streaked on eosin methylene blue agar medium (EMB) (oxoid CM69, UK). After an incubation period 18-24 h at 37 ± 2 °C, colonies with metallic sheen appearance were picked up and purified as E. coli isolates for a completed test.
Purified E. coli colonies obtained from previous assays were separately streaked onto selective differential sorbitol MacConkey agar media with cefixime- telurite supplement (CT-SMAC) (Oxoid, UK), which has a selective supplement (cefixime- telurite) for specific E. coli O157 isolation. After the incubation period, E. coli serogroup O157 colonies had a pale (colourless) appearance due to their inability to ferment the sorbitol [71]. These colonies were picked up, labeled with the source of sample and number, and underwent further investigations in this study.
Antibiotic susceptibility for E. coli O157 isolates
All presumptive purified E. coli O157:H7 were tested for their susceptibilities to the selected antibiotics by standard Kirby-Bauer disc diffusion method [72]. Thirteen antibiotic discs (Oxoid, Uk) were chosen as they are commonly used in treatment infections in local hospitals and recommended worldwide [48, 73]. Additionally, they represent different groups of antibiotics with different modes of action against E. coli isolates. The 13 chosen antibiotics include the following; cefoxitin (FOX, 30 µg), cefepime (FEP, 30 µg), ceftazidime (CAZ, 30 µg), ceftriaxon (CRO, 30 µg), amoxicillin/clavulanic acid (AMC, 30/10 µg), and imipenem (IPM, 10 µg) belonging to β-lactam antibiotics, piperacillin (PRL, 10 µg), gentamicin (CN, 10 µg), amikacin (AK, 30 µg),trimethoprim-sulfamethoxazole (SXT, 1.25/23.75 µg), ciprofloxacin (CIP, 5 µg), erythromycin (E, 15 µg), and polymixin (Pb30 µg). The test was performed as illustrated by [74] and the results were interpreted as resistant (R), intermediate (I) or sensitive (S), by measuring the diameter of the inhibition zone (DIZ) in mm around each disc in accordance with the recommendation of Clinical and Laboratory Standard Institute (CLSI, 2018). The antibiogram pattern of each isolate against the thirteen tested antibiotics was obtained and the multiple antibiotic resistance (MAR) index was calculated as follows:
Isolates with high MAR index were recorded as multidrug-resistant (MDR) isolates, especially if they showed resistance to more than two antibiotics from different groups [20].
Serological identification of MDR E. coli O157 isolates
According to the antibiogram pattern, the multidrug- resistant (MDR) isolates were serologically identified by slide agglutination according to [75] by using rapid monovalent O157and H7 antisera (DENKA SEIKEN Co., Japan). The isolate showed agglutination in O157 antiserum and H7 antiserum was confirmed as E. coli O157:H7, and they were selected to approve their carrying of Shiga toxin genes.
Amplification of Shiga toxin stx1 and stx2 virulence genes
DNA from E. coli O157:H7 isolates gave positive reaction for agglutination test with both anti-O157 and anti-H7 antibodies, and was extracted by using QIAamp DNA Mini kit (Qiagen, Düsseldorf, Germany, GmbH). Next, the isolates were subjected to molecular analysis for the presence of Shiga toxins (stx1 and stx2) as a potential virulence gene in E. coli O157:H7, by using multiplex PCR. The primer sequences were designed by [76], as illustrated in Table 5.
PCR was carried out in a 50 µL reaction containing 25 µL of EmeraldAmp GT PCR mastermix (2 × premix)(Takara, Japan), 1 µL of each primer (20 pmol), 13 µL of water, and 8 µL of DNA template. The reactions were performed in an Applied Biosystems 2720 thermal cycler (Biometra, Göttingen, Germany). The products of PCR were separated by electrophoresis on 1.5% (Wt/Vol) agarose gel (Applichem GmbH, Germany) in 1 × TBE buffer (10.78 g/L Tris buffer, 5.5 g/L Boric acid, 0.82 g/L EDTA, pH 8.3) at room temperature using gradients of 5 V/cm. For gel analysis, 20 µL of the products were loaded in each gel slot. A Gelpilot 100 bp Ladder (Qiagen GmbH, Germany) was used to determine the fragment sizes. The gel was photographed by a gel documentation system (Alpha Innotech, Biometra) and the data was analyzed using associated software [77].
Polymerase Chain Reaction (PCR) for amplification of 16S rRNA gene
PCR amplification was performed to confirm the identity of E. coli O157:H7 using universal 1492r primer [78], as in Table 5.
The extracted bacterial genome must be diluted before PCR performance by adding 495 μl of nuclease free water into 1.5 ml micro centrifuge tube and 5 μl of the supernatant from the PrepMan ultra extracted sample was added to get 1: 100 dilution; then, vortexed and mixed well. Thermo-scientific was used to amplify the first 500 base pairs of the 16S rRNA bacterial gene in the sample. PCR set up was performed by adding the following components: 12.5 μl of Dream Taq Green PCR Master Mix (2X) with Cat. No. K1081, 0.5 μl of Forward Primer (10 pmol), 0.5 μl of Reverse Primer (10 pmol), 10.5 μl Nuclease Free Water and 1.0 μl DNA Template; the tube was closed tightly and placed in the thermal cycler. PCR reaction conditions were initial denaturation at 95 °C for 4 min, 40 cycles at 95 °C for 30 s, 60 °C for 30 s and 72 °C for 40 s. Final extension at 72 °C for 10 min was done. The PCR product was run and visualized by loading 10 μl of the PCR product per lane on 1.8% agarose gel that stained by ethidium bromide against 100 bp DNA ladder as a marker using 1X TAE as a running buffer, and photographed by gel documentation system to identify its size.
Purification of PCR product
PCR products were purified according to the manufacturing procedure of Gene JET PCR Purification Kit (Thermo Scientific, Cat. No. K0701) as per the following steps: five microliters of the PCR reaction product was mixed with 2 μl of GeneJET reagent for a total of 7 μl of the reaction volume, and incubated at 37 °C for 15 min to degrade the remaining primers and nucleotides. Then, the sample was incubated at 80 °C for 15 min to inhibit the reagent reaction. The purified PCR product stored at -20 °C for sequencing.
DNA sequencing for the amplified gene
PCR products were sequenced using the standard Sanger method on ABI 3730XL DNA Sequencer at Macrogen sequencing services (Macrogen, Seol, South Korea for forward and reverse sequencing.
Sequence alignment and phylogenetic analysis
Pair wise and multiple DNA sequence alignment were carried out using CLUSTALW multiple sequence alignment programme version (7.0.9) (http://www.ebi/ac.uk/clustalw) [79]. Bootstrap neighbor joining tree was generated using MEGA version 3.1[80] from CLUSTALW alignment. Comparison with sequences in the Gene Bank database was achieved in BLASTN searches at the National center for Biotechnology Information site (htto:// ncbi. nlm.nih.gov).
The Gene-Bank nucleotide sequence accession numbers for partial sequences of 16S rRNA gene were generated in this study using DDBJ Center [81].
Statistical analysis
All statistics with a probability value of ≤ 0.05 were considered significant. The bacterial counts were normalized by transforming the data using log10. The MPN estimated by selecting the dilutions of positive tubes, and all the dilutions between them by using [82] formula;
where the sum is over the selected dilutions, and ∑gj = the number of positive tubes in the selected dilutions, ∑tjmj = the grams of sample in all tubes in the selected dilutions. The confidence 95% limits for any dilution test calculated by estimating the standard error of log10 (MPN) by the method of [83].
The mean value (from the three replicates) was plotted against time by linear regression analysis using cultural detection of E. coli. The slope of the line was calculated to obtain a growth rate for the organism (the unit for growth rate is log10 CFU).
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on request. The sequences of two strains analyzed were deposited in the National Library of Medicine, National Center for Biology Information (NCBI), GenBank nucleotide sequence database. The accession numbers assigned as LC666912 (https://www.ncbi.nlm.nih.gov/nuccore/LC666912) and LC666913 (https://www.ncbi.nlm.nih.gov/nuccore/LC666913).
Abbreviations
- E. coli :
-
Escherichia coli
- STEC:
-
Shiga toxin-producing Escherichia coli
- stx:
-
Shiga toxins
- MDR:
-
Multidrug-resistant
- MAR:
-
Multiple antibiotic resistance
- MPN:
-
Most probable number
- HUS:
-
Hemolytic-uremic syndrome
- HC:
-
Hemorrhagic colitis
- FCB:
-
Fecal coliform bacteria
- C:
-
chicken meat samples
- L:
-
luncheon samples
- H:
-
Hamburger samples
- CF:
-
Cattle faeces samples
- CT-SMAC:
-
Sorbitol MacConkey agar media with Cefixime-Telurite supplement
- MB:
-
MacConkey broth media
- EMB:
-
Eosin methylene blue medium
- TSB:
-
Tryptone Soya Broth media
- TBE:
-
Tris Borate EDTA buffer
- R:
-
Resistant
- I:
-
Intermediate
- S:
-
Sensitive
- DIZ:
-
Diameter of inhibition zone
- FOX:
-
Cefoxitin
- FEP:
-
Cefepime
- CAZ:
-
Ceftazidime
- CRO:
-
Ceftriaxon
- AMC:
-
Amoxicillin/clavulanic acid
- PRL:
-
Piperacillin
- IPM:
-
Imipenem
- AK:
-
Amikacin
- CN:
-
Gentamicin
- SXT:
-
Trimethoprim/sulfamethoxazole
- PB:
-
Polymixin
- CIP:
-
Ciprofloxacin
- E:
-
Erythromycin
References
Doyle MP. Escherichia coli O157: H7 and its significance in foods. Int J Food Microbiol. 1991;12(4):289–301.
Persad AK, Lejeune JT. Animal reservoirs of Shiga toxin-producing Escherichia coli. Enterohemorrhagic Escherichia coli and Other Shiga Toxin-Producing E coli 2015:211–230.
Mukherjee A, Cho S, Scheftel J, Jawahir S, Smith K, Diez-Gonzalez F. Soil survival of Escherichia coli O157: H7 acquired by a child from garden soil recently fertilized with cattle manure. J Appl Microbiol. 2006;101(2):429–36.
Smith JL, Fratamico PM, Gunther NW IV. Shiga toxin-producing Escherichia coli. Adv Appl Microbiol. 2014;86:145–97.
CfD Control. Prevention: Outbreaks of Escherichia coli O157: H7 infections among children associated with farm visits–Pennsylvania and Washington, 2000. MMWR Morbidity Mortality Weekly Report. 2001;50(15):293–7.
Boerlin P, McEwen SA, Boerlin-Petzold F, Wilson JB, Johnson RP, Gyles CL. Associations between virulence factors of Shiga toxin-producing Escherichia coli and disease in humans. J Clin Microbiol. 1999;37(3):497–503.
Griffin PJIotgt: Escherichia coli O157: H7 and other enterohemorrhagic Escherichia coli. 1995;739–761.
Law D. Virulence factors of Escherichia coli O157 and other Shiga toxin‐producing E. coli. J Appl Microbiol. 2000;88(5):729–745.
Scheutz F, Teel LD, Beutin L, Piérard D, Buvens G, Karch H, Mellmann A, Caprioli A, Tozzoli R, Morabito S, Strockbine NA. Multicenter evaluation of a sequence-based protocol for subtyping Shiga toxins and standardizing Stx nomenclature. J Clin Microbiol. 2012;50(9):2951–63.
Melton-Celsa A: Shiga toxin (Stx) classification, structure, and function. Microbiol Spectr 2: EHEC. In.: EHEC-0024–2013. https://doi.org/10.1128/microbiolspec.EHEC-0024-2013. 2014.
Etcheverria AI, Padola NL. Shiga toxin-producing Escherichia coli: factors involved in virulence and cattle colonization. Virulence. 2013;4(5):366–72.
Lindgren S, Melton A, O’Brien A. Virulence of enterohemorrhagic Escherichia coli O91: H21 clinical isolates in an orally infected mouse model. Infect Immun. 1993;61(9):3832–42.
Smith DL, Rooks DJ, Fogg P, Darby AC, Thomson NR, McCarthy AJ, Allison HE. Comparative genomics of Shiga toxin encoding bacteriophages. BMC Genomics. 2012;13(1):1–10.
Melton-Celsa AR. Shiga toxin (Stx) classification, structure, and function. Microbiol Spectrum. 2014;2(4):2.4. 06.
Rivas M, Chinen I, Guth BE: Enterohemorrhagic (Shiga toxin-producing) Escherichia coli. In: Escherichia coli in the Americas. Springer; 2016;97–123.
Armstrong GL, Hollingsworth J, Morris Jr JG. Emerging foodborne pathogens: Escherichia coli O157: H7 as a model of entry of a new pathogen into the food supply of the developed world. 1996.
Johnson RP, Clarke RC, Wilson JB, Read SC, Rahn K, Renwick SA, Sandhu KA, Alves D, Karmali MA, Lior H. Growing concerns and recent outbreaks involving non-O157: H7 serotypes of verotoxigenic Escherichia coli. J Food Prot. 1996;59(10):1112–22.
Olsen SJ, Miller G, Breuer T, Kennedy M, Higgins C, Walford J, McKee G, Fox K, Bibb W, Mead P. A waterborne outbreak of Escherichia coli O157: H7 infections and hemolytic uremic syndrome: implications for rural water systems. Emerg Infect Dis. 2002;8(4):370.
Da Costa PM, Loureiro L, Matos AJ. Transfer of multidrug-resistant bacteria between intermingled ecological niches: the interface between humans, animals and the environment. Int J Environ Res Public Health. 2013;10(1):278–94.
Araby E, Nada HG, El-Nour A, Salwa A, Hammad A. Detection of tetracycline and streptomycin in beef tissues using Charm II, isolation of relevant resistant bacteria and control their resistance by gamma radiation. BMC Microbiol. 2020;20(1):1-11
Rolain J-M. Food and human gut as reservoirs of transferable antibiotic resistance encoding genes. Front Microbiol. 2013;4:173.
Majowicz SE, Scallan E, Jones-Bitton A, Sargeant JM, Stapleton J, Angulo FJ, Yeung DH, Kirk MD. Global incidence of human Shiga toxin–producing Escherichia coli infections and deaths: a systematic review and knowledge synthesis. Foodborne Pathog Dis. 2014;11(6):447–55.
Chapman P, Malo AC, Ellin M, Ashton R, Harkin MJIjofm: Escherichia coli O157 in cattle and sheep at slaughter, on beef and lamb carcasses and in raw beef and lamb products in South Yorkshire, UK. 2001;64(1–2):139–150.
Fegan N, Higgs G, Vanderlinde P, Desmarchelier P. Enumeration of Escherichia coli O157 in cattle faeces using most probable number technique and automated immunomagnetic separation. Letters Appl Microbiol. 2004;38(1):56–9.
Beatty ME, Bopp CA, Wells JG, Greene KD, Puhr ND. Mintz EDJEid: Enterotoxin-producing Escherichia coli O169: H41. United States. 2004;10(3):518.
Devasia RA, Jones TF, Ward J, Stafford L, Hardin H, Bopp C, Beatty M, Mintz E, Schaffner W. Endemically acquired foodborne outbreak of enterotoxin-producing Escherichia coli serotype O169: H41. Am J Med. 2006;119(2):168. e167–168. e110.
Yang X, Bai X, Zhang J, Sun H, Fu S, Fan R, He X, Scheutz F, Matussek A, Xiong Y. Escherichia coli strains producing a novel Shiga toxin 2 subtype circulate in China. Int J Med Microbiol. 2020;310(1):151377.
Fatoki T, Ugboko H, Nwinyi O, Adetuyi O, Ibraheem O, Enibukun J. Computational Study of 16S rRNA of Microbe Cluster Implicated in Diarrhoeal: Phylogeny, Docking, and Dynamics; 2021.
Naylor SW, Gally DL, Low JC. Enterohaemorrhagic E. coli in veterinary medicine. Int J Medical Microbiol. 2005;295(6–7):419–441.
Akashi S, Joh K, Mori T, Tsuji A, Ito H, Hoshi H, Hayakawa T, Ihara J, Abe T, Hatori M, Nakamura T. A severe outbreak of haemorrhagic colitis and haemolytic uraemic syndrome associated withEscherichia coli 0157: H7 in Japan. Eur J Pediatr. 1994;153(9):650–655.
Licence K, Oates KR, Synge BA, Reid TM. An outbreak of E. coli O157 infection with evidence of spread from animals to man through contamination of a private water supply. Epidemiol Infect. 2001;126(1):135-8.
Crump JA, Sulka AC, Langer AJ, Schaben C, Crielly AS, Gage R, Baysinger M, Moll M, Withers G, Toney DM, Hunter SB. An outbreak of Escherichia coli O157: H7 infections among visitors to a dairy farm. New England J Med. 2002;347(8):555-60.
Bell BP, Goldoft M, Griffin PM, Davis MA, Gordon DC, Tarr PI, Bartleson CA, Lewis JH, Barrett TJ, Wells JG, Baron R. A multistate outbreak of escherichia coli o157: h7—associated bloody diarrhea and hemolytic uremic syndrome from hamburgers: the washington experience. Jama. 1994;272(17):1349-53.
Karch H, Bielaszewska M, Bitzan M, Schmidt H. Epidemiology and diagnosis of Shiga toxin-producing Escherichia coli infections. Diagnostic Microbiol Infect Dis. 1999;34(3):229-43.
Leininger DJ, Roberson JR, Elvinger F. Use of eosin methylene blue agar to differentiate Escherichia coli from other gram-negative mastitis pathogens. J Veterinary Diagnostic Investigation. 2001;13(3):273-5.
March SB, Ratnam SA. Sorbitol-MacConkey medium for detection of Escherichia coli O157: H7 associated with hemorrhagic colitis. J Clin Microbiol. 1986;23(5):869-72.
Chapman P, Siddons C, Harkin MJVR: Sheep as a potential source of verocytotoxin-producing Escherichia coli O157.[Correspondence]. 1996.
Galland JC, Hyatt DR, Crupper SS, Acheson DWJA, Microbiology E: Prevalence, antibiotic susceptibility, and diversity of Escherichia coli O157: H7 isolates from a longitudinal study of beef cattle feedlots. 2001;67(4):1619–1627.
Mora A, Blanco JE, Blanco M, Alonso MP, Dhabi G, Echeita A, González EA, Bernárdez MI, Blanco J. Antimicrobial resistance of Shiga toxin (verotoxin)-producing Escherichia coli O157: H7 and non-O157 strains isolated from humans, cattle, sheep and food in Spain. Res Microbiol. 2005;156(7):793–806.
Ranjbar R, Safarpoor Dehkordi F, Sakhaei Shahreza MH, Rahimi E. Prevalence, identification of virulence factors, O-serogroups and antibiotic resistance properties of Shiga-toxin producing Escherichia coli strains isolated from raw milk and traditional dairy products. Antimicrob Resistance Infect Control. 2018;7(1):1–1.
Fard A, Bokaeian M, Qureishi MJE-EMHJ, 14 , -,: Frequency of Escherichia coli O157: H7 in children with diarrhoea in Zahedan, Islamic Republic of Iran. 2008.
Ababu A, Endashaw D, Fesseha H. Isolation and antimicrobial susceptibility profile of Escherichia coli O157: H7 from raw milk of dairy cattle in Holeta district. Central Ethiopia Int J Microbiol. 2020;2020:1–8.
Kim JY, Kim SH, Kwon NH, Bae WK, Lim JY, Koo HC, Kim JM, Noh KM, Jung WK, Park KT, Park YH. Isolation and identification of Escherichia coli O157: H7 using different detection methods and molecular determination by multiplex PCR and RAPD. J Vet Sci. 2005;6(1):7–19.
Altalhi AD, Gherbawy YA, Hassan SA. Antibiotic resistance in Escherichia coli isolated from retail raw chicken meat in Taif, Saudi Arabia. Foodborne pathogens and disease. 2010;7(3):281-5.
Akond MA, Alam S, Hassan SM, Shirin M. Antibiotic resistance of Escherichia coli isolated from poultry and poultry environment of Bangladesh. Internet J Food Safety. 2009;11:19-23.
Collignon PC, Conly JM, Andremont A, McEwen SA, Aidara-Kane A, World Health Organization Advisory Group, Bogotá Meeting on Integrated Surveillance of Antimicrobial Resistance (WHO-AGISAR), Agerso Y, Andremont A, Collignon P, Conly J, Dang Ninh T. World Health Organization ranking of antimicrobials according to their importance in human medicine: a critical step for developing risk management strategies to control antimicrobial resistance from food animal production. Clin Infect Dis. 2016;63(8):1087-93.
Organization WH: WHO global principles for the containment of antimicrobial resistance in animals intended for food: report of a WHO consultation with the participation of the Food and Agriculture Organization of the United Nations and the Office International des Epizooties, Geneva, Switzerland 5-9 June 2000. In.: World Health Organization; 2000
Schroeder CM, Zhao C, DebRoy C, Torcolini J, Zhao S, White DG, Wagner DD, McDermott PF, Walker RD, Meng JJA et al. Antimicrobial resistance of Escherichia coli O157 isolated from humans, cattle, swine, and food. Appl Environment Microbiol. 2002;68(2):576–581.
Katsuda K, Kohmoto M, Mikami O, Uchida I. Antimicrobial resistance and genetic characterization of fluoroquinolone-resistant Mannheimia haemolytica isolates from cattle with bovine pneumonia. Vet Microbiol. 2009;139(1–2):74–9.
Wang G, Zhao T, Doyle MP. Fate of enterohemorrhagic Escherichia coli O157: H7 in bovine feces. Appl Environ Microbiol. 1996;62(7):2567–70.
Al–Dawmy FA, Yousif AA. Prevalence of E. coli O157: H7 in intestinal and urinary tract infection in children. Int J Adv Res. 2013;1(8):111-20.
Karmali MA, Petric M, Bielaszewska M. Evaluation of a microplate latex agglutination method (Verotox-F assay) for detecting and characterizing verotoxins (Shiga toxins) in Escherichia coli. J Clin Microbiol. 1999;37(2):396-9.
Kamel M, El-Hassan DG, El-Sayed A. Epidemiological studies on Escherichia coli O157: H7 in Egyptian sheep. Trop Anim Health Product. 2015;47:1161-7.
Ogden ID, MacRae M, Strachan NJ. Concentration and prevalence of Escherichia coli O157 in sheep faeces at pasture in Scotland. J Appl Microbiol. 2005;98(3):646–51.
Oporto B, Esteban J, Aduriz G, Juste R, Hurtado A: Escherichia coli O157: H7 and non-O157 Shiga toxin-producing E. coli in healthy cattle, sheep and swine herds in northern Spain. Zoonoses Public Health. 2008;55(2):73–81.
Rey J, Blanco JE, Blanco M, Mora A, Dahbi G, Alonso JM, Hermoso M, Hermoso J, Alonso MP, Usera MA, González EA. Serotypes, phage types and virulence genes of Shiga-producing Escherichia coli isolated from sheep in Spain. Veterinary Microbiol. 2003;94(1):47–56.
Paiba GA, Pascoe SJ, Wilesmith JW, Kidd SA, Byrne C, Ryan JB, Smith RP, McLaren IM, Jones YE, Chappell SA, Gibbens JC. Faecal carriage of verocytotoxin-producing Escherichia coli0157 in cattle and sheep at slaughter in Great Britain. Veterinary Record. 2002;150(19):593–8.
Blanco M, Blanco JE, Mora A, Dahbi G, Alonso MP, González EA, Bernárdez MI, Blanco J. Serotypes, Virulence Genes, and Intimin Types ofShiga Toxin (Verotoxin)-Producing Escherichia coli Isolatesfrom Cattle in Spain and Identification of a New Intimin VariantGene (eae-ξ). J Clin Microbiol. 2004;42(2):645-51.
Zweifel C, Kaufmann M, Blanco J, Stephan R. Significance of Escherichia coli O157 in sheep at slaughter in Switzerland. Schweizer Archiv fur Tierheilkunde. 2006;148(6):289-95.
Lim KG, Kang MI, Kim SK, Nam KW, Park HJ, Park JR, Cho KO, Lee BJ. Identification and characterization of Shiga toxin-producing Escherichia coli isolated from diarrhea in calves. Korean J Veterinary Res. 2006;46(2):135–42.
Nataro JP, Kaper JB. Diarrheagenic escherichia coli. Clin Microbiol Reviews. 1998;11(1):142–201.
Louise CB, Kaye SA, Boyd B, Lingwood CA, Obrig TG. Shiga toxin-associated hemolytic uremic syndrome: effect of sodium butyrate on sensitivity of human umbilical vein endothelial cells to Shiga toxin. Infect Immunity. 1995;63(7):2766–9.
Varma JK, Greene KD, Reller ME, DeLong SM, Trottier J, Nowicki SF, DiOrio M, Koch EM, Bannerman TL, York ST, Lambert-Fair MA. An outbreak of Escherichia coli O157 infection following exposure to a contaminated building. Jama. 2003;290(20):2709–12.
Tilden Jr J, Young W, McNamara AM, Custer C, Boesel B, Lambert-Fair MA, Majkowski J, Vugia D, Werner SB, Hollingsworth J, Morris Jr JG. A new route of transmission for Escherichia coli: infection from dry fermented salami. Am J Public Health. 1996;86(8_Pt_1):1142-5.
Tarr PI. Escherichia coli O157: H7: clinical, diagnostic, and epidemiological aspects of human infection. Clin Infect Dis. 1995;20(1):1–8.
Howie H, Mukerjee A, Cowden J, Leith J, Reid T. Investigation of an outbreak of Escherichia coli O157 infection caused by environmental exposure at a scout camp. Epidemiol Infect. 2003;131(3):1063–9.
Metlay JP, Powers JH, Dudley MN, Christiansen K, Finch RG. Antimicrobial drug resistance, regulation, and research. Emerg Infect Dis. 2006;12(2):183.
Iweriebor BC, Iwu CJ, Obi LC, Nwodo UU, Okoh AI. Multiple antibiotic resistances among Shiga toxin producing Escherichia coli O157 in feces of dairy cattle farms in Eastern Cape of South Africa. BMC Microbiol. 2015;15(1):1–9.
Sutton S. The most probable number method and its uses in enumeration, qualification, and validation. J Validation Technol. 2010;16(3):35–8.
Phyo SS, Yu SS, Saing KM. Bacteriological examination of bottled drinking water by MPN method. Haya: Saudi J Life Sci. 2019;4(7):227-32.
Szalanski AL, Owens CB, McKay T, Steelman CD. Detection of Campylobacter and Escherichia coli O157: H7 from filth flies by polymerase chain reaction. Med Vet Entomol. 2004;18(3):241–6.
Bauer AW Antibiotic susceptibility testing by a standardized single disc method. Am J Clin Pathol. 1966;45:149-158.
George DF, Gbedema SY, Agyare C, Adu F, Boamah VE, Tawiah AA, Saana SB. Antibiotic resistance patterns of Escherichia coli isolates from hospitals in Kumasi, Ghana. Int Scholarly Res Notices. 2012;2012.
Nada H, Hagag S, El-Tablawy S. Expression of dnak, groES and cps Genes in Irradiated Klebsiella pneumoniae Strains Isolated from UTI Egyptian Patients. Egypt J Radiation Sci Appl. 2018;31(2):185–93.
Kok T, Worswich D, Gowans E. Some serological techniques for microbial and viral infections. Practical Medical Microbiology (Collee, J.; Fraser, A.; Marmion, B. and Simmons, A., eds.), 14th ed., Edinburgh, Churchill Livingstone, UK. 1996:179-204.
Baron L, Santaniello A, Fontanella M, Lagos K, Fioretti A, Menna L. Presence of Shiga toxin-producing Escherichia coli O157: H7 in living layer hens. Lett Appl Microbiol. 2006;43(3):293–5.
Sambrook J, Fritsch EF, Maniatis T. Molecular cloning: a laboratory manual (No. Ed. 2). Cold spring harbor laboratory press. 1989.
Baron EJ, Miller JM, Weinstein MP, Richter SS, Gilligan PH, Thomson RB Jr, Bourbeau P, Carroll KC, Kehl SC, Dunne WM. A guide to utilization of the microbiology laboratory for diagnosis of infectious diseases: 2013 recommendations by the Infectious Diseases Society of America (IDSA) and the American Society for Microbiology (ASM) a. Clin Infect Dis. 2013;57(4):e22–121.
Thompson JD, Higgins DG, Gibson TJ. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994;22(22):4673–80.
Kumar S, Dhingra A, Daniell H. Stable transformation of the cotton plastid genome and maternal inheritance of transgenes. Plant Molecular Biol. 2004;56(2):203–16.
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215(3):403–10.
Thomas HA. Bacterial densities from fermentation tube tests. J Am Water Works Assoc. 1942;34(4):572–6.
Haldane JB. Sampling errors in the determination of bacterial or virus density by the dilution method. Epidemiol Infect. 1939;39(3):289–93.
Acknowledgements
The authors would like to express their special thanks to Retaj S. Elshourbagi (PharmD Program, Faculty of Pharmacy, Galala University) for her participation in final proofreading and language editing of this manuscript.
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB). This research did not receive any fund.
Author information
Authors and Affiliations
Contributions
A.A. and G.D. designed and supervised all steps in this study, in addition, revised the final manuscript. A.S.T collected samples, performed all microbiological assays, and identified the strains. H.G.N. conducted the tests with data analysis of the results, gave a point of view to discussion, and wrote the draft with the final manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Nada, H.G., El-Tahan, A.S., El-Didamony, G. et al. Detection of multidrug-resistant Shiga toxin-producing Escherichia coli in some food products and cattle faeces in Al-Sharkia, Egypt: one health menace. BMC Microbiol 23, 127 (2023). https://doi.org/10.1186/s12866-023-02873-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12866-023-02873-2