
Abstract
Background
B-box (BBX) family is a class of zinc finger transcription factors (TFs) that play essential roles in regulating plant growth, development, as well as abiotic stress. However, no systematic analysis of BBX genes has yet been conducted in alfalfa (Medica go sativa L.), and their functions have not been elucidated up to now.
Results
In this study, 28 MsBBX genes were identified from the alfalfa genome, which were clustered into 4 subfamilies according to an evolutionary tree of BBX proteins. Exon-intron structure and conserved motif analysis reflected the evolutionary conservation of MsBBXs in alfalfa. Collinearity analysis showed that segmental duplication promoted the expansion of the MsBBX family. Analysis of cis-regulatory elements suggested that the MsBBX genes possessed many growth/development-, light-, phytohormone-, and abiotic stress-related elements. MsBBX genes were differentially expressed in leaves, flowers, pre-elongated stems, elongated stems, roots and nodules, and most MsBBXs were remarkably induced by drought, salt and various plant growth regulators (ABA, JA, and SA). Further functional verification demonstrated that overexpressing of the MsBBX11 gene clearly promoted salt tolerance in transgenic Arabidopsis by regulating growth and physiological processes of seedlings.
Conclusions
This research provides insights into further functional research and regulatory mechanisms of MsBBX family genes under abiotic stress of alfalfa.
Similar content being viewed by others

Background
Abiotic stress seriously affects the whole life process of plants, which can cause retardation of growth and development, and reduction of yield and quality [1]. With long-term evolution process, plants have developed some adaptive mechanisms to environmental stress, including regulating the coordinated expression of various stress response genes, especially transcription factors (TFs) [1]. TFs regulate the transcription of target genes through specific interactions of their DNA-binding domains with target gene promoters [2]. The zinc finger TF family is further grouped into different subfamilies according to their structural and functional diversity. The B-box (BBX) proteins are a class of zinc finger TFs with diverse function, and have received widespread attention in plants [3].
BBX TFs have specialized tertiary structures that are stabilized through binding Zn ions. BBX proteins have a general structure that consists of one or two BBX domains (near the N-terminus), and some have a CCT (CONSTANS, CO-like and TOC1) domain (near the C-terminus) [4]. The N-terminal BBX domain participates in specific protein-protein interactions, and the CCT domain plays an essential role in regulating gene transcription [5, 6]. The BBX gene family was first identified in Arabidopsis thaliana, and 32 members were divided into five groups according to the number of BBX and CCT domains [7]. Subsequently, BBX genes were also characterized in various plants. For example, 30 OsBBXs, 31 SlBBXs, 64 MdBBXs, and 25 VvBBXs have been identified in rice [8], tomato [9], apple [10], and grapevine [11], respectively.
BBX proteins have been shown to play a role in seedling photomorphogenesis [4], flowering [12, 13], leaf senescence [14] and the shade avoidance response [15]. Several AtBBX genes, including CO, COL3, COL5 and COL9, can mediate flowering via the photoperiod pathway in Arabidopsis [16,17,18,19]. Studies have shown that BBX protein members integrate light signals perceived by plant photochromes and cryptic photoreceptors via the HY5-COP1 regulatory module, thereby influencing photomorphogenesis in seedlings [20, 21]. It has been proved that BBX proteins participate in the regulation of secondary metabolite biosynthesis, especially anthocyanins [22] and carotenoids [23]. In apple, MdMYB1 and MdMYB9 can positively regulate anthocyanin biosynthesis; however, MdBBX37 inhibits the binding of these genes to target genes by interacting with the MdMYB1 and MdMYB9 proteins, thus suppressing anthocyanin accumulation [24, 25]. In tomato, SlBBX20 promotes carotenoid accumulation through direct activation of the carotenoid biosynthesis enzyme PSY1 [23].
BBX proteins also play vital roles in response to abiotic stress and in the regulation of phytohormonal signaling in plants. BBX2 expression is consistently upregulated during long periods of cold stress in Arabidopsis [26]. BBX18 and BBX23 actively regulate the thermal morphogenesis of Arabidopsis by interacting with ELF3 and COP1, while their mutations cause thermo responsive hypocotyl shortening [27]. Heterologous expression of MdBBX10 in Arabidopsis significantly increased the drought and salt tolerance of plants by ABA signaling [28]. Recent studies have shown that the jasmonic acid (JA)-mediated cold stress response can be positively regulated by the BBX37 protein in apple, which is mainly attributed to the synergistic regulation of the BBX37-ICE 1-CBF module by JAZ [29]. In addition, BBX family genes function in phytohormone signaling pathways. For example, AtBBX18 (AtDBB1a) accelerates hypocotyl elongation by accumulating the content of gibberellin (GA) [30], whereas AtBBX20 (AtBZS1) negatively regulates the brassinosteroid signaling pathway [31]. MdBBX22 directly interacts with ABI5, the key regulator of the abscisic acid (ABA) signalling pathway, thereby inhibiting MdABI5 transcriptional activity [32]. However, the role of alfalfa MsBBX genes in the abiotic stress response and phytohormone signaling pathways remains to be investigated.
Alfalfa (Medicago sativa L.) represents the most important and widely distributed legume plant globally, and has been praised for its high protein content, nutritional quality, good palatability and strong adaptability [33]. However, alfalfa growth/development and production are severely restricted by environmental factors, especially water deficiency and salinity stress, thus affecting quality and yield. Although BBX family members play essential roles in many plant species, little research has been conducted on alfalfa BBX genes and their roles in various stress responses to date. The publication of alfalfa whole genome data facilitates a comprehensive detailed analysis of genes in the BBX family [34]. In the present study, we systematically analyzed the alfalfa BBX gene family at the whole genome level, including protein basic information, phylogenetic relationships, chromosomal distributions, gene structures, conserved domains and motifs, and cis-regulatory elements. Furthermore, we analyzed transcriptomic data and performed qRT-PCR analysis to investigate the expression of the alfalfa BBX genes. The function of the MsBBX11 gene in salt stress tolerance was identified by heterologous expression in Arabidopsis. Our results provide a basis for further exploration of the function of MsBBX genes and resistance breeding in alfalfa.
Results
Genome-wide identification of MsBBX genes in alfalfa
To identify the candidate members of BBX gene family in alfalfa, 32 AtBBXs and 30 OsBBXs were used as query sequences to screen protein database of alfalfa. After removing redundant sequences and performing domain identification, a total of 28 putative MsBBX genes containing the BBX domain (PF00643) were confirmed in the alfalfa genome and were named MsBBX1 to MsBBX28 according to their chromosomal position (Table 1). Analysis of the protein physicochemical properties showed that the MsBBX family members had an average of approximately 297 amino acids (aa) in length, ranging from 190 (MsBBX21/23) to 436 aa (MsBBX2). The molecular weights (MWs) of the MsBBX proteins ranged from 20.95 (MsBBX21) to 48.73 (MsBBX2) kDa, and the theoretical isoelectric points (pIs) ranged from 4.84 (MsBBX9) to 9.74 (MsBBX4). In addition, the aliphatic indices of the MsBBX proteins ranged from 52.52 (MsBBX6) to 73.80 (MsBBX22). The GRAVY values of all the proteins were negative, ranging from (-0.926) (MsBBX6) to (-0.342) (MsBBX18), implying that the MsBBXs are hydrophilic proteins. Subcellular localization prediction showed that all MsBBXs were located in the nuclei (Table 1).
Phylogenetic analysis of MsBBX proteins
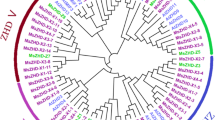
A neighbor-joining tree was constructed using MEGA to investigate the genetic evolution relation of the MsBBX gene family based on BBX proteins from alfalfa (28), Arabidopsis (32) and rice (30). As shown in Fig. 1, the 90 BBXs were divided into five subfamilies (I-V) depending on the sequence homology, and they were unequally distributed among the five subfamilies. Interestingly, no MsBBX members of alfalfa were found in subfamily V, which had the fewest BBX members with only 10 proteins. The results showed that the largest cluster was subfamily IV with 33 BBX members, including 15 MsBBXs, 8 AtBBXs and 10 OsBBXs. There were 20, 12 and 15 BBX members in subfamilies I, II and III, with 7, 5, and 1 MsBBX members, respectively. Concurrently, BBX proteins from alfalfa, Arabidopsis and rice in subfamilies I-IV were grouped into the same clade, suggesting that the BBX family was highly evolutionary conserved and might perform similar biological functions. The phylogenetic tree showed that the BBX proteins of alfalfa were more closely related to their orthologous in Arabidopsis than those in rice (Fig. 1).

Phylogenetic tree analysis of BBX proteins from alfalfa (MsBBX), Arabidopsis (AtBBX) and rice (OsBBX). The tree was divided into five clades represented by different colors. The bootstrap values are indicated at each node
Gene structure, motif, and conserved domain analysis of the MsBBX genes
To explore the structural features of the alfalfa MsBBX gene family, the exon-intron structure and conserved motifs were identified. The results showed that there were 2 to 5 exons and 1 to 4 introns in the MsBBX genes (Fig. 2A). We found that 11 MsBBX genes (39.3%) contained two exons, 13 genes (46.4%) had three exons, and 3 genes (10.7%) had four exons. In particular, MsBBX22 had five exons and four introns. However, all 28 MsBBX genes lacked the UTR region (Fig. 2A). Conserved motif prediction showed that ten distinct motifs (motifs 1–10) were discovered in MsBBX proteins (Fig. 2B). Among them, motifs 1, 2 and 6 were the top three motifs, and were present in 100%, 46.4% and 35.7%, respectively, of the MsBBX proteins, indicating that these motifs are the most critical components of MsBBXs. According to Fig. 2C, motif 1 and motif 2 are the B-box and CCT domains, respectively. The B-box domain was distributed among all the MsBBX proteins, with a total of 23 MsBBXs having two B-boxes, while the other proteins (MsBBX2, MsBBX4, MsBBX6, MsBBX19, and MsBBX20) had only one. In addition, we identified 13 MsBBXs containing a CCT domain (Fig. 2C). The number of motifs in the MsBBXs ranged from 3 to 6. Most of the MsBBX members (11) contained three motifs, and four members had six motifs. Furthermore, the MsBBX proteins that were closely related to the proteins in the phylogenetic tree had similar motif compositions. For instance, MsBBX11, MsBBX13, and MsBBX15 all contained 5 motifs, including motifs 1, 3, 6, and 10. The motif 9 only appeared in MsBBX2, MsBBX4, and MsBBX6 (Fig. 2B). Similar to the composition of motifs, the conserved domains were also distributed according to genetic relationships (Fig. 2C). Multiple sequence alignment showed the conserved domain locations in the MsBBX protein sequences, where all the MsBBXs shared the conserved B-box domain at the N-terminus, and some of the sequences also had a CCT domain at the C-terminus (Fig. S1).

Gene structure, conserved motif and conserved domain analysis of MsBBX genes. A Exon-intron structure analysis of MsBBX genes. The orange boxes and black lines indicate exons and introns, respectively. B Motif positions of MsBBX genes. Each motif is represented in a colored box. C Conserved domain analysis of MsBBX proteins. The green and yellow boxes indicate the B-box and CCT domains, respectively
Chromosome localization, gene duplication and collinearity analysis of MsBBX genes
The chromosomal positions and collinearity of the alfalfa MsBBX genes were mapped against published genome data. The 28 MsBBX genes were unevenly scattered across 20 out of the 32 chromosomes in alfalfa (Fig. S2, Fig. 3A). Each of the eight chromosomes (Chr1.1, 1.2, 1.4, 2.3, 3.3, 3.4, 4.2, 8.1) contained two MsBBX genes, and the remaining twelve chromosomes (Chr2.1, 3.1, 3.2, 4.1, 4.3, 4.4, 5.1, 5.3, 5.4, 7.1, 7.2) contained only one MsBBX gene. Gene duplication event analysis showed that no tandem duplications occurred in the alfalfa MsBBX gene family. Notably, a total of 24 gene pairs exhibited segmental duplication events, and these genes were distributed on chromosomes 1, 2, 3, 4, 5, and 8 (Table S1, Fig. 3A). Most MsBBX genes had 1–3 paralogous genes in alfalfa, while five MsBBXs (MsBBX11, MsBBX13, MsBBX15, MsBBX16, and MsBBX17) were found with 4 paralogous genes.
To explore the potential evolutionary relationships of the BBX genes between alfalfa and A. thaliana, O. sativa, or M. truncatula, a comparison of collinear maps were constructed. As shown in Fig. 3B and Table S2, a total of 23, 8 and 26 MsBBX genes showed syntenic relationships with Arabidopsis, O. sativa, and M. truncatula, respectively. Among these MsBBXs, 36, 9, and 40 pairs of orthologous genes were found between alfalfa and Arabidopsis, O. sativa, and M. truncatula, respectively. Most MsBBX genes were identified with 1–2 orthologous gene pairs in Arabidopsis, while MsBBX12 and MsBBX14 had three orthologous gene pairs. Except for MsBBX10 and MsBBX27, all the other MsBBX genes displayed syntenic relationships with M. truncatula, and MsBBX3 and MsBBX16 had three orthologous genes. Moreover, the largest number of collinear gene pairs was observed between alfalfa and M. truncatula, suggesting that the BBX proteins were highly conserved between the two legumes (Fig. 3B).

Chromosome distributions of MsBBX genes and synteny analysis between M. sativa and three other plant species. A Chromosomal location and duplication event analysis in the M. sativa genome. The segmental duplicated genes are connected by red curves. B Synteny analysis between M. sativa and A. thaliana, O. sativa, and M. truncatula. The grey lines indicate synteny blocks in M. sativa and the other species, while the red lines highlight the collinearity of BBX gene pairs
Analysis of cis-regulatory elements in MsBBX gene promoters
To better explore the potential regulatory mechanism of the MsBBX gene family, the cis-regulatory elements in the promoter sequences (2000 bp upstream of the start codon) of the MsBBX genes were analyzed using PlantCARE. Results showed that 39 types of cis-regulatory elements in the MsBBX promoter regions, with 12 (31%) related to stress response, nine (23%) related to growth and development, nine (23%) related to light responsiveness, and nine (23%) related to phytohormone response, respectively (Fig. 4, Table S3). All the MsBBXs contained these four categories of cis-regulatory elements. Stress-responsive elements were the most abundant elements, with MYC and MYB elements present in all the MsBBX family genes, ranging from 2 to 11 and 1 to 8, respectively. In addition, more than 78% of the MsBBX genes contained ARE and STRE of stress-related cis-elements (Fig. 4). In particular, stress and light response elements were most common in the MsBBX19 gene. The phytohormone-related elements identified in the MsBBX genes were associated with MeJA-responsive, ABA-responsive, IAA-responsive, GA-responsive, and SA-responsive (Table S3). Importantly, the ABRE involved in the ABA response, TGACG-motif (CGTCA-motif) involved in the JA response, and ERE involved in the ET response, appeared 77, 35 and 32 times in 26, 18 and 19 MsBBX genes, respectively, accounting for more than 64% of phytohormone responsive genes. Moreover, 87 and 54 G-box and Box4 elements involved in light responsiveness were found in 26 and 23 MsBBX genes, respectively. These results suggest that MsBBXs may play a key role in the alfalfa response to different environmental stresses and plant growth regulators.

Cis-regulatory element analysis of MsBBX genes. The gradient colors in the grid represent the number of cis-regulatory elements in the MsBBXs. The multicolor histogram indicates the number of different categories of cis-elements in these MsBBX genes
Expression profiles of MsBBX genes in different tissues in alfalfa
In order to study the expression patterns of MsBBX genes in alfalfa, we analyzed the transcriptome data of leaves, flowers, pre-elongated stems, elongated stems, roots and nodules in the NCBI database (Fig. S3). In the database, 15 MsBBX genes were found in different tissues of alfalfa, while no relevant information was found for the remaining 13 MsBBX genes. As shown in Fig. S3, most of the MsBBXs were highly expressed in leaves, flowers, pre-elongated stems and elongated stems, while their expression levels were lowest in nodules, suggesting that they play a role in the development of aboveground tissues. Interestingly, the expression level of MsBBX14 was the highest in the root tissues, indicating that MsBBX14 plays crucial roles in root development. Five MsBBX genes (MsBBX7, MsBBX15, MsBBX18, MsBBX21, and MsBBX27) were highly expressed in the flowers. MsBBX9 was more highly expressed in leaves than in other plant tissues, whereas the maximum expression of MsBBX10 and MsBBX17 occurred in pre-elongated stems. These results suggest that the MsBBX genes have tissue-specific expression profiles and functions during alfalfa development.
Expression profiles of MsBBX genes in alfalfa under different abiotic stresses
To further explore the expression profiles of MsBBX members under abiotic stress conditions, we downloaded the transcriptome data for alfalfa plants treated with drought and salt from the NCBI, and performed RNA-seq analysis. As shown in Fig. S4A, most of the MsBBX genes were positively induced by drought stress. The transcription levels of 8 MsBBXs (MsBBX7, MsBBX8, MsBBX11, MsBBX12, MsBBX16, MsBBX20, MsBBX26, and MsBBX28) peaked at 1 h and then decreased gradually with increasing drought duration. MsBBX2/18 and MsBBX4/21 were significantly upregulated at 3 and 12 h, respectively. However, MsBBX14 and MsBBX19 expression levels were significantly downregulated after drought stress. In addition, MsBBX15 expression was unaltered during drought treatment (Fig. S4A). Under salt treatment, the expression of most of the MsBBX transcripts changed except for that of MsBBX15 and MsBBX21 (Fig. S4B). The transcript levels of 21 MsBBX genes were upregulated to different degrees under salt treatment at different times. For instance, the transcript levels of MsBBX12, MsBBX16, MsBBX17, MsBBX20 and MsBBX26 significantly increased after 0.5 h of salt treatment, while the transcript levels of MsBBX2, MsBBX4, MsBBX11, MsBBX27 and MsBBX28 markedly increased after 1 h. The peak expression levels of MsBBX3 were observed after 24 h of salt stress. The expression of four MsBBX genes (MsBBX6, MsWBBX13, MsBBX14 and MsBBX19) decreased differentially during salt treatment (Fig. S4B). These results indicate that MsBBX genes may be involved in drought and salt stress responses in alfalfa.
We randomly selected six MsBBX genes (MsBBX3, MsBBX7, MsBBX8, MsBBX11, MsBBX20, and MsBBX28) that responded positively to drought and salt stress for qRT-PCR verification. As shown in Fig. 5, the six MsBBX genes exhibited diverse expression patterns during drought and salt stress treatments, and the patterns were largely consistent with the results of the transcriptome analysis. All the selected MsBBX genes were strongly induced by drought stress, and their expression was strongly elevated and peaked at 2 or 8 h. These MsBBX genes were also strongly induced by salt stress, and the expression levels reached a maximum at 2, 8 or 12 h (Fig. 5). In particular, the MsBBX11 gene was upregulated 27-fold at 2 h of salt stress treatment compared to the 0 h. Except for the MsBBX3 gene, the expression levels of all the selected MsBBX genes initially increased and subsequently decreased under salt stress. These results indicate that these MsBBX genes may participate in drought and salt adaptation in alfalfa.

Expression analysis of six selected genes under drought and salt stresses by qRT-PCR. Values are the means ± SEs (n = 9). Asterisks and double asterisks above the bars indicate significant differences: *P < 0.05; **P < 0.01
Analysis of MsBBX gene expression in response to plant growth regulator treatments in alfalfa
To identify hormone-responsive MsBBXs, we investigated the expression of MsBBX family genes in alfalfa treated with ABA at different times using RNA-seq data. The expression of most MsBBX genes was induced at different levels under ABA treatment (Fig. S5). Twenty-two MsBBX genes exhibited a positive response to ABA treatment, of which the expression of 17 MsBBXs first increased and then decreased. In particular, the expression of MsBBX11/12/17/22/28 and MsBBX4/15/24 increased dramatically after 1 and 3 h of ABA treatment, respectively. Compared with the control treatment, ABA treatment caused a gradual decrease in the transcript levels of MsBBX9, MsBBX13, MsBBX14 and MsBBX18 compared to 0 h. The expression level of MsBBX21 remained unchanged during ABA treatment (Fig. S5).
According to the RNA-seq analysis, six genes (MsBBX4, MsBBX11, MsBBX15, MsBBX17, MsBBX24 and MsBBX28) that positively responded to ABA treatment were analyzed by qRT-PCR at 0 h, 2 h, 4 h, 8 h, and 12 h after ABA, JA and SA treatments to investigate the response of MsBBX genes to plant growth regulators (Fig. 6). The expression of all the selected MsBBX genes was significantly induced by ABA, JA and SA at different treatment time intervals. The transcript levels of MsBBX4, MsBBX15 and MsBBX24 peaked after 4 h of ABA treatment, and that of MsBBBX4 increased almost 75-fold compared with that at 0 h. MsBBX11, MsBBX17 and MsBBX28 were highly induced in response to ABA treatment at 2 or 8 h. The expression trends of these MsBBX genes under ABA treatment were consistent with the transcriptome analysis results. All six MsBBX genes responded positively to JA treatment and reached maximum expression at 2, 4 or 8 h (Fig. 6). The expression level of the MsBBX genes showed a trend of first increasing and then decreasing under JA treatment conditions. MsBBX17 was highly expressed at 4 h and upregulated by 14-fold compared with that at 0 h. With the exception of MsBBX28, the expression of the selected genes initially increased and subsequently decreased under SA treatment. The expression of the MsBBX28 gene increased significantly (approximately 7-fold) at 12 h compared with that in the control. Notably, MsBBX15 was strongly induced in response to SA treatment at 4 h, reaching approximately 21-fold greater expression than that at 0 h (Fig. 6).

Expression analysis of six selected genes under ABA, JA and SA treatments by qRT-PCR. Values are the means ± SEs (n = 9). Asterisks and double asterisks above the bars indicate significant differences: *P < 0.05; **P < 0.01
Protein-protein interactions among the MsBBXs
To explore the comprehensive functions of MsBBXs in alfalfa, a protein interaction network was generated based on homologous proteins from Arabidopsis using the STRING database. A total of 12 MsBBX proteins were predicted to interact with each other (Fig. S6). The results showed that MsBBX23 had the most interactions with MsBBX proteins (seven), followed by MsBBX18 which interacted with six MsBBX proteins. MsBBX5 and MsBBX22 both have five interacting proteins and interact with each other. In addition, MsBBX2, MsBBX25 and MsBBX27 interacted separately with one MsBBX protein each, namely MsBBX22, MsBBX1 and MsBBX23, respectively. These results indicate that MsBBXs may function through interactions.
Subcellular localization of the MsBBX proteins
Prediction of the subcellular localization of MsBBX proteins using Plantm-PLoc revealed that all the MsBBXs were localized in the nuclei (Table 1). To further verify the prediction results and understand the functions of MsBBXs, we selected two MsBBX genes (MsBBX4 and MsBBX11) that were strongly induced by abiotic stress or plant growth regulators for transient expression in tobacco leaves. The results showed that the green fluorescent signals expressed by the MsBBX4-GFP and MsBBX11-GFP fusion vectors could be observed only in the nuclei, which was consistent with the predicted results (Fig. 7). These results suggest that MsBBX4 and MsBBX11 encode nuclear-localized proteins.

Subcellular localization of the MsBBX4 and MsBBX11 proteins. Images from left to right represent green fluorescent protein (GFP), bright field and an overlay (GFP and bright field) from the same sample. Scale bar = 25 μm
Overexpression of MsBBX11 in Arabidopsis confers tolerance to salt stress
To reveal the biological roles of the MsBBX genes, we selected a gene with high expression under salt stress, MsBBX11, from the qRT-PCR data of alfalfa for further study. MsBBX11 transgenic Arabidopsis plants were obtained by PPT screening and confirmed by semi-quantitative RT-PCR analysis. Two homozygous lines (OE1 and OE3) were randomly selected for further salt tolerance assays. Semi-quantitative RT-PCR result showed that MsBBX11 expression was detected in OE1 and OE3 lines but not in WT plants (Fig. 8A, Fig. S7). As shown in Fig. 8B, salt stress inhibited the root growth of both WT and transgenic lines. However, after 100, 125 or 150 mM NaCl stress, the transgenic plants exhibited higher primary root length than the WT (Fig. 8C). Correspondingly, the fresh weight of the transgenic plants was significantly higher than that of WT plants (Fig. 8D). Moreover, the OE1 and OE3 lines showed significantly higher cotyledon greening rates than the WT under 150 mM NaCl treatment (Fig. 8E).

Overexpression of MsBBX11 enhanced the salt tolerance of Arabidopsis during the seedling stage. A Semi-quantitative RT-PCR analysis of MsBBX11 expression levels in WT and transgenic lines. B Root growth phenotypes of Arabidopsis WT and transgenic seedlings vertically grown on MS medium supplemented with 0, 100, 125 or 150 mM NaCl. C Analysis of root length. D Analysis of fresh weight. E Analysis of cotyledon greening rate. Values are the means ± SE of three biological replicates. Asterisks and double asterisks above the bars indicate significant differences: *P < 0.05; **P < 0.01
To elucidate the role of MsBBX11 in salt resistance in soil, MsBBX11-overexpresing plants and WT were exposed to 300 mM NaCl stress for 10 days. As shown in Fig. 9A, there was no obvious difference in morphology between the transgenic plants and WT under normal growth conditions. However, upon exposure to NaCl, the WT plants exhibited more conspicuous leaf damage than the transgenic lines (Fig. 9A). The survival rates of OE1 and OE3 lines were 83.2% and 85.8%, respectively, while only 54.5% of the WT plants survived (Fig. 9B). Moreover, the fresh weight, Fv/Fm ratio, and chlorophyll content of the transgenic plants were higher than those of the WT plants under salt stress (Fig. 9C-E). Salt stress increased electrolyte leakage and the accumulation of MDA and H2O2, but these effects were significantly greater in WT than in transgenic plants (Fig. 9F-H). There were no significant differences in these indicators between the WT and transgenic lines under normal growth conditions.

Overexpression of MsBBX11 enhanced the salt tolerance of Arabidopsis at the vegetative stage. A Growth performance of the WT and transgenic plants before and after 300 mM NaCl treatment. Survival rate B, fresh weight C, Fv/FmD, chlorophyll content E, electrolyte leakage F, MDA content G, and H2O2 accumulation H of the WT and transgenic plants before and after salt treatment. Values are the means ± SEs of three biological replicates. Asterisks and double asterisks above the bars indicate significant differences: *P < 0.05; **P < 0.01
Discussion
BBX proteins belong to a super family of zinc-finger TFs that participate in plant growth, development, and response to abiotic stress and phytohormones [7]. To date, whole genome identification of BBX family members has been performed extensively in dicots and monocots, for example, Arabidopsis [4], O. sativa [8], tomato [9], tobacco [35] and Malus domestica [10]. However, the identification and functional analysis of the BBX gene family in alfalfa have not been reported. In this study, we used the whole genome sequence to perform a systematic bioinformatic identification and functional analysis of the BBX gene family in alfalfa. The results of this study will provide valuable information for further investigations of the functions of MsBBX members in alfalfa and will provide candidate genes for alfalfa stress tolerance breeding.
In this study, we identified 28 MsBBX family members from the alfalfa genome. Ma et al. [36] counted the number of BBX genes in 13 different plant species and approximately 30 BBX family members in each, indicating that the number of BBX genes in these plants, including alfalfa, was relatively stable. In contrast, there are 64 BBX genes in apple [10] and 19 in millet [11]. Yin et al. [37] reported that there was no direct relationship between the number of BBX family genes and the plant genome size, and we speculate that the remarkable variation may be caused by species-specific duplications or deletions during evolution. Previous studies have shown that the BBX genes in various plants are typically classified into 5 subfamilies [4, 11, 37]. In the present study, BBX proteins were divided into 5 subfamilies (I-V) according to sequence similarity to Arabidopsis, rice and alfalfa BBX proteins (Fig. 1). However, no alfalfa MsBBX proteins were grouped into subfamily V, indicating that the evolution of alfalfa MsBBX family may be different from that of other plants. A similar phenomenon was found in tobacco plants [35].
The diversity of gene structures typically plays an important role in the evolution of multiple gene families. BBX family genes contain one or two conserved B-box domains, and some possess a CCT domain in plants [4]. We also found similar results for MsBBX family genes (Fig. 2), indicating that the BBXs are relatively conserved among different species. Based on gene structure and motif analysis, the MsBBX genes in the same groups had similar intron/exon combinations and motif compositions (Fig. 2), suggesting that they may have similar biological functions. Generally, gene duplication events are the main drivers of new gene emergence and genome evolution. Tandem and segmental replication are the two main duplication patterns in plants [38]. In alfalfa, a total of 24 pairs of segmental duplications were found in the MsBBX gene family, but no tandem duplications occurred (Fig. 3A), implying that segmental duplications were particularly beneficial for the expansion of MsBBX family members. Similar results were reported by Ma et al. [3] during the evolution of the PeBBX gene family. In addition, many isogenous gene pairs were detected between alfalfa and Arabidopsis, O. sativa, and M. truncatula (Fig. 3B), suggesting the indispensable role of these genes in the evolution of the BBX family.
BBX proteins are functionally diverse in regulating plant growth, development and stress responses, which is further supported by the detection of numerous hormone- and stress-related elements in the promoter regions of the tomato and tobacco BBX genes [9, 35]. Promoter cis-regulatory elements regulate the transcription of specific genes in response to stress signals by binding to transcription factors [39]. In the present study, promoter elements associated with growth and development, the stress response and plant growth regulator response were found to be abundant among the MsBBX genes (Fig. 4), indicating that the MsBBX genes may actively participate in these physiological processes and stress resistance. In Arabidopsis, BBX32 can regulate the flowering pathway via interaction with CONSTANS-LIKE 3 (COL3)/BBX4 [40]. The direct interaction between BBX32 and BBX21 suppresses BBX21-HY5 and thus functions in light signaling [41]. A protein interaction network analysis suggested that the MsBBX proteins might synergistically regulate the biological processes of alfalfa through interactions.
Previous studies have reported that BBX6/COL5 accelerated Arabidopsis flowering by activating the transcription of FT under short-day conditions [18], while in contrast, BBX32/EIP6 regulated flowering in a manner independent of CO under long day conditions [42]. According to the transcriptome data of alfalfa, most MsBBXs were related to the growth and development of aboveground tissues in alfalfa (Fig. S3). Among them, the expression levels of MsBBX7/15/18/21/27 varied greatly among the flowers, indicating that these genes might play a critical role in the regulation of flower development. In addition, Ma et al. [36] found that CaBBX5 and CaBBX6 are involved in photomorphogenesis and are highly expressed highly in leaves. Similarly, MsBBX9 and MsBBX17 exhibited relatively high transcriptional activity in alfalfa leaves and stems, respectively, while MsBBX14 was highly expressed in roots, suggesting their potential involvement in seedling morphogenesis.
Although BBX genes have diverse functions, we concerned about their response to abiotic stress. Previous studies have reported that nine VvBBX genes were significantly upregulated in response to drought stress in berry [43]. In alfalfa, we found that most of the MsBBX genes positively responded to drought or salt stress (Fig. S4A, S4B), suggesting that these genes may have potential functions in plant drought or salt tolerance. The MdBBX10 gene has been proven to enhance the drought and salt tolerance of transgenic Arabidopsis [28]. This study revealed that several MsBBXs were responsive to both drought and salt stress, as verified by the qRT-PCR results for the six selected genes (Fig. 5). Therefore, it can be assumed that the MsBBX genes are positive regulators of drought and salt stress signaling in alfalfa.
Previous studies have shown that the transcription of AtBBX24 is positively related to salt stress signaling, and that the overexpression of AtBBX24 significantly increases salt stress resistance in Arabidopsis [44]. In this study, we further validated the function of the MsBBX11 gene, which actively responded to salt stress. It was found that overexpression of MsBBX11 in Arabidopsis promoted seedling growth and photosynthetic capacity, and reduced cell membrane damage and H2O2 accumulation (Figs. 8 and 9), thus conferring salt tolerance to the plants. The difference in transgene expression is an important factor determining the effectiveness of transgenic transformation, and is usually influenced by the sequence flanking the insertion site or other factors [45]. The expression level of the OE1 plants was relatively lower than that of the OE3 plants, and this difference might be related to the positional effect of the transgene or the specific insertion mode [45].
Plant BBX genes are also involved in hormone signal transduction. In Arabidopsis, BBX21 physically interacts with the HY5 or ABI5 proteins to repress ABI5 expression, thereby negatively regulating the inhibition of seed germination by ABA [46]. Recent studies have revealed that the BBX22-ABI5 interaction module negatively regulates chlorophyll degradation and leaf senescence through an ABA-dependent pathway [32]. MdBBX37 positively regulates JA-mediated cold stress tolerance through the JAZ-BBX37-ICE1-C BF pathway in apple [29]. In pepper, five BBX genes were significantly induced by SA treatment [36]. Most MsBBXs were upregulated under ABA treatment in alfalfa (Fig. S5). In addition, the expression of six selected genes (MsBBX4, MsBBX11, MsBBX15, MsBBX17, MsBBX24 and MsBBX28) was dramatically induced by ABA, JA and SA treatments (Fig. 6), which corresponded to phytohormone response elements in their promoter regions. It has been previously reported that MdBBX10 enhances abiotic stress tolerance through ABA signaling [28]. In this study, the MsBBX11 and MsBBX28 genes were positively induced by drought, salt, and plant growth regulator treatments simultaneously. However, whether MsBBXs regulate abiotic stress through hormone signaling remains to be further explored. Therefore, we speculate that MsBBXs may serve as positive regulators of ABA, JA, and SA signal transduction, participating in regulation of growth and abiotic stresses in alfalfa.
Conclusions
In the present study, 28 MsBBX genes were systematically explored in alfalfa and phylogenetically grouped into four subfamilies. The discovery of duplication and collinearity gene pairs provided valuable information about the evolutionary history of the MsBBX genes. We discovered that the MsBBX genes exhibit tissue specificity and that most MsBBXs may play important roles in aboveground tissue development. Furthermore, the expression of several MsBBX genes was significantly induced by drought, salt and hormone stress, suggesting that MsBBXs play essential roles in plant stress response. For instance, the MsBBX11 gene markedly improved the salt tolerance of transgenic Arabidopsis and can be used as a candidate gene for salt tolerance breeding in alfalfa. It will be of great interest to investigate the biological functions of these MsBBX genes and elucidate their detailed regulatory mechanisms in the future. This study lays an important foundation for creating stress resistant germplasms and breeding new varieties of alfalfa in the future.
Materials and methods
Plant materials
The alfalfa cultivar ‘Zhongmu No. 1’ was used in this study, and its seeds were provided by the Institute of Animal Science, Chinese Academy of Agricultural Sciences, China. Seeds of tobacco (Nicotiana benthamiana) and Arabidopsis thaliana Columbia-0 wide type (WT) used in this study were preserved in our laboratory (Laboratory of Forage Molecular Breeding, Ningxia University, China).
Genome-wide identification of alfalfa BBX genes
We downloaded the whole genome and annotation files of alfalfa from the website https://figshare.com/projects/whole_genome_sequencing_and_assembly_of_Medicago_sativa/66380 [34]. The reported BBX protein sequences of Arabidopsis thaliana and Oryza sativa were acquired from the TAIR website (https://www.arabidopsis.org/) and Phytozome13 database (https://phytozome-next.jgi.doe.gov/), respectively [47]. These proteins were queried against BBX gene family members from alfalfa in the BLASTP search. The Hidden Markov Model (HMM) profile of the B-box domain (PF00643) was used as the seed sequence to search the alfalfa genome [8]. After manually removing the redundant sequences, the candidate MsBBX family genes were further identified using the Conserved Domains Database (CDD) (http://www.ncbi.nlm.nih.gov/cdd/), SMART (https://smart.embl-heidelberg.de/) and Pfam database comparison (http://pfam.xfam.org/) [48].
Protein property analysis and phylogenetic analysis
TBtools software was used to acquire the genomic positions of the corresponding MsBBX genes from the alfalfa genome data [49]. The physical and chemical properties of the MsBBX proteins were predicted by ExPASy website (http://web.expasy.org/protparam/) [50]. Subcellular localization of the MsBBX proteins was determined by Plantm-PLoc (http://www.csbio.sjtu.edu.cn/bioinf/plant-multi/) [51]. Phylogenetic relationships of the BBX proteins among alfalfa, Arabidopsis and rice were analyzed based on the neighbor-joining method (1,000 bootstraps) with MEGA 7.0 software [52]. The image of the phylogenetic tree was then beautified using iTOL online software (https://itol.embl.de/) [53].
Gene structure, conserved motif, domain analysis and multiple sequence alignments
The exon-intron structure of MsBBX genes was obtained from the online website GSDS: http://gsds.gao-lab.org/ [54]. Conserved motifs of MsBBXs were identified using Multiple Expectation Maximization for Motif Elicitation (MEME) software [55]. The number of repetitions was set to any, the width of the conserved sites was restricted between 6 and 50, and the maximum number of motifs was set to 10. In addition, the conserved domains of the MsBBX proteins were analyzed by CDD databases (http://www.ncbi.nlm.nih.gov/cdd/). The ClustalW program was used to perform multiple sequence alignments of these MsBBX proteins [56].
Chromosomal localization and gene duplication analysis of the MsBBX gene family
The chromosomal localization information of BBX genes was retrieved from the genome files of different plants and visualized using TBtools software. The MsBBX gene duplication events and collinearity of BBX genes in alfalfa, Arabidopsis, rice and Medicago truncatula were determined by MCScanX software.
Cis-regulatory element and protein-protein interaction analysis of the MsBBX gene family
The sequences 2000 bp upstream of the start codon (ATG) of the MsBBX genes were extracted from the alfalfa genome database. The PlantCARE online tool (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/) was used to analyze the cis-regulatory elements [57]. The cis-acting element diagrams of MsBBX genes were drawn with TBtools software. The STRING database was used to predict the protein-protein interaction network of the MsBBX family genes based on their homologous in Arabidopsis [58].
Expression profiles of MsBBX genes with transcriptome data
We downloaded the transcriptome data for various tissues and stress treatments in alfalfa from the NCBI database (SRP055547, SRR7091780-7091794, and SRR7160313-7160357) [59,60,61]. This study analyzed six tissues including leaf, flower, pre-elongated stem, elongated stem, root and nodule and three abiotic stresses, salt, drought and ABA. The differential gene expression analysis was conducted using DESeq2 with |log2(fold change)| ≥ 1 and FDR < 0.01. TBtools software was used to complete the heatmap of MsBBX gene expression.
Plant growth conditions and treatments
Plants of the alfalfa cultivar ‘Zhongmu No. 1’ were grown hydroponically in a growth chamber at 23–26 °C and a photoperiod of 16 h light/8 h dark. After four weeks of incubation, the seedlings with consistent growth stages were separated into six groups: (1) control, (2) drought, (3) salt, (4) ABA, (5) JA, and (6) SA. The alfalfa seedlings were subsequently treated with Hoagland solution containing PEG6000 (20%), NaCl (200 mM), ABA (10 μM), SA (100 μM) or JA (100 μM) for 0, 2, 4, 8 or 12 h. Three independent replicates were set for each treatment time point (including control). After treatment, the leaves of alfalfa were frozen in liquid nitrogen and stored at -80 °C for gene expression analysis.
Gene expression pattern analysis of MsBBX genes by qRTPCR
The Eastep® Super total RNA Extraction kit (Promega, Shanghai, China) was used to extract the total RNA from each sample. RNA was reverse transcribed and first-strand cDNA was synthesized using a reverse transcription kit (Vazyme, Nanjing, China). qRT-PCR was carried out using ChamQ SYBR qPCR Master Mix (Vazyme, Nanjing, China) and the MsActin2 gene was used as an internal control [62]. All samples were run in three biological replicates, and each included three technical replicates. Relative expression levels of MsBBX genes were calculated using the 2−∆∆Ct method. The primer sequences and melting curves of the MsBBX genes are shown in Table S4 and Fig. S8.
Subcellular localization analysis
For subcellular location assays, the full-length of coding sequences without the stop codon of two selected MsBBX genes (MsBBX4 and MsBBX11) were amplified (specific primers are shown in Table S4) and cloned into the pCAMBIA1300-GFP vector, generating the pCAMBIA1300-MsBBX4/11-GFP fusion plasmids. The successfully constructed plasmids were subsequently introduced into A. tumefaciens strain GV3101, which was transiently expressed in tobacco leaves [63]. After 2 days of incubation in the dark, the GFP fluorescence signal was captured by a laser confocal microscope (Leica TCS SP8, Germany).
Plant transformation and transgenic plant generation
We transformed the A. tumefaciens strain GV3101 carrying the pCAMBIA1300-MSBX11-GFP recombinant vector into wild-type Arabidopsis using the floral dip method to obtain transgenic plants. Transformed Arabidopsis with overexpression of MsBBX11 were selected for 10 mg/L DL-phosphinothricin (PPT) and a total of 17 independent lines were generated. The homozygous transformants (T3) were further confirmed by semi-quantitative RT-PCR using specific primers (Table S4) and two lines (OE1 and OE3) were randomly selected for salt tolerance analysis.
Salt stress tolerance assays
For salt tolerance analysis of seedlings root elongation, Arabidopsis WT and transgenic lines seeds were grown vertically on 1/2 MS agar plates for 7 d and then transplanted to NaCl-containing 1/2 MS plates (0, 100 or 150 mM NaCl) for salt treatment. The primary root length of seedlings was determined after 7 days of growth. For the soil salinity tolerance assay, 7-day-old seedlings were transplanted into soil and watered with Hoagland solution. After two weeks of growth, the plants were irrigated with 300 mM NaCl solution for 10 d for salt treatment [64]. Thereafter, the plant phenotypes were photographed and the survival rates were calculated. The fresh weight of the rosette leaves was measured with a balance. The maximum quantum yield (Fv/Fm) of the leaves was measured after 30 min in the dark. The chlorophyll content was determined by 80% acetone according to Liu et al. [65]. Electrolyte leakage was analyzed according to Dahro et al. [66]. The malondialdehyde (MDA) content was measured using thiobarbituric acid (TBA) according to Puckette et al. [67]. The accumulation of hydrogen peroxide (H2O2) was spectrophotometrically determined according to Jiang and Zhang [68].
Statistical analysis
The data in the experiment was reported as means ± standard errors (SEs). The statistical significant differences between the control and treatment groups were determined by Student’s t-test at 5% (*P < 0.05) or 1% (**P < 0.01) probability levels with SPSS Statistical 20.0 software. Figures were produced using Sigmaplot software (version 12.5).
Data availability
All data generated or analyzed during the present study are available in the submitted manuscript and its supplementary material. The reference genome data and annotation information of alfalfa (Xinjiangdaye) were obtained from figshare data repository (https://figshare.com/projects/whole_genome_sequencing_and_assembly_of_Medicago_sativa/66380). The Arabidopsis and rice BBX protein sequences were downloaded from the TAIR (https://www.arabidopsis.org/) and Phytozome13 database (https://phytozome-next.jgi.doe.gov/), respectively. Transcriptome data for various tissues of alfalfa were downloaded from the NCBI database (SRP055547). Transcriptome data of alfalfa treated with salt, drought and ABA were downloaded from the NCBI database (SRR7091780-7091794 and SRR7160313-7160357).
References
Zhu JK. Abiotic stress signaling and responses in plants. Cell. 2016;167(2):313–24.
Zhang H, Zhu J, Gong Z, Zhu JK. Abiotic stress responses in plants. Nat Rev Genet. 2022;23(2):104–19.
Ma R, Chen J, Huang B, Huang Z, Zhang Z. The BBX gene family in Moso bamboo (Phyllostachys edulis): identification, characterization and expression profiles. BMC Genomics. 2021;22(1):533.
Khanna R, Kronmiller B, Maszle DR, Coupland G, Holm M, Mizuno T, Wu SH. The Arabidopsis B-box zinc finger family. Plant Cell. 2009;21(11):3416–20.
Qi Q, Gibson A, Fu X, Zheng M, Kuehn R, Wang Y, et al. Involvement of the N-terminal B-box domain of Arabidopsis BBX32 protein in interaction with soybean BBX62 protein. J Biol Chem. 2012;287(37):31482–93.
Yan H, Marquardt K, Indorf M, Jutt D, Kircher S, Neuhaus G, Rodríguez-Franco M. Nuclear localization and interaction with COP1 are required for STO/BBX24 function during photomorphogenesis. Plant Physiol. 2011;156(4):1772–82.
Gangappa SN, Botto JF. The BBX family of plant transcription factors. Trends Plant Sci. 2014;19(7):460–70.
Huang J, Zhao X, Weng X, Wang L, Xie W. The rice B-box zinc finger gene family: genomic identification, characterization, expression profiling and diurnal analysis. PLoS ONE. 2012;7(10):e48242.
Bu X, Wang X, Yan J, Zhang Y, Zhou S, Sun X, et al. Genome-wide characterization of B-Box gene family and its roles in responses to light quality and cold stress in tomato. Front Plant Sci. 2021;12:698525.
Liu X, Li R, Dai Y, Chen X, Wang X. Genome-wide identification and expression analysis of the B-box gene family in the apple (Malus domestica Borkh.) Genome. Mol Genet Genomics. 2018;293(2):303–15.
Zhang X, Zhang L, Ji M, Wu Y, Zhang S, Zhu Y, Yao J, Li Z, Gao H, Wang X. Genome-wide identification and expression analysis of the B-box transcription factor gene family in grapevine (Vitis vinifera L). BMC Genomics. 2021;22(1):221.
González-Schain ND, Díaz-Mendoza M, Zurczak M, Suárez-López P. Potato CONSTANS is involved in photoperiodic tuberization in a graft-transmissible manner. Plant J. 2012;70(4):678–90.
Lee YS, Jeong DH, Lee DY, Yi J, Ryu CH, Kim SL, et al. OsCOL4 is a constitutive flowering repressor upstream of Ehd1 and downstream of OsphyB. Plant J. 2010;63(1):18–30.
Liu Y, Chen H, Ping Q, Zhang Z, Guan Z, Fang W, Chen S, Chen F, Jiang J, Zhang F. The heterologous expression of CmBBX22 delays leaf senescence and improves drought tolerance in Arabidopsis. Plant Cell Rep. 2019;38(1):15–24.
Crocco CD, Holm M, Yanovsky MJ, Botto JF. Function of B-BOX under shade. Plant Signal Behav. 2011;6(1):101–4.
An H, Roussot C, Suárez-López P, Corbesier L, Vincent C, Piñeiro M, et al. CONSTANS acts in the phloem to regulate a systemic signal that induces photoperiodic flowering of Arabidopsis. Development. 2004;131(15):3615–26.
Datta S, Hettiarachchi GH, Deng XW, Holm M, Arabidopsis. CONSTANS-LIKE3 is a positive regulator of red light signaling and root growth. Plant Cell. 2006;18(1):70–84.
Hassidim M, Harir Y, Yakir E, Kron I, Green RM. Over-expression of CONSTANS-LIKE 5 can induce flowering in short-day grown Arabidopsis. Planta. 2009;230(3):481–91.
Cheng XF, Wang ZY. Overexpression of COL9, a CONSTANS-LIKE gene, delays flowering by reducing expression of CO and FT in Arabidopsis thaliana. Plant J. 2005;43(5):758–68.
Lin F, Jiang Y, Li J, Yan T, Fan L, Liang J, Chen ZJ, Xu D, Deng XW. B-BOX DOMAIN PROTEIN28 negatively regulates photomorphogenesis by repressing the activity of transcription factor HY5 and undergoes COP1-mediated degradation. Plant Cell. 2018;30(9):2006–19.
Heng Y, Lin F, Jiang Y, Ding M, Yan T, Lan H, Zhou H, Zhao X, Xu D, Deng XW. B-Box containing proteins BBX30 and BBX31, acting downstream of HY5, negatively regulate photomorphogenesis in Arabidopsis. Plant Physiol. 2019;180(1):497–08.
Bai S, Tao R, Tang Y, Yin L, Ma Y, Ni J, et al. BBX16, a B-box protein, positively regulates light-induced anthocyanin accumulation by activating MYB10 in red pear. Plant Biotechnol J. 2019;17(10):1985–97.
Xiong C, Luo D, Lin A, Zhang C, Shan L, He P, et al. A tomato B-box protein SlBBX20 modulates carotenoid biosynthesis by directly activating PHYTOENE SYNTHASE 1, and is targeted for 26S proteasome-mediated degradation. New Phytol. 2019;221(1):279–94.
Bai S, Saito T, Honda C, Hatsuyama Y, Ito A, Moriguchi T. An apple B-box protein, MdCOL11, is involved in UV-B- and temperature-induced anthocyanin biosynthesis. Planta. 2014;240(5):1051–62.
An JP, Wang XF, Espley RV, Lin-Wang K, Bi SQ, You CX, Hao YJ. An apple B-Box protein MdBBX37 modulates anthocyanin biosynthesis and hypocotyl elongation synergistically with MdMYBs and MdHY5. Plant Cell Physiol. 2020;61(1):130–43.
Hannah MA, Heyer AG, Hincha DK. A global survey of gene regulation during cold acclimation in Arabidopsis thaliana. PLoS Genet. 2005;1(2):e26.
Ding L, Wang S, Song ZT, Jiang Y, Han JJ, Lu SJ, Li L, Liu JX. Two B-Box domain proteins, BBX18 and BBX23, interact with ELF3 and regulate thermomorphogenesis in Arabidopsis. Cell Rep. 2018;25(7):1718–28.
Liu X, Li R, Dai Y, Yuan L, Sun Q, Zhang S, Wang X. A B-box zinc finger protein, MdBBX10, enhanced salt and drought stresses tolerance in Arabidopsis. Plant Mol Biol. 2019;99(4–5):437–47.
An JP, Wang XF, Zhang XW, You CX, Hao YJ. Apple B-box protein BBX37 regulates jasmonic acid mediated cold tolerance through the JAZ-BBX37-ICE1-CBF pathway and undergoes MIEL1-mediated ubiquitination and degradation. New Phytol. 2021;229(5):2707–29.
Wang Q, Zeng J, Deng K, Tu X, Zhao X, Tang D, Liu X. DBB1a, involved in gibberellin homeostasis, functions as a negative regulator of blue light-mediated hypocotyl elongation in Arabidopsis. Planta. 2011;233(1):13–23.
Fan XY, Sun Y, Cao DM, Bai MY, Luo XM, Yang HJ, et al. BZS1, a B-box protein, promotes photomorphogenesis downstream of both brassinosteroid and light signaling pathways. Mol Plant. 2012;5(3):591–00.
An JP, Zhang XW, Liu YJ, Zhang JC, Wang XF, You CX, Hao YJ. MdABI5 works with its interaction partners to regulate abscisic acid-mediated leaf senescence in apple. Plant J. 2021;105(6):1566–81.
Chen L, He F, Long R, Zhang F, Li M, Wang Z, Kang J, Yang Q. A global alfalfa diversity panel reveals genomic selection signatures in Chinese varieties and genomic associations with root development. J Integr Plant Biol. 2021;63(11):1937–51.
Chen H, Zeng Y, Yang Y, Huang L, Tang B, Zhang H, et al. Allele-aware chromosome-level genome assembly and efficient transgene-free genome editing for the autotetraploid cultivated alfalfa. Nat Commun. 2020;11(1):2494.
Song K, Li B, Wu H, Sha Y, Qin L, Chen X, Liu Y, Tang H, Yang L. The function of BBX gene family under multiple stresses in Nicotiana tabacum. Genes (Basel). 2022;13(10).
Ma J, Dai JX, Liu XW, Lin D. Genome-wide and expression analysis of B-box gene family in pepper. BMC Genomics. 2021;22(1):883.
Yin Y, Shi H, Mi J, Qin X, Zhao J, Zhang D et al. Genome-wide identification and analysis of the BBX gene family andits role in carotenoid biosynthesis in wolfberry (Lycium barbarum L). Int J Mol Sci. 2022;23(15).
Freeling M. Bias in plant gene content following different sorts of duplication: tandem, whole-genome, segmental, or by transposition. Annu Rev Plant Biol. 2009;60:433–53.
Singh K, Foley RC, Oñate-Sánchez L. Transcription factors in plant defense and stress responses. Curr Opin Plant Biol. 2002;5(5):430–36.
Tripathi P, Carvallo M, Hamilton EE, Preuss S, Kay SA. Arabidopsis B-BOX32 interacts with CONSTANS-LIKE3 to regulate flowering. Proc Natl Acad Sci USA. 2017;114(1):172–7.
Holtan HE, Bandong S, Marion CM, Adam L, Tiwari S, Shen Y, et al. BBX32, an Arabidopsis B-Box protein, functions in light signaling by suppressing HY5-regulated gene expression and interacting with STH2/BBX21. Plant Physiol. 2011;156(4):2109–23.
Park HY, Lee SY, Seok HY, Kim SH, Sung ZR, Moon YH. EMF1 interacts with EIP1, EIP6 or EIP9 involved in the regulation of flowering time in Arabidopsis. Plant Cell Physiol. 2011;52(8):1376–88.
Wei H, Wang P, Chen J, Li C, Wang Y, Yuan Y, Fang J, Leng X. Genome-wide identification and analysis of B-BOX gene family in grapevine reveal its potential functions in berry development. BMC Plant Biol. 2020;20(1):72.
Nagaoka S, Takano T. Salt tolerance-related protein STO binds to a myb transcription factor homologue and confers salt tolerance in Arabidopsis. J Exp Bot. 2003;54(391):2231–37.
Halweg C, Thompson WF, Spiker S. The Rb7 matrix attachment region increases the likelihood and magnitude of transgene expression in tobacco cells: a flow cytometric study. Plant Cell. 2005;17(2):418–29.
Xu D, Li J, Gangappa SN, Hettiarachchi C, Lin F, Andersson MX, Jiang Y, Deng XW, Holm M. Convergence of light and ABA signaling on the ABI5 promoter. PLoS Genet. 2014;10(2):e1004197.
He F, Shi YJ, Li JL, Lin TT, Zhao KJ, Chen LH, et al. Genome-wide analysis and expression profiling of Cation/H (+) exchanger (CAX) family genes reveal likely functions in cadmium stress responses in poplar. Int J Biol Macromol. 2022;204:76–88.
Li N, Xu R, Wang B, Wang J, Huang S, Yu Q, Gao J. Genome-wide identification and evolutionary analysis of the SRO gene family in tomato. Front Genet. 2021;12:753638.
Chen C, Chen H, Zhang Y, Thomas HR, Frank MH, He Y, Xia R. TBtools: an integrative toolkit developed for interactive analyses of big biological data. Mol Plant. 2020;13(8):1194–02.
Wilkins MR, Gasteiger E, Bairoch A, Sanchez JC, Williams KL, Appel RD, Hochstrasser DF. Protein identification and analysis tools in the ExPASy server. Methods Mol Biol. 1999;112:531–52.
Chou KC, Shen HB. Plant-mPLoc: a top-down strategy to augment the power for predicting plant protein subcellular localization. PLoS ONE. 2010;5(6):e11335.
Tamura K, Stecher G, Kumar S. MEGA11: molecular evolutionary genetics analysis version 11. Mol Biol Evol. 2021;38(7):3022–27.
Letunic I, Bork P. Interactive tree of life (iTOL) v5: an online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021;49(W1):W293–6.
Hu B, Jin J, Guo AY, Zhang H, Luo J, Gao G. GSDS 2.0: an upgraded gene feature visualization server. Bioinformatics. 2015;31(8):1296–7.
Bailey TL, Johnson J, Grant CE, Noble WS. The MEME suite. Nucleic Acids Res. 2015;43(W1):W39–49.
Thompson JD, Higgins DG, Gibson TJ. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994;22(22):4673–80.
Lescot M, Déhais P, Thijs G, Marchal K, Moreau Y, Van de Peer Y, Rouzé P, Rombauts S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002;30(1):325–7.
Chatterjee A, Paul A, Unnati GM, Rajput R, Biswas T, Kar T, Basak S, Mishra N, Pandey A, Srivastava AP. MAPK cascade gene family in Camellia sinensis: in-silico identification, expression profiles and regulatory network analysis. BMC Genomics. 2020;21(1):613.
O’Rourke JA, Fu F, Bucciarelli B, Yang SS, Samac DA, Lamb JF, et al. The Medicago sativa gene index 1.2: a web-accessible gene expression atlas for investigating expression differences between Medicago sativa subspecies. BMC Genomics. 2015;16(1):502.
Luo D, Zhou Q, Wu Y, Chai X, Liu W, Wang Y, Yang Q, Wang Z, Liu Z. Full-length transcript sequencing and comparative transcriptomic analysis to evaluate the contribution of osmotic and ionic stress components towards salinity tolerance in the roots of cultivated alfalfa (Medicago sativa L). BMC Plant Biol. 2019;19(1):32.
Luo D, Wu Y, Liu J, Zhou Q, Liu W, Wang Y, Yang Q, Wang Z, Liu Z. Comparative transcriptomic and physiological analyses of Medicago sativa L. indicates that multiple regulatory networks are activated during continuous ABA treatment. Int J Mol Sci. 2018;20(1).
Long R, Wang H, Shen Y, Kang J, Zhang T, Sun Y, Zhang Y, Li M, Yang Q. Molecular cloning and functional analysis of a salt-induced gene encoding an RNA-binding protein in alfalfa. Mol Breed. 2014;34:1465–73.
Li Y, Zhang S, Dong R, Wang L, Yao J, van Nocker S, Wang X. The grapevine homeobox gene VvHB58 influences seed and fruit development through multiple hormonal signaling pathways. BMC Plant Biol. 2019;19(1):523.
Li S, Liu J, An Y, Cao Y, Liu Y, Zhang J, Geng J, Hu T, Yang P. MsPIP2;2, a novel aquaporin gene from Medicago sativa, confers salt tolerance in transgenic Arabidopsis. Environ Exp Bot. 2019;165:39–52.
Liu JH, Inoue H, Moriguchi TJE, Botany E. Salt stress-mediated changes in free polyamine titers and expression of genes responsible for polyamine biosynthesis of apple in vitro shoots. Environ Exp Bot. 2008;62(1):28–35.
Dahro B, Wang F, Peng T, Liu JH. PtrA/NINV, an alkaline/neutral invertase gene of Poncirus trifoliata, confers enhanced tolerance to multiple abiotic stresses by modulating ROS levels and maintaining photosynthetic efficiency. BMC Plant Biol. 2016;16:76.
Puckette MC, Weng H, Mahalingam RJPP, Biochemistry. Physiological and biochemical responses to acute ozone-induced oxidative stress in Medicago truncatula. Plant Physiol Biochem. 2007;45(1):70–9.
Jiang M, Zhang JJP, Physiology C. Effect of abscisic acid on active oxygen species, antioxidative defence system and oxidative damage in leaves of maize seedlings. Plant cell Physiol. 2001;42:1265–73.
Funding
This research was supported by the National Natural Science Foundation of China (32101426), the Ningxia Natural Science Foundation (2023AAC05019); and the Inner Mongolia Pratacultural Technology Innovation Center Co., Ltd (CCPTZX2023B04).
Author information
Authors and Affiliations
Contributions
Conceptualization, SL and BF; methodology, SL and SG; software, SG and XL; validation, SL, SG, XG and XW; investigation, SG, XG, XW, YL, and JW; resources, SL and BF; data curation, XW, YL and JZ; funding acquisition, SL and BF; writing—original draft preparation, SL; visualization, SL and SG; supervision and manuscript revisions, BF and XG. All authors have read and agreed to the published version of the manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
The authors declare that all methods were carried out in accordance with relevant guidelines and regulations. The sampling of plant material was performed in compliance with institutional, national, and international guidelines. We conducted the experimental research on cultivated alfalfa in accordance with the IUCN Policy Statement on Research Involving Species at Risk of Extinction and the Convention on the Trade in Endangered Species of Wild Fauna and Flora. Alfalfa seeds of ‘Zhongmu No. 1’ were provided by the Institute of Animal Science, Chinese Academy of Agricultural Sciences, China. The tobacco and Arabidopsis thaliana Columbia-0 planting materials were obtained from our laboratory (Laboratory of Forage Molecular Breeding, Ningxia University, China).
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Supplementary Material 1: Table S1.
Paralogous gene pairs in segmental duplication events of alfalfa MsBBX genes. Table S2. Collinear genes of MsBBXs between alfalfa and Arabidopsis, alfalfa and O. sativa, alfalfa and M. truncatula. Table S3. The functions of Cis-regulatory elements in MsBBX gene promoters. Table S4. The primers used in this study
Supplementary Material 2: Fig. S1.
Multiple sequence alignments of the conserved domains of the MsBBX proteins. Fig. S2. Distribution and location of the MsBBX gene family on alfalfa chromosomes. Fig. S3. Transcriptome ananlysis of the expression patterns of the MsBBX genes in six tissues of alfalfa: leaf, flower, pre-elongated stem, elongated stem, root and nodule. Fig. S4. Expression profiles of the MsBBX genes in alfalfa under drought and salt stress from transcriptome data. Fig. S5. Expression profiles of the MsBBX genes in alfalfa under ABA treatment from transcriptome data. Fig. S6. Predicted protein-protein interaction networks of MsBBX proteins based on the interactions of their orthologs in Arabidopsis. Fig. S7. Semi-quantitative RT-PCR gel image of MsBBX11 expression levels in WT and transgenic lines (OE1 and OE3). Fig. S8. Melting curves for all primers used in the qRT-PCR assays.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Li, S., Guo, S., Gao, X. et al. Genome-wide identification of B-box zinc finger (BBX) gene family in Medicago sativa and their roles in abiotic stress responses. BMC Genomics 25, 110 (2024). https://doi.org/10.1186/s12864-024-10036-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12864-024-10036-4