
Abstract
Background
Chilling injury of mung bean (Vigna radiata (L.)) during the blooming and podding stages is a major agricultural threat in Northeast China. Uniconazole (UNZ) can alleviate water deficit stress in soybean and waterlogging stress in mung bean. However, there has been no report on the effect of UNZ application on the growth and transcriptomic profile of mung bean under chilling stress.
Results
UNZ application before chilling stress at the R1 stage alleviated the decline in mung bean yield. UNZ delayed the decrease in leaf chlorophyll content under chilling stress at the R1 stage and accelerated the increase in leaf chlorophyll content during the recovery period. Eighteen separate RNA-Seq libraries were generated from RNA samples collected from leaves exposed to six different treatment schemes. The numbers of DEGs specific for UNZ treatment between D1 + S vs. D1 and D4 + S vs. D4 were 708 and 810, respectively. GO annotations showed that photosynthesis genes were obviously enriched among the genes affected by chilling stress and UNZ application. KEGG pathway enrichment analysis indicated that 4 pathways (cutin, suberin and wax biosynthesis; photosynthesis; porphyrin and chlorophyll metabolism; and ribosome) were downregulated, while plant–pathogen interaction was upregulated, by chilling stress. UNZ application effectively prevented the further downregulation of the gene expression of members of these 4 KEGG pathways under chilling stress.
Conclusions
UNZ application effectively delayed the decrease in photosynthetic pigment content under chilling stress and accelerated the increase in photosynthetic pigment content during the recovery period, thus effectively limiting the decline in mung bean yield. UNZ application effectively prevented the further downregulation of the gene expression of members of 4 KEGG pathways under chilling stress and increased mung bean tolerance to chilling stress.
Similar content being viewed by others


Background
Mung bean (Vigna radiata L.) is an important and valuable legume crop due to its nutritional and health benefits. Mung beans contain significant amounts of proteins and phytochemicals with beneficial activities [1]. According to the Food and Agriculture Organization of the United Nations, in 2019, the global planting area of mung bean was 1649.7 thousand hectares, and the output was 26.982 million tons. Mung bean is one of the most important legume crops in China and is mainly produced in the Huang Huai River Basin and in Northeast China [2]. Mung bean originated in the tropics and is a thermophilic crop.
Cold stress can be classified as chilling (0–15 °C) or freezing (< 0 °C) stress, and both of these can cause significant crop loss. Chilling injury is a major agricultural hazard in the summer in Northeast China [3]. During the period from soybean sowing to maturity, periodic low-temperature chilling injury often occurs, mainly in the blooming and podding stages, which often reduces yield and quality [4,5,6,7]. Cold stress responses are influenced by the exposure duration [8], plant species [9] and plant development stage [7]. The exposure of mung bean to chilling results in high and irreversible electrolyte leakage [10]. The susceptibility of mung bean to chilling stress is related to the differential expression of cold-related genes [11, 12].
It has been well documented that plant growth regulators (PGRs) play important roles in crop production and in resistance to environmental stresses. Uniconazole (S-3307, UNZ), a plant growth retardant, has been increasingly applied in crops to increase their output and quality [13,14,15]. In a previous study, soybean plants were treated with UNZ at 50 mg L−1 at the beginning of bloom and then exposed to water deficit stress for 7 d. The chlorophyll content and photosynthesis rate were decreased by water deficit stress but remained higher in UNZ-treated stressed plants than in the stressed control. Biomass accumulation and seed yield were also increased in plants treated with UNZ compared with controls [15]. Mung bean plants were treated with UNZ at 50 mg L−1 at the beginning of bloom and at the beginning of seed development and then exposed to waterlogging for 5 days. The chlorophyll content and photosynthetic rate were decreased by waterlogging stress but remained higher in UNZ-treated plants than in the stressed control. Thus, UNZ could effectively alleviate yield reduction [16].
The genome of mung bean has been recently sequenced [17], which is a valuable resource for research on stress tolerance genes in plants. RNA sequencing (RNA-seq) analysis is a powerful method for transcriptome analysis that has been increasingly used to study transcriptomic responses to chilling/cold stress in many legume species [15, 18,19,20,21]. Transcriptome analysis has been conducted to analyze desiccation tolerance, adventitious rooting and bruchid resistance in mung bean [22,23,24]; however, the effect of UNZ on the growth and the transcriptomic profile of mung beans under chilling stress has not yet been investigated. In this work, mung bean plants were predicted to be harmed by chilling stress at the blooming and podding stages, and the application of UNZ could alleviate stress damage in soybean. The yield, photosynthetic pigment content and relevant transcriptome changes in mung bean in response to UNZ under low-temperature stress were studied. Thus, this study aimed to identify differentially expressed genes (DEGs) that influenced yield and photosynthetic pigments via transcriptome analysis, which would be useful for the application of UNZ in the production of mung bean. At the same time, it provides some reference information for mung bean cultivation in Heilongjiang and other cold regions.
Results
Effects of chilling stress and UNZ application on yield and yield components in mung bean seedlings
Pod number per plant and particle number per plant are two important yield components. The effect of UNZ on yield and yield components in mung bean under chilling stress at R1 stage in 2017 is shown in Table 1. The yields of the two mung bean varieties were significantly lower than those of CK after chilling treatment at stage R1. The pod number per plant, particle number per plant and yield followed the same pattern in the 2 varieties, specifically, CK > D1 > D2 > D3 > D4. The yield loss of Lvfeng 2 was more severe, which indicated that the different genotypes of mung bean had different tolerances to chilling stress. The yield loss in 2017 was similar to that in 2016 (data not shown). However, spraying UNZ at the R1 stage effectively alleviated the decline in mung bean yield.
Effects of chilling stress and UNZ application on photosynthetic pigments in mung bean at the R1 stage
Chlorophylls and carotenoids are the main photosynthetic pigments in plants. Photosynthetic pigments are involved in the process of absorbing and transferring energy. The change in photosynthetic pigment content can reflect the degree of chilling stress. The contents of chlorophyll a (Chl a) and total chlorophyll (Chl (a + b)) of Lvfeng 2 (L2) in L2-D and L2-D + S were significantly lower than those in the control from 1 to 8 d of treatment. However, the contents of chlorophyll b (Chl b) in L2-D and L2-D + S were significantly lower than those in the control from 3 to 7 d of treatment. The contents of total carotenoids (Car) in L2-D and L2-D + S were significantly lower than those in the control from 2 to 7 d treatment. The contents of Chl b and Car reached the normal level, that is, equivalent to the control, after 4 d of recovery. The contents of Chl a and Chl (a + b) in L2-D were significantly lower than those in L2-D + S from 1 to 8 d of treatment. However, the contents of Chl b in L2-D were significantly lower than those in L2-D + S from 3 to 7 d of treatment, and the contents of total Car in L2-D were significantly lower than those in L2-D + S from 2 to 4 d of treatment. Generally, the contents of Chl a, Chl b, Chl (a + b) and total Car decreased gradually with the extension of chilling stress and increased with the extension of recovery time but were always lower than those in the control (Fig. 1). The changes in the contents of Chl a, Chl b, total Chl and Car in Lvfeng 5 (Fig. S1) in L2-D and L2-D + S were similar to those in Lvfeng 2. Exogenous application of UNZ can effectively delay the decreases in the photosynthetic pigment content of leaves under chilling stress at the R1 stage and accelerate the increase in photosynthetic pigment content in leaves during the recovery period.

Effects of uniconazole on photosynthetic pigments in Lvfeng 2 under chilling stress at the R1 stage. Note: Data in the figure are the mean ± SD of four replicates. L2-CK, plants grown in the natural environment; L2-D, plants grown in the chilling environment; L2-D + S, plants grown in the chilling environment + 50 mg·L-1 UNZ; 1–4, time for chilling stress; 5–8: time for recovery in the natural environment. Different lowercase letters represent significant differences (p < 0.05) between the treatment and control
Transcriptome profiles of mung bean leaves under chilling stress and UNZ application
An overview of the RNA-seq reads derived from the eighteen libraries is presented in Table 2. A total of 133.08 Gb of clean reads was obtained, with an average of 22.18 Gb of reads for each sample. The average Q30 was 94.28%, and the GC content of each sample was uniform and stable, which indicated that the sequencing results were highly accurate.
The clean reads of each library were mapped to the reference genome sequence of mung bean. The mapped ratio of the libraries ranged from 92.22% to 93.28%, and the uniquely mapped ratio of the libraries ranged from 87.24% to 90.50%. The data were qualified for subsequent analysis. The correlation coefficient of the three biological repeats in each treatment was more than 0.9 (Fig. S2).
The numbers of DEGs between different pairs of treatments with FDR < 0.01 and fold change ≥ 2 are shown in Table S2. The number of DEGs between D1 + S and CK1 was 4 905, while the number of DEGs between D1 and CK1 was 4 025. The number of DEGs decreased to 3 266 in D4 + S vs. CK4 compared to the 4 023 DEGs in D4 vs. CK4.
The numbers of shared DEGs in mung bean leaves among these treatments were 11,134 and 11,000 for 1 day of chilling and 4 days of chilling stress, respectively. which may reflect that the expression of these genes was not influenced by UNZ or chilling stress. The numbers of DEGs specific to UNZ treatment between D1 + S and D1 and between D4 + S and D4 were 708 and 810, respectively (Fig. 2).

Venn diagram of the differentially expressed genes of mung bean leaves under chilling stress and UNZ application. Note: CK, plants grown in the natural environment; D, plants grown in the chilling environment; D + S, plants grown in the chilling environment + 50 mg·L-1 UNZ; 1 and 4, 1 d and 4 d of chilling stress
GO enrichment analysis of DEGs in mung bean leaves under chilling stress and UNZ application at the R1 stage
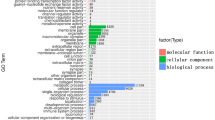
GO annotations were used to classify the possible functions of the mung bean genes. GO functional enrichment indicated that 1170, 1184, 2017, and 1385 DEGs in D1-CK1, D1 + S-CK1, D4-Ck4 and D4 + S-CK4, respectively, were classified into the three GO categories of biological process (BP), cellular component (CC), and molecular function (MF). The significantly enriched GO categories in CC, BP and MF are shown in Table 3. Only GO terms in the MF category were significantly enriched in D1 vs. CK1 and D1 + S vs. CK1 among the three categories. The number of enriched GO terms in the MF category was greater in D1 + S vs. CK1 than in D1 vs. CK1, which implies that the application of UNZ plays an important role in regulating MF gene expression under chilling stress. In D4 vs. CK4 and D4 + S vs. CK4, GO terms in each of the three categories were significantly enriched, and the number of GO terms in the CC category was the highest among the 3 categories. It is obvious that the cell components related to photosynthesis were most influenced by chilling stress.
KEGG enrichment analysis of DEGs in mung bean leaves under chilling stress and UNZ application at the R1 stage
For the KEGG pathway enrichment analysis [25], pathways with a Q value ≤ 0.05 were regarded as significantly changed in response to chilling stress with or without UNZ treatment. The top 20 enriched KEGG pathways are presented in Fig. 3. The pathways with significant enrichment among the DEGs were cutin, suberin and wax biosynthesis (Vra00073) in D1 vs. CK1; photosynthesis (Vra00195) in D4 vs. CK4; cutin, suberin and wax biosynthesis (Vra00073) and plant vs. pathogen interaction (Vra04626) in D1 + S vs. CK1; and photosynthesis (Vra00195), ribosome (Vra03010) and porphyrin and chlorophyll metabolism (Vra00860) in D4 + S vs. CK4.

Functional analysis of DEGs based on KEGG pathway annotations. Pathways with a Q-value ≤ 0.05 that were significantly enriched in DEGs in the comparison between the treatment and control at the flowering stages were analyzed with the KEGG database. The degrees of KEGG enrichment can be measured by the gene ratio, padj (multiple testing correction-adjusted p values), and number of genes enriched in this pathway. The right y-axis represents the KEGG pathway, and the x-axis shows the gene ratio, which denotes the ratio of the number of DEGs to the number of annotated genes enriched in this pathway
Differential expression of plant–pathogen interaction pathway genes
The expression levels of all plant–pathogen interaction genes except calcium-dependent protein kinase SK5 were upregulated after chilling stress with or without UNZ in all four treatments. These genes are important Ca2+ signaling genes, including 7 calcium-binding protein genes, 3 calcium-dependent protein kinase genes and 5 calmodulin calmodulin-like genes.
Two heat shock proteins, heat shock cognate protein and endoplasmin, were upregulated. One mitogen-activated protein kinase, pathogenesis-related gene transcriptional activator, respiratory burst oxidase homolog protein, somatic embryogenesis receptor kinase and WRKY transcription factor each were also upregulated.
The log2(fold change) (log2(FC)) values of most calmodulin and calmodulin-like proteins were positive in D1 + S vs. D1 and D1 + S vs. D1. In particular, calmodulin in D1 + S vs. D1 and calmodulin-like protein 11 in D1 + S vs. D1 and D4 + S vs. D4 were notably upregulated (log2(FC) > 1), indicating that UNZ treatment before chilling stress upregulated the expression of some Ca2+ signaling genes (Table 4).
Differential expression of ribosomal proteins
The ribosome is a large assembly of proteins and ribosomal RNAs (rRNAs) that functions to translate messenger RNAs (mRNAs) into proteins. Ribosome genes were significantly enriched in the D1 + S treatment. The expression levels of DEGs in this pathway are shown in Table S3. The expression levels of all ribosome genes were notably downregulated (log2(FC) > 1) after chilling stress with or without UNZ, except for 40S ribosomal protein S3a and 60S ribosomal protein L10-like, which were notably upregulated. The log2(FC) values of 23 of 26 genes were positive in D1 + S vs. D1, indicating that UNZ treatment effectively prevented further downregulation of the expression of ribosomal genes, while the log2(FC) values of only 11 of 26 genes were positive in D4 + S vs. D4, indicating that UNZ treatment did not effectively prevent ribosomal genes from being further downregulated with the extension of chilling stress.
Differential expression of cutin, suberin and wax biosynthesis genes
Cutin, suberin and wax, which are involved in cell wall structure, have the principal function of establishing the boundary between the cell and the environment [26]. The expression levels of all differentially expressed cutin, suberin and wax genes were obviously downregulated, except cytochrome P450 86A8, which was upregulated in the 4 treatments, indicating that the synthesis of cutin, suberin and wax was decreased (Table S4). Only peroxygenase 4 in D1 + S vs. D1 and ECERIFERUM 1 in D4 + S vs. D4 were obviously downregulated, indicating that UNZ treatment effectively prevented the expression of these two genes.
Differential expression of chlorophyll and photosynthesis biosynthesis genes
Chlorophyll is an important photosynthetic pigment in the chloroplasts of plants, and the metabolism of chlorophyll is an important factor in determining crop yield. The contents of Chl a, Chl b, total CHL and Car in leaves were found to increase after UNZ application. To investigate whether chlorophyll synthesis-related genes were involved in the increase in chlorophyll content, we further studied the expression patterns of regulatory enzymes involved in chlorophyll biosynthesis. One significantly enriched pathway among the DEGs was porphyrin and chlorophyll metabolism. The genes involved in this pathway and their expression are listed in Table 5. There are 17 enzymes required for chlorophyll biosynthesis, from glutamyl-tRNA to chlorophyll b [27, 28]. Ten genes were differentially expressed, of which 8 genes were downregulated in the 4 treatments. Staygreen protein and chlorophyll b reductase, which are associated with chlorophyll degradation, were found to be upregulated [29, 30]. The log2(FC) values of 8 and 9 of these ten genes were positive in D1 + S vs. D1 and D4 + S vs. D4, respectively, indicating that UNZ treatment prevented further downregulation of the expression of chlorophyll biosynthesis genes.
The majority of photosynthetic energy is harnessed via linear electron flow involving light-stimulated electron transfer between two reaction centers, PSI and PSII [31]. According to the results of transcriptome sequencing, 31 genes that were differentially regulated in the 4 treatments were annotated as photosystem genes, and their expression levels are listed in Table 6. Nine genes were detected for PSI eleven genes, including 3 oxygen-evolving enhancer proteins, were detected for PSII; and nine genes, including cytochrome b6-f complex iron-sulfur subunit, plastocyanin and ferredoxin-NADP, ferredoxin reductase, photosynthetic NDH subunit of luminal location and ATP synthase, were detected for photosynthetic electron transfer. The log2(FC) values of 15 and 27 of these 31 genes were positive in D1 + S vs. D1 and D4 + S vs. D4, respectively, indicating that UNZ treatment prevented further downregulation of the expression of photosynthetic genes under chilling stress and improved the cold tolerance of mung bean.
Quantitative Real-time PCR verification
Ten randomly selected cold tolerance unigenes were selected to verify the reliability and accuracy of our transcriptome data using qRT–PCR. The gene IDs and their functional annotations are listed in Table S1. The expression patterns determined by qRT–PCR were compared with those of the RNA-seq assay. The qRT–PCR and RNA-seq results showed a positive correlation coefficient (Pearson coefficient R2 = 0.911) (Fig. 4).

The correlation of RNA-seq and qRT–PCR results
Discussion
Effects of chilling stress and UNZ application on yield and photosynthetic pigments in mung bean
Low-temperature stress can cause changes in membrane phase and membrane permeability in plants, and it can also destroy enzyme systems, cause metabolic disorders, reduce energy supplies, inhibit photosynthesis, promote the accumulation of toxic substances, and affect plant growth and biomass production [32]. Different mung bean varieties differ in their tolerance to cold stress [2, 12]. UNZ has been increasingly applied in crops to increase crop output and quality [13,14,15, 33]. Two major cultivated varieties, Lvfeng 2 and Lvfeng 5, were selected to evaluate the influence of UNZ application under chilling stress. Our results demonstrated that chilling stress for 1 day and 4 days significantly decreased the yields and chlorophyll content in the two mung bean varieties, although different genotypes of mung bean had different tolerances to chilling stress. Soybean plants under water deficit stress and mung bean plants under waterlogging stress were treated with UNZ, and the chlorophyll content and seed yield were increased compared to those of the control [16, 34]. Our results were consistent with the findings in soybean and mung bean plants. Application of UNZ before chilling stress increased the yields and chlorophyll content significantly in two mung bean varieties compared to the chilling stress treatments. These results indicated that the application of UNZ alleviated chilling stress and that UNZ-induced tolerance to stress was related to changes in photosynthesis and thus increased the yield of mung bean.
Transcription patterns of chlorophyll and photosynthesis in response to chilling stress and UNZ application
Chlorophyll is an important photosynthetic pigment in the chloroplasts of plants that performs the essential process of harvesting light energy in antenna systems [35]. Chlorophyll metabolism is an important factor in determining the photosynthetic rate and affects crop yield. To date, several studies have focused on the involvement of UNZ in photosynthesis [13, 14, 16, 34], but the expression of chlorophyll synthesis genes after the application of UNZ has been studied only in duckweed [14]. In this study, a genome-wide transcriptomic analysis method was used to investigate the metabolism of key enzymes involved in the chlorophyll biosynthesis pathway in mung bean. Eight of the ten DEGs required for chlorophyll biosynthesis from glutamyl-tRNA to chlorophyll b [27] were dramatically downregulated in the 4 treatments, and two chlorophyll degradation genes were upregulated [28, 29]. The log2(FC) of most chlorophyll biosynthesis genes was positive in D1 + S vs. D1 and D4 + S vs. D4, indicating that UNZ treatment prevented further downregulation of the expression of chlorophyll biosynthesis genes by chilling stress. UNZ enhanced the chlorophyll content and upregulated the expression of key enzymes involved in the chlorophyll biosynthesis pathway. These results were consistent with the upregulation of chlorophyll biosynthesis genes after UNZ application [14, 16, 36, 37].
Transcription patterns of KEGG pathways in response to chilling stress and UNZ application
Mung bean plants often upregulate the expression of some cold-related genes (CORs) to cope with low temperatures. The number of upregulated CORs with a fold change ≥ 3 or downregulated CORs with a fold change ≤ 0.3 genes in mung bean-resistant varieties was 2 times greater than that in susceptible varieties. These CORs are involved in photosynthesis, cellular redox homeostasis, disease resistance and membrane stabilization [11]. Mung bean cell protection and stress-regulated genes were found to respond to low temperatures. More than 489 uniESTs among 1,198 ESTs were involved in metabolic activity, photomorphogenesis, photosynthesis and/or developmental programs [12]. These studies were not conducted at the global transcriptomic level. The use of transcriptomics to quantify the nearly complete set of cellular transcripts allows quantitative and qualitative differences in gene expression to be determined for a specific developmental stage or stress condition [38]. In this study, RNA-seq was used to study the molecular mechanisms underlying UNZ-induced low-temperature tolerance in mung bean. The significantly enriched pathways were cutin, suberin and wax biosynthesis; plant–pathogen interaction; ribosome; porphyrin and chlorophyll metabolism; and photosynthesis.
It is well known that Ca2+ acts as a key messenger in regulating cold stress signal transduction pathways [39]. Here, the expression of important Ca2+ signaling genes, including reactive oxygen species (ROS), Ca2+-dependent protein kinases (CDPKs), mitogen-activated protein kinase (MAPK) cascades and transcription factors (TFs), was found to be upregulated after chilling stress with or without UNZ treatment. The results were consistent with those in other plants exposed to low temperature [15, 40,41,42,43]. UNZ treatment before chilling stress upregulated the expression of some Ca2+ signal genes and promoted the tolerance of mung bean to chilling stress.
The plant cuticle is the first protective barrier against environmental stress. Most cuticle-associated genes, including those involved in cuticle lipid synthesis, export of cuticular lipids, and regulation of plant cuticle development, are upregulated under low-temperature treatment [44]. In our experiment, most of these genes were downregulated after chilling stress with or without UNZ application, but UNZ treatment effectively prevented further downregulation of the expression of these genes. Our results were obviously different from those showing that most cuticle-associated genes exhibited higher expression levels under cold conditions in Thellungiella salsuginea [44]. The reason may be that the plants were treated with different temperatures and at different stages. Thellungiella salsuginea was stressed by exposure to 4 ℃ at the preflowering stage, whereas in our experiment, mung beans were stressed by exposure to 15 ℃ at the flowering stage.
Conclusions
The application of UNZ affected yield and photosynthetic parameters. UNZ effectively delayed the decrease in the photosynthetic pigment content of leaves under chilling stress and accelerated the increase in photosynthetic pigment content in leaves during the recovery period, thus effectively alleviating the decline in mung bean yield. Genes in four pathways (cutin, suberin and wax biosynthesis; photosynthesis; porphyrin and chlorophyll metabolism; and ribosome) were downregulated, while genes related to plant–pathogen interaction were upregulated, by chilling stress. UNZ treatment effectively altered the expression patterns of these genes and increased mung bean tolerance to chilling stress.
Methods
Plant materials and UNZ treatments
Two cultivars of mung bean, Lvfeng 2 and Lvfeng 5, were provided by the germplasm bank of the National Coarse Cereals Engineering Technology Research Center for use as materials. The two cultivars were planted in pots and grown to the outset of the flowering stage (R1) in a greenhouse at Heilongjiang Academy of Agricultural Sciences in 2017. Three treatments were established, and each treatment was repeated 4 times. The plants were sprayed with water and maintained at the natural temperature for 4 days (CK); then, the plants were sprayed with water, and 36 h later, the plants were shifted to 15 ℃ for 1 d, 2 d, 3 d or 4 d (D1, D2, D3, D4) and then returned to the warmer temperature to grow for 4 days; the plants were sprayed with 50 mg·L−1 UNZ (Jiangxi Nongda Ruite Chemical Technology Co., Ltd., CAS: 76714–83-5), and 36 h later, the plants were shifted to 15 ℃ to grow for 1 d, 2 d, 3 d and 4 d (D1 + S, D2 + S, D3 + S, D4 + S) and then returned to the warmer temperature to grow for 4 d [6, 7]. The plants of the 3 treatments were grown under natural conditions until maturity. The lowest external temperature of each day during the 4 days of chilling stress was above 20 °C, as monitored by an EM50 micro meteorological monitoring system.
Yield components and chlorophyll measurement
After mung bean was grown to maturity, the pod number per plant, particle number per plant, and yield were measured at the maturity stage (R8). The mung bean grain was dried to constant weight, and the final yield was calculated according to the determined 14% water content. Each treatment consisted of 4 replicates with 10 plants per replicate. The mung bean leaves at each day of chilling stress and recovery were taken as the materials to determine the chlorophyll content. Fresh leaves (0.1 g) were immersed in 10 mL alcohol and kept at room temperature for 24 h and shielded from light for 1 d after chilling stress. The leaf chlorophyll a, chlorophyll b and carotenoid concentrations in the supernatant of the solution were measured using a spectrophotometer at 663 and 645 nm and 470 nm, respectively [45, 46].
RNA extraction, cDNA library construction and sequencing
Lvfeng 2 was used as the experimental material. There were 6 treatments (CK1, D1 and D1 + S and CK4, D4 and D4 + S), which are described in the plant materials and UNZ treatments section. Each treatment consisted of 3 replicates. Sample collection was carried out between 8:00 and 9:00 a.m. Mixed samples of three leaves each in the CK, D and D + S treatments were subjected to RNA extraction (Invitrogen TRIzol Reagent of RNA extraction kit 15,596,018). A total of eighteen libraries were constructed using a NEBNext Ultra RNA Library Prep Kit for Illumina (NEB, USA) and sequenced using an Illumina HiSeqTM2000 (Beijing Biomarker Technologies Co.).
Assembly and functional annotation
The raw sequencing reads were cleaned by removing adaptors and low-quality reads. After filtering, Bowtie software (version 2.2.5) was used to map the clean reads to the mung bean (VC1973A) reference genome (463.638 Mbp) [47], and then RSEM [48] was used to estimate the expression levels with the FPKM value. To identify DEGs, the criteria applied were an FDR (false discovery rate) less than 0.01 and an absolute value of log2 ratio of at least 1. The screened DEGs were analyzed mainly by GO function (http://geneontology.org/system [49] and KEGG pathway enrichment (https://www.kegg.jp/kegg/kegg1.html) [25]. GO annotations of DEGs and GO functions mapped to the corresponding secondary features based on unigene GO annotation were extracted, and a histogram was drawn [49]. The KEGG pathway enrichment analysis was implemented via KOBAS2.0 (http://kobas.cbi.pku.edu.cn/home.do) [25].
Quantitative real-time PCR verification
Quantitative real-time PCR (qRT–PCR) was performed to examine the expression patterns shown by the RNA-seq analysis. Ten genes were selected on the basis of their potential functions in RNA-seq. The sequences of each primer are shown in Table S1. RNA (1 μg) from each treatment was treated with DNAse I (Invitrogen), translated into first-strand cDNA with TransScript One-Step gDNA Removal (Super Script) and cDNA Synthesis SuperMix (Transgene Biotech), and then the cDNA was stored at − 20 °C for subsequent analysis. Each PCR contained a 20 μl mixture consisting of 2 μl of cDNA, 10 μl of 2 × TransStart Top Green qPCR SuperMix, and 0.4 μl of the forward and reverse primers. All qRT-PCRs were performed in three technical replicates in a Bio–Rad CFX96 thermocycler and performed in two steps: predenaturation for 2 min at 94 °C, followed by 45 cycles of denaturation for 2 s at 94 °C and annealing/extension for 15 s at 60 °C. The relative expression level was calculated by the 2−ΔΔCt method [50] with the actin gene as an internal standard. Each measurement included three biological and three technical replicates.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from Mendeley Data, v1 (http://dx.doi.org/10.17632/v9b8pcxk56.1).
Abbreviations
- UNZ:
-
Uniconazole
- DEGs:
-
Differentially expressed genes
- Chl a:
-
Chlorophyll a
- Chl b:
-
Chlorophyll b
- Chl (a + b):
-
Total chlorophyll
- Car:
-
Carotenoids
References
Tang D, Dong Y, Ren H, Li L, He C. A review of phytochemistry, metabolite changes, and medicinal uses of the common food mung bean and its sprouts (Vigna radiata). Chem Cent J. 2014;8:4.
Wang LX, Cheng XZ, Wang SH. Advances in research on genetic resources, breeding and genetics of mung bean (Vigna radiata L.). Acta Agron Sin. 2009;42:1519–27.
Fang YH, Li J, Zhou F. Research progress and prospect of summer freezing damage’s impact factors in northeast China. Chinese Agri Sci Bull. 2013;29:194–200.
Wang P, Song HX, Ma SY, Tao D, Yuan Y, Song LQ, et al. Effects of low temperature at blooming on podding and seed filling in soybean varieties. Chinese J Oil Crop Sci. 2000;22:31–3.
Li HN, Zhu LJ, Li MQ, Jiang LX, Ren CY, Gao XN. Construction of soybean chilling damage indicator and its evolution characteristics in northeast China. J Appl Meteorol Sci. 2021;32:491–503.
Funatsuki H, Matsuba S, Kawaguchi K, Murakami T, Sato Y. Methods for evaluation of soybean chilling tolerance at the reproductive stage under artificial climatic conditions. Plant Breed. 2004;123:558–63.
Ohnishi S, Miyoshi T, Shirai S. Low temperature stress at different flower developmental stages affects pollen development, pollination, and pod set in soybean. Environ Exp Bot. 2010;69:56–62.
Ercoli L, Mariotti M, Masoni A, Arduini I. Growth responses of sorghum plants to chilling temperature and duration of exposure. Eur J Agron. 2004;21:93–103.
Carvallo MA, Pino MT, Jeknić Z, Zou C, Doherty CJ, Shiu SH, et al. A comparison of the low temperature transcriptomes and CBF regulons of three plant species that differ in freezing tolerance: Solanum commersonii, Solanum tuberosum, and Arabidopsis thaliana. J Exp Bot. 2011;62:3807–19.
Chang MY, Chen SL, Lee CF, Chen YM. Cold-acclimation and root temperature protection from chilling injury in chilling-sensitive mung bean (Vigna radiata L.) seedlings. Bot Bull Acad Sin. 2001;42:53–60.
Chen LR, Ko CY, Folk WR, Lin TY. Chilling susceptibility in mung bean varieties is associated with their differentially expressed genes. Bot Stud. 2017;58:7.
Chen LR, Markhart AH, Shanmugasundaram S, Lin TY. Early developmental and stress responsive ESTs from mung bean, Vigna radiata (L.) Wilczek, seedlings. Plant Cell Rep. 2008;27:535–52.
Wu C, Sun J, Zhang A, Liu W. Dissipation and enantioselective degradation of plant growth retardants paclobutrazol and Uniconazole in open field, greenhouse, and laboratory soils. Environ Sci Technol. 2013;47:843–9.
Liu Y, Fang Y, Huang MJ, Jin YL, Sun JL, Tao X, et al. Uniconazole induced starch accumulation in the bioenergy crop duckweed (Landoltia punctata) I: transcriptome analysis of the effects of uniconazole on chlorophyll and endogenous hormone biosynthesis. Biotechnol Biofuels. 2015;8:57.
Liu C, Yang X, Yan Z, Fan YJ, Feng GJ, Liu DJ. Analysis of differential gene expression in cold-tolerant vs. cold-sensitive varieties of snap bean (Phaseolus vulgaris L.) in response to low temperature stress. Genes Genom. 2019;41:1445–55.
Zhang MC, Duan LS, Tian XL, He ZP, Li JM, Wang BM, et al. Uniconazole-induced tolerance of soybean to water deficit stresses in relation to changes in photosynthesis, hormones and antioxidant system. J Plant Physiol. 2007;164:709–17.
Kang YJ, Kim SK, Kim MY, Lestari P, Kim KH, Ha BK, et al. Genome sequence of mung bean and insights into evolution within Vigna species. Nat Commun. 2014;5:5443.
Aghamirzaie D, Batra D, Heath LS, Schneider A, Grene R, Collakova E. Transcriptome-wide functional characterization reveals novel relationships among differentially expressed transcripts in developing soybean embryos. BMC Genom. 2015;16:928.
Xu SC, Liu N, Mao WH, Hu QZ, Wang GF, Gong YM. Identification of chilling-responsive microRNAs and their targets in vegetable soybean (Glycine Max L.). Sci Rep. 2016;6:26619.
Tan H, Huang H, Tie M, Tang Y, Lai Y, Li H. Transcriptome profiling of two asparagus bean (Vigna unguiculata subsp. sesquipedalis) cultivars differing in chilling tolerance under cold stress. PLoS One. 2016;11:e0151105.
Yin X, Hiraga S, Hajika M, Nishimura M, Komatsu S. Transcriptomic analysis reveals the flooding tolerant mechanism in flooding tolerant line and abscisic acid treated soybean. Plant Mol Biol. 2017;93:1–18.
Li SW, Leng Y, Shi RF. Transcriptomic profiling provides molecular insights into hydrogen peroxide-induced adventitious rooting in mung bean seedlings. BMC Genom. 2017;18:1–23.
Liu MS, Kuo CY, Ko CY, Wu DC, Li KY, Lin WJ. Genomic and transcriptomic comparison of nucleotide variations for insights into bruchid resistance of mung bean (Vigna radiata [L.] R. Wilczek). BMC Plant Biol. 2016;16:46.
Tian X, Li S, Liu Y, Liu X. Transcriptomic profiling reveals metabolic and regulatory pathways in the desiccation tolerance of Mung bean (Vigna radiata [L.] R. Wilczek). Front Plant Sci. 2016;21:1921.
Xie C, Mao X, Huang J, Wei L. A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011;39(Web Server):W316–22.
Beisson F, Li-Beisson Y, Pollard M. Solving the puzzles of cutin and suberin polymer biosynthesis. Curr Opin Plant Biol. 2012;15:329–37.
Beale SI. Enzymes of chlorophyll biosynthesis. Photosynth Res. 1999;60:43–73.
Cornah JE, Terry MJ, Smith AG. Green or red: what stops the traffic in the tetrapyrrole pathway? Trends Plant Sci. 2003;8:224–30.
Horie Y, Ito H, Kusaba M, Tanaka R, Tanaka A. Participation of Chlorophyll b reductase in the initial step of the degradation of light-harvesting chlorophyll a/b-protein complexes in Arabidopsis. J Biol Chem. 2009;284:17449–56.
Park SY, Yu JW, Park JS, Li JJ, Yoo SC, Lee NY, et al. The senescence-induced staygreen protein regulates chlorophyll degradation. Plant Cell. 2007;19:1649–64.
Livingston AK, Cruz JA, Kohzuma K, Dhingr A, Kramera DM. An Arabidopsis mutant with high cyclic electron flow around photosystem I (hcef) involving the NADPH dehydrogenase complex. Plant Cell. 2010;22:221–33.
Theocharis A, Clément C, Barka EA. Physiological and molecular changes in plants grown at low temperatures. Planta. 2012;235:1091–105.
Liu CJ, Feng NJ, Zheng DF, Cui HQ, Sun FD, Gong XW. Uniconazole and diethyl aminoethyl hexanoate increase soybean pod setting and yield by regulating sucrose and starch content. J Sci Food Agr. 2018;99:748–58.
Yu Q, Feng NJ, Wang SY, Zuo GQ, Zheng DF. Effects of S3307 on the photosynthesis and yield of mung bean at R1 and R5 stages under waterlogging stress. Acta Agron Sin. 2019;45:1080–9.
Fromme P, Melkozernov A, Jordan P, Krauss N. Structure and function of photosystem I: interaction with its soluble electron carriers and external antenna systems. FEBS Lett. 2003;555:40–4.
Ashraf M, Harris PJC. Photosynthesis under stressful environments: an overview. Photosynthetica. 2013;51:163–90.
Allen DJ, Ort DR. Impacts of chilling temperatures on photosynthesis in warm-climate plants. Trends Plant Sci. 2001;6:36–42.
Wang Z, Gerstein M, Snyder M. RNA-Seq: a revolutionary tool for transcriptomics. Nat Rev Genet. 2009;10:57–63.
Reddy AS, Ali GS, Celesnik H, Day IS. Coping with stresses: roles of calcium- and calcium/calmodulin-regulated gene expression. Plant Cell. 2011;23:2010–32.
Ren L, Sun J, Chen S, Gao J, Dong B, Liu Y, et al. A transcriptomic analysis of Chrysanthemum nankingense provides insights into the basis of low temperature tolerance. BMC Genom. 2014;15:844.
Wang XC, Zhao QY, Ma CL, Zhang ZH, Yang YJ. Global transcriptome profiles of Camellia sinensis during cold acclimation. BMC Genom. 2013;14:415.
Thomashow MF. Role of cold-responsive genes in plant freezing tolerance. Plant Physiol. 1998;118:1–8.
Yokotani N, Sato Y, Tanabe S, Chujo T, Shimizu T, Okada K, et al. WRKY76 is a rice transcriptional repressor playing opposite roles in blast disease resistance and cold stress tolerance. J Exp Bot. 2013;64:5085–97.
He J, Tang S, Yang D, Chen Y, Ling L, Zou Y, et al. Chemical and transcriptomic analysis of cuticle lipids under cold stress in Thellungiella salsuginea. Int J Mol Sci. 2019;20:4519.
Lichtenthaler HK. Chlorophylls and carotenoids-pigments of photosynthetic biomembranes. Methods Enzymol. 1987;148C:350–82.
Strain HH, Svec WA. Extraction, separation, estimation and isolation of the chlorophylls. The Chlorophylls. 1966;1:22–66.
Langmead B, Trapnell C, Pop M, Salzberg SL. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009;10:R25.
Li B, Dewey CN. RSEM: accurate transcript quantification from RNA-seq data with or without a reference genome. BMC Bioinformatics. 2011;12:323.
Ye J, Fang L, Zheng H, Zhang Y, Chen J, Zhang Z, et al. WEGO: a web tool for plotting GO annotations. Nucleic Acids Res. 2006;34(Web Server):W293–7.
Livak K, Schmittgen T. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods. 2001;25(4):402–8.
Acknowledgements
Not applicable.
Funding
This research was supported by the National Natural Science Foundation of China (31871576) and the Project of Enhancing School with Innovation of Guangdong Ocean University (230420006).
Author information
Authors and Affiliations
Contributions
HHQ and ZDF contributed to the bioinformatics analysis and writing of the manuscript; FNJ designed the experiment and determined the chlorophyll content; SXF participated in the chilling stress and UNZ treatment and RNA-seq experiments; ZLM cultivated the mung bean plants and determined the yields; ZDF designed the experiment and provided funding. The author(s) read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The study was performed in accordance with relevant institutional, national, and international guidelines and legislation.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Table S1.
Selected genes and primers forquantitative qRT–PCR.
Additional file 2: Table S2.
Effect of uniconazole on DEGs in mungbean leaves under chilling stress at the Rl stage.
Additional file 3: Table S3.
Expression level of genes associatedwith ribosome (Vra03010) in mung bean.
Additional file 4: Table S4.
Expression level of genes associatedwith cutin, suberin and wax biosynthesis(Vra00073) in mung bean.
Additional file 5: Figure S1.
Effects of uniconazole on photosynthetic pigments in Lvfeng 5 under chillingstress at the R1 stage.
Additional file 6: Figure S2.
Pearsoncorrelation between samples.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Hu, H., Feng, N., Shen, X. et al. Transcriptomic analysis of Vigna radiata in response to chilling stress and uniconazole application. BMC Genomics 23, 205 (2022). https://doi.org/10.1186/s12864-022-08443-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12864-022-08443-6