
Abstract
Progress into the understanding of immunopathology in rheumatoid arthritis is reviewed in the present article with regard to pro-inflammatory cytokine production, cell activation and recruitment, and osteoclastogenesis. Studies highlight the potential importance of T helper 17 cells and regulatory T cells in driving and suppressing inflammation in rheumatoid arthritis, respectively, and highlight other potential T-cell therapeutic targets. The genetic associations of the HLA shared epitope alleles with antibodies to citrullinated peptides in rheumatoid arthritis patients indicate that T cells are providing help to B cells to produce autoantibodies, and there is increasing evidence that these autoantibodies are pathogenic in rheumatoid arthritis.
Similar content being viewed by others

Introduction
Rheumatoid arthritis (RA) is an autoimmune disease characterised by chronic inflammation of the joint. Although the precise pathogenesis of RA remains unclear, T cells, B cells, macrophages, neutrophils and synovial fibroblasts are central to the mechanisms of joint inflammation and disease progression. The genetic association of HLA-DR1 and HLA-DR4 with RA suggests that the disease is at least partially driven by T cells. The role of T cells has not, however, been conclusively demonstrated in the pathogenesis of RA – although the success of abatacept (CTLA-4Ig) in clinical trials [1] implies that rheumatoid T cells are important in driving the inflammatory process, and thus T cells could be targeted in clinical therapy.
The role of B cells in RA pathology has been highlighted by the clinical improvements in RA patients receiving B-cell-depleting therapies such as rituximab, an anti-CD20 antibody [2], and the increased interest in the role of autoantibodies in RA. In addition to producing antibodies, proinflammatory cytokines and chemokines, B cells efficiently act as antigen-presenting cells themselves and thus influence T-cell activation and expansion [3, 4].
In the present review we look at recent developments in the immunobiology of RA, with focus on the role of T cells and B cells, the products they produce, including cytokines and autoantibodies, and the genetic factors potentially involved in their regulation and function.
T cells
In contrast to the clearly defined role of macrophage-derived cytokines such as TNFα in the pathogenesis of RA, the relevance and contribution of the T cells is not clear and has been challenged [5]. In particular, the expectation that the increased T cells in the synovium are a result of clonal expansion to a given antigen has not been established. An HLA-restricted T-cell response to antigen is suggested, since over 80% of Caucasian RA sufferers have a shared epitope (SE) conserved across the HLA-DR1 and HLA-DR4 haplotypes (0101, 0401, 0404 and 1402) [6]. No overall consensus has been reached, however, on the potential auto-antigens involved. T-cell responses to collagen type II, heat shock proteins and microbial antigens have been reported in a small proportion of RA patients (reviewed in [7]), and more recently autoantibodies to deiminated 'citrullinated' peptides have been described, suggesting that they may be important autoantigens in this disease. This aside, the concordance for disease in identical twins is still less than 15%, suggesting other factors are of major importance.
Rheumatoid T cells have an unusual phenotype. While these cells maintain a highly activated phenotype indicated by high expression of CD69, transferrin receptor and HLA-DR, they are nonetheless hyporesponsive to antigenic stimulation [8–10]. Brennan and colleagues demonstrated that the spontaneous TNFα production in RA synovium was largely T-cell dependent [11], suggesting that regulation of T-cell function might be important to control the disease. It is thus not surprising that treatment with a nondepleting anti-CD4 antibody (keliximab) has some clinical efficacy in RA patients [12–14]. Owing to unacceptable side effects, however, anti-CD4 clinical trials were not pursued [15]. Clinical trials with abatacept, on the other hand, look more promising [1, 16, 17]. Abatacept inhibits activation of T cells by blocking the interaction between CD28 on T cells and B7 on antigen-presenting cells. In recent phase III clinical trials, abatacept showed a similar disease-modifying efficacy as infliximab treatment, the most successful treatment so far, in RA patients with an inadequate response to methotrexate [18]. Furthermore, abatacept has less adverse effect than infliximab, suggesting it is biologically safer and a more tolerised treatment [18, 19].
In addition to blocking the interaction between T cells and antigen-presenting cells, there are several other targeting possibilities for T-cell-based intervention including prevention of T-cell infiltration, inhibition of T effector cell activation and induction of regulatory T cells.
Cellular trafficking and cross-talk
An extensive array of cytokines, chemokines and adhesion molecules has been detected in the synovium of patients with RA and considered of importance in the migration of cells to the synovium (reviewed in [20]). A recent study by Kop and colleagues show that neutralisation of CD97, a member of the epidermal growth factor seven-span transmembrane family of TM7 adhesion receptors, increases resistance to collagen-induced arthritis (CIA) in mice, indicating that interaction between CD97 and its ligands may be involved in cell migration in arthritis [21]. CD97 is expressed by inflammatory cells, mainly leukocytes, in RA synovium [22]; the ligands for CD97 (CD55, chondroitin sulphate B, and α5β1) are also expressed in this tissue [22, 23].
Although antigen-dependent T-cell responses may be important in initiating the inflammatory response during arthritis, there is evidence that antigen-independent responses also play a role in RA. The RA synovial T cells can activate human monocytes/macrophages in a contact-dependent manner to induce the expression of inflammatory cytokines, including TNFα [24, 25]. A recent study further demonstrated that RA synovial T cells induce monocyte CC chemokine production (monocyte chemoattractant protein 1, macrophage inflammatory protein 1 alpha, macrophage inflammatory protein 1 beta and RANTES) and CXC chemokine production (IL-8, growth-related gene product alpha and IP-10) in a contact-dependent manner. This effector function was also shared by T cells activated with a cytokine cocktail (IL-2, IL-6 and TNFα) [26]. Furthermore, Tran and colleagues reported that T cells activated by IL-2, IL-6 and TNFα also induce fibroblast-like synoviocytes to produce inflammatory cytokines such as IL-6 and IL-8 in a cell contact-dependent manner [27]. These studies provide further evidence that T cells can be important drivers of chronic inflammation through antigen-independent mechanisms.
Cross-talk between natural killer cells and monocytes also results in the sustained stimulation of TNFα production. Natural killer cells activated by IL-15 activate monocytic cells to synthesise TNFα in a contact-dependent manner; in turn, monocytic cells induce CD69 expression and IFNγ production in natural killer cells, an effect mediated by β integrins and membrane-bound IL-15. IFNγ further increased production of membrane-bound IL-15 in monocytic cells, and neutralising membrane-bound IL-15 and β2 integrins inhibited TNFα production – suggesting that membrane-bound IL-15 and integrins are important in the cross-talk between β2 natural killer cells and monocytes [28]. The pathogenic role of IL-15 in RA has been confirmed in a phase I/II trial with anti-IL-15 therapy in RA patients [29].
T helper 17 cells
There is increasing evidence that IL-17 plays a role in the immunopathology of RA. This proinflammatory cytokine is produced by T helper (Th) 17 cells, which represent a recently discovered CD4+ effector T-cell linage distinct from Th1 cells, Th2 cells and regulatory T (Treg) cells. IL-17 has pleiotrophic effects on many cell types including macrophages, fibroblasts, epithelial cells, endothelial cells and mesenchymal cells, where it induces upregulation of nuclear factor kappa B and HLA class I as well as neutrophil chemo-kines and cytokines such as TNFα, IL-1β, IL-6 and granulocyte–macrophage colony-stimulating factor [30–33]. Important in RA pathogenesis are the effects of IL-17 in driving osteoclastogenesis leading to bone resorption. All of these effects together lead to joint destructions and chronic inflammation [33, 34]. Human RA cells expressing high levels of IL-17 are present in the synovium and circulation [34–40], and IL-17 mRNA levels in synovial membranes are predictive of joint damage progression in RA [41].
Recent work by Fasth and colleagues suggests that CD4+CD28nullcells, if activated within the synovial membrane, may potentially act as a negative regulator for the differentiation of Th17 cells in RA. An increased frequency of CD4+ T cells lacking expression of CD28 has been reported in peripheral blood of RA patients (reviewed in [42]). Fasth and colleagues more recently reported that CD4+CD28null T cells were infrequent in synovial membrane and synovial fluid, despite significant frequencies in the circulation of RA patients [43]. It is interesting to note that CD4+CD28null cells in synovial fluid were able to produce high levels of IFNγ upon antigenic stimulation. This T-cell-derived cytokine is rarely found in synovial membranes [44], and has been reported to block the differentiation of Th17 cells [45]. Nevertheless, the ability of CD4+CD28null cells to regulate the differentiation of Th17 within rheumatoid synovium remains to be demonstrated.
A role for IL-17 in experimental arthritis has been demonstrated. CIA is suppressed in IL-17-deficient mice, and administration of neutralising anti-IL17 antibodies significantly reduces the severity of CIA (reviewed in [46]). Autoimmune arthritis in SKG mice also appears to be highly dependent on the CD4+ T cells secreting IL-17 [47]. The SKG mouse strain spontaneously develops T-cell-mediated autoimmune arthritis, which clinically and immunologically resembles RA due to a mutation of the gene encoding ZAP-70, a key signal transduction molecule in T cells [48]. SKG mice develop thymus-produced self-reactive T cells that are constantly activated in the periphery, and that proliferate and differentiate to Th17 cells. In vivo development and expansion of Th17 cells, and consequently arthritis, were dependent on IL-6 produced by either T cells or non-T cells. IL-17 or IL-6 deficiency completely inhibited arthritis, whereas IFNγ deficiency exacerbated the disease [47]. A genetic polymorphism might therefore contribute to thymic generation of potentially arthritogenic self-reactive T cells, which form a cytokine milieu that facilitates differentiation into self-reactive Th17 cells.
Recent data, however, suggest that IL-17 production is regulated differently and has a somewhat different effect in humans compared with mice. While development of mouse Th17 cells require transforming growth factor beta (TGFβ) plus IL-6, human Th17 cell development appears to be independent of these cytokines but requires IL-23 and IL-1β [49–53].
T cells and osteoclastogenesis
T cells are important contributors to the pathogenesis of bone erosion in RA through induction of osteoclastogenesis. Miranda-Carus and colleagues recently showed that autologous T-cell monocyte cocultures derived from peripheral blood of patients with early RA, but not from healthy control individuals, resulted in osteoclast differentiation dependent on RANKL – which is expressed by activated T cells and RA synovial fibroblasts [54, 55] – and augmented by IL-15, IL-17, TNFα and IL-1β [56]. Other studies have identified Th17 cells as the exclusive osteoclastogenic T-cell subset among the CD4+ T-cell lineages [34, 57].
Yago and colleagues further reported that recombinant human IL-23 was able to induce osteoclastogenesis in macrophage-colony-stimulating factor-differentiated human peripheral blood mononuclear cells, and the process was independent of RANKL but dependent on osteoprogestin, IL-17 and TNFα. Furthermore, anti IL-23 treatment significantly improved inflammation and bone erosion in a CIA model in rat [58]. This further demonstrated a direct involvement of T cells in pathogenesis of RA.
T-cell activation and apoptosis
Regulation of T-cell apoptosis is critical for lymphocyte homeostasis and immune function. Inhibition of T-cell apoptosis in the synovium of patients with established RA was first described in 1995 [59]. Raza and colleagues recently showed that inhibition of synovial fluid leukocyte apoptosis in the earliest clinically apparent phase of RA distinguishes this from other early arthritides [60]. Patients with early RA had significantly lower levels of neutrophil apoptosis than patients who developed non-RA persistent arthritis and those with resolving disease course. Similarly, lymphocyte apoptosis was absent in patients with early RA whereas it was seen in patients with other early arthritides. The mechanism for this inhibition of apoptosis may relate to the high levels of antiapoptotic cytokines (IL-2, IL-4, IL-15, granulocyte–macrophage colony-stimulating factor, granulocyte colony-stimulating factor) found in the early rheumatoid joint [60].
Apoptosis proceeds through two major pathways: the intrinsic pathway is triggered by cellular stress, specifically mitochondrial stress caused by factors such as DNA damage and heat shock; and the extrinsic pathway is triggered by molecules released by other cells binding to transmembrane death receptors on the target cell to induce apoptosis (reviewed in [61]). A study in the K/BxN serum transfer model of inflammatory arthritis indicates that the proapoptotic protein Bid, an intermediary for the extrinsic and intrinsic apoptotic pathways, is important in the development of inflammatory arthritis. Mice lacking Bid display increased arthritis associated with more inflammation, pannus formation, bone destruction and infiltrating leukocytes. Furthermore there are fewer apoptotic cells in the joints of Bid-/-compared with wildtype mice, suggesting that the failure to resolve arthritis in Bid-/- mice may be due to an inability to delete autoreactive cells in the joint [62].
Most recently, the transcriptional activity in synoviocytes was investigated with a focus on the transcription factor Forkhead box class O isoforms, which are targets of the PI3 kinase/PKB signalling pathways and play emerging roles in the regulation of inflammatory responses. The Forkhead box class O isoforms are expressed in RA synovial tissue, and a strong negative correlation between inactivation (phosphorylation) of Forkhead box class O 4 in RA synovial tissue and increased serum C-reactive protein levels and raised erythrocyte sedimentation rate in RA patients has been demonstrated [63, 64]. The Forkhead box class O isoforms may thus be involved in regulating homeostasis and inflammation in autoimmune diseases.
Regulatory T cells
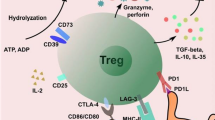
Treg cells inhibit proliferation and cytokine production of conventional T cells, including self-reactive T cells, thereby controlling inflammatory responses and contributing to the maintenance of self-tolerance (reviewed in [65]). Although the frequency of CD4+CD25+ Treg cells is higher in the synovium fluid than in peripheral blood of RA patients, there is still persistent inflammation in the joint [66–70], suggesting that the Treg cells are ineffective in controlling inflammatory responses. There is increasing evidence that the suppressive function of these Treg cells is defective. CD4+CD25high Treg cells isolated from patients with active RA express reduced levels of transcription factor forkhead box P3 (FoxP3) – which plays a major role in the function of Treg cells [71, 72] – that poorly suppress cytokine secretion from T cells and monocytes, and that do not convey a suppressive phenotype to effector CD4+CD25- T cells [73, 74]. A recent study by Valencia and colleagues suggests that TNFα, which is produced in RA synovium, inhibits the suppressive activity CD4+CD25+ Treg cells via signalling through TNF receptor II [73]. Interestingly, treatment with anti-TNF antibody (infliximab) increases FoxP3 expression in CD4+CD25high Treg cells and restores their suppressive function [73, 74]. Eliminating TNFα by antibody therapy might therefore be beneficial not only by directly suppressing proinflammtory processes but also by restoring the suppressive function of Treg cells.
Nadkarni and colleagues' work further suggests that anti-TNF therapy in RA patients generates a newly differentiated population of Treg cells, which compensates for the defective natural Treg cells [75]. The authors showed that infliximab treatment induced differentiation of a Treg cell population expressing FoxP3 and low levels of CD62L through conversion of CD4+CD25- T cells. The natural CD62L+ Treg cells remained defective in infliximab-treated patients, whereas the infliximab-induced CD62L- Treg cells mediated suppression via TGFβ and IL-10 [75].
Using experimental animal models, it has been demonstrated that depletion of CD25+ T cells before or after the induction of arthritis leads to exacerbation of arthritis with increased cellular and humoral responses, and that transfer of CD4+CD25+ Treg cells at the time of induction of arthritis decreases the severity of disease. Transfer of Treg cells, however, appears unable to cure established chronic arthritis in animal models, suggesting that therapies increasing the number of Treg cells may not be sufficient to suppress ongoing inflammation in RA (reviewed in [76]). A novel immunoregulatory T-cell population was recently discovered in mice. Charbonnier and colleagues demonstrated that vaccination with immature dendritic cells suppresses CIA in mice and induces tolerance by expansion of an immuno-regulatory TCRβ+CD49b+ T-cell population [77].
IL-6 and TGFβ together induce differentiation of pathogenic Th17 cells from naïve T cells in mice [49]. TGFβ is also a critical differentiation factor for generation of Treg cells [78], whereas IL-6, which is expressed in the RA synovium, was shown to inhibit TGFβ-induced generation of FoxP3+ Treg cells [49]. The authors suggest not only that there is a functional antagonism between Th17 and Treg cells, but that these cells arise in a mutually exclusive fashion depending on whether they are activated in the presence of TGFβ or TGFβ plus IL-6.
B cells
B-cell depletion therapy with rituximab, an anti-CD20 monoclonal antibody, provides evidence that the proinflammatory response in RA is dependent on the presence of B cells. CD20 is a B-cell surface antigen expressed only on pre-B cells and mature B cells that is lost before differentiation of B cells into plasma cells. A single course of two infusions of rituximab, alone or in combination with either cyclophospha-mide or continued methotrexate, provided significant improvement in disease symptoms in RA patients [2]. Depletion of B cells may inhibit many different immunological responses as B cells are able to internalise, to process and to present antigens via MHC class II molecules to T cells, leading to T-cell activation and subsequent macrophage activation and further TNFα production.
In some RA patients, synovial B cells undergo differentiation and proliferation within extrafollicular germinal centres consisting of T-cell and B-cell aggregates [79]. A study using human synovium–SCID mouse chimeras showed that B cells are important to the formation of these germinal centres and follicular CD4+ T cells [4]. Moreover, activated B cells can produce proinflammatory cytokines and chemokines, and experiments in animal models of arthritis have demonstrated that activation of B cells via Toll-like receptors play a significant role in the development of arthritis (reviewed in [3]). Hence, there are many possible mechanisms and strategies of B-cell-directed therapies in autoimmune diseases. In the present review we shall focus on the role of B cells as plasma cells and on the increased interest in the role of autoantibodies in RA.
Autoantibodies
Several autoantibodies have been described in RA, but only rheumatoid factor (RF), antibodies to citrullinated antigens, and antibodies to immunoglobulin binding protein have shown sufficient sensitivity and specificity to be considered clinically useful (reviewed in [80]). RF is detectable in 70% to 80% of RA patients, but is also detectable in up to 10% of normal individuals and in other systemic diseases [80]. Antibodies to autoantigens modified by citrullination through deimination of arginine to citrulline are present in about two-thirds of all RA patients, but are rare (<2%) in healthy individuals and are relatively rare in other inflammatory conditions [81, 82].
Although antibodies against citrullinated proteins are specific and predictive markers for rheumatoid arthritis, the pathologic relevance of these antibodies remains unclear. A recent study of the mouse CIA model demonstrated that antibodies against citrullinated proteins are involved in the pathogenesis of autoimmune arthritis. Kuhn and colleagues found that antibodies against both type II collagen and cyclic citrullinated peptide (CCP) appeared early after immunisation with type II collagen, before joint swelling was observed. When mice were tolerised with a citrulline-containing peptide prior to type II collagen challenge, a significantly reduced disease severity and incidence compared with control mice was demonstrated [83].
The controversy of whether autoantibodies contribute to, or are secondary to, the pathogenesis of RA was also recently addressed in a passive transfer model with mice deficient in the low-affinity inhibitory Fc receptor FcγRIIB. Petkova and colleagues showed that plasma or serum from patients with active RA induces inflammation and histological lesions in FCγRIIB-/- mice consistent with arthritis, and it was caused by the IgG-rich fraction. In contrast, serum from normal blood donors did not induce arthritis. This suggests that humoral immunity can contribute directly to autoimmune arthritis [84]. Autoantibodies will be discussed further in the context of genetics, since studies suggest that the SE alleles of the HLA-DR gene are strongly associated with anti-CCP-positive RA but not with anti-CCP-negative RA.
Genetic risk factors
RA is a complex autoimmune disease that appears to be caused by small individual effects of many common genes rather than rare mutations of single genes, and a number of gene variations have been associated with autoimmunity and RA. The SE alleles of the HLA-DR gene comprise the major genetic risk factor for RA, whereas smoking is the major known environmental risk factor (reviewed in [85]). Other polymorphic genes thought to be involved in RA include protein tyrosine phosphatase N22 (PTPN22), CTLA4, peptidyl arginine deiminase type IV and macrophage migration inhibitory factor (reviewed in [86]). More recently, single-nucleotide polymorphisms (SNPs) within signal transducer and activator of transcription 4 (STAT4) [87, 88] and TNF receptor-associated factor 1 (TRAF1)-C5 regions [89, 90] were found to be associated with seropositive RA and other autoimmune diseases.
Protein tyrosine phosphatase N22
A polymorphism that results in a substitution of arginine with tryptophan (R620W) in the PTPN22 gene has been associated with RA in European and North American populations [91–93]. The amino acid substitution in PTPN22 (R620W) affects the gene's interaction with Src tyrosine kinases involved in regulation of T-cell receptor signalling in lymphocytes [94]. A recent study found that the PTPN22 variant R620W is associated with increased titres of IgG autoantibodies to an immunodominant conformational epitope (C1III) of type II collagen in early RA [91]. The study also found that anti-C1III titres were higher in RA patients harbouring alleles of the RA-associated HLA-DRB1 SE than in those lacking this SE. The allelic variants encoding the binding pocket for peptide presentation (SE) to T cells and a functional domain of a negative regulator of T-cell receptor signalling (PTPN22*620W), respectively, synergise in early RA to break the self-tolerance towards C1III, an evolutionary conserved cartilage determinant.
The PTPN22 1858 SNP is also associated with future development of RA and has been shown to be a better predictor of RA than the HLA-SE [95]. Moreover, Johansson and colleagues found that there was an association between PTPN22 1858 and anti-CCP antibodies and that the combination gives a specificity of 100% for diagnosing RA [95]. In a German cohort, the frequency of the PTPN22 1858 polymorphism was higher in male RA patients compared with female RA patients, indicating that this genetic contribution to pathogenesis might be more prominent in men [96]. Moreover, an association between PTPN22 and RA has been found in South Asians in the United Kingdom [97] but not in a Japanese population [98], suggesting that the PTPN22 gene is associated with RA only in specific genetic groups.
Signal transducer and activator of transcription 4
A genome-wide screen for RA-susceptible genes identified a region of 52 Mb genomic DNA on chromosome 2q that was associated with a risk of RA [99]. Fine mapping of this region in North American and Swedish populations recently revealed that four SNPs within the third intron of STAT4 were associated with risk of RA, rs7574865 being the most significant [88]. An association of rs7574865 and susceptibility of RA has subsequently also been found in a Korean population [87].
STAT4 is an intracellular molecule transducing signals triggered by IL-12, type I interferons and IL-23 (reviewed in [100]), and regulates the differentiation of Th1 and Th17 cells [101], the two lymphocyte subsets thought to be involved in the pathogenesis of many inflammatory diseases (reviewed in [102, 103]). Perhaps it is not surprising that the same rs7574865 SNP was also associated with susceptibility of lupus (systemic lupus erythematosus), suggesting a common pathway of pathogenesis of autoimmune diseases [88].
TNF receptor-associated factor 1-C5
The SNPs in the region of the TRAF1-C5 locus on chromosome 9 have been associated with susceptibility and severity of RA in Dutch, Swedish and North American populations [89, 90]. This genetic risk factor, however, was not identified in a genome-wide association study performed by the Welcome Trust Case Control Consortium [104]. Association of SNPs within the TRAF1-C5 region with disease susceptibility and severity, however, is predominant in an autoantibody-positive subset of RA patients, suggesting this genetic risk factor is confined to a specific RA phenotype [89, 90].
Shared epitope alleles
The SE alleles have been shown to increase cellular susceptibility to oxidative stress, which has been implicated in RA [105]. Ling and colleagues showed that the SE acts as an allele-specific ligand that activates nitric oxide-mediated pro-oxidative signalling in nearby cells, thereby increasing cell vulnerability to oxidative damage [106]. This activation may contribute to disease susceptibility and to severity of disease.
Recent studies suggest that SE alleles are strongly associated with anti-CCP-positive RA but not with anti-CCP-negative RA, and are indeed more strongly associated with anti-CCP than with RA itself [107, 108]. van der Helm-van Mil and colleagues further showed that the presence/absence of SE alleles correlates with the levels of anti-CCP antibodies, suggesting that the SE alleles act as classic immune response genes for the development of anti-CCP antibodies [107].
There are also data suggesting that anti-CCP and the RF status are independent severity factors for RA, with SE alleles playing a secondary role at most. RF and anti-CCP were strongly associated with radiographic severity of disease, and patients with both RF and anti-CCP expressed the most severe disease, suggesting both these factors may have important influence and pathways that lead to joint damage. The anti-CCP status was also strongly associated with the SE alleles and a clear gene dose–effect was observed. The magnitude of this effect was most striking in RF-negative patients, which supports the view that the association of SE with radiographic severity may be indirect and due to an association with anti-CCP [109].
There is increasing evidence that smoking is an environmental risk factor that, in the context of HLA-DR SE genes, may trigger RA-specific immune reactions to citrullinated proteins. A recent study by Klareskog and colleagues found that previous smoking is dose-dependently associated with occurrence of anticitrulline antibodies in RA patients and that the presence of SE genes was a risk factor only for anti-citrulline-positive RA, and not for anticitrulline-negative RA. The combination of smoking history and the presence of double copies of HLA-DR SE genes increased the risk for RA 21-fold compared with the risk among nonsmokers carrying no SE genes. Moreover, positive immunostaining for citrullinated proteins was recorded in bronchoalveolar lavage cells from smokers but not in those from nonsmokers [108]. The gene–environment interaction between smoking and SE leading to autoantibodies has subsequently been reproduced in the Leiden case–control study [110] and in the Danish case–control study [111]. Unlike these studies, a recent study from North America could not, however, confirm an interaction between smoking, SE genes and anti-CCP, indicating that environmental factors other than smoking may also be associated with citrullination and RA [112].
Conclusion
Over the past couple of years advances have been made in the understanding of the involvement of T cells in the immunopathology of autoimmune diseases, with a focus on proinflammatory cytokine production, cell recruitment and osteoclastogenesis. In vitro investigations of Th17 cells have resulted in a better understanding of the T-cell inflammatory response and/or T-cell cytokine-driven inflammatory response in RA and their role in promoting osteoclastogenesis. In vivo studies with anti-TNF treatment indicated that Treg cells may be important in controlling inflammation. These investigations have identified new mechanisms of pathogenesis of RA and have opened new possibilities for future therapeutic interventions. In addition, the genetic associations of the HLA-SE alleles with antibodies to citrullinated peptides in RA patients support a role of B cells in the pathogenesis of RA, although the precise mechanisms still remain unclear.
Abbreviations
- CCP:
-
cyclic citrullinated peptide
- CIA:
-
collagen-induced arthritis
- FoxP3:
-
forkhead box P3
- IFN:
-
interferon
- IL:
-
interleukin
- PTPN22:
-
protein tyrosine phosphatase N22
- RA:
-
rheumatoid arthritis
- RF:
-
rheumatoid factor
- SE:
-
shared epitope
- SNP:
-
single-nucleotide polymorphism
- STAT4:
-
signal transducer and activator of transcription 4
- TGFβ:
-
transforming growth factor beta
- Th cells:
-
T helper cells
- TNF:
-
tumour necrosis factor
- TRAF-1:
-
TNF receptor-associated factor 1
- Treg cells:
-
regulatory T cells.
References
Ruderman EM, Pope RM: The evolving clinical profile of abatacept (CTLA4-Ig): a novel costimulatory modulator for the treatment of rheumatoid arthritis. Arthritis Res Ther. 2005, 7 (Suppl 2): S21-S25. 10.1186/ar1688.
Edwards JC, Szczepanski L, Szechinski J, Filipowicz-Sosnowska A, Emery P, Close DR, Stevens RM, Shaw T: Efficacy of B-cell-targeted therapy with rituximab in patients with rheumatoid arthritis. N Engl J Med. 2004, 350: 2572-2581. 10.1056/NEJMoa032534.
Martinez-Gamboa L, Brezinschek HP, Burmester GR, Dorner T: Immunopathologic role of B lymphocytes in rheumatoid arthritis: rationale of B cell-directed therapy. Autoimmun Rev. 2006, 5: 437-442. 10.1016/j.autrev.2006.02.004.
Takemura S, Klimiuk PA, Braun A, Goronzy JJ, Weyand CM: T cell activation in rheumatoid synovium is B cell dependent. J Immunol. 2001, 167: 4710-4718.
Firestein GS, Zvaifler NJ: How important are T cells in chronic rheumatoid synovitis?: II. T cell-independent mechanisms from beginning to end. Arthritis Rheum. 2002, 46: 298-308. 10.1002/art.502.
Stastny P: Association of the B-cell alloantigen DRw4 with rheumatoid arthritis. N Engl J Med. 1978, 298: 869-871.
Cope AP, Sonderstrup G: Evaluating candidate autoantigens in rheumatoid arthritis. Springer Semin Immunopathol. 1998, 20: 23-39. 10.1007/BF00831997.
Malone DG, Wahl SM, Tsokos M, Cattell H, Decker JL, Wilder RL: Immune function in severe, active rheumatoid arthritis. A relationship between peripheral blood mononuclear cell proliferation to soluble antigens and synovial tissue immunohistologic characteristics. J Clin Invest. 1984, 74: 1173-1185. 10.1172/JCI111526.
Emery P, Panayi GS, Welsh KI, Cole BC: Relationship of HLA-DR4 to defective cellular immunity in rheumatoid arthritis using PPD, and mycoplasma and lectin mitogens. J Rheumatol. 1985, 12: 859-864.
Cope AP, Londei M, Chu NR, Cohen SB, Elliott MJ, Brennan FM, Maini RN, Feldmann M: Chronic exposure to tumor necrosis factor (TNF) in vitro impairs the activation of T cells through the T cell receptor/CD3 complex; reversal in vivo by anti-TNF antibodies in patients with rheumatoid arthritis. J Clin Invest. 1994, 94: 749-760. 10.1172/JCI117394.
Brennan FM, Hayes AL, Ciesielski CJ, Green P, Foxwell BM, Feldmann M: Evidence that rheumatoid arthritis synovial T cells are similar to cytokine-activated T cells: involvement of phosphatidylinositol 3-kinase and nuclear factor kappaB pathways in tumor necrosis factor alpha production in rheumatoid arthritis. Arthritis Rheum. 2002, 46: 31-41. 10.1002/1529-0131(200201)46:1<31::AID-ART10029>3.0.CO;2-5.
Schulze-Koops H, Davis LS, Haverty TP, Wacholtz MC, Lipsky PE: Reduction of Th1 cell activity in the peripheral circulation of patients with rheumatoid arthritis after treatment with a non-depleting humanized monoclonal antibody to CD4. J Rheumatol. 1998, 25: 2065-2076.
Choy EH, Connolly DJ, Rapson N, Jeal S, Brown JC, Kingsley GH, Panayi GS, Johnston JM: Pharmacokinetic, pharmacodynamic and clinical effects of a humanized IgG1 anti-CD4 monoclonal antibody in the peripheral blood and synovial fluid of rheumatoid arthritis patients. Rheumatology (Oxford). 2000, 39: 1139-1146. 10.1093/rheumatology/39.10.1139.
Mason U, Aldrich J, Breedveld F, Davis CB, Elliott M, Jackson M, Jorgensen C, Keystone E, Levy R, Tesser J, Totoritis M, Truneh A, Weisman M, Wiesenhutter C, Yocum D, Zhu J: CD4 coating, but not CD4 depletion, is a predictor of efficacy with primatized monoclonal anti-CD4 treatment of active rheumatoid arthritis. J Rheumatol. 2002, 29: 220-229.
Choy EH, Panayi GS, Emery P, Madden S, Breedveld FC, Kraan MC, Kalden JR, Rascu A, Brown JC, Rapson N, Johnston JM: Repeatcycle study of high-dose intravenous 4162W94 anti-CD4 humanized monoclonal antibody in rheumatoid arthritis. A randomized placebo-controlled trial. Rheumatology (Oxford). 2002, 41: 1142-1148. 10.1093/rheumatology/41.10.1142.
Kremer JM, Dougados M, Emery P, Durez P, Sibilia J, Shergy W, Steinfeld S, Tindall E, Becker JC, Li T, Nuamah IF, Aranda R, Moreland LW: Effects of abatacept in patients with methotrexate-resistant active rheumatoid arthritis: a randomized trial. Ann Intern Med. 2006, 144: 865-876.
Genant HK, Peterfy CG, Westhovens R, Becker JC, Aranda R, Vratsanos G, Teng J, Kremer JM: Abatacept inhibits structural damage progression in rheumatoid arthritis: results from the long-term extension of the AIM trial. Ann Rheum Dis. 2007 [Epub ahead of print].
Schiff M, Keiserman M, Codding C, Songcharoen S, Berman A, Nayiager S, Saldate C, Li T, Aranda R, Becker JC, Lin C, Cornet PL, Dougados M: Efficacy and safety of abatacept or infliximab versus placebo in ATTEST: a phase III, multicenter, randomized, double-blind, placebo-controlled study in patients with rheumatoid arthritis and an inadequate response to methotrexate. Ann Rheum Dis. 2007 [Epub ahead of print].
Kremer JM, Dougados M, Emery P, Durez P, Sibilia J, Shergy W, Steinfeld S, Tindall E, Becker JC, Li T, Nuamah IF, Aranda R, Moreland LW: Treatment of rheumatoid arthritis with the selective costimulation modulator abatacept: twelve-month results of a phase iib, double-blind, randomized, placebo-controlled trial. Arthritis Rheum. 2005, 52: 2263-2271. 10.1002/art.21201.
Sweeney SE, Firestein GS: Rheumatoid arthritis: regulation of synovial inflammation. Int J Biochem Cell Biol. 2004, 36: 372-378. 10.1016/S1357-2725(03)00259-0.
Kop EN, Adriaansen J, Smeets TJ, Vervoordeldonk MJ, van Lier RA, Hamann J, Tak PP: CD97 neutralisation increases resistance to collagen-induced arthritis in mice. Arthritis Res Ther. 2006, 8: R155-10.1186/ar2049.
Hamann J, Wishaupt JO, van Lier RA, Smeets TJ, Breedveld FC, Tak PP: Expression of the activation antigen CD97 and its ligand CD55 in rheumatoid synovial tissue. Arthritis Rheum. 1999, 42: 650-658. 10.1002/1529-0131(199904)42:4<650::AID-ANR7>3.0.CO;2-S.
Sharif M, Osborne DJ, Meadows K, Woodhouse SM, Colvin EM, Shepstone L, Dieppe PA: The relevance of chondroitin and keratan sulphate markers in normal and arthritic synovial fluid. Br J Rheumatol. 1996, 35: 951-957. 10.1093/rheumatology/35.10.951.
Sebbag M, Parry SL, Brennan FM, Feldmann M: Cytokine stimulation of T lymphocytes regulates their capacity to induce monocyte production of tumor necrosis factor-alpha, but not interleukin-10: possible relevance to pathophysiology of rheumatoid arthritis. Eur J Immunol. 1997, 27: 624-632. 10.1002/eji.1830270308.
McInnes IB, Leung BP, Sturrock RD, Field M, Liew FY: Interleukin-15 mediates T cell-dependent regulation of tumor necrosis factor-alpha production in rheumatoid arthritis. Nat Med. 1997, 3: 189-195. 10.1038/nm0297-189.
Beech JT, Andreakos E, Ciesielski CJ, Green P, Foxwell BM, Brennan FM: T-cell contact-dependent regulation of CC and CXC chemokine production in monocytes through differential involvement of NFκB: implications for rheumatoid arthritis. Arthritis Res Ther. 2006, 8: R168-10.1186/ar2077.
Tran CN, Lundy SK, White PT, Endres JL, Motyl CD, Gupta R, Wilke CM, Shelden EA, Chung KC, Urquhart AG, Fox DA: Molecular interactions between T cells and fibroblast-like synoviocytes. Role of membrane tumor necrosis factor-α on cytokine-activated T cells. Am J Pathol. 2007, 171: 1588-1598. 10.2353/ajpath.2007.070004.
Gonzalez-Alvaro I, Dominguez-Jimenez C, Ortiz AM, Nunez-Gonzalez V, Roda-Navarro P, Fernandez-Ruiz E, Sancho D, Sanchez-Madrid F: Interleukin-15 and interferon-gamma participate in the cross-talk between natural killer and monocytic cells required for tumour necrosis factor production. Arthritis Res Ther. 2006, 8: R88-10.1186/ar1955.
Baslund B, Tvede N, Danneskiold-Samsoe B, Larsson P, Panayi G, Petersen J, Petersen LJ, Beurskens FJ, Schuurman J, van de Winkel JG, Parren PW, Gracie JA, Jongbloed S, Liew FY, McInnes IB: Targeting interleukin-15 in patients with rheumatoid arthritis: a proof-of-concept study. Arthritis Rheum. 2005, 52: 2686-2692. 10.1002/art.21249.
Huang SH, Frydas S, Kempuraj D, Barbacane RC, Grilli A, Boucher W, Letourneau R, Madhappan B, Papadopoulou N, Verna N, De Lutiis MA, Iezzi T, Riccioni G, Theoharides TC, Conti P: Interleukin-17 and the interleukin-17 family member network. Allergy Asthma Proc. 2004, 25: 17-21.
Ryu S, Lee JH, Kim SI: IL-17 increased the production of vascular endothelial growth factor in rheumatoid arthritis synoviocytes. Clin Rheumatol. 2006, 25: 16-20. 10.1007/s10067-005-1081-1.
Jovanovic DV, Di Battista JA, Martel-Pelletier J, Jolicoeur FC, He Y, Zhang M, Mineau F, Pelletier JP: IL-17 stimulates the production and expression of proinflammatory cytokines, IL-β and TNF-α, by human macrophages. J Immunol. 1998, 160: 3513-3521.
Kolls JK, Linden A: Interleukin-17 family members and inflammation. Immunity. 2004, 21: 467-476. 10.1016/j.immuni.2004.08.018.
Kotake S, Udagawa N, Takahashi N, Matsuzaki K, Itoh K, Ishiyama S, Saito S, Inoue K, Kamatani N, Gillespie MT, Martin TJ, Suda T: IL-17 in synovial fluids from patients with rheumatoid arthritis is a potent stimulator of osteoclastogenesis. J Clin Invest. 1999, 103: 1345-1352. 10.1172/JCI5703.
Chabaud M, Durand JM, Buchs N, Fossiez F, Page G, Frappart L, Miossec P: Human interleukin-17: a T cell-derived proinflammatory cytokine produced by the rheumatoid synovium. Arthritis Rheum. 1999, 42: 963-970. 10.1002/1529-0131(199905)42:5<963::AID-ANR15>3.0.CO;2-E.
Hwang SY, Kim HY: Expression of IL-17 homologs and their receptors in the synovial cells of rheumatoid arthritis patients. Mol Cell. 2005, 19: 180-184.
Raza K, Falciani F, Curnow SJ, Ross EJ, Lee CY, Akbar AN, Lord JM, Gordon C, Buckley CD, Salmon M: Early rheumatoid arthritis is characterized by a distinct and transient synovial fluid cytokine profile of T cell and stromal cell origin. Arthritis Res Ther. 2005, 7: R784-R795. 10.1186/ar1733.
Katz Y, Nadiv O, Beer Y: Interleukin-17 enhances tumor necrosis factor alpha-induced synthesis of interleukins 1,6, and 8 in skin and synovial fibroblasts: a possible role as a 'fine-tuning cytokine' in inflammation processes. Arthritis Rheum. 2001, 44: 2176-2184. 10.1002/1529-0131(200109)44:9<2176::AID-ART371>3.0.CO;2-4.
Chabaud M, Fossiez F, Taupin JL, Miossec P: Enhancing effect of IL-17 on IL-1-induced IL-6 and leukemia inhibitory factor production by rheumatoid arthritis synoviocytes and its regulation by Th2 cytokines. J Immunol. 1998, 161: 409-414.
Ziolkowska M, Koc A, Luszczykiewicz G, Ksiezopolska-Pietrzak K, Klimczak E, Chwalinska-Sadowska H, Maslinski W: High levels of IL-17 in rheumatoid arthritis patients: IL-15 triggers in vitro IL-17 production via cyclosporin A-sensitive mechanism. J Immunol. 2000, 164: 2832-2838.
Kirkham BW, Lassere MN, Edmonds JP, Juhasz KM, Bird PA, Lee CS, Shnier R, Portek IJ: Synovial membrane cytokine expression is predictive of joint damage progression in rheumatoid arthritis: a two-year prospective study (the DAMAGE study cohort). Arthritis Rheum. 2006, 54: 1122-1131. 10.1002/art.21749.
Lundy SK, Sarkar S, Tesmer LA, Fox DA: Cells of the synovium in rheumatoid arthritis. T lymphocytes. Arthritis Res Ther. 2007, 9: 202-10.1186/ar2107.
Fasth AE, Snir O, Johansson AA, Nordmark B, Rahbar A, Af Klint E, Bjorkstrom NK, Ulfgren AK, van Vollenhoven RF, Malmstrom V, Trollmo C: Skewed distribution of proinflammatory CD4+CD28null T cells in rheumatoid arthritis. Arthritis Res Ther. 2007, 9: R87-10.1186/ar2286.
Smeets TJ, Dolhain R, Miltenburg AM, de Kuiper R, Breedveld FC, Tak PP: Poor expression of T cell-derived cytokines and activation and proliferation markers in early rheumatoid synovial tissue. Clin Immunol Immunopathol. 1998, 88: 84-90. 10.1006/clin.1998.4525.
Harrington LE, Hatton RD, Mangan PR, Turner H, Murphy TL, Murphy KM, Weaver CT: Interleukin 17-producing CD4+ effector T cells develop via a lineage distinct from the T helper type 1 and 2 lineages. Nat Immunol. 2005, 6: 1123-1132. 10.1038/ni1254.
Lubberts E, Koenders MI, van den Berg WB: The role of T-cell interleukin-17 in conducting destructive arthritis: lessons from animal models. Arthritis Res Ther. 2005, 7: 29-37. 10.1186/ar1478.
Hirota K, Hashimoto M, Yoshitomi H, Tanaka S, Nomura T, Yamaguchi T, Iwakura Y, Sakaguchi N, Sakaguchi S: T cell self-reactivity forms a cytokine milieu for spontaneous development of IL-17+ Th cells that cause autoimmune arthritis. J Exp Med. 2007, 204: 41-47. 10.1084/jem.20062259.
Sakaguchi N, Takahashi T, Hata H, Nomura T, Tagami T, Yamazaki S, Sakihama T, Matsutani T, Negishi I, Nakatsuru S, Sakaguchi S: Altered thymic T-cell selection due to a mutation of the ZAP-70 gene causes autoimmune arthritis in mice. Nature. 2003, 426: 454-460. 10.1038/nature02119.
Bettelli E, Carrier Y, Gao W, Korn T, Strom TB, Oukka M, Weiner HL, Kuchroo VK: Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells. Nature. 2006, 441: 235-238. 10.1038/nature04753.
Mangan PR, Harrington LE, O'Quinn DB, Helms WS, Bullard DC, Elson CO, Hatton RD, Wahl SM, Schoeb TR, Weaver CT: Transforming growth factor-beta induces development of the T(H)17 lineage. Nature. 2006, 441: 231-234. 10.1038/nature04754.
Veldhoen M, Hocking RJ, Atkins CJ, Locksley RM, Stockinger B: TGFβ in the context of an inflammatory cytokine milieu supports de novo differentiation of IL-17-producing T cells. Immunity. 2006, 24: 179-189. 10.1016/j.immuni.2006.01.001.
Wilson NJ, Boniface K, Chan JR, McKenzie BS, Blumenschein WM, Mattson JD, Basham B, Smith K, Chen T, Morel F, Lecron JC, Kastelein RA, Cua DJ, McClanahan TK, Bowman EP, de Waal Malefyt R: Development, cytokine profile and function of human interleukin 17-producing helper T cells. Nat Immunol. 2007, 8: 950-957. 10.1038/ni1497.
Chen Z, Tato CM, Muul L, Laurence A, O'Shea JJ: Distinct regulation of interleukin-17 in human T helper lymphocytes. Arthritis Rheum. 2007, 56: 2936-2946. 10.1002/art.22866.
Kotake S, Udagawa N, Hakoda M, Mogi M, Yano K, Tsuda E, Takahashi K, Furuya T, Ishiyama S, Kim KJ, Saito S, Nishikawa T, Takahashi N, Togari A, Tomatsu T, Suda T, Kamatani N: Activated human T cells directly induce osteoclastogenesis from human monocytes: possible role of T cells in bone destruction in rheumatoid arthritis patients. Arthritis Rheum. 2001, 44: 1003-1012. 10.1002/1529-0131(200105)44:5<1003::AID-ANR179>3.0.CO;2-#.
Shigeyama Y, Pap T, Kunzler P, Simmen BR, Gay RE, Gay S: Expression of osteoclast differentiation factor in rheumatoid arthritis. Arthritis Rheum. 2000, 43: 2523-2530. 10.1002/1529-0131(200011)43:11<2523::AID-ANR20>3.0.CO;2-Z.
Miranda-Carus ME, Benito-Miguel M, Balsa A, CoboIbanez T, Perez de Ayala C, Pascual-Salcedo D, Martin-Mola E: Peripheral blood T lymphocytes from patients with early rheumatoid arthritis express RANKL and interleukin-15 on the cell surface and promote osteoclastogenesis in autologous monocytes. Arthritis Rheum. 2006, 54: 1151-1164. 10.1002/art.21731.
Sato K, Suematsu A, Okamoto K, Yamaguchi A, Morishita Y, Kadono Y, Tanaka S, Kodama T, Akira S, Iwakura Y, Cua DJ, Takayanagi H: Th17 functions as an osteoclastogenic helper T cell subset that links T cell activation and bone destruction. J Exp Med. 2006, 203: 2673-2682. 10.1084/jem.20061775.
Yago T, Nanke Y, Kawamoto M, Furuya T, Kobashigawa T, Kamatani N, Kotake S: IL-23 induces human osteoclastogenesis via IL-17 in vitro, and anti-IL-23 antibody attenuates collagen-induced arthritis in rats. Arthritis Res Ther. 2007, 9: R96-10.1186/ar2297.
Firestein GS, Yeo M, Zvaifler NJ: Apoptosis in rheumatoid arthritis synovium. J Clin Invest. 1995, 96: 1631-1638. 10.1172/JCI118202.
Raza K, Scheel-Toellner D, Lee CY, Pilling D, Curnow SJ, Falciani F, Trevino V, Kumar K, Assi LK, Lord JM, Gordon C, Buckley CD, Salmon M: Synovial fluid leukocyte apoptosis is inhibited in patients with very early rheumatoid arthritis. Arthritis Res Ther. 2006, 8: R120-10.1186/ar2009.
Arnold R, Brenner D, Becker M, Frey CR, Krammer PH: How T lymphocytes switch between life and death. Eur J Immunol. 2006, 36: 1654-1658. 10.1002/eji.200636197.
Scatizzi JC, Hutcheson J, Bickel E, Haines GK, Perlman H: Pro-apoptotic Bid is required for the resolution of the effector phase of inflammatory arthritis. Arthritis Res Ther. 2007, 9: R49-10.1186/ar2204.
Kuo CC, Lin SC: Altered FOXO1 transcript levels in peripheral blood mononuclear cells of systemic lupus erythematosus and rheumatoid arthritis patients. Mol Med. 2007, 13: 561-566. 10.2119/2007-00021.Kuo.
Ludikhuize J, de Launay D, Groot D, Smeets TJ, Vinkenoog M, Sanders ME, Tas SW, Tak PP, Reedquist KA: Inhibition of fork-head box class O family member transcription factors in rheumatoid synovial tissue. Arthritis Rheum. 2007, 56: 2180-2191. 10.1002/art.22653.
Rouse BT: Regulatory T cells in health and disease. J Intern Med. 2007, 262: 78-95. 10.1111/j.1365-2796.2007.01836.x.
Liu MF, Wang CR, Fung LL, Lin LH, Tsai CN: The presence of cytokine-suppressive CD4+CD25+ T cells in the peripheral blood and synovial fluid of patients with rheumatoid arthritis. Scand J Immunol. 2005, 62: 312-317. 10.1111/j.1365-3083.2005.01656.x.
van Amelsfort JM, Jacobs KM, Bijlsma JW, Lafeber FP, Taams LS: CD4(+)CD25(+) regulatory T cells in rheumatoid arthritis: differences in the presence, phenotype, and function between peripheral blood and synovial fluid. Arthritis Rheum. 2004, 50: 2775-2785. 10.1002/art.20499.
Mottonen M, Heikkinen J, Mustonen L, Isomaki P, Luukkainen R, Lassila O: CD4+CD25+ T cells with the phenotypic and functional characteristics of regulatory T cells are enriched in the synovial fluid of patients with rheumatoid arthritis. Clin Exp Immunol. 2005, 140: 360-367. 10.1111/j.1365-2249.2005.02754.x.
Cao D, Malmstrom V, Baecher-Allan C, Hafler D, Klareskog L, Trollmo C: Isolation and functional characterization of regulatory CD25brightCD4+ T cells from the target organ of patients with rheumatoid arthritis. Eur J Immunol. 2003, 33: 215-223. 10.1002/immu.200390024.
Cao D, van Vollenhoven R, Klareskog L, Trollmo C, Malmstrom V: CD25brightCD4+ regulatory T cells are enriched in inflamed joints of patients with chronic rheumatic disease. Arthritis Res Ther. 2004, 6: R335-R346. 10.1186/ar1192.
Fontenot JD, Gavin MA, Rudensky AY: Foxp3 programs the development and function of CD4+CD25+ regulatory T cells. Nat Immunol. 2003, 4: 330-336. 10.1038/ni904.
Allan SE, Passerini L, Bacchetta R, Crellin N, Dai M, Orban PC, Ziegler SF, Roncarolo MG, Levings MK: The role of 2 FOXP3 isoforms in the generation of human CD4+ Tregs. J Clin Invest. 2005, 115: 3276-3284. 10.1172/JCI24685.
Valencia X, Stephens G, Goldbach-Mansky R, Wilson M, Shevach EM, Lipsky PE: TNF downmodulates the function of human CD4+CD25hi T-regulatory cells. Blood. 2006, 108: 253-261. 10.1182/blood-2005-11-4567.
Ehrenstein MR, Evans JG, Singh A, Moore S, Warnes G, Isenberg DA, Mauri C: Compromised function of regulatory T cells in rheumatoid arthritis and reversal by anti-TNFα therapy. J Exp Med. 2004, 200: 277-285. 10.1084/jem.20040165.
Nadkarni S, Mauri C, Ehrenstein MR: Anti-TNF-α therapy induces a distinct regulatory T cell population in patients with rheumatoid arthritis via TGF-β. J Exp Med. 2007, 204: 33-39. 10.1084/jem.20061531.
Bayry J, Siberil S, Triebel F, Tough DF, Kaveri SV: Rescuing CD4+CD25+ regulatory T-cell functions in rheumatoid arthritis by cytokine-targeted monoclonal antibody therapy. Drug Discov Today. 2007, 12: 548-552. 10.1016/j.drudis.2007.05.002.
Charbonnier LM, van Duivenvoorde LM, Apparailly F, Cantos C, Han WG, Noel D, Duperray C, Huizinga TW, Toes RE, Jorgensen C, et al: Immature dendritic cells suppress collagen-induced arthritis by in vivo expansion of CD49b+ regulatory T cells. J Immunol. 2006, 177: 3806-3813.
Chen ZM, O'Shaughnessy MJ, Gramaglia I, Panoskaltsis-Mortari A, Murphy WJ, Narula S, Roncarolo MG, Blazar BR: IL-10 and TGF-β induce alloreactive CD4+CD25- T cells to acquire regulatory cell function. Blood. 2003, 101: 5076-5083. 10.1182/blood-2002-09-2798.
Schroder AE, Greiner A, Seyfert C, Berek C: Differentiation of B cells in the nonlymphoid tissue of the synovial membrane of patients with rheumatoid arthritis. Proc Natl Acad Sci USA. 1996, 93: 221-225. 10.1073/pnas.93.1.221.
Mewar D, Wilson AG: Autoantibodies in rheumatoid arthritis: a review. Biomed Pharmacother. 2006, 60: 648-655. 10.1016/j.biopha.2006.09.002.
Schellekens GA, Visser H, de Jong BA, van den Hoogen FH, Hazes JM, Breedveld FC, van Venrooij WJ: The diagnostic properties of rheumatoid arthritis antibodies recognizing a cyclic citrullinated peptide. Arthritis Rheum. 2000, 43: 155-163. 10.1002/1529-0131(200001)43:1<155::AID-ANR20>3.0.CO;2-3.
Hoffman IE, Peene I, Cebecauer L, Isenberg D, Huizinga TW, Union A, Meheus L, De Bosschere K, Hulstaert F, Veys EM, et al: Presence of rheumatoid factor and antibodies to citrullinated peptides in systemic lupus erythematosus. Ann Rheum Dis. 2005, 64: 330-332. 10.1136/ard.2004.022111.
Kuhn KA, Kulik L, Tomooka B, Braschler KJ, Arend WP, Robinson WH, Holers VM: Antibodies against citrullinated proteins enhance tissue injury in experimental autoimmune arthritis. J Clin Invest. 2006, 116: 961-973. 10.1172/JCI25422.
Petkova SB, Konstantinov KN, Sproule TJ, Lyons BL, Awwami MA, Roopenian DC: Human antibodies induce arthritis in mice deficient in the low-affinity inhibitory IgG receptor FcγRIIB. J Exp Med. 2006, 203: 275-280. 10.1084/jem.20051951.
Klareskog L, Padyukov L, Ronnelid J, Alfredsson L: Genes, environment and immunity in the development of rheumatoid arthritis. Curr Opin Immunol. 2006, 18: 650-655. 10.1016/j.coi.2006.06.004.
Orozco G, Rueda B, Martin J: Genetic basis of rheumatoid arthritis. Biomed Pharmacother. 2006, 60: 656-662. 10.1016/j.biopha.2006.09.003.
Lee HS, Remmers EF, Le JM, Kastner DL, Bae SC, Gregersen PK: Association of STAT4 with rheumatoid arthritis in the Korean population. Mol Med. 2007, 13: 455-460.
Remmers EF, Plenge RM, Lee AT, Graham RR, Hom G, Behrens TW, de Bakker PI, Le JM, Lee HS, Batliwalla F, Li W, Masters SL, Booty MG, Carulli JP, Padyukov L, Alfredsson L, Klareskog L, Chen WV, Amos CI, Criswell LA, Seldin MF, Kastner DL, Gregersen PK: STAT4 and the risk of rheumatoid arthritis and systemic lupus erythematosus. N Engl J Med. 2007, 357: 977-986. 10.1056/NEJMoa073003.
Plenge RM, Seielstad M, Padyukov L, Lee AT, Remmers EF, Ding B, Liew A, Khalili H, Chandrasekaran A, Davies LR, Li W, Tan AK, Bonnard C, Ong RT, Thalamuthu A, Pettersson S, Liu C, Tian C, Chen WV, Carulli JP, Beckman EM, Altshuler D, Alfredsson L, Criswell LA, Amos CI, Seldin MF, Kastner DL, Klareskog L, Gregersen PK: TRAF1-C5 as a risk locus for rheumatoid arthritis – a genomewide study. N Engl J Med. 2007, 357: 1199-1209. 10.1056/NEJMoa073491.
Kurreeman FA, Padyukov L, Marques RB, Schrodi SJ, Seddighzadeh M, Stoeken-Rijsbergen G, van der Helm-van Mil AH, Allaart CF, Verduyn W, Houwing-Duistermaat J, Alfredsson L, Begovich AB, Klareskog L, Huizinga TW, Toes RE: A candidate gene approach identifies the TRAF1/C5 region as a risk factor for rheumatoid arthritis. PLoS Med. 2007, 4: e278-10.1371/journal.pmed.0040358.
Burkhardt H, Huffmeier U, Spriewald B, Bohm B, Rau R, Kallert S, Engstrom A, Holmdahl R, Reis A: Association between protein tyrosine phosphatase 22 variant R620W in conjunction with the HLA-DRB1 shared epitope and humoral autoimmunity to an immunodominant epitope of cartilage-specific type II collagen in early rheumatoid arthritis. Arthritis Rheum. 2006, 54: 82-89. 10.1002/art.21498.
Begovich AB, Carlton VE, Honigberg LA, Schrodi SJ, Chokkalingam AP, Alexander HC, Ardlie KG, Huang Q, Smith AM, Spoerke JM, Conn MT, Chang M, Chang SY, Saiki RK, Catanese JJ, Leong DU, Garcia VE, McAllister LB, Jeffery DA, Lee AT, Batliwalla F, Remmers E, Criswell LA, Seldin MF, Kastner DL, Amos CI, Sninsky JJ, Gregersen PK: A missense single-nucleotide polymorphism in a gene encoding a protein tyrosine phosphatase (PTPN22) is associated with rheumatoid arthritis. Am J Hum Genet. 2004, 75: 330-337. 10.1086/422827.
Hinks A, Barton A, John S, Bruce I, Hawkins C, Griffiths CE, Donn R, Thomson W, Silman A, Worthington J: Association between the PTPN22 gene and rheumatoid arthritis and juvenile idiopathic arthritis in a UK population: further support that PTPN22 is an autoimmunity gene. Arthritis Rheum. 2005, 52: 1694-1699. 10.1002/art.21049.
Bottini N, Vang T, Cucca F, Mustelin T: Role of PTPN22 in type 1 diabetes and other autoimmune diseases. Semin Immunol. 2006, 18: 207-213. 10.1016/j.smim.2006.03.008.
Johansson M, Arlestig L, Hallmans G, Rantapaa-Dahlqvist S: PTPN22 polymorphism and anti-cyclic citrullinated peptide antibodies in combination strongly predicts future onset of rheumatoid arthritis and has a specificity of 100% for the disease. Arthritis Res Ther. 2006, 8: R19-10.1186/ar1868.
Pierer M, Kaltenhauser S, Arnold S, Wahle M, Baerwald C, Hantzschel H, Wagner U: Association of PTPN22 1858 single-nucleotide polymorphism with rheumatoid arthritis in a German cohort: higher frequency of the risk allele in male compared to female patients. Arthritis Res Ther. 2006, 8: R75-10.1186/ar1945.
Mastana S, Gilmour A, Ghelani A, Smith H, Samanta A: Association of PTPN22 with rheumatoid arthritis among South Asians in the UK. J Rheumatol. 2007, 34: 1984-1986.
Ikari K, Momohara S, Inoue E, Tomatsu T, Hara M, Yamanaka H, Kamatani N: Haplotype analysis revealed no association between the PTPN22 gene and RA in a Japanese population. Rheumatology (Oxford). 2006, 45: 1345-1348. 10.1093/rheumatology/kel169.
Jawaheer D, Seldin MF, Amos CI, Chen WV, Shigeta R, Etzel C, Damle A, Xiao X, Chen D, Lum RF, Monteiro J, Kern M, Criswell LA, Albani S, Nelson JL, Clegg DO, Pope R, Schroeder HW, Bridges SL, Pisetsky DS, Ward R, Kastner DL, Wilder RL, Pincus T, Callahan LF, Flemming D, Wener MH, Gregersen PK: Screening the genome for rheumatoid arthritis susceptibility genes: a replication study and combined analysis of 512 multicase families. Arthritis Rheum. 2003, 48: 906-916. 10.1002/art.10989.
Frucht DM: IL-23: a cytokine that acts on memory T cells. Sci STKE. 2002, 2002: PE1-
Mathur AN, Chang HC, Zisoulis DG, Stritesky GL, Yu Q, O'Malley JT, Kapur R, Levy DE, Kansas GS, Kaplan MH: Stat3 and Stat4 direct development of IL-17-secreting Th cells. J Immunol. 2007, 178: 4901-4907.
Bettelli E, Oukka M, Kuchroo VK: T(H)-17 cells in the circle of immunity and autoimmunity. Nat Immunol. 2007, 8: 345-350. 10.1038/ni0407-345.
Kikly K, Liu L, Na S, Sedgwick JD: The IL-23/Th(17) axis: therapeutic targets for autoimmune inflammation. Curr Opin Immunol. 2006, 18: 670-675. 10.1016/j.coi.2006.09.008.
Genome-wide association study of 14,000 cases of seven common diseases and 3,000 shared controls. Nature. 2007, 447: 661-678. 10.1038/nature05911.
Murdoch C, Muthana M, Lewis CE: Hypoxia regulates macrophage functions in inflammation. J Immunol. 2005, 175: 6257-6263.
Ling S, Li Z, Borschukova O, Xiao L, Pumpens P, Holoshitz J: The rheumatoid arthritis shared epitope increases cellular susceptibility to oxidative stress by antagonizing an adenosine-mediated anti-oxidative pathway. Arthritis Res Ther. 2007, 9: R5-10.1186/ar2111.
van der Helm-van Mil AH, Verpoort KN, Breedveld FC, Huizinga TW, Toes RE, de Vries RR: The HLA-DRB1 shared epitope alleles are primarily a risk factor for anti-cyclic citrullinated peptide antibodies and are not an independent risk factor for development of rheumatoid arthritis. Arthritis Rheum. 2006, 54: 1117-1121. 10.1002/art.21739.
Klareskog L, Stolt P, Lundberg K, Kallberg H, Bengtsson C, Grunewald J, Ronnelid J, Harris HE, Ulfgren AK, Rantapaa-Dahlqvist S, Eklund A, Padyukov L, Alfredsson L: A new model for an etiology of rheumatoid arthritis: smoking may trigger HLA-DR (shared epitope)-restricted immune reactions to autoantigens modified by citrullination. Arthritis Rheum. 2006, 54: 38-46. 10.1002/art.21575.
Mewar D, Coote A, Moore DJ, Marinou I, Keyworth J, Dickson MC, Montgomery DS, Binks MH, Wilson AG: Independent associations of anti-cyclic citrullinated peptide antibodies and rheumatoid factor with radiographic severity of rheumatoid arthritis. Arthritis Res Ther. 2006, 8: R128-10.1186/ar2017.
Linn-Rasker SP, van der Helm-van Mil AH, van Gaalen FA, Kloppenburg M, de Vries RR, le Cessie S, Breedveld FC, Toes RE, Huizinga TW: Smoking is a risk factor for anti-CCP antibodies only in rheumatoid arthritis patients who carry HLA-DRB1 shared epitope alleles. Ann Rheum Dis. 2006, 65: 366-371. 10.1136/ard.2005.041079.
Pedersen M, Jacobsen S, Klarlund M, Pedersen BV, Wiik A, Wohlfahrt J, Frisch M: Environmental risk factors differ between rheumatoid arthritis with and without auto-antibodies against cyclic citrullinated peptides. Arthritis Res Ther. 2006, 8: R133-10.1186/ar2022.
Lee HS, Irigoyen P, Kern M, Lee A, Batliwalla F, Khalili H, Wolfe F, Lum RF, Massarotti E, Weisman M, Bombardier C, Karlson EW, Criswell LA, Vlietinck R, Gregersen PK: Interaction between smoking, the shared epitope, and anti-cyclic citrullinated peptide: a mixed picture in three large North American rheumatoid arthritis cohorts. Arthritis Rheum. 2007, 56: 1745-1753. 10.1002/art.22703.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Rights and permissions
About this article
Cite this article
Andersson, A.K., Li, C. & Brennan, F.M. Recent developments in the immunobiology of rheumatoid arthritis. Arthritis Res Ther 10, 204 (2008). https://doi.org/10.1186/ar2370
Published:
DOI: https://doi.org/10.1186/ar2370