
Abstract
Background
Plasmodium vivax is the predominant species of human malaria parasites present in China. Although sulphadoxine-pyrimethamine (SP) and chloroquine (CQ) have been widely used for malaria treatment in China, the resistance profiles of these drugs are not available. Analysis of dihydrofolate reductase (dhfr), dihydropteroate synthase (dhps), and multidrug resistance (mdr-1) gene mutations in P. vivax isolates is a valuable molecular approach for mapping resistance to SP and CQ. This study investigates the prevalence of pvdhfr, pvdhps, and pvmdr-1 of P. vivax clinical isolates from China and provides baseline molecular epidemiologic data on SP- and CQ-associated resistance in P. vivax.
Methods
Plasmodium vivax clinical isolates were collected from two malaria-endemic regions of China, subtropical (Xishuangbanna, Yunnan province) and temperate (Bozhou, Anhui province), from 2009 to 2012. All isolates were analysed for single nucleotide polymorphism haplotypes in pvdhfr, pvdhps, and pvmdr-1 using direct DNA sequencing.
Results
In pvdhfr, 15% of Xishuangbanna isolates carried wild-type (WT) allele, whereas the majority of isolates carried mutant genes with substitutions at five codons. Eight mutant haplotypes of pvdhfr were detected, while limited polymorphism of pvdhfr was found in Bozhou isolates. A size polymorphism was present in pvdhfr, with the three-repeat type being the most predominate in both Xishuangbanna (79%) and Bozhou (97%) isolates. In pvdhps, mutations at four codons were detected in Xishuangbanna isolates leading to six haplotypes, including WT allele, single-mutation, double-mutation, and triple-mutation alleles. All Bozhou isolates carried WT pvhdps. In pvmdr-1, isolates from Xishuangbanna carried mutations at codons Y976F and F1076L, whereas all isolates from Bozhou had only a single mutation at codon F1076L.
Conclusions
Plasmodium vivax isolates from subtropical and temperate zones of China are shown to have dramatically different frequencies and patterns of mutations in pvdhfr, pvdhps, and pvmdr-1. Whereas P. vivax populations in subtropical China are highly resistant to SP and CQ, those in the temperate zone may still be susceptible to SP and CQ. This information is useful for establishing treatment policy and provides a baseline for molecular surveillance of drug-resistant P. vivax in these areas.
Similar content being viewed by others

Background
Plasmodium vivax infects approximately 100 million people annually and endangers 40% of the world’s population [1]. Malaria has been a significant public health problem in China until recently, predominantly in two areas of China: subtropical zone (Yunnan and Hainan provinces) and temperate zone (such as Anhui, Henan, Hunan, Hubei, and Jiangsu provinces). Due to China’s national malaria control programme launched in 1978, the intensity and scale of vivax malaria outbreaks have decreased dramatically through vector control and drug treatment of febrile individuals [2]; however, malaria incidence rose steadily between 2000 and 2006 [3]. In subtropical China, Yunnan is one of the two provinces with year-round local transmission of P. vivax and Plasmodium falciparum and has suffered one of the highest malaria morbidity and mortality rates in China. In temperate zone of Central China, outbreaks of malaria have been reported in Anhui and Henan provinces. In 2006, malaria prevalence increased considerably, with the highest numbers of malaria cases reported in Anhui province [4]. Although malaria incidence is low in China, the areas and populations at risk are large. P. vivax malaria accounted for > 95% of all malaria cases reported in China in 2007 [5], having become the sole parasite species and responsible for more than 90% of re-emerged malaria cases reported in temperate zone of China in 2009 [6]. In 2010, a countrywide malaria elimination policy was launched by the Ministry of Health of P. R. China with the goal of eliminating malaria by 2015 in China—with the exception of the border region in Yunnan province, and to completely eliminate malaria from China by 2020. Therefore, there is an urgent need to monitor vivax malaria transmission and drug resistance in subtropical and temperate zones of China.
Resistance to common anti-malarial drugs, such as chloroquine (CQ), has been reported for P. vivax around the world, including Indonesia [7], Myanmar [8], India [9], Vietnam [10], Turkey [11], and Ethiopia [12]. CQ and primaquine have been used as first-line therapies for radical cure of vivax malaria in China for the past 60 years, and clinical failures or reduced efficacy of CQ-primaquine drug combinations for treating vivax malaria had been reported at the Yunnan-Myanmar border [13], in Yunnan [14] and Anhui [15] provinces. Although sulphadoxine-pyrimethamine (SP) is rarely used to treat patients with P. vivax malaria, pyrimethamine, a component of the two combination regimens [Maloprim® (plus dapsone) and Fansidar® (plus sulphadoxine)], has been widely used for malaria prophylaxis in China between the mid-1960s and early 1990s [16]. This may place P. vivax under the selection of SP drug stress in China, especially in the subtropical zone where P. vivax and P. falciparum mixed-species infections were common. P. vivax malaria re-emerged in many counties of China in recent years, possibly caused by social and environmental changes and intrinsic differences in parasites with long or short relapse patterns [17–20]. Even changes in mosquito strains or species could favour a subpopulation of vivax parasites and lead to changes in the parasite population structure [21], and the emergence and spread of drug-resistant malaria should, of course, be held accountable. Studies in other P. vivax-endemic regions such as Vietnam highlight the geographic heterogeneity and temporal dynamics of drug resistance in P. vivax[10], raising further concerns about the presence of clinical resistance to SP and CQ in these areas of China and highlighting the importance of continued surveillance of CQ drug efficacy.
Molecular genetic markers of resistance are useful for monitoring the emergence and spread of anti-malarial drug resistance; a better understanding of the mechanisms of drug action and resistance are essential for fulfilling the promise of eradicating malaria [22]. Currently, several genetic markers, including dihydrofolate reductase (dhfr), dihydropteroate synthase (dhps), and multidrug resistance (mdr-1), have been used to study the prevalence and spread of SP- and CQ-resistant P. vivax[23–26]; however, molecular epidemiologic information on P. vivax parasite resistance to CQ and primaquine in China is limited. This study investigated the frequencies and patterns of mutations in pvdhfr, pvdhps, and pvmdr-1 linked to SP and CQ resistance in P. vivax isolates from Yunnan and Anhui provinces of China, and the results provide important information for molecular surveillance of drug-resistant P. vivax in these areas.
Methods
Study areas
The study was conducted in areas in subtropical (Xishuangbanna prefecture, Yunnan province) and temperate (Bozhou city, Anhui province) zones of China (Figure 1). Xishuangbanna prefecture had a population of around 113,000 in 2010. It is located in southern Yunnan (the south-west area of China), bordering Myanmar in the west and Laos and Vietnam in the south, in an area between 21°08’ and 22°36’ latitude and 99°56’ and 101°50’ longitude. Its climate is characterized by a cool, dry winter, and a warm, wet summer. It has a distinct seasonal climate characterized by rainy season from May to October and dry season from November to May, an average temperature of 18–22°C and average rainfall of 1100–2400 mm per year.

Map of Yunnan in subtropical zone and Anhui province in temperate zone of China. Shown are the locations of Xishuangbanna prefecture and Bozhou city where P. vivax isolates were collected. Scale bar = 400 kilometers.
Xishuangbanna was a highly endemic area for both P. falciparum and P. vivax malaria. Five confirmed Anopheles species are widely distributed throughout Yunnan province, with the predominant malaria vectors being Anopheles minimus and Anopheles dirus[27]. Transmission in the study area is seasonal, and malaria incidence characterized by peaks of infection occurring from June to July and again in October to November [18]. Bozhou city’s population was around 485,000 in 2010. It is located in the north-west of Anhui (temperate zone of China) bordering six provinces: Jiangsu in the south-east, Shandong in the north, Hubei in the west, Henan in north-west, Jiangxi in the south, and Zhejiang in the north-east, located in an area between 32°51’ and 35°05’ latitude and 115°53’ and 116°49’ longitude. The area has a distinct seasonal climate characterized by humid summer from June to September and dry winter from October to December, with an average temperature of 14–17°C and average rainfall of 500–750 mm per year [16]. Bozhou city was an endemic area for P. vivax malaria, and the main malaria vector was Anopheles sinensis and the secondary vector Anopheles anthropophagus.
Blood sample collection
Blood samples were collected from patients with symptomatic P. vivax malaria admitted to local hospitals for anti-malarial drug treatment. Inclusion criterion was patients infected with P. vivax but no other human malaria species as confirmed by peripheral smear examination. Preparation and staining of blood slides were done following the World Health Organization procedures for basic malaria microscopy. Thick and thin blood smears were checked after staining with Giemsa. A total of 114 P. vivax clinical isolates (blood samples) were collected from patients with symptomatic P. vivax malaria admitted to local hospitals for anti-malarial drug treatment (n = 53) in Xishuangbanna, Yunnan province, from June 2011 to October 2012 and (n = 61) in Bozhou city, Anhui province, from June 2009 to October 2010 (collected earlier than those of Xishuangbanna samples). Two patients from Bozhou city had a travel history outside of Anhui; all others had no history of travel to neighbouring provinces or countries. All the Xishuangbanna isolates were collected from the border of Myanmar. After informed consent from all adults or legal guardians of children, a 1.0 mL sample of whole blood was collected from vivax malaria patients in EDTA tubes and stored at -20°C until DNA extraction. This study was reviewed and approved by the Ethics Committee of Sun Yat-sen University, China (IRB 2009A0101).
DNA extraction for pvdhfr, pvdhps, and pvmdr-1
Plasmodium vivax genomic DNA was extracted from 200 μL of each blood sample using a DNA blood kit following the manufacturer’s instructions (Takara Bio Inc., Shiga, Japan) with minor modifications. The DNA was dissolved in TE buffer (10 mM Tris–HCl, 0.1 M EDTA, pH 8.0) and stored at -20°C until use. The quality of total DNA was analysed by running 5 μL of each DNA sample on a 1.0% agarose gel stained with ethidium bromide and visualized under ultraviolet illumination.
Polymerase chain reaction (PCR) amplification for pvdhfr, pvdhps, and pvmdr-1
To amplify pvdhfr, pvdhps, and pvmdr-1, a nested PCR amplification method was used following previously reported protocols with some minor modifications [15, 16, 28]. Oligonucleotide primers and cycling conditions are listed in Table 1. All amplification reactions were carried out in a total volume of 25 μL containing 17.0 μL of dH2O, 1.0 μL of each primer (10 pM), 1.5 μL of MgCl2 (25 mM), 0.5 μL of Taq polymerase (5 U/μL), 0.5 μL of dNTP mixture (10 mM each), and 2.5 μL of 10 × PCR buffer following the manufacturer’s instructions (BioBasic Inc., Markham, ON, Canada). Primary amplification reactions were initiated with 1.0 μL of template genomic DNA prepared from the blood samples, and 1.0 μL of primary reaction amplification was used in secondary amplification reactions. The amplified nested PCR products were resolved on 1.0% agarose gel, and the sizes of the PCR products were determined using a 100-bp DNA ladder (NewProbe, Beijing, China). The nested PCR products were stored at -20°C until analysis.
Sequence analysis of pvdhfr, pvdhps, and pvmdr-1
The nested PCR products of pvdhfr, pvdhps, and pvmdr-1 from the above samples were directly sequenced in both directions, using an ABI PRISM3730 DNA sequencer from Sangon Biotech (Shanghai, China). Nucleotide and amino acid sequences of pvdhfr, pvdhps, and pvmdr-1 were aligned and compared with the following published sequences of pvdhfr (accession no. X98123), pvdhps (accession no. AY186730), and pvmdr-1 (accession no. AY618622) using Clustal W of the BioEdit 7.0 program.
Statistical analysis
Statistical significance was determined with SPSS software (version 13.0). U test was used to compare the prevalence of infection between collection sites. P < 0.05 was considered statistically significant.
Results
Prevalence and patterns of mutant pvdhfr in P. vivax isolates
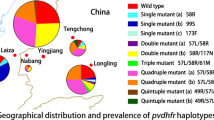
All blood samples (n = 114) were shown to be infected with P. vivax only by microscopy. The pvdhfr gene was successfully amplified from all 114 P. vivax isolates. Mutations at codons F57I/L, S58R, T61M, H99R/S, and S117N/T were detected in the 53 Xishuangbanna isolates, accounting for 60%, 62%, 62%, 19%, and 74% of the isolates examined, respectively (Table 2); no mutations at pvdhfr positions I13 or I173 were found in the Xishuangbanna isolates. Only H99S and S117N were detected in the 61 Bozhou isolates, with frequencies of 40% and 57% isolates, respectively. Mutant codon S117N was the most prevalent in both Xishuangbanna and Bozhou areas, accounting for 74% and 57% of isolates examined, respectively. Haplotype analysis of pvdhfr for all Xishuangbanna isolates revealed eight distinct allelic forms (Table 3), including the wild-type (WT) allele (I13/F57/S58/T61/H99/S117/I173), two single-mutant alleles (99S and 117N), three double-mutant alleles (99S/117N, 99R/117N, and 58R/99N), and two quadruple mutant alleles (57I/58R/61M/117T and 57L/58R/61M/117T). Of the eight allelic variants, the most prevalent allelic variants were quadruple-mutant alleles (57L/58R/61M/117T) (53%, 28/53) among Xishuangbanna isolates, followed by WT allele (15%, 8/53) and a single-mutant allele H99S (11%, 6/53). The remaining five allelic variants were only detected in 21% (11/53) of P. vivax isolates examined from Xishuangbanna. Four allelic variants [WT allele, single-mutant alleles (99S and 117N), and a double-mutant allele (99S/117N)] were detected in 61 isolates from Bozhou, with a single-mutant allele (117N) being predominant (42%, 26/61) (Table 3).
In addition to the above point mutations, variations at the central tandem repeat region between amino acid positions 88 and 105 of pvdhfr were also detected. Analysis of repeat variant in pvdhfr by sequencing showed that most (89%, 101/114) of the isolates from both areas contained three copies of the GGDNXX repeat that could be divided into type A, B, and C based on substitutions in the middle repeat (Figure 2A). The rest of the isolates (11%, 13/114) had two repeats (type D). The majority of parasites from both Xishuangbanna and Bozhou isolates (79% and 97%, respectively) had the three-copy repeat type. However, the distribution and frequency of tandem repeats in pvdhfr in Xishuangbanna isolates were different from those of Bozhou isolates (Figure 2B). Type A (60%) was the dominant allele in Xishuangbanna, whereas type C (56%) was the most common allele among the Bozhou isolates.

Alignment of central tandem repeat region between amino acid positions 88 and 105 in the pvdhfr gene and frequencies of central tandem repeat types circulating from Xishuangbanna and Bozhou. A) Sequences were constructed to the published amino acid sequences (accession no. X98123). Dots (. . . .) and dashes (---) represent identical residues and deletions, respectively. Amino acid changes resulting from nucleotide substitutions are shown in bold. B) Frequencies of four central tandem repeat types obtained from 53 P. vivax Xishuangbanna isolates (black) and 61 Bozhou isolates (grey).
Prevalence and patterns of mutant pvdhps in P. vivax isolates
All 114 isolates from Xishuangbanna and Bozhou were successfully amplified for pvdhps. Sequence analysis revealed that point mutations were detected at codons S382A, A383G, K512E, and A553G, but not at V585 in pvdhps in Xishuangbanna isolates (Table 2). Among the above four mutants, codon S382A was the most prevalent, accounting for 79%, followed by codons S382A and A553G with frequency of 32% and 28% in all Xishuangbanna isolates, respectively; the rest of the mutant codon K512E accounted for only 2% of the Xishuangbanna isolates. Haplotype analysis of pvdhps in Xishuangbanna isolates showed six distinct allelic forms, including WT allele (S382/A383/K512/A553/V585), one single-mutant allele (383G), two double-mutant alleles (S382A/A383G and 383G/553G), and two triple-mutant alleles (S382A/A383G/A553G and 383G/512E/585G) (Table 3). The most common pvdhps alleles in Xishuangbanna isolates were the double-mutant allele 382A/383G, followed by single-mutant allele 383G, double-mutant allele 383G/553G, and WT mutant, accounting for 26%, 25%, 21%, and 21%, respectively. However, no mutation was detected in any of the Bozhou P. vivax isolates (Tables 2 and 3).
Prevalence and patterns of mutant pvmdr-1 in P. vivax isolates
All 114 samples were successfully amplified for pvmdr-1, and mutations at codons Y976F and F1076L with the frequencies of 4% and 70%, respectively, were detected (Table 2); however, different mutant alleles were found in the parasite populations from Xishuangbanna and Bozhou. Mutant codon F1076L was detected from the isolates of both Xishuangbanna and Bozhou (36% and 100%, respectively); whereas mutant codon Y976F was found only in Xishuangbanna isolates (9%). Three haplotypes existed among the parasites: WT (Y976/ F1076), single-mutant allele (1076L), and double-mutant allele (976F/1076L), with WT being the predominant type (65%, 34/53) in Xishuangbanna isolates (Table 3) and the single-mutant allele (1076L) being the only genotype (100%, 61/61) in the Bozhou isolates (Table 3).
Haplotypes of combined pvdhfr, pvdhps, and pvmdr-1 allelic variants
Analysing variants of the three anti-malarial drug genetic markers (pvdhfr, pvdhps, and pvmdr-1) in combination identified a total of 28 distinct haplotypes in the 114 isolates (24.5%) from both Xishuangbanna and Bozhou. Among the haplotypes, 26 were found in Xishuangbanna isolates (22.8%), four were found in Bozhou isolates (3.5%), and two (Types 2 and 10) were found in both regions (Additional file 1). The two most prevalent haplotypes were Types 21 and 22, accounting for 11.3% and 13.2% of Xishuangbanna isolates, respectively; the remainder haplotypes were distributed at low frequency (1.9% to 7.5%), although the predominant haplotype in Bozhou isolates was Type 7 (42.6%, 26/61), followed by Types 10, 2, and 13. Disregarding the mutations at codon 99 in pvdhfr and at codon 1076 in pvmdr-1, which were considered to be not related with the anti-malarial drugs, 4.4% of the 114 isolates (including Types 16, 19, and 26) had mutations in all the three genes, pvdhfr, pvdhps, and pvmdr-1, simultaneously, whereas 31.6% of the 114 isolates (including Types 8, 14–19, and 21–28) had mutations in the SP drug-related genes (dhfr and dhps) which were found in Xishuangbanna but not Bozhou isolates.
Discussion
Emergence and spread of CQ- or SP-resistant P. vivax strains has been reported in many malarial regions around the world, especially in Southeast Asia, which is responsible for the increasing morbidity and mortality of P. vivax[29]. The use of anti-malarial drugs exhibits enormous geographic heterogeneity in subtropical and temperate zones of China. With increasing movement of human populations, drug-resistant P. vivax populations and the parasite transmission pattern in the regions are also changing constantly. The emergence and spread of drug-resistant strains of human Plasmodium species may be partly responsible for re-emergence of malaria in China. SP was widely used for malaria prophylaxis between the mid-1960s and early 1990s, and CQ-primaquine is still the first-line anti-malarial drug for treating P. vivax malaria in China [16]. Molecular markers have been validated as tools for surveillance of resistance. Analysis of point mutations in these marker genes thus serves as a valuable molecular approach for mapping drug resistance and monitoring malaria-control measures [30]. In this study, the profile of mutations in marker genes associated with CQ and antifolate drug resistance among the P. vivax parasites obtained from patients of the subtropical (Xishuangbanna in Yunnan) and temperate (Bozhou in Anhui) zones of China were determined to better understand the current and changing patterns of CQ- and SP-resistant P. vivax in different malaria-endemic areas. The P. vivax parasite populations in subtropical and temperate zones were shown to differ dramatically in pvdhfr, pvdhps, and pvmdr-1 allele frequencies, i.e., P. vivax populations in subtropical zone are mostly resistant to SP and are likely more tolerant to CQ, whereas the majority of P. vivax populations in the temperate zone are still effectively susceptible to SP and CQ.
The long history of CQ use has exposed P. vivax to this drug pressure continually in China. Clinical failures after standard CQ treatment were reported in four cases of P. vivax malaria in Yunnan province in 1996 [14]. A trend of gradual decline of in vitro sensitivity to CQ had also been documented between 2005 and 2008 in some areas of China, especially at the Yunnan-Myanmar border and in central China [31], and reduced efficacy of CQ combinations for treating vivax malaria patients from 2007 to 2008 was reported at the Yunnan-Myanmar border [13]. The relatively high frequency (9%) of pvmdr-1 Y976F mutation among the Xishuangbanna isolates is consistent with the previous reports of declining sensitivity to CQ. The Y976F mutation had been reported in P. vivax isolates from many malaria-endemic regions around the world, including Indonesia [26], Thailand [32], Myanmar [32], and Mauritania [33, and is associated with an increase in CQ IC50 values in vitro[15]. Cases of CQ-resistant P. vivax malaria have been reported in the areas of Yunnan bordering Myanmar, Laos, and Vietnam [8, 10]. The long history of CQ use, as well as frequent population movement across the borders, may contribute to the CQ-resistant P. vivax detected in Xishuangbanna. However, the Y976F mutation was not detected in any Bozhou isolates, which is consistent with the results showing that CQ is still effective in killing P. vivax parasites from the temperate zone in an in vitro schizont maturation inhibition assay [15]. The presence of Y976F in pvmdr-1 in Xishuangbanna isolates suggests a trend toward decreased CQ sensitivity.
This study detected the F1076L mutation in pvmdr-1 in 100% of Bozhou isolates, which is consistent with previous reports from Central China (100%) [26], the Republic of Korea (100%) [26], Mauritania (100%) [26], and Madagascar (100%) [34]; however, only 36% of Xishuangbanna isolates had the F1076L mutation. It has been reported that the F1076L substitution may not be linked to drug resistance but may rather be a geographic variant [34].
SP inhibits the activities of dhps and dhfr in the folate biosynthesis pathway of both P. falciparum and P. vivax parasites, resulting in a synergistic anti-malarial effect [35]. Resistance to antifolate drugs in both parasite species is found to be involved with point mutations in dhps and dhfr[23]. Although P. vivax infections are not generally treated with SP, the parasite is often exposed to the drug due to mixed infections and misdiagnosis [36]. In this study, five mutations at amino acid positions 57, 58, 61, 99, and 117 and seven mutated alleles in pvdhfr were detected in Xishuangbanna isolates, while only mutations at positions 99 and 117 of pvdhfr and three mutated alleles in pvdhfr were found in Bozhou isolates. It has been reported that double mutation S58R and S117N in pvdhfr may first arise under drug pressure and move toward development of resistance to SP [23]. Interestingly, approximately 62% of Xishuangbanna isolates was found to have double mutations at S58R/S117N, whereas no Bozhou isolates were detected with these mutations. Instead, 39% of Bozhou isolates had a single S117N mutation, indicating that the P. vivax isolate circulating in Xishuangbanna may be under stronger drug pressure than those in Bozhou. The most prevalent double mutations of S58R/S117N in Xishuangbanna isolates were similar to those in previous reports from East Timor [37], the Philippines [37], Vietnam [37], and Pakistan [38]. Clinical studies have shown that patients carrying triple and quadruple pvdhfr mutant vivax parasites were more likely associated with SP treatment failure than those with WT parasites [39, 40]. The triple and quadruple mutations in pvdhfr have been detected in Yunnan [16], Hainan [16], and Guizhou [41] provinces of China’s subtropical zone, and in some other countries including Indonesia [40], Papua New Guinea [40] and India [42]. Similarly, 61% of Xishuangbanna isolates had quadruple mutant alleles (57L/58R/61M/117T and 57I/58R/61M/117T), strongly suggesting that many Xishuangbanna P. vivax isolates are resistant to sulphadoxine drugs. The triple and quadruple mutations were not detected in isolates in Bozhou as well as in other temperate areas of China (such as Wuhe county in Anhui province) [41], Thailand [43], Vietnam [43], Korea [43], Afghanistan [44], Pakistan [45], and Iran [46], indicating that Bozhou P. vivax isolates may still be sensitive to antifolate drugs.
Among mutated codons in pvdhfr, the T61M mutation was mostly linked to the S117T mutation [23]. In the present study, two mutations at T61M/S117T with quadruple mutated alleles (57L/58R/61M/117T and 57I/58R/61M/117T) were detected in Xishuangbanna isolates. The observation is consistent with the early reports from Thailand [47], but is in contrast to other reports from the temperate zone of China (Wuhe county in Anhui province) that most T61M mutations were found as single mutations and arose independently [41]. In the current study, the three-copy repeat type was the predominate type in both Xishuangbanna (79%) and Bozhou (97%) isolates, which was similar to the findings from India [42], Afghanistan [44], Iran [46], and Wuhe county in Anhui province and Luodian county in Guizhou province of China [41]. The association between pvdhfr mutations and tandem repeat polymorphisms has been extensively investigated and used to predict the prevalence of drug-resistant malaria around the world [43]. Therefore, the observations from this study suggest increasing prevalence of antifolate-resistant parasites in the Xishuangbanna region. Additionally, no isolates were detected with the four-copy repeat type from either Xishuangbanna or Bozhou, in contrast to other reports showing that quadruple-mutant dhfr alleles were exclusively associated with the four-copy repeat type in Thailand [23], India [42], and Myanmar [48]. Whether the four-copy repeat type contributes to drug resistance requires further investigation.
It has been suggested that mutations at codons 382, 383, 512, 553, and 585 in pvdhps gene are related to reduced sensitivity to sulfadoxine [46, 49]. In the present study, four pvdhps mutations at codons S382A (32%), A383G (79%), K512E (2%), and A553G (28%) and five mutant-allelic types were detected in Xishuangbanna isolates, including 25% single-mutant allele (A383G), 47% double-mutant allele (S382A/A383G and A383G/A553G), and 8% triple-mutant allele (S382A/A383G/A553G and A383G/K512E/A553G), strongly suggesting that Xishuangbanna P. vivax isolates were highly resistant to sulphadoxine. The high prevalence of mutant pfdhps alleles is similar to a previous report in Yunnan province of China (Nu River) [16]. In contrast, all isolates from Bozhou carried the WT allele, which is similar to reports from Wuhe county in Anhui province [31, 41] and Luodian county in Guizhou province [41] of China, Mauritania [33], Iran [38], and Afghanistan [44]. The absence of mutations in pvdhps in this study indicated that P. vivax populations in Bozhou may still be effectively susceptible to sulphadoxine.
Molecular epidemiologic studies in different areas have shown dramatically different mutation rates in pvdhfr and pvdhps, which may be attributed to different drug-selection pressures or the intrinsic differences among endemic strains of P. vivax. It is important to take into account the presence of A383G mutation of pvdhps along with double pvdhfr mutations, as SP drug-treatment failure was more frequently associated with multiple mutations in pvdhfr and pvdhps[50]; when the P. falciparum parasite carries mutant alleles of dhfr and dhps, clinical effectiveness of SP is compromised [51]. In the current study, disregarding the mutations at codon 99 of pvdhfr, which is not considered to be related to the anti-malarial drugs, 31.6% of 114 isolates (including Types 8, 14 to 19, and 21 to 28) contained the mutations in pvdhfr and pvdhps and 4.4% of 114 isolates (including Types 16, 19, and 26) mutated simultaneously in the three drug-resistant genes pvdhfr, pvdhps, and pvmdr-1 were found in the Xishuangbanna isolates but not in Bozhou isolates. Although SP was not recommended to treat P. vivax malaria in China due to the intensive use of antifolate drugs for treating P. falciparum infections, P. vivax has been under SP drugs pressure through mixed infections and/or incorrect diagnosis in Yunnan. In contrast, without the introduction of sulphadoxine in Anhui—where P. vivax is the only or most predominant malaria parasite, P. vivax is not under sulphadoxine but rather pyrimethamine drug pressure. Therefore, the different usage of antifolate drug in Yunnan and Anhui may lead to the high prevalence of pvdhfr and pvdhps mutated alleles in Xishuangbanna isolates and relatively low prevalence of these mutations in Bozhou isolates. In tropical area, in addition to longer transmission period, short-term relapses are more common (1–2 months), leading to increased expose of P. vivax parasites to sub-therapeutic concentrations of SP and CQ. Regardless, the results from this study indicates that the high prevalence of multiple mutations of pvdhfr, pvdhps, and pvmdr-1 genes may further reduce the sensitivity to SP and CQ in P. vivax populations in Xishuangbanna of Yunnan, while the P. vivax population may still be susceptible to SP and CQ in Bozhou of central China due to the absence of multiple mutations in pvdhfr, pvdhps, and pvmdr-1.
Conclusions
Results from the current study indicate that the prevalence and patterns of mutant pvdhfr, pvdhps, and pvmdr-1 exhibit enormous geographic heterogeneity in subtropical (Xishuangbanna, Yunnan) and temperate (Bozhou, Anhui) zones of China. P. vivax populations in subtropical zone of China are likely resistant to SP and CQ, whereas in temperate zone of China they are still relatively susceptible to SP and CQ, which provides valuable information for monitoring drug resistance in different malaria-endemic areas of China and for assessing the appropriateness of the current national anti-malarial drug policy. The increased resolution afforded by combination of molecular genetics and geographic information systems (GIS) tools has the potential to provide insights into the epidemiology, evolution, and ecology of these parasites in the future.
Abbreviations
- CQ:
-
Chloroquine
- dhfr :
-
Dihydrofolate reductase gene
- dhps :
-
Dihydropteroate synthase gene
- mdr-1 :
-
Multidrug resistance gene
- PCR:
-
Polymerase chain reaction
- SP:
-
Sulphadoxine-pyrimethamine
- WT:
-
Wild type.
References
Bright AT, Manary MJ, Tewhey R, Arango EM, Wang T, Schork NJ, Yanow SK, Winzeler EA: A high resolution case study of a patient with recurrent Plasmodium vivax infections shows that relapses were caused by meiotic siblings. PLoS Negl Trop Dis. 2014, 8: e2882-10.1371/journal.pntd.0002882.
Tang LH, Qian HL, Xu SH: Malaria and its control in the People’s Republic of China. Southeast Asian J Trop Med Public Health. 1991, 22: 467-476.
Zhu G, Xia H, Zhou H, Li J, Lu F, Liu Y, Cao J, Gao Q, Sattabongkot J: Susceptibility of Anopheles sinensis to Plasmodium vivax in malarial outbreak areas of central China. Parasit Vectors. 2013, 6: 176-10.1186/1756-3305-6-176.
Zhou S, Huang F, Tang L, Zheng X, Shen Y, Su Y, Huang G: Study on the spatial distribution of malaria in Yellow River and Huai River areas based on the “Kriging” method (in Chinese). Journal of Pathogen Biology. 2007, 3: 204-207.
Zhou S, Wang Y, Fang W, Tang LH: Malaria situation in the People’s Republic of China in 2007. Chin Parasitol Parasit Dis. 2008, 29: 401-403.
Zhou S, Wang Y, Xia ZG: Malaria situation in the People’s Republic of China in 2009. Chin Parasitol Parasit Dis. 2011, 29: 1-3.
Sumawinata IW, Leksana B, Sutamihardja A, Subianto B, Fryauff DJ, Baird JK: Very high risk of therapeutic failure with chloroquine for uncomplicated Plasmodium falciparum and P. vivax malaria in Indonesian Papua. Am J Trop Med Hyg. 2003, 68: 416-420.
Guthmann JP, Pittet A, Lesage A, Imwong M, Lindegardh N, Min Lwin M, Zaw T, Annerberg A, de Radiguès X, Nosten F: Plasmodium vivax resistance to chloroquine in Dawei, southern Myanmar. Trop Med Int Health. 2008, 13: 91-98. 10.1111/j.1365-3156.2007.01978.x.
Garg M, Gopinathan N, Bodhe P, Kshirsagar NA: Vivax malaria resistant to chloroquine: case reports from Bombay. Trans R Soc Trop Med Hyg. 1995, 89: 656-657. 10.1016/0035-9203(95)90432-8.
Phan GT, de Vries PJ, Tran BQ, Le HQ, Nguyen NV, Nguyen TV, Heisterkamp SH, Kager PA: Artemisinin or chloroquine for blood stage Plasmodium vivax malaria in Vietnam. Trop Med Int Health. 2002, 7: 858-864. 10.1046/j.1365-3156.2002.00948.x.
Kurcer MA, Simsek Z, Kurcer Z: The decreasing efficacy of chloroquine in the treatment of Plasmodium vivax malaria, in Sanliurfa, south-eastern Turkey. Ann Trop Med Parasitol. 2006, 100: 109-113. 10.1179/136485906X86284.
Teka H, Petros B, Yamuah L, Tesfaye G, Elhassan I, Muchohi S, Kokwaro G, Aseffa A, Engers H: Chloroquine-resistant Plasmodium vivax malaria in Debre Zeit, Ethiopia. Malar J. 2008, 7: 220-10.1186/1475-2875-7-220.
Liang GL, Sun XD, Wang J, Zhang ZX: Sensitivity of Plasmodium vivax to chloroquine in Laza City, Myanmar. Chin J Parasitol Parasitic Dis. 2009, 27: 175-176.
Yang XM, Yang MQ, Huang JW: Clinical research of the sensitivity of Plasmodium vivax to chloroquine. Chinese J Parasitol Parasit Dis. 1996, 9: 226-227.
Lu F, Lim CS, Nam DH, Kim K, Lin K, Kim TS, Lee HW, Chen JH, Wang Y, Sattabongkot J, Han ET: Genetic polymorphism in pvmdr-1 and pvcrt-o genes in relation to in vitro drug susceptibility of Plasmodium vivax isolates from malaria-endemic countries. Acta Trop. 2011, 117: 69-75. 10.1016/j.actatropica.2010.08.011.
Ding S, Ye R, Zhang D, Sun X, Zhou H, McCutchan TF, Pan W: Anti-folate combination therapies and their effect on the development of drug resistance in Plasmodium vivax. Sci Rep. 2013, 3: 1008-
Clements AC, Barnett AC, Cheng ZW, Snow RW, Zhou HN: Space-time variation of malaria incident in Yunnan province, China. Malar J. 2009, 8: 180-10.1186/1475-2875-8-180.
Hui FM, Xu B, Chen ZW, Cheng X, Liang L, Huang HB, Fang LQ, Yang H, Zhou HN, Yang HL, Zhou XN, Cao WC, Gong P: Spatio-temporal distribution of malaria in Yunnan Province, China. Am J Trop Med Hyg. 2009, 81: 503-509.
Gao HW, Wang LP, Liang S, Liu YX, Tong SL, Wang JJ, Li YP, Wang XF, Yang H, Ma JQ, Fang LQ, Cao WC: Change in rainfall drives malaria re-emergence in Anhui province, China. PLoS One. 2012, 7: e43686-10.1371/journal.pone.0043686.
Zhou S, Zhang S, Wang J, Zheng X, Huang F, Li W, Xu X, Zhang HW: Spatial correlation between malaria cases and water-bodies in Anopheles sinensis dominated areas of Huang-Huai plain, China. Parasit Vectors. 2012, 5: 106-10.1186/1756-3305-5-106.
Huang B, Huang S, Su XZ, Guo H, Xu Y, Xu F, Hu X, Yang Y, Wang S, Lu F: Genetic diversity of Plasmodium vivax population in Anhui province of China. Malar J. 2014, 13: 13-10.1186/1475-2875-13-13.
Yuan J, Cheng KC, Johnson RL, Huang R, Pattaradilokrat S, Liu A, Guha R, Fidock DA, Inglese J, Wellems TE, Austin CP, Su XZ: Chemical genomic profiling for antimalarial therapies, response signatures, and molecular targets. Science. 2011, 333: 724-729. 10.1126/science.1205216.
Imwong M, Pukrittayakamee S, Rénia L, Letourneur F, Charlieu JP, Leartsakulpanich U, Looareesuwan S, White NJ, Snounou G: Novel point mutations in the dihydrofolate reductase gene of Plasmodium vivax: evidence for sequential selection by drug pressure. Antimicrob Agents Chemother. 2003, 47: 1514-1521. 10.1128/AAC.47.5.1514-1521.2003.
Gregson A, Plowe CV: Mechanisms of resistance of malaria parasites to antifolates. Pharmacol Rev. 2005, 57: 117-145. 10.1124/pr.57.1.4.
Marfurt J, de Monbrison F, Brega S, Barlollat L, Müller I, Sie A, Goroti M, Reeder JC, Beck HP, Picot S, Genton B: Molecular markers of in vivo Plasmodium vivax resistance to amodiaquine plus sulfadoxine-primethamine: mutations in pvdhfr and pvmdr-1. J Infect Dis. 2008, 198: 409-417. 10.1086/589882.
Brega S, Meslin B, de Monbrison F, Severini C, Gradoni L, Udomsangpetch R, Sutanto I, Peyron F, Picot S: Identification of the Plasmodium vivax mdr-like gene (pvmdr1) and analysis of single-nucleotide polymorphisms among isolates from different areas of endemicity. J Infect Dis. 2005, 191: 272-277. 10.1086/426830.
Cui L, Yan G, Sattabongkot J, Cao Y, Chen B, Chen X, Fan Q, Fang Q, Jongwutiwes S, Parker D, Sirichaisinthop J, Kyaw MP, Su XZ, Yang H, Yang Z, Wang B, Xu J, Zheng B, Zhong D, Zhou G: Malaria in the Greater Mekong Subregion: heterogeneity and complexity. Acta Trop. 2012, 121: 227-239. 10.1016/j.actatropica.2011.02.016.
Barnadas C, Kent D, Timinao L, Iga J, Gray LR, Siba P, Mueller I, Thomas PJ, Zimmerman PA: A new high-throughput method for simultaneous detection of drug resistance associated mutations in Plasmodium vivax dhfr, dhps and mdr-1 genes. Malar J. 2011, 10: 282-10.1186/1475-2875-10-282.
Tjitra E, Anstey NM, Sugiarto P, Warikar N, Kenangalem E, Karyana M, Lampah DA, Price RN: Multidrug-resistant Plasmodium vivax associated with severe and fatal malaria: a prospective study in Papua, Indonesia. PLoS Med. 2008, 5: e128-10.1371/journal.pmed.0050128.
Garg S, Saxena V, Lumb V, Pakalapati D, Boopathi PA, Subudhi AK, Chowdhury S, Kochar SK, Kochar DK, Sharma YD, Das A: Novel mutations in the antifolate drug resistance marker genes among Plasmodium vivax isolates exhibiting severe manifestations. Exp Parasitol. 2012, 132: 410-416. 10.1016/j.exppara.2012.09.018.
Lu F, Wang B, Cao J, Sattabongkot J, Zhou H, Zhu G, Kim K, Gao Q, Han ET: Prevalence of drug resistance-associated gene mutations in Plasmodium vivax in central China. Korean J Parasitol. 2012, 50: 379-384. 10.3347/kjp.2012.50.4.379.
Imwong M, Pukrittayakamee S, Pongtavornpinyo W, Nakeesathit S, Nair S, Newton P, Nosten F, Anderson TJ, Dondorp A, Day NP, White NJ: Gene amplification of the multidrug resistance 1 gene of Plasmodium vivax isolates from Thailand, Laos, and Myanmar. Antimicrob Agents Chemother. 2008, 52: 2657-2659. 10.1128/AAC.01459-07.
Mint Lekweiry K, Ould Mohamed Salem Boukhary A, Gaillard T, Wurtz N, Bogreau H, Hafid JE, Trape JF, Bouchiba H, Ould Ahmedou Salem MS, Pradines B, Rogier C, Basco LK, Briolant S: Molecular surveillance of drug-resistant Plasmodium vivax using pvdhfr, pvdhps and pvmdr-1 markers in Nouakchott, Mauritania. J Antimicrob Chemother. 2012, 67: 367-374. 10.1093/jac/dkr464.
Barnadas C, Ratsimbasoa A, Tichit M, Bouchier C, Jahevitra M, Picot S, Ménard D: Plasmodium vivax resistance to chloroquine in Madagascar: clinical efficacy and polymorphisms in pvmdr-1 and pvcrt-o genes. Antimicrob Agents Chemother. 2008, 52: 4233-4240. 10.1128/AAC.00578-08.
Hawkins VN, Joshi H, Rungsihirunrat K, Na-Bangchang K, Sibley CH: Antifolates can have a role in the treatment of Plasmodium vivax. Trends Parasitol. 2007, 23: 213-222. 10.1016/j.pt.2007.03.002.
Alam MT, Bora H, Bharti PK, Saifi MA, Das MK, Dev V, Kumar A, Singh N, Dash AP, Das B, Wajihullah , Sharma YD: Similar trends of pyrimethamine resistance-associated mutations in Plasmodium vivax and P. falciparum. Antimicrob Agents Chemother. 2007, 51: 857-863. 10.1128/AAC.01200-06.
Auliff A, Wilson DW, Russell B, Gao Q, Chen N, le Anh N, Maguire J, Bell D, O’Neil MT, Cheng Q: Amino acid mutations in Plasmodium vivax dhfr and dhps from several geographical regions and susceptibility to antifolate drugs. Am J Trop Med Hyg. 2006, 75: 617-621.
Zakeri S, Motmaen SR, Afsharpad M, Djadid ND: Molecular characterization of antifolates resistance-associated genes (dhfr and dhps) in Plasmodium vivax isolates from the Middle East. Malar J. 2009, 8: 20-10.1186/1475-2875-8-20.
Tjitra E, Baker J, Suprianto S, Cheng Q, Anstey NM: Therapeutic efficacies of artesunate- sulfadoxine-pyrimethamine and chloroquine-sulfadoxine-pyrimethamine in vivax malaria pilot studies: relationship to Plasmodium vivax dhfr mutations. Antimicrob Agents Chemother. 2002, 46: 3947-3953. 10.1128/AAC.46.12.3947-3953.2002.
Hastings MD, Maguire JD, Bangs MJ, Zimmerman PA, Reeder JC, Baird JK, Sibley CH: Novel Plasmodium vivax dhfr alleles from the Indonesian archipelago and Papua, New Guinea: association with pyrimethamine resistance determined by a Saccharomyces cerevisiae expression system. Antimicrob Agents Chemother. 2005, 49: 733-740. 10.1128/AAC.49.2.733-740.2005.
Miao M, Yang Z, Cui L, Ahlum J, Huang Y, Cui L: Different allele prevalence in the dihydrofolate reductase and dihydropteroate synthase genes in Plasmodium vivax populations from China. Am J Trop Med Hyg. 2010, 83: 1206-1211. 10.4269/ajtmh.2010.10-0259.
Prajapati SK, Joshi H, Dev V, Dua VK: Molecular epidemiology of Plasmodium vivax anti-folate resistance in India. Malar J. 2011, 10: 102-10.1186/1475-2875-10-102.
Feng L, Chae SL, Deok HN, Kwonkee K, Khin L, Tong SK, Hyeong WL, Junhu C, Yue W, Jestsumon S, Eun TH: Mutations in the antifolate-resistance-associated genes dihydrofolate reductase and dihydropteroate synthase in Plasmodium vivax isolates from malaria-endemic countries. Am J Trop Med Hyg. 2010, 83: 474-479. 10.4269/ajtmh.2010.10-0004.
Zakeri S, Afsharpad M, Ghasemi F, Raeisi A, Safi N, Butt W, Atta H, Djadid ND: Molecular surveillance of Plasmodium vivax dhfr and dhps mutations in isolates from Afghanistan. Malar J. 2010, 9: 75-10.1186/1475-2875-9-75.
Zakeri S, Afsharpad M, Ghasemi F, Raeisi A, Kakar Q, Atta H, Djadid ND: Plasmodium vivax: prevalence of mutations associated with sulfadoxine–pyrimethamine resistance in Plasmodium vivax clinical isolates from Pakistan. Exp Parasitol. 2010, 127: 167-172.
Afsharpad M, Zakeri S, Pirahmadi S, Djadid ND: Molecular assessment of dhfr/dhps mutations among Plasmodium vivax clinical isolates after introduction of sulfadoxine/pyrimethamine in combination with artesunate in Iran. Infect Genet Evol. 2012, 12: 38-44. 10.1016/j.meegid.2011.10.003.
Rungsihirunrat K, Sibley CH, Mungthin M, Na-Bangchang K: Geographical distribution of amino acid mutations in Plasmodium vivax DHFR and DHPS from malaria endemic areas of Thailand. Am J Trop Med Hyg. 2008, 78: 462-467.
Na BK, Lee HW, Moon SU, In TS, Lin K, Maung M, Chung GT, Lee JK, Kim TS, Kong Y: Genetic variations of the dihydrofolate reductase gene of Plasmodium vivax in Mandalay Division, Myanmar. Parasitol Res. 2005, 96: 321-325. 10.1007/s00436-005-1364-0.
Korsinczky M, Fischer K, Chen N, Baker J, Rieckmann K, Cheng Q: Sulfadoxine resistance in Plasmodium vivax is associated with a specific amino acid in dihydropteroate synthase at the putative sulfadoxine-binding site. Antimicrob Agents Chemother. 2004, 48: 2214-2222. 10.1128/AAC.48.6.2214-2222.2004.
Imwong M, Pukrittayakamee S, Cheng Q, Moore C, Looareesuwan S, Snounou G, White NJ, Day NPJ: Limited Polymorphism in the Dihydropteroate Synthetase Gene (dhps) of Plasmodium vivax Isolates from Thailand. Antimicrob Agents Chemother. 2005, 49: 4393-4395. 10.1128/AAC.49.10.4393-4395.2005.
Staedke SG, Sendagire H, Lamola S, Kamya MR, Dorsey G, Rosenthal PJ: Relationship between age, molecular markers, and response to sulphadoxine–pyrimethamine treatment in Kampala. Uganda Trop Med Int Health. 2004, 9: 624-629. 10.1111/j.1365-3156.2004.01239.x.
Acknowledgements
We thank Brenda Rae Marshall, DPSS, NIAID, for editing. This research was supported by the Fogarty International Center of the National Institutes of Health (NIH, USA) under Award Number R01TW008151, and the Division of Intramural Research, National Institute of Allergy and Infectious Diseases, NIH. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
XZS and FL designed the study and wrote the manuscript. BH, SH, and HL carried out the field work. BH and SH performed experiments, analysed data, and wrote the manuscript. XT and JY performed the preliminary data analysis. All authors read and approved the final manuscript.
Bo Huang, Shiguang Huang contributed equally to this work.
Electronic supplementary material
12936_2014_3378_MOESM1_ESM.docx
Additional file 1: Frequency distribution of SNPs in combination of pvdhfr , pvdhps , and pvmdr-1 haplotypes associated with SP and CQ in P. vivax isolates. Description: The data provided frequency distribution of SNPs in combination of pvdhfr, pvdhps, and pvmdr-1 haplotypes. (DOCX 34 KB)
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

Cite this article
Huang, B., Huang, S., Su, Xz. et al. Molecular surveillance of pvdhfr, pvdhps, and pvmdr-1 mutations in Plasmodium vivax isolates from Yunnan and Anhui provinces of China. Malar J 13, 346 (2014). https://doi.org/10.1186/1475-2875-13-346
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1475-2875-13-346