
Abstract
Background
Anti-malarial drug resistance has emerged as one of the biggest challenges confronting the worldwide effort to control malaria. The appearance of chloroquine and multi-drug resistance had devastating effects on therapeutic efficacy of former first-line agents. Artemisinin has proven to be an excellent therapeutic alternative to fill the void in chemotherapeutic options left by resistance mechanisms. At the time of introduction, no resistance to artemisinins had been recorded, and artemisinins demonstrated excellent parasite reduction rates. In an attempt to protect artemisinin efficacy, the World Health Organization (WHO) made artemisinin-based combination therapy (ACT) its official first-line treatment recommendation for uncomplicated Plasmodium falciparum in 2006. In Brazil, artemether/lumefantrine became the Brazilian Malaria Control Programme's official treatment recommendation in 2007. The sarco/endoplasmic reticulum Ca2+ - ATPase ortholog of P. falciparum (pfatp 6) has been suggested as one of the targets of artemisinins. Consequently, pfatp 6 gene polymorphisms are being investigated as markers of artemisinin resistance elsewhere. The goal of this work was to describe the molecular profile of pfatp 6 in P. falciparum isolates from different localities in the Amazonas State.
Methods
DNA polymorphisms of the pfatp6 gene in 80 P. falciparum isolates from 11 municipalities of the Amazonas State (Western Brazilian Amazon), before and after the introduction of ACT in the Brazilian anti-malarial guidelines, were analysed by automatic sequencing. Mutations in the pfatp6 gene were searched using Mutation Surveyor v3.25 software.
Results
The P. falciparum pfatp6 gene presented polymorphisms at codons 37, 630 and 898. The R37K mutation was found in 16% of the samples, A630S in 32% and I898I in 52%. No S769N mutation, however, was detected in the analysed samples.
Conclusion
Despite the small number of samples, data presented here provide baseline information about polymorphisms of pfatp6 gene before and after exposure to ACT in a low transmission area, which will help to infer drug selection pressure in this area in the future.
Similar content being viewed by others

Background
The Brazilian Amazon Region is responsible for 99.8% of all reported malarial cases in the Brazil, where socioeconomic and environmental conditions favor the proliferation of the Anopheles darlingi mosquito. In the State of Amazonas, where this study was conducted, 32,566 cases were reported from January to July in 2010, making it the second most malaria-affected state in Brazil after Pará (51,697 cases). Plasmodium vivax accounts for 83.7% of these, but Plasmodium falciparum accounts for the majority of the remaining cases [1].
Plasmodium falciparum resistance to anti-malarial drugs, especially chloroquine and pyrimethamine-sulphadoxine, has emerged as one of the biggest challenges to be faced in malaria control [2]. Artemisinin-based combination therapy (ACT) is now the WHO recommended strategy for preventing the development of drug resistance, and, in Brazil, artemether-lumefantrine was subsequently adopted by the Brazilian Ministry of Health's first-line treatment recommendation for uncomplicated falciparum infections. The efficacy of this strategy, however, has become threatened by the discovery of delayed artemisinin sensitivity on the Thai-Cambodian border [3]. The identification and monitoring of indicators of artemisinin resistance is essential for preservation of ACT efficacy [4]. Two genes, originally shown to modulate sensitivity to chloroquine in P. falciparum, have been investigated in the context of artemisinin resistance: pfmdr1 and pfcrt, but failed to correlate with the clinical phenotype of delayed parasite clearance. A previous study, however, with P. falciparum suggested that a sarcoplasmic and endoplasmic reticulum Ca2+-ATPase (SERCA)-type protein encoded by the gene pfatp6 might be the primary target of these drugs [5] (Figure 1), and mutations in this gene may alter P. falciparum's sensitivity to artemisinin.

Pfatpase6 --Membrane protein topology and aminoacid substitutions, evidencing the 769 polymorphism found in samples from the French Guyana (in the Eastern Amazon).
Jambou et al. reported a significant decrease in in vitro sensitivity to artemether in P. falciparum isolates from French Guiana, located along Brazil's northern border [6]. This reduced efficacy was associated with a S769N polymorphism in the PfATPase6 protein. Although the significance of this mutation has not been determined, these observations could indicate that the S769N mutation could be a marker of drug resistance. French Guyana's proximity to Brazil makes migration of drug resistant parasite populations highly possible [7].
Other polymorphisms have been identified in the pfatp6 gene: the double mutation E431K, A623E in Senegal, I89T in Thailand [8], H243Y in Central Africa [9] and T2694 in São Tomé and Principe [10]. In 2008, Dahlstrom et al. [11], studying P. falciparum isolates from East and West Africa, identified 33 single nucleotide polymorphisms (SNPs), three of which were found in a frequency higher than 5% in codons H431K, N569K and A630S.
Data on the pfatp6 gene in Brazilian isolates is scarce. Recently, three mutations were described in samples from the State of Pará, in nucleotide positions 110, 1916 and 2694 [7]. This study reports pfatp6 gene polymorphisms in P. falciparum isolates collected before and after the introduction of ACT in eleven endemic municipalities.
Methods
Study design
This is an ecological study aimed at evaluating changes in the molecular profile of P. falciparum between two distinct time points: before and after the introduction of ACT in Brazil.
Samples
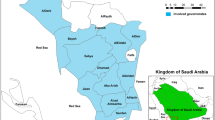
Plasmodium falciparum field isolates collected between 2000 and 2005 (60 samples; 10 per year) were obtained from the FMT-HVD Laboratory of Malaria. Samples from between 2009 and 2010 (20 samples; 10 per year) were prospectively collected. Samples from both time points come from the same eleven endemic municipalities of the Amazonas State (Figure 2). Samples were collected as follows: two from Autazes, one from Barcelos, one from Borba, three from Careiro, three from Coari, two from Guajará, two from Humaitá, six from Itacoatiara, 53 samples from Manaus, one from São Gabriel da Cachoeira and six from Tefé. Isolates were collected from patients with uncomplicated P. falciparum infections before unsupervised anti-malarial treatment (quinine/doxycycline from 2000 to 2005 and artemether/lumefantrine from 2009 to 2010). As a tertiary care and referral center for infectious diseases in the State of Amazonas, the Fundação de Medicina Tropical Dr. Heitor Vieira Dourado (FMT-HVD) routinely follows all patients diagnosed with malaria for 42 days since the middle 1990s. Patients come to the outpatient clinics on days 0, 3, 7, 14, 28 and 42 for clinical evaluation and thick blood smear collection, as part of a protocol to characterize anti-malarial resistance. Diagnosis was carried out by microscopic examination of Giemsa-stained thick blood films as recommended by the Brazilian Malaria Control Programme. After confirming P. falciparum mono-infection, 5 mL of venous blood were collected into an EDTA-containing tube. The tube was centrifuged to separate plasma, and erythrocytes were stored at -20°C until DNA extraction.

Map of the Amazonas State showing the municipalities where samples originated (1. São Gabriel da Cachoeira; 2. Barcelos; 3. Manaus; 4. Itacoatiara; 5. Careiro; 6. Coari; 7. Tefé; 8. Humaitá; 9. Guajará; 10. Autazes; 11. Borba).
DNA extraction
Extraction of DNA from patient blood samples infected with P. falciparum was carried out using a Genomic DNA Purification kit (Wizard®, USA) according to the manufacturer's protocol.
PCR amplification and sequencing
Five pairs of oligonucleotide primers (Table 1) were designed for amplification of five genomic sequences between nucleotides 28 to 3078 of the pfatp6 gene. The fragments were amplified by PCR and the quality of PCR products generated for each fragment was determined by 1% agarose gel electrophoresis and visualized by staining with ethidium bromide. The DNA concentration was measured by NanoDrop® 2000 (Thermo Scientific). The template for DNA sequencing following purification was carried with a commercial kit (SV Gel and PCR Clean-Up System, Wizard®, USA).
Sequencing reactions were carried out using an ABI 3130xl genetic analyzer (Applied Biosystems®, USA) as specified by manufacturer's protocol. DNA sequences were generated from both sense and antisense primers, aligned to check for genetic polymorphism and compared to the reference sequence of clone 3D7 (accession PFA0310c in http://www.plasmodb.org). PCR primers and reaction conditions have been published elsewhere [10].
Data analysis
Sequence quality was assessed using FinchTV® v.1.4.0 (Geospiza®) software. Analysis of polymorphisms was performed in Mutation Surveyor® v.3.97 (SoftGenetics®, LLC).
Ethical approval
This study, which used retrospective samples (2000-2005) and prospectively collected samples (2009-2010), received ethical approval from the Ethics Committee Board of the FMT-HVD (protocol 3080-08).
Results
Samples from 80 patients with acute P. falciparum malaria ranging from 13 to 80 years old (71% of which were males) were studied. No instances of treatment failure or recrudescence were observed in the 42-day follow-up period.
PCR amplification and sequencing of P. falciparum genomic DNA was successful in all isolates included in this study. A total of three polymorphisms in the pfatp6 gene were identified: two non-synonymous (R37K and A630S) and one synonymous (I898I). The mutation at codon 37 lead to the amino acid change Arginine → Lysine. While the second mutation at codon 630 lead to the amino acid change Alanine → Serine. The third mutation at codon 898, however, did not lead to any amino acid change. The prevalence of each mutation was: 16% for R37K, 32% for A630S and 52% for I898I (Figure 3). The G2306A mutation (encoding S769N) was absent in all the samples.

Polymorphism frequencies by year in the pfatp6 gene from samples from different municipalities of the Amazonas State (10 samples analysed per year).
Discussion
The genetic basis of resistance to anti-malarial drugs can be investigated in several ways. This study describes for the first time a pfatp6 gene sequence polymorphism in Amazonas State's samples in two distinct periods: before and after the introduction of artemisinins. No clear change in polymorphism distribution, however, was observed between the two periods. The major limitation of this study was the small number of samples due to the declining number of P. falciparum infections. This decline has been particularly pronounced since the introduction of ACTs as first-line treatment in 2007.
A study of the susceptibility to artemisinins and derivatives and molecular analysis of SNPs in Africa, found a silent mutation T2694A in isolates of São Tomé and Príncipe [10]. In this present study, 40 samples from different municipalities presented this mutation, and were similarly found to be phenotypically insignificant.
In 2008, Ferreira et al. also published a similar paper with Brazilian samples [7]. They described the analysis of four SNPs in isolates from Pará in nucleotide positions: 110, 1916, 2306 and 2694. In the present study, no mutations were detected in nucleotides 1916 and 2306. The mutations in nucleotides 110 and 2694, however, were observed with frequencies of 16% and 52% respectively.
Polymorphisms similar to the third observed mutation at nucleotide 1888 in codon 630 in 32% of isolates, were detected in Nigerian samples (A630S), with a frequency of 1.1% [12]. The association between the mutation occurrence and external factors (environment and human host) was tested by bivariate analysis and showed no correlation.
The G2306A mutation (encoding S769N), found in the French Guiana (Eastern Amazon) [6] was absent in the present samples. That is probably explained by the fact that not much immigration occurs from that region to the Amazonas State (Western Brazilian Amazon). Actually the Amazon represents a wide and diverse area with distinct transmission scenarios and still with reduced population mobility, which may parallel the low dissemination of locally originated mutations [13].
Pfmdr1 copy number was not assessed in this study. Though pfmdr1 copy number is well correlated with decreased therapeutic efficacy of many drugs and found in South America, mefloquine-based ACT is not a first-line agent in Brazil. Therefore, pfmdr1 is less likely to be under mefloquine pressure, despite the recent observation that in vitro response to lumefantrine could also be associated to pfmdr1 mutations [14]. More genes, however, in larger samples from a representative portion of the Amazon should be urgently investigated.
Considering the implication of pfatp6 in artemisinin resistance, the molecular variability of this gene should be carefully monitored in areas where P. falciparum poses a public health threat. Losing artemisinin efficacy would have a devastating effect on malaria control and treatment efforts, because there is no equivalent oral therapeutic. The gravity of these implications warrants intensive phenotypic and molecular monitoring of field isolates.
Conclusion
Pfatp6 S769N mutation, a potential molecular marker of artemisinin resistance, was not present in the few analysed samples, as well as changes in polymorphism prevalence in P. falciparum isolates from Amazonas State since the introduction of ACT in 2007. These data provide a baseline level of genetic variation in a potential candidate gene for artemisinin resistance and will be of value in future resistance monitoring studies. Identification and monitoring of polymorphisms and mutations that confer drug resistance to P. falciparum is essential to the success of malaria prevention programmes.
References
The epidemiological situation of malaria in Brazil. [http://www.saude.gov.br/svs]
Bray PG, Ward SA, O'Neill PM: Quinolines and artemisinin: chemistry, biology and history. Curr Top Microbiol Immunol. 2005, 295: 3-38. 10.1007/3-540-29088-5_1.
Dondorp AM, Nosten F, Yi P, Das D, Phyo AP, Tarning J, Lwin KM, Ariey F, Hanpithakpong W, Lee SJ, Ringwald P, Silamut K, Imwong M, Chotivanich K, Lim K, Herdman T, An SS, Yeung S, Singhasivanon P, Day NPJ, Lindegardh N, Socheat D, White NJ: Artemisinin resistance in Plasmodium falciparum malaria. N Engl J Med. 2009, 361: 455-467. 10.1056/NEJMoa0808859.
Afonso A, Hunt P, Cheesman S, Alves AC, Cunha CV, Rosario V, Cravo P: Malaria parasites can develop stable resistance to artemisinin but lack mutations in candidate genes atp6 (encoding the sarcoplasmic and endoplasmic reticulum Ca2+ ATPase), tctp, mdr1, and cg10. Antimicrob Agents Chemother. 2006, 50: 480-489. 10.1128/AAC.50.2.480-489.2006.
Eckstein-Ludwig U, Webb RJ, Van Goethem ID, East JM, Lee AG, Kimura M, O'Neill PM, Bray PG, Ward SA, Krishna S: Artemisinins target the SERCA of Plasmodium falciparum. Nature. 2003, 424: 957-961. 10.1038/nature01813.
Jambou R, Legrand E, Niang M, Khim N, Lim P, Volney B, Ekala MT, Bouchier C, Esterre P, Fandeur T, Mercereau-Puijalon O: Resistance of Plasmodium falciparum field isolates to in-vitro artemether and point mutations of the SERCA-type PfATPase6. Lancet. 2005, 366: 1960-1963. 10.1016/S0140-6736(05)67787-2.
Ferreira ID, Martinelli A, Rodrigues LA, do Carmo EL, Rosario VE, Povoa MM, Cravo P: Plasmodium falciparum from Para state (Brazil) shows satisfactory in vitro response to artemisinin derivatives and absence of the S769N mutation in the SERCA-type PfATPase6. Trop Med Int Health. 2008, 13: 199-207. 10.1111/j.1365-3156.2007.01991.x.
Price RN, Uhlemann AC, Brockman A, McGready R, Ashley E, Phaipun L, Patel R, Laing K, Looareesuwan S, White NJ, Nosten F, Krishna S: Mefloquine resistance in Plasmodium falciparum and increased pfmdr1 gene copy number. Lancet. 2004, 364: 438-447. 10.1016/S0140-6736(04)16767-6.
Cojean S, Hubert V, Le Bras J, Durand R: Resistance to dihydroartemisinin. Emerg Infect Dis. 2006, 12: 1798-1799. 10.3201/eid1211.060903.
Ferreira ID, Lopes D, Martinelli A, Ferreira C, Rosario VE, Cravo P: In vitro assessment of artesunate, artemether and amodiaquine susceptibility and molecular analysis of putative resistance-associated mutations of Plasmodium falciparum from Sao Tomé and Principe. Trop Med Int Health. 2007, 12: 353-362. 10.1111/j.1365-3156.2006.01789.x.
Dahlstrom S, Veiga MI, Ferreira P, Martensson A, Kaneko A, Andersson B, Bjorkman A, Gil JP: Diversity of the sarco/endoplasmic reticulum Ca(2+)-ATPase orthologue of Plasmodium falciparum (PfATP6). Infect Genet Evol. 2008, 8: 340-345. 10.1016/j.meegid.2008.02.002.
Ibrahim ML, Khim N, Adam HH, Ariey F, Duchemin JB: Polymorphism of PfATPase in Niger: detection of three new point mutations. Malar J. 2009, 8: 28-10.1186/1475-2875-8-28.
Griffing SM, Mixson-Hayden T, Sridaran S, Alam MT, McCollum AM, Cabezas C, Marquino Quezada W, Barnwell JW, Oliveira AM, Lucas C, Arrospide N, Escalante AA, Bacon DJ, Udhayakumar V: South American Plasmodium falciparum after the malaria eradication era: clonal population expansion and survival of the fittest hybrids. PLoS One. 2011, 6: e23486-10.1371/journal.pone.0023486.
Nkhoma S, Nair S, Mukaka M, Molyneux ME, Ward SA, Anderson TJ: Parasites bearing a single copy of the multi-drug resistance gene (pfmdr-1) with wild-type SNPs predominate amongst Plasmodium falciparum isolates from Malawi. Acta Trop. 2009, 111: 78-81. 10.1016/j.actatropica.2009.01.011.
Acknowledgements
The study received the financial support of PRONEX Malaria Network, funded by the Brazilian Ministry of Science and Technology (MCT), National Council for Scientific and Technological Development (CNPq), Brazilian Ministry of Health (DECIT/SCTIE/MS) and the Research Support Foundation from the Amazonas State (FAPEAM) (grant number 555.666/2009-3). MVGL, WMRO and MGZ are level 2 fellows from CNPq. Patients are also acknowledged for their participation, as well as Nelida Sanchez for technical help and Carolina Bustamante for the creation of the map.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
LWB performed laboratory work and drafted the manuscript. ALLA helped in laboratory work and CMCO was responsible for sequencing reactions. GCM, MGCA, MVGL, WMRO, CO and MGZ participated in its design, coordination and elaborated the final version of manuscript. All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Brasil, L.W., Areas, A.L., Melo, G.C. et al. Pfatp6 molecular profile of Plasmodium falciparum isolates in the western Brazilian Amazon. Malar J 11, 111 (2012). https://doi.org/10.1186/1475-2875-11-111
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1475-2875-11-111