
Abstract
Background
Fermentation of lignocellulosic biomass is an attractive alternative for the production of bioethanol. Traditionally, the yeast Saccharomyces cerevisiae is used in industrial ethanol fermentations. However, S. cerevisiae is naturally not able to ferment the pentose sugars D-xylose and L-arabinose, which are present in high amounts in lignocellulosic raw materials.
Results
We describe the engineering of laboratory and industrial S. cerevisiae strains to co-ferment the pentose sugars D-xylose and L-arabinose. Introduction of a fungal xylose and a bacterial arabinose pathway resulted in strains able to grow on both pentose sugars. Introduction of a xylose pathway into an arabinose-fermenting laboratory strain resulted in nearly complete conversion of arabinose into arabitol due to the L-arabinose reductase activity of the xylose reductase. The industrial strain displayed lower arabitol yield and increased ethanol yield from xylose and arabinose.
Conclusion
Our work demonstrates simultaneous co-utilization of xylose and arabinose in recombinant strains of S. cerevisiae. In addition, the co-utilization of arabinose together with xylose significantly reduced formation of the by-product xylitol, which contributed to improved ethanol production.
Similar content being viewed by others

Introduction
Large interest in biofuel ethanol production from renewable sources has led in numerous attempts to construct microbial strains that efficiently ferment lignocellulose hydrolysate to ethanol [1, 2]. The optimal micro-organism must be robust, ethanol and inhibitor tolerant, viable at low pH, and produce high ethanol yields from all sugars present in lignocellulose [3]. The baker's yeast Saccharomyces cerevisiae is very suitable for industrial ethanol production, however, it cannot utilize pentoses that represent a significant fraction of the sugars present in lignocellulosic raw materials. As examples, 16% xylan and 5% arabinan are found in grass, 19% xylan and 3% arabinan are found in corn stover, and as high as 15% arabinan and 19% xylan are found in wheat bran [4]. Utilization of all available substrate is a prerequisite for an economically feasible process, and even small increases in substrate utilization may significantly improve the overall process [5].
Several genetic engineering strategies have been used in attempt to enable xylose or arabinose utilization by S. cerevisiae. Xylose fermentation has been achieved by expressing Pichia stipitis xylose reductase (XR) and xylitol dehydrogenase (XDH) together with overexpression of the endogenous xylulokinase (XK) [6–8]. Xylose growth and fermentation have also been achieved by expressing xylose isomerase from bacterial [9] or fungal [10] sources. Arabinose utilization in S. cerevisiae has been enabled by expressing bacterial [11] or fungal [12] arabinose pathways. Efficient growth and fermentation of arabinose was achieved with high-level expression of the bacterial arabinose pathway consisting of Bacillus subtilis L-arabinose isomerase (AraA), Escherichia coli L-ribulokinase (AraB) and E. coli L-ribulose-5-P 4-epimerase (AraD), together with the endogenous pentose-transporting permease gene GAL2, followed by selection for improved arabinose growth by sequential transfer in arabinose medium [11].
To the best of our knowledge, no S. cerevisiae strain with demonstrated co-utilization of xylose and arabinose has previously been developed. E. coli is naturally able to utilize both xylose and arabinose, and it has been engineered to redirect the product formation towards ethanol [13]. Inversely, co-utilization of xylose and arabinose was achieved in the ethanol-producing bacterium Zymomonas mobilis by introducing the bacterial arabinose and xylose utilization pathways [14]. However, the industrial use of bacteria for the fermentation of lignocellulose hydrolysates remains problematic due to their low tolerance towards inhibitors [15].
In this work, we developed and characterized yeast strains co-consuming and co-fermenting both pentose sugars. The XR, XDH and XK activities together with the bacterial arabinose pathway were first introduced in a S. cerevisiae laboratory CEN.PK strain [16] to demonstrate the functional co-utilization of the two pentoses. A method for multiple integration of genes in the ribosomal DNA (rDNA) [17] was applied to construct stable high-level expression of the arabinose genes in industrial yeast strains. Using this tool, an arabinose and xylose utilizing industrial strain based on the xylose-fermenting strain TMB 3400 [18] was developed.
Results
Development of a S. cerevisiae laboratory strain co-consuming D-xylose and L-arabinose
To demonstrate functional co-utilization of xylose and arabinose by recombinant yeast, the metabolic pathways for both xylose and arabinose utilization were expressed in a S. cerevisiae laboratory strain. The construct was based on the arabinose-fermenting strain JBY25-4M [11] (Table 1), which contains the bacterial arabinose pathway consisting of the genes for L-arabinose isomerase (AraA), L-ribulokinase (AraB) and L-ribulose-5-P 4-epimerase (AraD) as well as the overexpressed endogenous galactose transporter gene (GAL2), each on a separate multicopy plasmid, and which also has beneficial mutations resulting from prolonged culturing in arabinose medium [11]. One mutation resulted in the amplification of the TAL1 gene [11], but other unknown mutations are possibly also present. Because the strain JBY25-4M can retain the four plasmids only in the presence of a selection pressure for arabinose utilization, the plasmids of JBY25-4M were removed by cultivation in rich medium before further genetic modification. In the resulting strain, the xylose pathway genes were introduced using the integrative vector YIpXR/XDH/XK [7] (Table 1) which contains the P. stipitis xylose pathway genes coding for XR (XYL1), XDH (XYL2) and the endogenous XK (XKS1), selecting for histidine prototrophy. The resulting strain was named BWY02.X. Presence of the functional xylose pathway was verified by xylose growth. A prototrophic control strain without the arabinose pathway was constructed by introducing three empty plasmids, p424H7, p425H7, and p426H7 [11] (Table 1) without the arabinose genes into BWY02.X, resulting in strain BWY02.Xp. To generate a xylose- and arabinose-consuming strain, three multicopy plasmids carrying the three bacterial arabinose pathway genes were then introduced in BWY02.X. B. subtilis AraA gene, mutated E. coli AraB gene and E. coli AraD gene were introduced in BWY02.X. using plasmids YEpURAaraA (Table 1), YEparaB [11] and YEparaD [11], respectively, selecting for prototrophy and growth on arabinose. The resulting strain was named BWY02.XA.
The functional xylose pathway in BWY02.XA was verified by xylose growth. BWY02.XA grew aerobically on 50 g/l xylose with a growth rate of 0.013 h-1 (data not shown). Similar in vitro activities of XR and XDH were obtained for BWY02.XA and the xylose-growing control strain TMB3001 [7]. For BWY02.XA, the XR activity was 0.08 ± 0.01 U mg protein-1 and the XDH activity was 1.36 ± 0.07 U mg protein-1, whereas the activities for TMB 3001 were 0.10 ± 0.01 U mg protein-1 and 1.01 ± 0.08 U mg protein-1, respectively. Very low activities of 0.007 ± 0.002 U mg protein-1 (XR) and 0.004 ± 0.001 U mg protein-1 (XDH) were measured in the negative control strain CEN.PK113-7D. The presence of an active arabinose pathway in BWY02.XA was demonstrated by aerobic growth on arabinose. In vitro activities for the arabinose pathway enzymes could not be measured due to the reagents required not being commercially available. BWY02.XA grew in aerobic shake-flask cultures with 50 g/l arabinose at a growth rate of 0.05 h-1 until an OD620 nm of about 6, whereas the control strain BWY02.Xp did not grow on arabinose (Figure 1A). BWY02.XA utilized arabinose and xylose simultaneously in aerobic shake-flask cultures containing a mixture of 50 g/l xylose and 50 g/l arabinose. Arabinose was the preferred substrate, with 35% of the arabinose and 15% of the xylose being consumed within 95 hours (data not shown).

Aerobic growth of S. cerevisiae strains in YNB medium with 50 g/l arabinose as the sole carbon source. A. Laboratory strains BWY02.Xp (◆) (xylose pathway and empty multicopy plasmids) and BWY02.XA (■) (xylose and arabinose pathways). B. Industrial strains TMB 3400 (■) (xylose pathway), TMB 3061 (◆)(xylose and arabinose pathways), and TMB 3063 (▲) (xylose and arabinose pathways).
Genetic tools for stable genomic integration of the arabinose pathway genes in industrial S. cerevisiae strains
Previously, the best arabinose utilization by a laboratory strain has been obtained by expressing the AraA and AraD genes from multi-copy plasmids and the L-ribulokinase gene AraB either in a mutated form with reduced activity or from a single copy centromeric vector [11]. In order to reproduce this pattern in industrial yeast strains, genetic tools aiming at multiple chromosomal integration of the AraA and AraD genes were constructed. The rDNA region of the S. cerevisiae genome was chosen as target for multiple integration because the rDNA region is present at about 140 copies in the yeast genome [19] and the strategy has previously been successfully applied [20–22]. Plasmid prDNA2 (Figure 2B) containing two adjacent 500 bp long sequences from the S. cerevisiae rDNA was first constructed (see Materials and Methods). The AraA and AraD genes were separately cloned in prDNA2, resulting in two plasmids prDNA-AraA and prDNA-AraD (Figure 2B), respectively. The plasmids were then used as a template for creating PCR-fragments to be used for transformation, consisting of either of the genes flanked by the rDNA-sequences. Instead of the more commonly used linearized plasmids, PCR-fragments were preferred for yeast transformation to avoid introducing unnecessary antibiotic markers or other vector sequences, as well as to keep the introduced sequence as short as possible.

Plasmids used in construction of the industrial strains TMB 3061 and TMB 3063. A. Integrative vector YIpAraB. B. Multicopy plasmid prDNA2 and the derivative plasmids prDNAAraA and prDNAAraD that are used as templates for PCR-amplification.
Construction of a stable industrial S. cerevisiae strain co-consuming xylose and arabinose
The chromosomal integration of the arabinose pathway genes was applied on the xylose-fermenting industrial strain TMB 3400 [18]. First, single integration of the mutated E. coli L-ribulokinase gene AraB [11] was performed on TMB 3400 using the vector YIpAraB (Figure 2A) linearized by Eco81I, resulting in integration in the TRP1-locus. Transformants were selected on geneticin, and the presence of the AraB gene was confirmed by PCR. The resulting yeast strain was named TMB 3060. For multiple integration, the AraA and AraD genes flanked by the rDNA target sequences were PCR-amplified from plasmids prDNA-AraA or prDNA-AraD (Figure 2B) and both PCR fragments were transformed simultaneously into TMB 3060. Since introduction of these two genes completed the arabinose utilization pathway, the transformants were selected for growth on arabinose. One transformant was purified and the resulting strain was named TMB 3061.
To verify functional arabinose utilization in TMB 3061, the aerobic arabinose growth rates of TMB 3400 and TMB 3061 were compared. TMB 3061 grew with a maximum specific growth rate of 0.06 h-1 in YNB medium with 50 g/l arabinose, whereas the parental strain TMB 3400 did not grow on arabinose (Figure 1B). Growth of TMB 3061 stopped at an OD620 nm of about 3, indicating presence of a limiting step in the existing metabolic pathway. In aerobic shake-flask cultures using YNB medium with a mixture of 25 g/l xylose and 25 g/l arabinose as carbon sources, TMB 3061 consumed arabinose and xylose simultaneously, xylose being the preferred sugar (Figure 3). No arabinose consumption, but nearly complete xylose consumption was observed for TMB 3400 (data not shown).

Growth of S. cerevisiae TMB 3061 (xylose and arabinose-growing industrial strain) in mineral medium containing a mixture of 25 g/l arabinose and 25 g/l xylose in aerobic batch culture and co-consumption of arabinose and xylose. Left axis: arabinose (▲), xylose (■) (g/l), right axis: OD620 (◆).
In order to further improve arabinose utilization, TMB 3061 was re-transformed with the AraA gene encoding L-arabinose isomerase, since previous work on JBY25-4M showed that the number of the plasmid carrying the AraA gene exceeded the numbers of the other plasmids, indicating that L-arabinose isomerase activity may be limiting for growth on arabinose [11]. Transformants with improved arabinose growth were selected in liquid YNB medium containing 20 g/l arabinose. After three rounds of sequential transfer to new liquid cultures, aliquots were plated on arabinose plates and single colonies were purified and analyzed for arabinose growth. The best clone was named TMB 3063. Strain TMB 3063 grew on 50 g/l arabinose with a slightly reduced growth rate of 0.04 h-1, but reached a significantly increased final OD620 nm of 6 (Figure 1B), which is the same as for the laboratory strain BWY02.XA carrying the arabinose pathway genes on multicopy vectors (Figure 1A).
The effect of the presence of the arabinose pathway on xylose utilization was also tested. TMB 3061 and TMB 3063 grew on xylose with growth rates of 0.03 h-1 and 0.012 h-1, respectively, compared with 0.08 h-1 for the parental strain TMB 3400. The decrease in xylose growth could not be explained by a decrease in the XR or XDH levels, since both arabinose-growing strains TMB 3061 and TMB 3063 had similar XR and XDH activities than the parental strain TMB 3400. The XR activities were 0.13 ± 0.01 and 0.10 ± 0.01 for TMB 3061 and TMB 3063, respectively, compared with 0.09 ± 0.00 for TMB 3400. The XDH activities were 0.52 ± 0.03 and 0.81 ± 0.08 for TMB 3061 and TMB 3063, respectively, and 0.59 ± 0.08 for TMB 3400.
Fermentation of xylose and arabinose
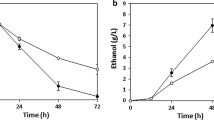
Xylose and arabinose fermentation by strain BWY02.XA was independently investigated in anaerobic batch fermentation in defined mineral medium containing either 20 g/l glucose and 20 g/l xylose or 20 g/l glucose and 20 g/l arabinose (Table 2, Figure 4A–B). Glucose was included to support biomass formation, since BWY02.XA did not grow anaerobically on xylose or arabinose. Product yields were determined in the pentose phase of the fermentation, i.e. after glucose depletion. About 20% of the arabinose was consumed during the arabinose fermentation (Figure 4A) with a rate of 0.03 g arabinose h-1 g cells-1 (Table 2), whereas nearly all xylose was consumed during the xylose fermentation (Figure 4B) with a rate of 0.1 g xylose h-1 g cells-1 (Table 2). Ethanol was produced from xylose with the yield of 0.23 g ethanol g consumed xylose-1during the xylose phase of the xylose fermentation (Table 2), whereas no ethanol was produced from arabinose.

Anaerobic batch fermentations by recombinant S. cerevisiae in defined mineral medium with 20 g/l glucose and 20 g/l arabinose (A), 20 g/l glucose and 20 g/l arabinose (B) or 20 g/l glucose, 20 g/l xylose and 20 g/l arabinose (C-E) as carbon sources. A. B. C. BWY02.XA (xylose and arabinose-growing laboratory strain); D. TMB 3400 (xylose-growing industrial strain); E. TMB 3063 (xylose and arabinose-growing industrial strain). Symbols: ○ glucose; □ xylose; △ arabinose; × xylitol; + arabitol; ▲ glycerol; ◆ ethanol; ■ biomass.
Co-fermentation of arabinose and xylose was studied by performing anaerobic batch fermentation using a mixture of 20 g/l glucose, 20 g/l xylose and 20 g/l arabinose as carbon sources. The laboratory strain BWY02.XA and the industrial strains TMB 3400 and TMB 3063 were tested in this set-up (Figure 4C–4E). The specific xylose and arabinose consumption rates and product yields calculated from the pentose phase of the fermentation are summarized in Table 2. Strain BWY02.XA (Figure 4C) consumed arabinose and xylose simultaneously with approximately the same consumption rates of ca. 0.04 g h-1 g cells-1 (Table 2). Nearly half of both arabinose and xylose was consumed (Figure 4C). The xylitol yield on consumed xylose was 0.33 g xylitol g xylose-1, whereas the arabitol yield on consumed arabinose was about 1 g arabitol g arabinose-1 (Table 2).
TMB 3400 (Figure 4D) consumed nearly all available xylose (about 20 g/l) and about 3 g/l of the arabinose, whereas TMB 3063 (Figure 4E) consumed slightly less xylose (16 g/l) but much more arabinose (11 g/l). TMB 3400 and TMB 3063 had xylose consumption rates of 0.066 g xylose h-1 g cells-1 and 0.042 g xylose h-1 g cells-1, respectively. The arabinose consumption rate of TMB 3400 was 0.010 g arabinose h-1 g cells as compared to 0.029 g arabinose h-1 g cells for TMB 3063 (Table 2). Considerable amount of xylose was consumed in the presence of glucose in both strains, whereas arabinose consumption started only after glucose depletion (Figure 4E). In both strains, ethanol was produced from pentose sugars. The final ethanol concentration was highest for the xylose- and arabinose-fermenting TMB 3063 (Table 2), despite a reduced xylose consumption rate. Reduced xylitol and arabitol formation was observed in TMB 3063 (Table 2), accompanied by improved ethanol yield from pentoses.
Discussion
D-Xylose or L-arabinose-consuming S. cerevisiae strains have previously been independently developed by introduction of heterologous enzymes necessary for the assimilation of either of the sugars [6, 7, 10–12]. However, co-fermentation of the two pentose sugars by S. cerevisiae has not, to the best of our knowledge, previously been reported. We have introduced both the arabinose and the xylose pathways in S. cerevisiae strains to investigate the effects and possible limitations of the combined metabolism of the two pentoses. Xylose utilization was enabled by overexpression of the P. stipitis genes coding for XR and XDH as well as the endogenous XK through chromosomal integration. Arabinose assimilation was enabled through heterologous expression of the bacterial arabinose pathway consisting of L-arabinose isomerase (AraA), L-ribulokinase (AraB) and L-ribulose-5-P-4-epimerase (AraD) genes.
The combination of xylose and arabinose pathways was first tested in a laboratory strain. However, industrial fermentation of lignocellulosic substrates requires robust industrial S. cerevisiae strains because chemical detoxification of the industrial substrates is not economically feasible [5]. The xylose-fermenting strain TMB 3400 has been found to be inhibitor tolerant [23] and an efficient ethanol producer in undetoxified lignocellulose hydrolysate [24]. Therefore, the first industrial arabinose and xylose co-fermenting S. cerevisiae strain was constructed by introducing the bacterial arabinose pathway in TMB 3400 [18].
AraB was introduced as a single copy in both strain backgrounds to avoid the negative effects of kinase overexpression [25, 26]. Since AraA and AraD were needed in high levels [11], multicopy plasmids were used for high-level expression of AraA and AraD in the laboratory strain, generating strain BWY02.XA. For the industrial strain, where plasmid usage was not applicable, a method for multiple chromosomal integration in the rDNA region was applied. The first resultant arabinose-growing strain TMB 3061 displayed limited arabinose growth compared with the strain BWY02.XA, whereas the further improved strain TMB 3063 with additional copies of the L-arabinose isomerase gene reached the growth performance of the plasmid-based laboratory strain BWY02.XA.
Co-utilization of arabinose and xylose was demonstrated with the laboratory strain BWY02.XA carrying the arabinose and the xylose pathways. BWY02.XA as well as the industrial strains TMB 3061 and TMB 3063 grew aerobically on both sugars and co-consumed both sugars simultaneously in aerobic batch culture and anaerobic batch fermentation. The simultaneous consumption of xylose and arabinose demonstrated that co-utilization of pentose sugars in S. cerevisiae differed from the case of glucose and xylose utilization, where glucose is usually consumed first and xylose consumption starts only when glucose is at least partially depleted [18]. Arabinose was the preferred sugar for BWY02.XA in aerobic cultivation on a mixture of xylose and arabinose, whereas xylose was the preferred sugar for TMB 3061. This is possibly due to the differences in gene copy numbers between the strains, since the laboratory strain carries the arabinose genes on multicopy plasmids, whereas the genes have been chromosomally integrated in TMB 3061.
Anaerobic fermentation of arabinose and xylose was tested both independently (Figure 4A–B) and as a mixture (Figure 4C) for the laboratory strain BWY02.XA. Nearly all xylose was consumed in the xylose fermentation, and ethanol was produced from xylose. In contrast, only 20% of the arabinose was consumed in the arabinose fermentation. The consumed arabinose was almost completely converted to arabitol. This is most likely due to the L-arabinose reductase activity of the P. stipitis XR [27]. XR has similar Km values for both xylose and arabinose [27, 28]. Moreover, no arabitol formation was reported in the arabinose-fermenting strain JBY25-4M [11] that did not contain the xylose pathway enzymes. In a mixture of arabinose and xylose, the xylose consumption rate, the xylitol yield and the final ethanol concentration were reduced compared with the fermentation of xylose only. The reduced ethanol level could be seen as a consequence of the reduced xylose consumption rate observed for this strain in the presence of arabinose. The reduced xylose consumption rate may in turn have resulted from the competition of the two pentose sugars for reduction by XR. It is also possible that the capacity of the pentose phosphate pathway is limiting pentose consumption. In addition, there may be competition for the unspecific pentose transporters in S. cerevisiae [29]. For example, Gal2p has been shown to transport L-arabinose [30] as well as D-xylose [29]. Thus, further improvement should be possible by overexpressing the unspecific transporter GAL2 [11], which is known to improve arabinose utilization, or by overexpressing a specific transporter for either xylose [31] or arabinose.
Introduction of the arabinose pathway in the industrial strain TMB 3400 resulted in reduced growth on xylose in strains TMB 3061 and TMB 3063, which could not be explained by decreased activities of the xylose pathway enzymes XR and/or XDH. Instead, we believe that the reduced growth results from the metabolic load observed when expressing multiple heterologous enzymes at high levels [32].
The industrial strain TMB 3063 had an improved fermentation performance compared to the parental strain TMB 3400 in fermentation of mixture of glucose, xylose and arabinose. Arabinose and xylose were consumed simultaneously under anaerobic conditions by TMB 3063 and about half of the available arabinose was now used. The ethanol yield from pentose was nearly doubled in TMB 3063, and the final ethanol concentration was increased by 15% compared to TMB 3400, despite the reduced xylose consumption of TMB 3063. The improvement in ethanol yield was accompanied by a decrease in xylitol and arabitol yields in TMB 3063. The decrease in xylitol formation in TMB 3063, as well as in the laboratory strain, is possibly a result of the competing reduction of L-arabinose by XR, which could result in larger fraction of xylose being reduced with NADH. Thus, additional genetic modification of the xylose and arabinose utilization pathways to optimize the amounts of the heterologous enzymes expressed could improve the co-fermentation of xylose and arabinose.
Conclusion
The novel arabinose- and xylose-growing strains displayed arabinose and xylose co-consumption under both aerobic and anaerobic conditions. The functional co-consumption of arabinose and xylose was demonstrated with laboratory strain BWY02.XA. Industrial xylose- and arabinose-growing strains TMB 3061 and TMB 3063 were constructed, of which TMB 3063 reached arabinose growth performance similar to BWY02.XA. Under anaerobic conditions, TMB 3063 consumed highest amount of arabinose, however, with arabitol was a considerable by-product. At the same time, the consumption of arabinose reduced the xylitol yield by 70%, which resulted in an overall increase in ethanol formation. The results show that arabinose and xylose can be co-consumed by recombinant S. cerevisiae, but efficient ethanolic fermentation of L-arabinose will require further genetic modification.
Materials and methods
Strains and media
Plasmids and yeast strains used in this study are summarized in Table 1. E. coli strain DH5α (Life Technologies, Rockville, MD, USA) was used for cloning and grown in LB-medium [33] with 100 mg/l ampicillin or 40 mg/l kanamycin. In aerobic batch cultivations, S. cerevisiae was grown in Yeast Nitrogen Base medium (YNB) (6.7 g/l Difco Yeast Nitrogen Base without amino acids; Becton, Dickinson and Company, Sparks, MD, USA) supplemented with 20 g/l glucose, 50 g/l xylose or 50 g/l arabinose as carbon sources and buffered at pH 5.5 with 10.21 g/l potassium hydrogen phthalate. When the sugar concentration in the medium exceeded 20 g/l, the concentration of YNB was doubled. In anaerobic fermentations, defined mineral medium [34] was used, supplied with 0.4 g/l Tween 80 and 0.01 g/l ergosterol and 20 g/l glucose, 20 g/l xylose and/or 20 g/l arabinose as carbon sources. Plate cultures were made on YPD (10 g/l yeast extract, 20 g/l peptone, 20 g/l glucose, 20 g/l agar) or YNB (6.7 g/l Difco YNB, 30 g/l agar, 20 g/l glucose, 50 g/l arabinose or 50 g/l xylose). When needed, 200 mg/l geneticin (Life Technologies, Rockville, Md.) was added to YPD plates.
Molecular biology techniques
Standard molecular biology techniques were used [33]. The calcium chloride method was used for bacterial transformation [35] and the lithium acetate method was used for yeast transformation [36]. Plasmids were isolated from bacteria with Quantum Prep kit (Bio-Rad, Hercules, USA). Yeast chromosomal DNA was extracted with Easy-DNA Kit (Invitrogen, Groningen, The Netherlands). DNA sequencing was performed with Abi-Prism BigDye cycle sequencing kit (Applied Biosystems, Weiterstadt, Germany). PCR was performed with the following program: 95°C for 5 min, 45 cycles of 95°C for 30 s, 50°C for 30 s and 72°C for 1 min 20 s and 72°C for 7 min. Taq-polymerase (Fermentas, Vilnius, Lithuania) was used in analytical PCR and PWO polymerase (Roche Diagnostics AB, Bromma, Sweden) in preparative PCR. T4-nucleotide ligase and restriction enzymes were purchased from Fermentas (Vilnius, Lithuania).
Plasmid construction
Plasmid YEpURAaraA (Table 1) was constructed by subcloning the B. subtilis araA gene flanked by the truncated HXT7 promoter [37] and the CYC1 terminator from plasmid YEparaA [11] into plasmid p426H7 [11].
For the construction of an integrative vector carrying the mutated E. coli AraB gene [11], the KanMX antibiotic marker [38] was first cloned into the plasmid YEparaB [11] which already contained the AraB gene. A fragment with the KanMX-marker flanked by loxP-sequences was amplified from plasmid pUG6 [38] with primers containing the ApaI restriction site and cloned in YEparaB [11]. The 2 μ-sequence and the ampicillin resistance marker in the resultant plasmid were removed by cutting with Eco105I and Eco31I followed by self-ligation of the plasmid, resulting in plasmid YIpAraB (Table 1, Figure 2A). The E. coli colonies carrying the plasmid were selected for kanamycin resistance.
Construction of the template plasmids prDNAAraA and prDNAAraD
To enable cloning of the bacterial AraA and AraD genes in between flanking sequences of S. cerevisiae rDNA sequence, a plasmid containing two adjacent regions from the S. cerevisiae rDNA was constructed. Two PCR-products were amplified from S. cerevisiae CEN.PK 113-5D rDNA NTS2 region [39] with primers containing restriction sites for KpnI (5'-AGTCGGTACC GATATGGAATGGTTGGCGAAG-3') and ApaI (5'-TCAGCCCGGG TGGCTTCCTATGCTAAATCCC-3') or SacI (5'-AGTCGAGCTC TGTTAAGTATACATGTATATATTGC-3') and SacII (5'-AGTCCCGCGG CGTTGCAAAGATGGGTTGAAAG-3'). The ~500 bp long PCR products were digested with the corresponding restriction enzymes and cloned sequentially into pBluescript SK-(Stratagene, La Jolla, CA, USA). The resulting plasmid was named prDNA2 (Figure 2B).
AraA and AraD genes were independently cloned between the rDNA sequences of prDNA2 to create a template for obtaining PCR-products consisting of either of the genes flanked by the rDNA sequences. Fragments containing AraA or AraD gene, flanked by the truncated HXT7-promoter [37] and the CYC1-terminator, were digested from plasmids YEparaA or YEparaD [11] with Eco52I and ApaI. The fragments were cloned into prDNA2, resulting in plasmids prDNAAraA and prDNAAraD (Figure 2B), respectively.
Aerobic cultivation
Pre-cultures were grown until late exponential phase in YNB medium with 20 g/l glucose except for strain BWY02.XA, which was grown on 20 g/l arabinose. Cells were washed with sterile water and inoculated at an OD620 nm of 0.2 in YNB medium with either 50 g/l arabinose, 50 g/l xylose, 20 g/l glucose, or a mixture of 25 g/l arabinose and 25 g/l xylose. 50 ml cultures were grown in 500 ml baffled shake flasks at 30°C with 180 rpm agitation in a shaker incubator (Gallenkamp INR-200, Leicester, UK). The growth rates were measured at least in duplicate with standard deviations of less than 10%.
Anaerobic batch fermentation
Anaerobic batch fermentation was performed in Biostat® bioreactors (B. Braun Biotech International, Melsungen, Germany). Precultures were performed as described above. Different combinations of 20 g/l glucose, 20 g/l xylose and/or 20 g/l arabinose were used as carbon sources. The fermentations were inoculated at an OD620 nm of about 0.2, run at 30°C, with 200 rpm stirring and pH 5.5 maintained by addition of 3 M KOH. Anaerobic conditions were maintained by sparging with nitrogen gas (containing less than 5 ppm O2, AGA, Malmö, Sweden) with the flow rate of 0.2 l min-1 controlled with a mass flow-meter (Bronkhorst, HI-TECH, Ruurlo, The Netherlands). Carbon dioxide and oxygen concentrations in the exhaust gas were measured with a Carbon Dioxide and Oxygen Monitor type 1308 (Bruel & Kjaer, Copenhagen, Denmark) in order to follow the course of the fermentation and verify anaerobiosis. The experiments were performed in duplicate.
Analyses
Concentrations of glucose, xylose, glycerol, and acetic acid were analyzed by HPLC (Beckman Instruments, Fullerton, CA, USA) using three in series connected Aminex HPX-87H ion exchange columns (Bio-Rad, Hercules, USA). The mobile phase was 5 mM H2SO4, temperature 45°C and the flow rate 0.6 ml/min. The concentrations of arabinose, arabitol and xylitol were analyzed with HPLC (Waters, Milford, Massachusetts, USA) using a HPX-87P (Bio-Rad, Hercules, USA) ion exchange column at 85°C with water as the mobile phase at a flow rate of 0.5 ml/min. Refractive index detector (RID-6A, Shimadzu, Kyoto, Japan) was used for quantification in both HPLC systems. The concentration of ethanol was calculated from the degree of reduction balance due to evaporation of the produced ethanol from the reactors. Triplicate measurements of cell dry weight were performed by filtering a known volume of the culture through a pre-weighed nitrocellulose filter with 0.45 μm pores. The filters were dried in a microwave oven and weighed.
XR and XDH activity
Cell free extracts were made from cells grown on YNB medium containing glucose. The cells were harvested in exponential growth phase, washed with sterile water and treated with yeast protein extraction reagent Y-PER according to the manufacturer's protocol (Pierce, Rockford, IL, USA). Protein concentrations were determined with Micro-BCA™ kit (Pierce, Rockford, IL, USA) using COBAS MIRA plus (Roche, Mannheim, Germany) automatic analyzer. XR and XDH activities were determined as previously described [40, 41].
Contributions of authors
KK constructed and characterized the industrial strains, participated in the design of the industrial strains, performed data interpretation and drafted the manuscript.
BW constructed and characterized the laboratory strains.
EB participated in design of the study, design of the laboratory strains and commented the manuscript.
BHH participated in design of the study and commented the manuscript.
MFGG participated in design of the study, participated in the design of the industrial strains, performed data interpretation and assisted in drafting the manuscript.
All authors have read and approved the manuscript.
References
Dien BS, Cotta MA, Jeffries TW: Bacteria engineered for fuel ethanol production: current status. Appl Microbiol Biotechnol. 2003, 63: 258-266. 10.1007/s00253-003-1444-y.
Hahn-Hägerdal B, Wahlbom CF, Gardonyi M, van Zyl WH, Cordero Otero RR, Jonsson LJ: Metabolic engineering of Saccharomyces cerevisiae for xylose utilization. Adv Biochem Eng Biotechnol. 2001, 73: 53-84.
Bothast RJ, Nichols NN, Dien BS: Fermentations with new recombinant organisms. Biotechnol Prog. 1999, 15: 867-875. 10.1021/bp990087w.
Hayn M, Steiner W, Klinger R, Steinmüller H, Sinner M, Esterbauer H: Basic research and pilot studies on the enzymatic conversion of lignocellulosics. Bioconversion of forest and agricultural plant residues. Edited by: Saddler JN. 1993, Wallingford UK, CAB international, 33-72.
von Sivers M, Zacchi G: Ethanol from lignocellulosics: a review of the economy. Biores Technol. 1996, 56: 131-140. 10.1016/0960-8524(96)00018-1.
Toivari MH, Aristidou A, Ruohonen L, Penttilä M: Conversion of xylose to ethanol by recombinant Saccharomyces cerevisiae: importance of xylulokinase (XKS1) and oxygen availability. Metab Eng. 2001, 3: 236-249. 10.1006/mben.2000.0191.
Eliasson A, Christensson C, Wahlbom CF, Hahn-Hägerdal B: Anaerobic xylose fermentation by recombinant Saccharomyces cerevisiae carrying XYL1, XYL2, and XKS1 in mineral medium chemostat cultures. Appl Environ Microbiol. 2000, 66: 3381-3386. 10.1128/AEM.66.8.3381-3386.2000.
Tantirungkij M, Nakashima N, Seki T, Yoshida T: Construction of xylose-assimilating Saccharomyces cerevisiae. J Ferment Bioeng. 1993, 75: 83-88. 10.1016/0922-338X(93)90214-S.
Karhumaa K, Hahn-Hägerdal B, Gorwa-Grauslund MF: Investigation of limiting metabolic steps in the utilization of xylose by recombinant Saccharomyces cerevisiae using metabolic engineering. Yeast. 2005, 22: 359-368. 10.1002/yea.1216.
Kuyper M, Harhangi HR, Stave AK, Winkler AA, Jetten MS, de Laat WT, den Ridder JJ, Op den Camp HJ, van Dijken JP, Pronk JT: High-level functional expression of a fungal xylose isomerase: the key to efficient ethanolic fermentation of xylose by Saccharomyces cerevisiae?. FEMS Yeast Res. 2003, 4: 69-78. 10.1016/S1567-1356(03)00141-7.
Becker J, Boles E: A modified Saccharomyces cerevisiae strain that consumes L-arabinose and produces ethanol. Appl Environ Microbiol. 2003, 69: 4144-4150. 10.1128/AEM.69.7.4144-4150.2003.
Richard P, Verho R, Putkonen M, Londesborough J, Penttilä M: Production of ethanol from L-arabinose by Saccharomyces cerevisiae containing a fungal L-arabinose pathway. FEMS Yeast Res. 2003, 3: 185-189. 10.1016/S1567-1356(02)00184-8.
Ingram LO, Conway T, Clark DP, Sewell GW, Preston JF: Genetic engineering of ethanol production in Escherichia coli. Appl Environ Microbiol. 1987, 53: 2420-2425.
Mohagheghi A, Evans K, Chou YC, Zhang M: Cofermentation of glucose, xylose, and arabinose by genomic DNA-integrated xylose/arabinose fermenting strain of Zymomonas mobilis AX101. Appl Biochem Biotechnol. 2002, 98-100: 885-898. 10.1385/ABAB:98-100:1-9:885.
Lin Y, Tanaka S: Ethanol fermentation from biomass resources: current state and prospects. Appl Microbiol Biotechnol. 2006, 69: 627-642. 10.1007/s00253-005-0229-x.
Entian KD, Kötter P: Yeast gene analysis. Edited by: Brown JPA, and Tuite MF. 1998, San Diego, California, Academic Press, 26: 431-449. Yeast mutant and plasmid collections, Methods in Microbiology,
Szostak JW, Wu R: Insertion of a genetic marker into the ribosomal DNA of yeast. Plasmid. 1979, 2: 536-554. 10.1016/0147-619X(79)90053-2.
Wahlbom CF, van Zyl WH, Jönsson LJ, Hahn-Hägerdal B, Otero RR: Generation of the improved recombinant xylose-utilizing Saccharomyces cerevisiae TMB 3400 by random mutagenesis and physiological comparison with Pichia stipitis CBS 6054. FEMS Yeast Res. 2003, 3: 319-326. 10.1016/S1567-1356(02)00206-4.
Schweizer E, MacKechnie C, Halvorson HO: The redundancy of ribosomal and transfer RNA genes in Saccharomyces cerevisiae. J Mol Biol. 1969, 40: 261-277. 10.1016/0022-2836(69)90474-4.
Fujii T, Kondo K, Shimizu F, Sone H, Tanaka J, Inoue T: Application of a ribosomal DNA integration vector in the construction of a brewer's yeast having alpha-acetolactate decarboxylase activity. Appl Environ Microbiol. 1990, 56: 997-1003.
Lin L, Ma Y, Chien HR, Hsu W: Construction of an amylolytic yeast by multiple integration of the Aspergillus awamori glucoamylase gene into a Saccharomyces cerevisiae chromosome. Enzyme Microb Technol. 1998, 23: 360-365. 10.1016/S0141-0229(98)00058-1.
Nieto A, Prieto JA, Sanz P: Stable high-copy-number integration of Aspergillus oryzae alpha-AMYLASE cDNA in an industrial baker's yeast strain. Biotechnol Prog. 1999, 15: 459-466. 10.1021/bp9900256.
Sonderegger M, Jeppsson M, Larsson C, Gorwa-Grauslund MF, Boles E, Olsson L, Spencer-Martins I, Hahn-Hägerdal B, Sauer U: Fermentation performance of engineered and evolved xylose-fermenting Saccharomyces cerevisiae strains. Biotechnol Bioeng. 2004, 87: 90-98. 10.1002/bit.20094.
Öhgren K, Bengtsson O, Gorwa-Grauslund MF, Galbe M, Hahn-Hägerdal B, Zacchi G: Simultaneous saccharification and co-fermentation of glucose and xylose in steam pretreated corn stover at high fiber concentration with recombinant Saccharomyces cerevisiae TMB 3400. submitted
Johansson B, Christensson C, Hobley T, Hahn-Hägerdal B: Xylulokinase overexpression in two strains of Saccharomyces cerevisiae also expressing xylose reductase and xylitol dehydrogenase and its effect on fermentation of xylose and lignocellulosic hydrolysate. Appl Environ Microbiol. 2001, 67: 4249-4255. 10.1128/AEM.67.9.4249-4255.2001.
Teusink B, Walsh MC, van Dam K, Westerhoff HV: The danger of metabolic pathways with turbo design. Trends Biochem Sci. 1998, 23: 162-169. 10.1016/S0968-0004(98)01205-5.
Verduyn C, Van Kleef R, Frank J, Schreuder H, Van Dijken JP, Scheffers WA: Properties of the NAD(P)H-dependent xylose reductase from the xylose-fermenting yeast Pichia stipitis. Biochem J. 1985, 226: 669-677.
Jeppsson M, Träff K, Johansson B, Hahn-Hägerdal B, Gorwa-Grauslund MF: Effect of enhanced xylose reductase activity on xylose consumption and product distribution in xylose-fermenting recombinant Saccharomyces cerevisiae. FEMS Yeast Res. 2003, 3: 167-175. 10.1016/S1567-1356(02)00186-1.
Hamacher T, Becker J, Gardonyi M, Hahn-Hägerdal B, Boles E: Characterization of the xylose-transporting properties of yeast hexose transporters and their influence on xylose utilization. Microbiology. 2002, 148: 2783-2788.
Kou SC, Christensen MS, Cirillo VP: Galactose transport in Saccharomyces cerevisiae. II. Characteristics of galactose uptake and exchange in galactokinaseless cells. J Bacteriol. 1970, 103: 671-678.
Leandro MJ, Goncalves P, Spencer-Martins I: Two glucose/xylose transporter genes from the yeast Candida intermedia: first molecular characterization of a yeast xylose/H + symporter. Biochem J. 2006
Görgens JF, van Zyl WH, Knoetze JH, Hahn-Hägerdal B: The metabolic burden of the PGK1 and ADH2 promoter systems for heterologous xylanase production by Saccharomyces cerevisiae in defined medium. Biotechnol Bioeng. 2001, 73: 238-245. 10.1002/bit.1056.
Sambrook J, Fritch E, Maniatis T: Molecular cloning: a laboratory manual. 2nd ed. 1989, Cold Spring Harbor, NY, Cold Spring Harbor Laboratory Press
Jeppsson M, Bengtsson O, Franke K, Lee H, Hahn-Hägerdal B, Gorwa-Grauslund MF: The expression of a Pichia stipitis xylose reductase mutant with higher K(M) for NADPH increases ethanol production from xylose in recombinant Saccharomyces cerevisiae. Biotechnol Bioeng. 2006, 93: 665-673. 10.1002/bit.20737.
Dagert M, Ehrlich SD: Prolonged incubation in calcium chloride improves the competence of Escherichia coli cells. Gene. 1979, 6: 23-28. 10.1016/0378-1119(79)90082-9.
Gietz RD, Schiestl RH, Willems AR, Woods RA: Studies on the transformation of intact yeast cells by the LiAc/SS-DNA/PEG procedure. Yeast. 1995, 11: 355-360. 10.1002/yea.320110408.
Hauf J, Zimmermann FK, Muller S: Simultaneous genomic overexpression of seven glycolytic enzymes in the yeast Saccharomyces cerevisiae. Enzyme Microb Technol. 2000, 26: 688-698. 10.1016/S0141-0229(00)00160-5.
Guldener U, Heck S, Fielder T, Beinhauer J, Hegemann JH: A new efficient gene disruption cassette for repeated use in budding yeast. Nucleic Acids Res. 1996, 24: 2519-2524. 10.1093/nar/24.13.2519.
Skryabin KG, Eldarov MA, Larionov VL, Bayev AA, Klootwijk J, de Regt VC, Veldman GM, Planta RJ, Georgiev OI, Hadjiolov AA: Structure and function of the nontranscribed spacer regions of yeast rDNA. Nucleic Acids Res. 1984, 12: 2955-2968.
Smiley KL, Bolen PL: Demonstration of D-xylose reductase and D-xylitol dehydrogenase in Pachysolen tannophilus. Biotech Lett. 1982, 4: 607-610. 10.1007/BF00127793.
Rizzi M, Harwart K, Erlemann P, Bui-Thahn NA, Dellweg H: Purification and properties of the NAD+-xylitol-dehydrogenase from the yeast Pichia stipitis. J Ferment Bioeng. 1989, 67: 20-24. 10.1016/0922-338X(89)90080-9.
Acknowledgements
We thank Valeria Wallace Salinas for assistance in cloning. KK was financed by the Swedish Energy Agency. BW was partly financed by a Marie-Curie Fellowship grant (QCIM, QLK3-CT-2001-60077).
Author information
Authors and Affiliations
Corresponding author
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Karhumaa, K., Wiedemann, B., Hahn-Hägerdal, B. et al. Co-utilization of L-arabinose and D-xylose by laboratory and industrial Saccharomyces cerevisiae strains. Microb Cell Fact 5, 18 (2006). https://doi.org/10.1186/1475-2859-5-18
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1475-2859-5-18