
Abstract
Background
TP53 is one of major tumour suppressor genes being essential in preservation of genome integrity. Two very common polymorphisms have been demonstrated to contribute to cancer susceptibility and tumour behaviour. The purpose of this study was to evaluate the role of Arg72Pro and PIN3 Ins16bp polymorphisms in TP53 gene as genetic susceptibility and predictive markers to breast cancer.
Methods
We analysed DNA samples from 264 breast cancer patients and 440 controls, for TP53 Arg72Pro and PIN3 Ins16bp polymorphisms using PCR-RFLP.
Results
We observed that women with A2A2 genotype have increased risk for developing breast cancer, either in women with or without familial history (FH) of the disease (OR = 4.40, 95% CI 1.60–12.0; p = 0.004; OR = 3.88, 95% CI 1.18–12.8; p = 0.026, respectively). In haplotype analysis, statistically significant differences were found between TP53 Arg-A2 haplotype frequencies and familial breast cancer cases and the respective control group (OR = 2.10, 95% CI 1.08–4.06; p = 0.028). Furthermore, both TP53 polymorphisms are associated with higher incidence of lymph node metastases.
Conclusion
Our findings suggest TP53 PIN3 Ins16bp polymorphism as a real risk modifier in breast cancer disease, either in sporadic and familial breast cancer. Furthermore, both TP53 polymorphisms are associated with higher incidence of lymph node metastases.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Background
Breast cancer have been associated with well-established risk factors, such as high estrogen exposure, environmental factors (e. g. diet and ionizing radiation) and family history [1, 2]. Family history of breast cancer is a particularly important high risk factor for this disease. Two genes were identified as the major susceptibility genes in high risk families, namely BRCA1 and BRCA2. However, these genes account for only a minority of the overall family risk of breast cancer [3]. Furthermore, approximately only 10% of all breast cancer cases exhibit a familial pattern of incidence [4, 5]. In this way, the remaining familial and sporadic risk may be due to common low to moderate penetrance genetic variants, which are also referred as genetic polymorphisms. One strong candidate for genetic susceptibility factor to familial and/or sporadic breast cancer is the TP53 gene. This gene is frequently somatically mutated in breast cancer [6, 7] and TP53 germline mutations are associated with increased risk for developing diverse malignancies, including 25–30% of hereditary breast cancer cases associated with Li-Fraumeni syndrome [8]. Furthermore, based on its pivotal role in DNA damage repair and its physical and functional interactions with BRCA1 and BRCA2 proteins [9, 10], TP53 seems to be a strong candidate breast cancer predisposition.
The TP53 tumour suppressor gene, also designated the guardian of the genome, is essential in preservation of genome integrity. From the numerous biological functions of p53 protein, inhibition of cell cycle progression, DNA repair and apoptosis are the major cellular pathways where it is involved [6].
TP53 gene mutations are widely detected in breast cancer, being correlated with specific clinical phenotypes [11, 12].
Predisposition to several human cancers has been associated with genetic polymorphisms, which may represent an important contribution to cancer susceptibility and tumour behaviour [13–16]. Several polymorphisms have been identified within TP53 gene, both in non-coding and coding regions [17]. One of the most well studied TP53 gene polymorphism is Arg72Pro, located in codon 72 on exon 4, leading to arginine-proline substitution, which in its turn results in a structural alteration of the protein [18]. Another common polymorphism is 16 base pair (bp) duplication in intron 3 of the TP53 gene (PIN3 Ins16bp).
In this case-control study, we hypothesize that the two common polymorphisms of TP53 gene play a role either apoptosis, cell cycle control efficiency, as well as DNA repair capacity, which ultimately may contribute to an increase of breast cancer susceptibility within familial and/or sporadic cases, as well as represent an additional tool for prognosis prediction.
Methods
Study Population
We analysed a total of 264 DNA breast cancer cases: 73 unrelated familial breast cancer cases were selected from the Oncology and Surgical Departments from S. João Hospital at Porto and Vigo Hospital, and 191 unrelated sporadic breast cancer cases were recruited from IPO-Porto (Oncology Portuguese Institute), during 1998–2003, from patients that were receiving treatment. All cases were histological confirmed at the Department of Pathology. Clinico-pathological parameters were obtained when possible from hospital clinical records. Familial case group presented a mean age of 42.07 years, with an age range of 24–77 years. The high-risk familial breast cancer group, also designated by us as family history (FH) breast cancer cases, included women with the follow features, based on the Breast Cancer Linkage Consortium criteria [19]: early onset (≤40 years) and/or bilaterality; or more than three cases of breast cancer in the family; or more than one case of ovarian cancer in the family; or more than two first-degree relatives involved; or male breast cancer. These high-risk breast cancer cases are BRCA1/BRCA2 mutations non-carriers [20]. Sporadic cases group (with no presence of FH) presented a mean age of 53.41 years, with an age range of 41–88 years. Control women were randomly selected from blood banks in the same region during the same time period as the cases were collected. The selection criteria include no prior history of cancer, and controls were frequency matched to the cases by age (± 5 years). A total of 440 healthy women presenting a median age of 42.29 years and an age range of 21–85 years, were used as control group of familial breast cancer cases. From the above control group 216 healthy women were selected according to the age of diagnosis of the sporadic breast cancer patients group (higher than 40 years), with a median age of 53.05 and an age range of 41–85, being used as control group of sporadic breast cancer cases. All participants provided informed consent. Ethical approval was obtained by the specific institutions ethical committees.
Laboratory Methods
Genomic DNA was isolated from lymphocytes of peripherical blood using Puregene® DNA Purification Kit (Gentra Systems, Minneapolis, USA). All the polymorphisms were assessed by PCR-RFLP technique. TP53 Arg72Pro polymorphism (rs1042522) was detected by amplifying genomic DNA with the forward primer 5'-GAA GAC CCA GGT CCA GAT GA-3' and the reverse primer 5'-CTG CCC TGG TAG GTT TTC TG-3'. The PCR amplification parameters were 32 cycles each of 30 sec at 94°C, 30 sec at 54°C, and 30 sec at 72°C. The 152 bp PCR product was digested with Bsh1236I (Fermentas, Ontario, Canada) at 37°C overnight. Digested products were separated by electrophoresis in a 3% agarose gel (Seakem® LE Agarose, Rockland, USA) and visualized by ethidium bromide staining. Wild type alleles resulted in 50 and 102 bp fragments and the variant alleles resulted in 152 bp fragment following restriction enzyme digestion. TP53 PIN3 Ins16bp polymorphism (rs17878362) was detected by amplifying genomic DNA with the forward primer 5'-CTG AAA ACA ACG TTC TGG TA-3' and the reverse primer 5'-AAG GGG GAC TGT AGA TGG GTG-3'. The PCR amplification parameters were 32 cycles each of 30 sec at 94°C, 30 sec at 60°C, and 30 sec at 72°C. The PCR product was separated by electrophoresis in a 4% agarose gel (Seakem® LE Agarose, Rockland, USA) and visualized by ethidium bromide staining. Wild type alleles, designated A1 allele (no duplication) resulted in 119 bp fragment and the variant alleles, designated A2 allele (with 16 bp duplication) resulted in 135 bp fragment.
To ensure quality control of all genotyping results, 10% of the samples was randomly selected and sequenced using an ABI automated sequencer. We obtained a 98% of concordance between the genotyping results acquire by the two methods.
Statistical Analysis
Analysis of data was performed using the computer software SPSS version 14.0 (SPSS Inc., Chicago, USA). Chi-square (χ2 test) analysis was used to compare categorical variables. Whenever necessary, the Fisher test was used when number of samples was equal or inferior to 5. A 5% level of significance was used in the analysis. The Odds Ratio (OR) and its 95% confidence interval (CI) were calculated to measure the association between TP53 polymorphic genotypes and breast cancer risk. Logistic regression analysis was used to calculate the adjusted OR and 95% CI for the influence of TP53 genotypes in the risk of breast cancer, adjusted for age and/or FH. Whenever appropriate, the observed number of each genotype in control groups were compared with that expected for a population in the Hardy-Weinberg Equilibrium by using a goodness of fit χ2 test. The frequencies of expected haplotypes were estimated by using the statistical methodologies implemented by HPlus software [21], which infers haplotypes based on expectation-maximization with a modified progressive ligation computational algorithm. The nonparametric test, Kruskal-Wallis test, was used to compare mean age of diagnosis between the different genotype polymorphisms.
Results
The distribution of the genotype frequencies in PIN3 Ins16bp polymorphisms among control group (p = 0.478) and in Arg72Pro and PIN3 Ins16bp among control subgroup (p = 0.082 and p = 0.294) is in agreement with those expected under Hardy-Weinberg equilibrium, excepted for Arg72Pro in the overall control group (p = 0.013).
Concerning TP53 Arg72Pro polymorphism in the familial breast cancer cases, frequencies of Arg72Arg, Arg72Pro and Pro72Pro were 53.4%, 34.2% and 12.3%, respectively. In sporadic breast cancer, 56.0%, 34.9% and 9.1% were homozygous for 72Arg allele, heterozygous and homozygous for 72Pro allele, respectively. No statistically significant associations were found between the TP53 Arg72Pro polymorphism and risk of familial and sporadic breast cancer risk (Table 1).
Frequencies of TP53 PIN3 Ins16bp polymorphism genotypes were 65.7% to A1A1, 21.4% to A1A2 and 12.9% to A2A2, in familial breast cancer cases. Regarding sporadic breast cancer group, we observed 63.9%, 29.3% and 6.8% frequencies for homozygous for A1 allele, heterozygous and homozygous for A2 allele, respectively. We observed that A2A2 genotype carriers with positive FH were at a 4.40-fold (95% CI = 1.60–12.0; p = 0.004) increased risk of breast cancer compared with the respective control group. Moreover, statistically significant differences were observed in A2A2 genotype frequencies comparing sporadic breast cancer cases and respective control group (p = 0.026). Our results showed that carriers of A2A2 genotype with no FH present an increased risk of breast cancer (OR = 3.88, 95% CI 1.18–12.8).
We investigated haplotype effects of the two polymorphisms studied in breast cancer risk (Table 2). Compared the common TP53 Arg-A1 haplotype with the other expected haplotypes; we only observed statistically significant differences regarding TP53 Arg-A2 haplotype between the familial breast cancer cases and respective control group (p = 0.028). Carriers of TP53 Arg-A2 haplotype and presence of FH of breast cancer presented an increased risk of develop breast cancer (OR= 2.10; 95% CI 1.08–4.06).
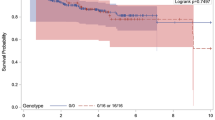
We examined the relationship between age at onset and genotypes and found a positive correlation in the FH group. The mean age of FH patients group with A2A2 genotype was 33.43 (± 8.08) years, whereas the mean age of patients with A1A1 and A1A2 genotypes was 42.44 (± 12.14) and 44.80 (± 10.85) years, respectively (Kruskal Wallis test p = 0.056). Therefore, the carrier's status of A2A2 genotype was associated with an earlier age at onset cancer with respect to the patients with A1 genotypes. However, this difference was in the frontier of statistically significant, possibly because of the smaller size of the group (7 patients to Pro/Pro genotype). No association was observed relating age at onset and Arg72Pro polymorphism (Kruskal Wallis test p = 0.747).
The analysis of the TP53 polymorphisms with respect to some clinical pathological showed a significant association of Pro or A2 genotypes with the presence of lymph node metastases (p = 0.009 and p < 0.001, respectively, adjusted for age and breast cancer family history, using logistic regression analysis).
Discussion
Breast cancer is an heterogeneous disease, as sustained by wide variable morphological appearance, many risks factors and distinct gene expression profile [2, 22]. Common genetic alterations (e.g. polymorphisms), with possible effects on function and/or protein expression, within genes involved in essential cellular pathways, such as carcinogen metabolism, DNA repair, cell cycle control and cell proliferation, could predispose individuals to cancer [15, 23–25], including breast cancer [15, 26–29].
The TP53 is one of the major tumour suppressor genes which carry out essential functions in preservation of genome integrity. Thus, when the cell is under stress, particularly stress which will involve DNA damage, p53 promotes growth arrest, allowing the cell to repair the DNA lesions. If the damage is excessively hazardous, then p53 will lead to cell apoptosis. Several genetic polymorphisms have been described in TP53 gene [18] and some of these variants seem to confer different functions among the p53 [30–32].
In the present study, we evaluated two separate TP53 polymorphisms, Arg72Pro and PIN3 Ins16bp, in two groups of breast cancer, familial and sporadic cases, as well as in matching control groups. The allelic frequencies of our control group for the different polymorphisms are in accordance with earlier reports from European populations [16, 33]. We found a deviation from Hardy-Weinberg equilibrium in overall group of controls for Arg72Pro P53 polymorphism genotypes. Hardy-Weinberg equilibrium depends on a series of features about the tested population, including, for example the sample population size, random mating, no migration, no genetic drift and no selection taking place [34]. Thus, this deviation could be due to chance or violation of these assumptions, being the possibility of genotyping errors lower, since 10% our control sample genotyping were confirmed by direct sequence and its were similar with other European populations.
Concerning the codon 72 TP53 polymorphism (Arg72Pro), we did not find any association between this polymorphism and breast cancer. Our results are in agreement with other studies [33, 35, 36], however, the literature remains highly controversial regarding the role of this polymorphism in breast cancer risk [37–42]. One study showed that TP53 72Pro variant induces transcription activation more efficiently than TP53 72Arg variant [40]. On the other hand, other authors revealed that TP53 72Pro variant induce cell cycle arrest better than 72Arg [31]. Other studies have showed that TP53 72Arg variant is more efficient in inducing apoptosis [32, 42]. Beside apoptosis and cell cycle control, p53 protein seems to be crucial in the regulation of the different DNA repair pathways [43]. A recent study demonstrated the influence of TP53 Arg72Pro in DNA repair capacity, showing that TP53 72Pro variant activates several TP53 dependent target genes involved in DNA repair and DNA damage repair much more efficiently than the 72Arg variant expressing cells [30]. These contradictory results could be explained by the differential effects of this alteration in p53 function. Several in vitro evidences have demonstrated that both TP53 Arg72Pro variants may selectively regulate specific cellular functions.
In TP53 PIN3 polymorphism, our findings suggest an association of A2A2 genotype and increased breast cancer risk among women with FH and sporadic breast cancer, suggesting that this polymorphism contributed to enhance susceptibility for breast cancer among Portuguese population, regardless of the presence of FH. Our results are supported by previously reported studies suggesting an association of PIN3 A2 genotypes with breast cancer risk [44]. Although, the biological effect of the TP53 PIN3 Ins16bp polymorphism is currently unclear, theoretically, this polymorphism could affect mRNA splicing, altering the coding regions and therefore being implicated in regulation of gene expression and DNA-protein interactions, resulting in a defective protein [45, 46]. Until now, just a single study had show PIN3 A2 allele presents reduced mRNA stability [47].
The linkage disequilibrium between TP53 polymorphisms region could be an important factor affecting the incidence of cancer in general [48, 49], and breast cancer, in particular [42, 44, 50]. Thus, haplotype analysis would be important to confirm the significance of this variant on breast cancer susceptibility. A statistical significant association was found between Arg-A2 haplotype and breast cancer susceptibility among women with FH of breast cancer. On the other hand, a recent study has found that Pro-A1 haplotype individuals present increased breast cancer risk, however, in BRCA2 mutation carriers [42]. Nevertheless, other reports have also demonstrated a positive association of Arg-A2 haplotype with cancer [44, 48]. Moreover, functional studies have shown that, in a specific haplotype combination, A2 allele is associated with decreased apoptotic and DNA repair capacity [33, 48].
Our findings suggest the Pro/Pro and A2A2 TP53 genotypes as predictor factors for the presence of lymph node metastases, being in agreement with previously functional studies in the biological consequences of these variations in P53 protein functions [40, 47].
The natural history of breast cancer can be influenced by several factors. We hypothesize that under the influence of TP53 genetic polymorphisms, chronic exposure to higher levels of several endogenous (e.g. estrogens) and exogenous breast carcinogens resulting in consequent higher accumulation of DNA damage during an individual's lifetime, may alter the age at onset of disease. Moreover, it has been suggested that TP53 polymorphisms are associated to familial breast cancer by the age of 50 years [33]. Our results are consistent with this hypothesis, since TP53 PIN3 Ins16bp polymorphism seems to influence directly the age to onset of familial breast cancer.
Conclusion
In conclusion, our findings suggest TP53 PIN3 Ins16bp polymorphism as a real risk modifier in breast cancer disease, either in sporadic and familial breast cancer. Subsequently, these results will be crucial in the characterization of the genetic breast cancer susceptibility profile, within familial breast cancer cases non-carriers of BRCA1/BRCA2 mutations. Furthermore, our findings suggest TP53 Arg72Pro and PIN3 Ins16bp polymorphisms as predictive factors of presence of lymph node metastases.
Abbreviations
- FH:
-
Family history
- PCR:
-
Polymerase chain reaction
- RFLP:
-
Restriction fragment length polymorphism
- OR:
-
Odds ratio
- CI:
-
Confidence interval
- OS:
-
Estimate overall survival.
References
DeBruin LS, Josephy PD: Perspectives on the chemical etiology of breast cancer. Environ Health Perspect. 2002, 110 Suppl 1: 119-128.
Dumitrescu RG, Cotarla I: Understanding breast cancer risk -- where do we stand in 2005?. J Cell Mol Med. 2005, 9: 208-221. 10.1111/j.1582-4934.2005.tb00350.x.
Dapic V, Carvalho MA, Monteiro AN: Breast cancer susceptibility and the DNA damage response. Cancer Control. 2005, 12: 127-136.
Ford D, Easton DF, Stratton M, Narod S, Goldgar D, Devilee P, Bishop DT, Weber B, Lenoir G, Chang-Claude J, Sobol H, Teare MD, Struewing J, Arason A, Scherneck S, Peto J, Rebbeck TR, Tonin P, Neuhausen S, Barkardottir R, Eyfjord J, Lynch H, Ponder BA, Gayther SA, Zelada-Hedman M, .: Genetic heterogeneity and penetrance analysis of the BRCA1 and BRCA2 genes in breast cancer families. The Breast Cancer Linkage Consortium. Am J Hum Genet. 1998, 62: 676-689. 10.1086/301749.
Lichtenstein P, Holm NV, Verkasalo PK, Iliadou A, Kaprio J, Koskenvuo M, Pukkala E, Skytthe A, Hemminki K: Environmental and heritable factors in the causation of cancer--analyses of cohorts of twins from Sweden, Denmark, and Finland. N Engl J Med. 2000, 343: 78-85. 10.1056/NEJM200007133430201.
Feki A, Irminger-Finger I: Mutational spectrum of p53 mutations in primary breast and ovarian tumors. Crit Rev Oncol Hematol. 2004, 52: 103-116. 10.1016/j.critrevonc.2004.07.002.
Gasco M, Yulug IG, Crook T: TP53 mutations in familial breast cancer: functional aspects. Hum Mutat. 2003, 21: 301-306. 10.1002/humu.10173.
Olivier M, Goldgar DE, Sodha N, Ohgaki H, Kleihues P, Hainaut P, Eeles RA: Li-Fraumeni and related syndromes: correlation between tumor type, family structure, and TP53 genotype. Cancer Res. 2003, 63: 6643-6650.
Jonkers J, Meuwissen R, van der GH, Peterse H, van , Berns A: Synergistic tumor suppressor activity of BRCA2 and p53 in a conditional mouse model for breast cancer. Nat Genet. 2001, 29: 418-425. 10.1038/ng747.
Ongusaha PP, Ouchi T, Kim KT, Nytko E, Kwak JC, Duda RB, Deng CX, Lee SW: BRCA1 shifts p53-mediated cellular outcomes towards irreversible growth arrest. Oncogene. 2003, 22: 3749-3758. 10.1038/sj.onc.1206439.
Xu Y, Yao L, Ouyang T, Li J, Wang T, Fan Z, Lin B, Lu Y, Xie Y: p53 Codon 72 polymorphism predicts the pathologic response to neoadjuvant chemotherapy in patients with breast cancer. Clin Cancer Res. 2005, 11: 7328-7333. 10.1158/1078-0432.CCR-05-0507.
Tommiska J, Eerola H, Heinonen M, Salonen L, Kaare M, Tallila J, Ristimaki A, von SK, Aittomaki K, Heikkila P, Blomqvist C, Nevanlinna H: Breast cancer patients with p53 Pro72 homozygous genotype have a poorer survival. Clin Cancer Res. 2005, 11: 5098-5103. 10.1158/1078-0432.CCR-05-0173.
Medeiros R, Vasconcelos A, Costa S, Pinto D, Lobo F, Morais A, Oliveira J, Lopes C: Linkage of angiotensin I-converting enzyme gene insertion/deletion polymorphism to the progression of human prostate cancer. J Pathol. 2004, 202: 330-335. 10.1002/path.1529.
Medeiros R, Vasconcelos A, Costa S, Pinto D, Morais A, Oliveira J, Lopees C: Steroid hormone genotypes ARStuI and ER325 are linked to the progression of human prostate cancer. Cancer Genet Cytogenet. 2003, 141: 91-96. 10.1016/S0165-4608(02)00660-X.
Pinto D, Vasconcelos A, Costa S, Pereira D, Rodrigues H, Lopes C, Medeiros R: HER2 polymorphism and breast cancer risk in Portugal. Eur J Cancer Prev. 2004, 13: 177-181. 10.1097/01.cej.0000130015.91525.c7.
Santos AM, Sousa H, Pinto D, Portela C, Pereira D, Catarino R, Duarte I, Lopes C, Medeiros R: Linking TP53 codon 72 and P21 nt590 genotypes to the development of cervical and ovarian cancer. Eur J Cancer. 2006, 42: 958-963. 10.1016/j.ejca.2006.01.015.
Olivier M, Eeles R, Hollstein M, Khan MA, Harris CC, Hainaut P: The IARC TP53 database: new online mutation analysis and recommendations to users. Hum Mutat. 2002, 19: 607-614. 10.1002/humu.10081.
Pietsch EC, Humbey O, Murphy ME: Polymorphisms in the p53 pathway. Oncogene. 2006, 25: 1602-1611. 10.1038/sj.onc.1209367.
Pathology of familial breast cancer: differences between breast cancers in carriers of BRCA1 or BRCA2 mutations and sporadic cases. Breast Cancer Linkage Consortium. Lancet. 1997, 349: 1505-1510. 10.1016/S0140-6736(96)10109-4.
Duarte F, Cameselle-Teijeiro JF, Soares R, Seixas C, Cortizo-Torres ME, Perez-Villanueva J, Schmitt FC: [Analysis of mutations in genes BRCA1 and BRCA2 among patients with breast and ovarian cancer in northern Portugal and Galicia]. Rev Clin Esp. 2002, 202: 259-263.
Zhao LP, Li LL, Khalid N: A method for assessing disease associations with SNP haplotypes and environmental variables in case-control. Am J Hum Genet. 2003, 72: 1231-1250. 10.1086/375140.
Perou CM, Sorlie T, Eisen MB, van de RM, Jeffrey SS, Rees CA, Pollack JR, Ross DT, Johnsen H, Akslen LA, Fluge O, Pergamenschikov A, Williams C, Zhu SX, Lonning PE, Borresen-Dale AL, Brown PO, Botstein D: Molecular portraits of human breast tumours. Nature. 2000, 406: 747-752. 10.1038/35021093.
Medeiros R, Vasconcelos A, Costa S, Pinto D, Ferreira P, Lobo F, Morais A, Oliveira J, Lopes C: Metabolic susceptibility genes and prostate cancer risk in a southern European population: the role of glutathione S-transferases GSTM1, GSTM3, and GSTT1 genetic polymorphisms. Prostate. 2004, 58: 414-420. 10.1002/pros.10348.
Costa S, Pinto D, Morais A, Vasconcelos A, Oliveira J, Lopes C, Medeiros R: Acetylation genotype and the genetic susceptibility to prostate cancer in a southern European population. Prostate. 2005, 64: 246-252. 10.1002/pros.20241.
Costa S, Pinto D, Pereira D, Vasconcelos A, fonso-Lopes C, Osorio T, Lopes C, Medeiros R: Importance of xeroderma pigmentosum group D polymorphisms in susceptibility to ovarian cancer. Cancer Lett. 2006, 246: 324-330. 10.1016/j.canlet.2006.03.014.
Gaudet MM, Bensen JT, Schroeder J, Olshan AF, Terry MB, Eng SM, Teitelbaum SL, Britton JA, Lehman TA, Neugut AI, Ambrosone CB, Santella RM, Gammon MD: Catechol-O-methyltransferase haplotypes and breast cancer among women on Long Island, New York. Breast Cancer Res Treat. 2006, 99: 235-240. 10.1007/s10549-006-9205-0.
Shin A, Shu XO, Cai Q, Gao YT, Zheng W: Genetic polymorphisms of the transforming growth factor-beta1 gene and breast cancer risk: a possible dual role at different cancer stages. Cancer Epidemiol Biomarkers Prev. 2005, 14: 1567-1570. 10.1158/1055-9965.EPI-05-0078.
Dufloth RM, Costa S, Schmitt F, Zeferino LC: DNA repair gene polymorphisms and susceptibility to familial breast cancer in a group of patients from Campinas, Brazil. Genet Mol Res. 2005, 4: 771-782.
Lee KM, Choi JY, Park SK, Chung HW, Ahn B, Yoo KY, Han W, Noh DY, Ahn SH, Kim H, Wei Q, Kang D: Genetic polymorphisms of ataxia telangiectasia mutated and breast cancer risk. Cancer Epidemiol Biomarkers Prev. 2005, 14: 821-825. 10.1158/1055-9965.EPI-04-0330.
Siddique M, Sabapathy K: Trp53-dependent DNA-repair is affected by the codon 72 polymorphism. Oncogene. 2006, 25: 3489-3500. 10.1038/sj.onc.1209405.
Pim D, Banks L: p53 polymorphic variants at codon 72 exert different effects on cell cycle progression. Int J Cancer. 2004, 108: 196-199. 10.1002/ijc.11548.
Dumont P, Leu JI, Della PAC, George DL, Murphy M: The codon 72 polymorphic variants of p53 have markedly different apoptotic potential. Nat Genet. 2003, 33: 357-365. 10.1038/ng1093.
Wang-Gohrke S, Becher H, Kreienberg R, Runnebaum IB, Chang-Claude J: Intron 3 16 bp duplication polymorphism of p53 is associated with an increased risk for breast cancer by the age of 50 years. Pharmacogenetics. 2002, 12: 269-272. 10.1097/00008571-200204000-00012.
Lewis R: Human Genetics. 2001, New York, McGraw-Hill, 12: 228-232. 4
Mabrouk I, Baccouche S, El-Abed R, Mokdad-Gargouri R, Mosbah A, Said S, Daoud J, Frikha M, Jlidi R, Gargouri A: No evidence of correlation between p53 codon 72 polymorphism and risk of bladder or breast carcinoma in Tunisian patients. Ann N Y Acad Sci. 2003, 1010: 764-770. 10.1196/annals.1299.137.
Suspitsin EN, Buslov KG, Grigoriev MY, Ishutkina JG, Ulibina JM, Gorodinskaya VM, Pozharisski KM, Berstein LM, Hanson KP, Togo AV, Imyanitov EN: Evidence against involvement of p53 polymorphism in breast cancer predisposition. Int J Cancer. 2003, 103: 431-433. 10.1002/ijc.10834.
Bonafe M, Ceccarelli C, Farabegoli F, Santini D, Taffurelli M, Barbi C, Marzi E, Trapassi C, Storci G, Olivieri F, Franceschi C: Retention of the p53 Codon 72 Arginine Allele Is Associated with a Reduction of Disease-Free and Overall Survival in Arginine/Proline Heterozygous Breast Cancer Patients. Clin Cancer Res. 2003, 9: 4860-4864.
Ohayon T, Gershoni-Baruch R, Papa MZ, Distelman MT, Eisenberg BS, Friedman E: The R72P P53 mutation is associated with familial breast cancer in Jewish women. Br J Cancer. 2005, 92: 1144-1148. 10.1038/sj.bjc.6602451.
Buyru N, Tigli H, Dalay N: P53 codon 72 polymorphism in breast cancer. Oncol Rep. 2003, 10: 711-714.
Thomas M, Kalita A, Labrecque S, Pim D, Banks L, Matlashewski G: Two polymorphic variants of wild-type p53 differ biochemically and biologically. Mol Cell Biol. 1999, 19: 1092-1100.
Martin AM, Kanetsky PA, Amirimani B, Colligon TA, Athanasiadis G, Shih HA, Gerrero MR, Calzone K, Rebbeck TR, Weber BL: Germline TP53 mutations in breast cancer families with multiple primary cancers: is TP53 a modifier of BRCA1?. J Med Genet. 2003, 40: e34-10.1136/jmg.40.4.e34.
Osorio A, Martinez-Delgado B, Pollan M, Cuadros M, Urioste M, Torrenteras C, Melchor L, Diez O, de la HM, Velasco E, Gonzalez-Sarmiento R, Caldes T, Alonso C, Benitez J: A haplotype containing the p53 polymorphisms Ins16bp and Arg72Pro modifies cancer risk in BRCA2 mutation carriers. Hum Mutat. 2006, 27: 242-248. 10.1002/humu.20283.
Sengupta S, Harris CC: p53: traffic cop at the crossroads of DNA repair and recombination. Nat Rev Mol Cell Biol. 2005, 6: 44-55. 10.1038/nrm1546.
Weston A, Pan CF, Ksieski HB, Wallenstein S, Berkowitz GS, Tartter PI, Bleiweiss IJ, Brower ST, Senie RT, Wolff MS: p53 haplotype determination in breast cancer. Cancer Epidemiol Biomarkers Prev. 1997, 6: 105-112.
Mattick JS: Introns: evolution and function. Curr Opin Genet Dev. 1994, 4: 823-831. 10.1016/0959-437X(94)90066-3.
Mattick JS: Rna Regulation: a new genetics?. Nat Rev Genet. 2004, 5: 316-10.1038/nrg1321.
Gemignani F, Moreno V, Landi S, Moullan N, Chabrier A, Gutierrez-Enriquez S, Hall J, Guino E, Peinado MA, Capella G, Canzian F: A TP53 polymorphism is associated with increased risk of colorectal cancer and with reduced levels of TP53 mRNA. Oncogene. 2004, 23: 1954-1956. 10.1038/sj.onc.1207305.
Wu X, Zhao H, Amos CI, Shete S, Makan N, Hong WK, Kadlubar FF, Spitz MR: p53 Genotypes and Haplotypes Associated With Lung Cancer Susceptibility and Ethnicity. J Natl Cancer Inst. 2002, 94: 681-690.
Birgander R, Sjalander A, Rannug A, Alexandrie AK, Sundberg MI, Seidegard J, Tornling G, Beckman G, Beckman L: P53 polymorphisms and haplotypes in lung cancer. Carcinogenesis. 1995, 16: 2233-2236. 10.1093/carcin/16.9.2233.
Sjalander A, Birgander R, Hallmans G, Cajander S, Lenner P, Athlin L, Beckman G, Beckman L: p53 polymorphisms and haplotypes in breast cancer. Carcinogenesis. 1996, 17: 1313-1316. 10.1093/carcin/17.6.1313.
Pre-publication history
The pre-publication history for this paper can be accessed here:http://www.biomedcentral.com/1471-2407/8/32/prepub
Acknowledgements
The authors would like to thank to Oncology and Surgical Department from S. João Hospital at Porto, and the Pathology Departments of the different Hospitals involved. We would like to thank to Imunohemotherapy Department of S. Marcos Hospital at Braga and to Clínica Laboratorial Dr. Edgar Botelho Moniz for their helpful assistance in the management of controls. We gratefully acknowledge FLAD (Fundação Luso-Americana para o Desenvolvimento), the Minister of Health of Portugal (Comissão de Fomento da Investigacão em Cuidados de Saúde: CFICS- 226/01), Astra Zeneca Foundation and Calouste Gulbenkian Foundation by financial funding of this work. We also gratefully acknowledge the Minister of Science and Superior Education-FCT (Fundação para a Ciência e Tecnologia: SFRH/BD/9758/2003) by financial support of individual grant for Doctoral degree of the first author.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The author(s) declare that they have no competing interests.
Authors' contributions
The SC was the principal investigator; contribute to data and samples collection, developed study design, experimental plan and implementation, statistical analysis, and drafted the manuscript. DP contributed to data and samples collection and critical revision of the manuscript. DP, HR and JCT were critical to data and samples collection. RM contributed to study design, samples collection, data analysis and critical review of the manuscript. FS contributed to conception of the study hypotheses, study design and critical review of the manuscript. In addition, all the authors read and approved the final submitted manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Costa, S., Pinto, D., Pereira, D. et al. Importance of TP53 codon 72 and intron 3 duplication 16bppolymorphisms in prediction of susceptibility on breast cancer. BMC Cancer 8, 32 (2008). https://doi.org/10.1186/1471-2407-8-32
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1471-2407-8-32