
Abstract
Background
Antimicrobial peptides (AMP) are important elements of the first line of defence against pathogens in animals. NK-lysin is a cationic AMP that plays a critical role in innate immunity. The chicken NK-lysin gene has been cloned and its antimicrobial and anticancer activity has been described but its location in the chicken genome remains unknown. Here, we mapped the NK-lysin gene and examined the distribution of a functionally significant single nucleotide polymorphism (SNP) among different chicken inbred lines and heritage breeds.
Results
A 6000 rad radiation hybrid panel (ChickRH6) was used to map the NK-lysin gene to the distal end of chromosome 22. Two additional genes, the adipocyte enhancer-binding protein 1-lik e gene (AEBP1) and the DNA polymerase delta subunit 2-like (POLD2) gene, are located in the same NW_003779909 contig as NK-lysin, and were thus indirectly mapped to chromosome 22 as well. Previously, we reported a functionally significant SNP at position 271 of the NK-lysin coding sequence in two different chicken breeds. Here, we examined this SNP and found that the A allele appears to be more common than the G allele in these heritage breeds and inbred lines.
Conclusions
The chicken NK-lysin gene mapped to the distal end of chromosome 22. Two additional genes, AEBP1 and POLD2, were indirectly mapped to chromosome 22 also. SNP analyses revealed that the A allele, which encodes a peptide with a higher antimicrobial activity, is more common than the G allele in our tested inbred lines and heritage breeds.
Similar content being viewed by others


Background
The chicken is an important animal for several reasons. In addition to being a major source of protein in the world, it is valuable to the understanding of genome evolution because of its relationship to mammals. The chicken genome sequence assembly was completed in 2004 with a six-fold whole genome shotgun (Sanger) coverage. It was the first avian genome to be sequenced [1] and, therefore, holds a place in comparative genomics as a prototypic avian genome. Subsequent sequencing and mapping have improved upon that first build, and efforts continue in search of missing and/or unassembled sequences, primarily on the smaller microchromosomes and the sex chromosomes. Nevertheless, sequence segments remain misplaced in the genome assembly [2] or unmapped.
Radiation hybrid (RH) panels are useful mapping tools to determine the location and order of genes, and to aid the assembly of genome sequences. RH panels are available for several domestic animal species including cow [3], pig [4], horse [5], dog [6], cat [7], mouse [8], chicken [9] and duck [10]. The ChickRH6 radiation hybrid panel was produced in 2002 [9] and has been used to construct consensus chromosome RH maps of the chicken genome and a limited number of chicken or GGA (Gallus gallus) chromosome maps have been published to date, namely GGA2 [11], GGA4 [12], GGA5 [13], GGA7 [14], GGA14 [15], GGA15 [16], GGA16 [17] and GGA25 [18].
NK-lysin is a cationic peptide with antibacterial activity that was originally isolated from porcine intestinal tissue. Extensive research has been conducted on the structure and antimicrobial activities of NK-lysin isolated from different species [19–23]. The chicken NK-lysin gene was cloned in 2006 [24], and its antimicrobial activity against Eimeria sporozocites was reported [25]. Previously, we identified a single nucleotide polymorphism (SNP) in the chicken NK-lysin gene and discovered that the encoded protein variants are differentially cytotoxic for several bacteria and cell lines derived from human cancers [26]. However, to date the location of NK-lysin on the chicken genome remains unknown. Here, we used the ChickRH6 panel to map the NK-lysin gene on the chicken genome, and also examined its polymorphism and allele distribution among diverse heritage breeds and inbred lines of chicken.
Methods
Genotyping
Different heritage chicken breeds available publically and several experimental inbred lines were surveyed. The heritage chicken breed samples came as pooled blood from eight to 26 animals, each. Three to seven individual samples were used for each inbred line. DNA (50 ng) was used for PCR amplification with a forward primer of cNKL QF3 and reverse primer of cNKL QR2 (Table 1), and Sanger sequencing was carried out with the BigDye Termination kit, version 1 (ABI, Foster City, CA). To estimate allele frequencies from pooled blood samples, mixtures of known amounts of pure A and G allele DNA at ratios of 1:0, 3:1, 2:1, 1:1, 1:2 and 0:1 were prepared as a standard reference for quantitative PCR products. The peak area and heights were measured to approximate the ratio of alleles in pooled DNA from the heritage breed samples.
Radiation hybrid panel genotyping and map construction
The chicken NK-lysin gene was physically mapped using the INRA Chicken RH panel (ChickRH6) containing 90 hamster-chicken hybrid cell lines [9]. DNA from each line, along with control chicken and hamster DNA, were analysed for the presence or absence of NK-lysin and other markers by PCR (polymerase chain reaction) in 96-well microtiter plates. PCR was performed with the primers listed in Table 1. The PCR reactions were conducted with an initial denaturation step of 95°C for 5 min, followed by 35 cycles of 95°C for 30 s, 62°C for 30 s, 72°C for 30 s, and a final elongation step of 5 min at 72°C. Each marker was run at least twice on the RH panel to insure reproducibility. The amplified product was typed and scored as present (1), absent (0), or ambiguous (2) as previously described [9].
Chromosome assignment was done and mapping was performed by including our genotyping data in a larger dataset composed of 10 143 markers for the chicken genome. This dataset is composed of 2663 markers (genes and microsatellites) from the ChickRH database (http://chickrh.toulouse.inra.fr) and 7480 SNPs genotyped on the ChickRH panel using the Illumina GoldenGate assay at the Centre National de Génotypage (CNG), Evry, France. RH mapping was conducted using the Carthagene software [27]. We assumed random breakage along the chromosomes and equi-probable retention of fragments. RH map was constructed in three steps: (1) a two-point analysis identified markers linked together with a LOD score greater than 8 and defined RH groups from these data; (2) using all the markers from the linkage group corresponding to GGA22, multipoint analyses were done to build a framework map using a LOD threshold of 3; and (3) additional markers were added by calculating their location relative to the framework markers. Finally the map design was created using MapChart 2.0 [28].
Results
Single nucleotide polymorphism genotyping
Previously, we identified a SNP at nucleotide 271 of the NK-lysin coding sequence and discovered that the encoded protein variants have different cytotoxicities for bacteria and anticancer activity [26]. Here, we genotyped this SNP among 32 heritage breeds and 10 inbred lines. The DNA from the heritage breeds was obtained from pooled blood samples from eight to 26 animals. We compared the A and G peak heights based on the sequencing chromatogram to estimate allele frequency (Figure 1). A single A peak was detected in eight breeds and a single G peak was detected only in two of the 32 breeds (Table 2). Twenty-two of the 32 breeds analysed by pooled DNA revealed peaks for both A and G alleles. Eleven breeds showed A and G chromatogram peaks of similar height, indicating that the frequencies of the A and G alleles in the pooled DNA were approximately equal. Seven samples had a peak at least twice as high for allele A than for allele G. Only four of the 22 breeds carried G as the major allele. Thus, the A allele is more common than the G allele across all tested heritage breeds (Table 2).

Sequencing chromatogram of pooled DNA. DNA from homozygous chicken (A and G allele) was mixed in 1:0, 3:1, 2:1, 1:1, 1:2 and 0:1 A to G ratios and used as PCR template and sequenced; each chromatogram peak was compared to peaks from pooled DNA samples of heritage breeds and shows the estimated A to G ratio.
We also genotyped 10 inbred lines, eight from single animals and two from five pooled individuals. The White Leghorn line 6, Fayoumi M-5.1 and Fayoumi M-15.2 carried the G allele, while White Leghorn line 7, Leghorn Ghs-6, Leghorn Ghs-13, Spanish 21.1 and Leghorn line 8–15.1 carried the A allele. Two pooled inbred lines, line 0 and line 15Is, had the A allele (Table 3). There was no evidence of heterozygosity in any of the tested inbred lines. Thus, the A allele which has the higher antimicrobial activity was more common than the G allele across all tested heritage breeds and inbred lines.
Mapping of the chicken NK-lysin gene
In the Gallus_gallus-4.0 Primary Assembly, the unmapped contig NW_003779909 contains the chicken NK-lysin along with two additional genes, adipocyte enhancer-binding protein 1-like (AEBP1) and DNA polymerase delta subunit 2-like (POLD2). We used the 6000 rad ChickRH6 panel to physically map the NK-lysin gene in the chicken genome. A retention frequency (RF) of 27.7% for NK-lysin was observed based on 25 positive PCR bands across the ChickRH6 panel. This RF value is within the range (6.8% - 55.7%) observed in other studies reported for this panel [13, 29, 30].
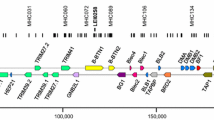
Using two-point analysis, the NK-lysin gene was included in a linkage group of 104 markers corresponding to microchromosome GGA22. After multipoint analysis, the framework map was composed of 23 markers covering 351.8 cR. Three additional markers AEBP1, LRRTM4 (leucine-rich repeat transmembrane neuronal 4) and ADRA1A (adrenoceptor alpha 1A) were integrated at their best possible locations on the comprehensive map (Figure 2).

Radiation hybrid mapping of the NK-lysin gene and other markers on chromosome GGA22. The GGA22 RH map (left) is 352 cR long and is aligned to the chicken genome assembly (right); the location of the NK-lysin gene is indicated at the distal end of chromosome GGA22.
Discussion
SNPs are probably the most abundant genetic variants in animal genomes and these variations can be associated with or even underlie phenotypic traits, including disease susceptibility. Previously, we identified a SNP at nucleotide 271 of the NK-lysin coding sequence and discovered that the encoded protein variants have different properties and also different cytotoxicities for bacteria and anticancer activity [26].
The objective of this study was to map the NK-lysin gene in the chicken genome and to evaluate the distribution of the SNP alleles in common chicken breeds and inbred lines. Some breeds or lines were available only as pooled DNA from multiple animals of that line. However, quantitative analysis of pooled DNA samples is recognized as a legitimate approach to approximate allele frequencies of SNPs [31]. Thus, we analyzed the peak area and height of sequencing chromatograms and compared these to a reference standard of known amounts of mixed A and G allele DNA. Since allele frequencies can only be estimated, we are unable to conclude that any breed or line is fixed for either the A or G allele. We can, however, predict very low frequencies of the minor allele in breeds for which we only detected a single peak. These analyses revealed that the A allele is more common than the G allele across all tested heritage breeds and inbred lines.
RH mapping analysis revealed that the NK-lysin gene is located in the distal region of chromosome GGA22. The RH map produced here is in agreement with the sequence assembly over the length of GGA22, extending it slightly with the addition of NK-lysin. Chromosome GGA22 is a microchromosome, approximately 4 Mb long. Microchromosomes represent about one-third of the total avian genome size, and have been found to have a higher gene density than macrochromosomes [32]. Because microchromosomes are not easy to identify cytogenetically and because they lack microsatellite markers, it is difficult to localize genes on specific microchromosomes. Thus, many of the small linkage groups in the chicken genome that have not been placed on the genetic map or genome assembly are assumed to be located on the microchromosomes [32]. RH mapping analysis revealed that the NK-lysin gene was located very near to the NUDCD3 (NudC domain containing 3) gene on GGA22. Our data place NK-lysin, and the 10 kb contig NW_003779909, distal to NUDCD3, the most telomeric gene marker in the sequence assembly. Its location near the telomere may explain the previous difficulty in placing this contig in the assembled chicken genome sequence.
Granulysin (GNLY), the human counterpart of NK-lysin, is located on human chromosome 2 between the genes SFTPB (surfactant-protein B) and ATOH8 (atonal homolog 8). This genomic organization is well conserved on cattle chromosome 11, pig chromosome 3, horse chromosome 15, chimpanzee chromosome 2 and dog chromosome 17. While there is no known SFTPB gene in the chicken genome, the NUDCD3 gene at the telomeric end of GGA22 is located on human chromosome 7 and bovine chromosome 4. The chicken contig NW_003779909 contains the genes NK-lysin, AEBP1 and POLD2, which are all three located on the same human chromosome 7 and bovine chromosome 4. Thus, a segment that contains these three mammalian genes is conserved in chicken, but the NK-lysin gene disrupts the otherwise conserved synteny.
Conclusions
Previous reports have described the cloning of the chicken NK-lysin gene [24] and its antimicrobial and anticancer activity [26] but its location in the chicken genome was unknown. Here, we used ChickRH6 to localize the NK-lysin gene in the chicken genome at the distal end of GGA22. Two additional genes, AEBP1 and POLD2, are located in the same NW_003779909 contig, and thus were also indirectly mapped to GGA22. Previously, we reported a functionally significant SNP in the NK-lysin coding sequence of two different chicken breeds. Here, we examined this SNP among a large number of different inbred lines and heritage breeds and found that the A allele, which has the higher antimicrobial activity, was more common than the G allele in our tested inbred lines and heritage breeds.
References
International Chicken Genome Sequencing Consortium: Sequence and comparative analysis of the chicken genome provide unique perspectives on vertebrate evolution. Nature. 2004, 432: 695-716.
Morisson M, Denis M, Milan D, Klopp C, Leroux S, Bardes S, Pitel F, Vignoles F, Gerus M, Fillon V, Douaud M, Vignal A: The chicken RH map: current state of progress and microchromosome mapping. Cytogenet Genome Res. 2007, 117: 14-21.
Womack JE, Johnson JS, Owens EK, Rexroad CE, Schlapfer J, Yang YP: A whole-genome radiation hybrid panel for bovine gene mapping. Mamm Genome. 1997, 8: 854-856.
Yerle M, Pinton P, Robic A, Alfonso A, Palvadeau Y, Delcros C, Hawken R, Alexander L, Beattie C, Schook L, Milan D, Gellin J: Construction of a whole-genome radiation hybrid panel for high-resolution gene mapping in pigs. Cytogenet Cell Genet. 1998, 82: 182-188.
Chowdhary BP, Raudsepp T, Kata SR, Goh G, Millon LV, Allan V, Piumi F, Guerin G, Swinburne J, Binns M, Lear TL, Mickelson J, Murray J, Antczak DF, Womack JE, Skow LC: The first-generation whole-genome radiation hybrid map in the horse identifies conserved segments in human and mouse genomes. Genome Res. 2003, 13: 742-751.
Priat C, Hitte C, Vignaux F, Renier C, Jiang Z, Jouquand S, Cheron A, André C, Galibert F: A whole-genome radiation hybrid map of the dog genome. Genomics. 1998, 54: 361-378.
Menotti-Raymond M, David VA, Roelke ME, Chen ZQ, Menotti KA, Sun S, Schäffer AA, Tomlin JF, Agarwala R, O'Brien SJ, Murphy WJ: Second-generation integrated genetic linkage/radiation hybrid maps of the domestic cat (Felis catus). J Hered. 2003, 94: 95-106.
McCarthy LC, Terrett J, Davis ME, Knights CJ, Smith AL, Critcher R, Schmitt K, Hudson J, Spurr NK, Goodfellow PN: A first-generation whole genome-radiation hybrid map spanning the mouse genome. Genome Res. 1997, 7: 1153-1161.
Morisson M, Lemiere A, Bosc S, Galan M, Plisson-Petit F, Pinton P, Delcros C, Feve K, Pitel F, Fillon V, Yerle M, Vignal A: ChickRH6: a chicken whole-genome radiation hybrid panel. Genet Sel Evol. 2002, 34: 521-533.
Rao M, Morisson M, Faraut T, Bardes S, Feve K, Labarthe E, Fillon V, Huang Y, Li N, Vignal A: A duck RH panel and its potential for assisting NGS genome assembly. BMC Genomics. 2012, 13: 513-
Leroux S, Dottax M, Bardes S, Vignoles F, Feve K, Pitel F, Morisson M, Vignal A: Construction of a radiation hybrid map of chicken chromosome 2 and alignment to the chicken draft sequence. BMC Genomics. 2005, 6: 12-
Rabie TS, Crooijmans RP, Morisson M, Andryszkiewicz J, van der Poel JJ, Vignal A, Groenen MA: A radiation hybrid map of chicken Chromosome 4. Mamm Genome. 2004, 15: 560-569.
Pitel F, Abasht B, Morisson M, Crooijmans RP, Vignoles F, Leroux S, Feve K, Bardes S, Milan D, Lagarrigue S, Groenen MA, Douaire M, Vignal A: A high-resolution radiation hybrid map of chicken chromosome 5 and comparison with human chromosomes. BMC Genomics. 2004, 5: 66-
Morisson M, Jiguet-Jiglaire C, Leroux S, Faraut T, Bardes S, Feve K, Genet C, Pitel F, Milan D, Vignal A: Development of a gene-based radiation hybrid map of chicken Chromosome 7 and comparison to human and mouse. Mamm Genome. 2004, 15: 732-739.
Morisson M, Leroux S, Jiguet-Jiglaire C, Assaf S, Pitel F, Lagarrigue S, Bardes S, Feve K, Faraut T, Milan D, Vignal A: A gene-based radiation hybrid map of chicken microchromosome 14: comparison to human and alignment to the assembled chicken sequence. Genet Sel Evol. 2005, 37: 229-251.
Jennen DG, Crooijmans RP, Kamps B, Acar R, van der Poel JJ, Groenen MA: Comparative map between chicken chromosome 15 and human chromosomal region 12q24 and 22q11-q12. Mamm Genome. 2003, 14: 629-639.
Solinhac R, Leroux S, Galkina S, Chazara O, Feve K, Vignoles F, Morisson M, Derjusheva S, Bed'hom B, Vignal A, Fillon V, Pitel F: Integrative mapping analysis of chicken microchromosome 16 organization. BMC Genomics. 2010, 11: 616-
Douaud M, Feve K, Gerus M, Fillon V, Bardes S, Gourichon D, Dawson DA, Hanotte O, Burke T, Vignoles F, Morisson M, Tixier-Boichard M, Vignal A, Pitel F: Addition of the microchromosome GGA25 to the chicken genome sequence assembly through radiation hybrid and genetic mapping. BMC Genomics. 2008, 9: 129-
Andersson M, Gunne H, Agerberth B, Boman A, Bergman T, Sillard R, Jornvall H, Mutt V, Olsson B, Wigzell H, Dagerlind A, Boman HG, Gudmundsson GH: NK-lysin, a novel effector peptide of cytotoxic T and NK cells. Structure and cDNA cloning of the porcine form, induction by interleukin 2, antibacterial and antitumour activity. EMBO J. 1995, 14: 1615-1625.
Pena SV, Hanson DA, Carr BA, Goralski TJ, Krensky AM: Processing, subcellular localization, and function of 519 (granulysin), a human late T cell activation molecule with homology to small, lytic, granule proteins. J Immunol. 1997, 158: 2680-2688.
Wang Q, Bao B, Wang Y, Peatman E, Liu Z: Characterization of a NK-lysin antimicrobial peptide gene from channel catfish. Fish Shellfish Immunol. 2006, 20: 419-426.
Davis EG, Sang Y, Rush B, Zhang G, Blecha F: Molecular cloning and characterization of equine NK-lysin. Vet Immunol Immunopathol. 2005, 105: 163-169.
Espino AM, Hillyer GV: Molecular cloning of a member of the Fasciola hepatica saposin-like protein family. J Parasitol. 2003, 89: 545-552.
Hong YH, Lillehoj HS, Dalloul RA, Min W, Miska KB, Tuo W, Lee SH, Han JY, Lillehoj EP: Molecular cloning and characterization of chicken NK-lysin. Vet Immunol Immunopathol. 2006, 110: 339-347.
Hong YH, Lillehoj HS, Siragusa GR, Bannerman DD, Lillehoj EP: Antimicrobial activity of chicken NK-lysin against Eimeria sporozoites. Avian Dis. 2008, 52: 302-305.
Lee MO, Kim EH, Jang HJ, Park MN, Woo HJ, Han JY, Womack JE: Effects of a single nucleotide polymorphism in the chicken NK-lysin gene on antimicrobial activity and cytotoxicity of cancer cells. Proc Natl Acad Sci USA. 2012, 109: 12087-12092.
de Givry S, Bouchez M, Chabrier P, Milan D, Schiex T: CARHTA GENE: multipopulation integrated genetic and radiation hybrid mapping. Bioinformatics. 2005, 21: 1703-1704.
Voorrips RE: MapChart: software for the graphical presentation of linkage maps and QTLs. J Hered. 2002, 93: 77-78.
Tian Y, Lu LZ, Fu Y, Tao ZR, Shen JD, Wang DQ, Yuan AP, Yin ZZ: Assignment of CCR7 gene to chicken chromosome 27 by radiation hybrid panel mapping. J Zhejiang Univ Sci B. 2007, 8: 314-317.
Tian Y, Lu L, Fu Y, Zhao J, Zhang C, Yuan Q, Shen J: Assignment of Dicer gene to chicken chromosome 5 by radiation hybrid panel mapping. Biochem Genet. 2007, 45: 239-243.
Olsson C, Liljedahl U, Syvanen AC: Quantitative analysis of SNPs in pooled DNA samples by solid-phase minisequencing. Meth Molec Biol. 2003, 212: 167-176.
Groenen MA, Cheng HH, Bumstead N, Benkel BF, Briles WE, Burke T, Burt DW, Crittenden LB, Dodgson J, Hillel J, Lamont S, Ponce de Leon A, Soller M, Takahashi H, Vignal A: A consensus linkage map of the chicken genome. Genome Res. 2000, 10: 137-147.
Acknowledgements
This work was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the ministry of education (2012R1A6A3A03039557).
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
MOL and JEW designed the experiment and MOL, and YH performed initial RH mapping. HHC, WMM, SJL, HSL, SHL provided DNA and information on the origins of stocks and strains and contributed to writing the manuscript. MM, AV and EY constructed the refined GGA22 RH map and assigned NK-lysin. All authors have read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

Cite this article
Lee, M.O., Yang, E., Morisson, M. et al. Mapping and genotypic analysis of the NK-lysin gene in chicken. Genet Sel Evol 46, 43 (2014). https://doi.org/10.1186/1297-9686-46-43
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1297-9686-46-43