
Abstract
This review article provides data on the current state of the pathogenesis peculiarities of body and lung inflammation (pneumonia) under the influence of damaging factors of various nature: infectious agents, chemical toxicants, as well as incorporated radionuclides, etc. The peculiarities of inflammation itself, as a typical pathological process, are considered. Information on mediators that induce the so-called pro-resolving phase of inflammation manifestations is given. Approaches to the neuroimmune correction of non-specific inflammation are substantiated. Data on the following alternative approaches to the correction of nonspecific inflammation are summarized: factors of the coagulation system, modulators of the integrated stress response, and modulators of sigma-1 receptors. Based on the data presented, general directions for the treatment of nonspecific pneumonia are formulated, including reflexogenic and anti-inflammatory therapy in combination with multimodal drugs, as well as pro-resolving therapy in combination with drugs that prevent fibrosis.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
INTRODUCTION
Infectious agents, chemical toxicants, and incorporated radionuclides (hereinafter referred to as damaging factors) mediate specific and non-specific respiratory disorders as a result of aerogenic exposure. As this takes place, the formation of an inflammatory reaction acts as a key trigger mediating their damaging effect. The severity and intensity of this process depends on the characteristics of the bioavailability of the damaging factors and on the size of the inhaled aerosol particles: the larger the particles of the inhaled aerosol, the lower the probability of their deposition in the lower respiratory tract (Cargnello Roux, 2011). The geometry of various regions of the tracheobronchial tree plays an important role in the aerosol deposition profile. The diameter determines the necessary displacement of a particle before it contacts the epithelial lining, the cross-section determines the speed of air movement (Basharin et al., 2022), and the nature of bronchial branching affects the mixing between inhaled and reserve air (Chepur et al., 2019; Codagnone et al., 2018).
In the case of aerosol intake, the basis for subsequent interactions of a damaging factor with the body is the level of the neuroimmunoendocrine reaction, which determines the intensity of nonspecific inflammatory processes, often resulting in death or pronounced morphological and functional changes in the lung tissue, significantly reducing the quality of life, the associated illnesses are: chronic obstructive pulmonary disease, diffuse pulmonary fibrosis, respiratory failure etc. Therefore, the tactics for prevention and treatment of injured individuals with this pathology should include emergency, if possible, inhalation administration of drugs that prevent the induction of trigger mechanisms of the inflammatory reaction, level the initial phase of the inflammation and accelerate the formation of an adaptive complex of structural and functional rearrangements.
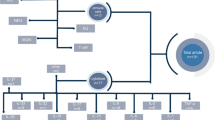
To determine the possible directions of therapy for inhalation lung injuries, one should take into account the molecular and cellular mechanisms of induction, maintenance, and resolution of inflammation inherent in the effect of a wide range of damaging factors. At the moment, the scheme for the correction of nonspecific pneumonia (pulmonoprotection) is based on the use of anti-inflammatory and pro-resolving therapy, as well as on blocking pathological reflex reactions (Fig. 1).

The main directions of therapy for nonspecific pneumonia caused by adverse environmental factors.
CORRECTION OF THE INFLAMMATORY REACTION ITSELF
It is known that the interaction of the body with a damaging factor initiates the release of pro-inflammatory mediators. Thus, direct damage to cells by chemical toxicants leads to the release of molecular fragments of DAMPs (damage-associated molecular patterns) associated with the damage, while damage caused by a biopathogen releases molecular fragments of PAMPs (pathogen-associated molecular patterns) associated with the pathogen. There is also a universal system of molecular signals associated with disruption of cellular homeostasis, gene expression, inflammation, allergy, and tissue repair. The above signal molecules are represented by proteins, RNA, DNA, lipopolysaccharides (LPSs), exo- and endotoxins, various glycans that activate pattern recognition receptors (toll-like receptors TLR1–11, NOD-like receptors NBS-LRR (nucleotide binding site leucine-rich repeat), RIG-I-like receptors (RLR)), membrane 2'‑5'-oligoadenylate synthetases, protein kinases activated by double-stranded RNA, etc. (Chepur et al., 2019; Di Salvo et al., 2021). The combined or isolated action of these molecular patterns determines the induction of inflammation. Some xenobiotics (phlogogens) can predetermine their activation without direct tissue damage, presumably through the stress molecules MICA, MICB, RAE-1, ULBP1, ULBP2, and ULBP3 with high affinity for killer-inhibiting receptors of NK cells (Zingoni et al., 2018). It is believed that the tissue- and stress-specific representation, concentration, and duration of the effect of these signals program the inflammatory response and determine its specificity. The circumstances mentioned above dictate the need to identify the most significant predictors of nonspecific inflammation.
Regardless of the nature of the damaging factor, disruption of cell integrity leads to the release into the extracellular environment of a significant amount of cytosolic and intranuclear contents, including conservative hydrophobic domains, the alarmins. Alarmins are represented by defensins, cathelicidins, eosinophil neurotoxin, nuclear non-histone protein B1 (HMGB1), heat shock proteins HSP, nucleic acids, histones, and nucleosomes. Acting as ligands for specific receptors—TLRs, glycan-binding receptor RAGE (receptor for advanced glycation endproducts), phagocytosis receptors, including MerTK and MFG-E8, C-type lectin receptor (CLR), scavenger receptor (SR), etc.—alarmins induce an inflammatory response (Anfinogenova et al., 2020). Thus, massive tissue necrosis during surgical operations leads to a systemic inflammatory reaction, which ends in death in 4–7% of cases (Bakulina et al., 2017). Alarmin receptors act as an associating link in response to damaging stimuli, and the impact on them is justified from the viewpoint of the universality of the correction of nonspecific inflammation.
The influence of DAMPs is mediated by a significant representation of receptors, among which TLR4 plays a significant role. Demonstrating the ability to receive a huge number of DAMPs, including bacterial LPS, TLR4 serves as a universal trigger for the development of nonspecific inflammation, the reinforcement of which is determined by cells of the human immune system. Existing in the form of homo- and heterodimeric complexes with receptors of other subtypes, which are located both on the cytolemma and intracellularly in the cytoplasm, TLR4 triggers the mechanisms of innate immunity. The ability to bind to all adapter proteins determines the main role of TLR4 in the formation of nonspecific inflammation, including cytokine secretion, the production of pro-inflammatory IL-13 and IL-18, as well as autophagy and apoptosis processes (Bakulina et al., 2017).
TLR4s are involved in the pathogenesis of chronic inflammatory diseases, acute sepsis, and the life-threatening complications of dangerous infections (Perrin-Cocon et al., 2017). Thus, TLR4 gene knockout protects rodents from death as a result of impaired myocardial contractility caused by Bacillus anthracis toxin (Kandadi et al., 2012). It has been proven (Imai et al., 2008) that the severity of acute lung injury under the influence of damaging factors depends on the degree of activation of the innate immunity signaling pathway TLR4–TRIF–TRAF6, where TLR4 is toll-like receptor 4, TRIF is TIR-domain-containing adapter-inducing interferon-, and TRAF6 is TNF receptor-associated factor 6. In turn, the use of TLR4 antagonists prevents the secretion of pro-inflammatory cytokines, LPS-induced dendritic cell migration, and also reduces acute lung injury and mortality associated with viral infection.
μ-Оpioid receptor antagonists (naloxone and naltrexone) can be used as TLR4 blockers. Their effectiveness against TLR4-dependent pathology, neuropathic pain or addiction, has been demonstrated in a number of studies (Watkins et al., 2014; Wang et al., 2016). Currently, the dextrorotatory isomer of naltrexone is considered the most promising means of correcting the hyperimmune response (Selfridge et al., 2015). The ability to block the cascade of PAMP-associated LPS-dependent stimuli in relation to TLR4 is associated with this isomer.
The release of mediators into the bloodstream promotes the recruitment of neutrophils and monocytes of bone marrow origin, which migrate to the focus of inflammation, where they undergo differentiation. In the focus of inflammation, epithelial and immunocompetent cells begin to produce cytokines and chemokines: MCP-1 (monocyte chemoattractant protein 1), CCR2 (C-C motif chemokine receptor type 2), CXCL-1 (C-X-C motif chemokine ligand 1), among which the priority role in the development and progression of the inflammatory process belongs to TNF-α and IL-1β. As a result of the action of proteolytic enzymes, TNF-α is released from its association with the cell membrane and interacts with the receptor of the same name on the surface of macrophages, dendritic cells, and T lymphocytes and activates the apoptosis domain through the mitogen-activated protein kinase p38 MAPK and NF-κB. MAPK activates the Ets transcription factors and, as a result, the expression of the fas genes, resulting in an increase in the production of class II pro-inflammatory proteins (DP, DQ, DR) of HLA antigens of the T-cell receptor, AP-1, which regulates the transcription of cyclin 1D and the pro-inflammatory growth factors NGF, EGF, PDGF, etc. (Cargnello and Roux, 2011). The result of the activation of neutrophils and monocytes is the accumulation of reactive oxygen and nitrogen species.
In terms of screening effective pulmonary protectors, it is possible to use a model of lung damage by cigarette smoke, based on the general pathogenetic mechanisms of lung damage inherent in many damaging factors (Fig. 2).

Pathogenetic links of lung damage by cigarette smoke.
Among the molecular targets, the impact on which can contribute to the prevention and treatment of nonspecific pneumonia, it is advisable to mark out the following (Yao et al., 2008):
(1) Cell signaling inhibitors: type B (PDE4B) and type 4 (roflumilast, cilomilast, GRC3886 and GSK842470) phosphodiesterase inhibitors; p38-MAPK inhibitors (SB 203580, SB 239063 and RWJ 67657, SD282, GSK-681323, GSK-85633); IKK-2 inhibitors (under development); and inhibitors of phosphoinositide-3-kinase PI3K of δ- and γ-types (LY294002).
(2) Cytokine and chemokine inhibitors: CCL2 (MCP-1) and CCR2 antagonists (CCX915, INCB3284, antibodies ABN912, INCB8696, JNJ-27553292, SKL-2841, and INCB3344); CXCL1, CXCL8 antagonists (SB-265610, SCH 527123); TNF- inhibitors (infiximab, etanercept, adalumimab).
(3) Modifiers of HDAC histone deacetylases (vorinostat and romidepsin).
(4) Antiproteinases: neutrophil elastase inhibitors (ZD0892, AZD3342, α-1-antitrypsin); inhibitors of MMP-1, -2, -9, -12 (macrophage) types (marimastat, BMS-561392, and GW3333).
PHARMACEUTICALS THAT INDUCE THE PRO-RESOLVING PHASE OF THE INFLAMMATORY REACTION
Despite the fact that resolution is the outcome of inflammation of any etiology, this period is characterized by an active process regulated by special pro-resolving lipid mediators (SPMs). Their synthesis begins at the moment of neutrophil activation (minutes, hours) and continues until the functional and structural restoration of lung tissue (days, weeks). SPMs reduce the severity of the inflammatory response without depression of the immune system, protect tissues from damage, accelerate the removal of etiological factors of inflammation and apoptotic leukocytes, helping to restore homeostasis (Krishnamoorthy et al., 2018). SPMs are formed as a result of the fermentation of fatty acids: lipoxins are formed from arachidonic acid; E-series resolvins, from eicosapentaenoic acid; and D-series resolvins, protectins, and maresins, from docosahexaenoic acid. Under certain conditions, highly active conjugates can be formed, for example, SPM-sulfide or SPM-acetyl derivatives (Chiang and Serhan, 2020).
A large number of recent publications devoted to resolvins are associated, among other things, with the prospects for their use in the treatment of COVID-19, since a hyperinflammatory reaction, up to a cytokine storm, is one of the main causes of high mortality during coronavirus infection (Panigrahy et al., 2020). The basis for this was the results of the use of pro-resolving therapy on models of lung tissue pathology, these results showed that the use of resolvin D1 is the most promising. Thus, it was shown (Xia et al., 2019) that resolvin D1, when administered intraperitoneally, reduced ventilation-associated damage to the lungs of mice, the severity of emphysema and chronic inflammation in the model of chronic (Hsiao et al., 2015) and acute (Hsiao et al., 2013) exposure to cigarette smoke. Resolvin D1 contributed to the resolution of pneumonia using a model of infection caused by Pseudomonas aeruginosa (Codagnone et al., 2018) or a model of LPS-induced lung injury (Wang et al., 2014). Resolvin D1 limited septic lung injury in mice induced by ligation and puncture of the caecum (Zhuo et al., 2018).
The most important feature of the biochemical transformations of resolvins is associated with their sensitivity to the acetylating effect of aspirin. Thus, in a model of paraquat-induced lung injury in mice, aspirin-induced resolvin D1 had a protective effect by reducing oxidative stress, inflammatory response, and pulmonary edema (Hu et al., 2019), and in a model of pneumonia of mixed etiology, it reduced inflammation and infection of the lungs (Wang et al., 2017).
In the resolution of inflammation, in addition to resolvins, lipoxins, and maresins, the following proteins are also involved: annexin A1, and TIM-4 (T-cell immunoglobulin and mucin domain 4), the action of which is realized due to the modification of the signaling pathway (PI3K associated with ATP) and the work of intracellular regulators of inflammatory reactions (the protein kinase B (PKB) family and serine-threonine specific protein kinase). It was demonstrated that by binding to the specific N-formyl peptide receptor type 2 (FPR2) of leukocyte membranes, annexin A1 reduces epithelial adhesion, processes of leukocyte migration, chemotaxis, and phagocytosis induced by N-formylmethionine-containing oligopeptides (including products of arachidonic acid) (Schloer et al., 2019).
The above data indicate the perspectives of using resolvins, in particular, resolvin D1, as agents that induce the pro-resolving phase of nonspecific inflammation.
NEUROIMMUNE CORRECTION OF NONSPECIFIC PNEUMONIA
The acute impact of damaging factors and, most importantly, toxic substances on the lungs is caused not only by a violation of tissue integrity due to direct cytolytic action, but also by hyperactivation of the body’s protective functions, such as mucus secretion, cough, bronchiospasm, edema, and neurogenic inflammation. The intensity of reflex reactions can be reduced by opening the reflex arc represented by the afferent, central, and efferent phases. This approach is successfully implemented using β2-adrenergic agonists and M3-selective anticholinergic agents as means of urgent therapy for injuries with pulmonary toxicants (De Virgiliis and Di Giovanni, 2020).
The respiratory epithelium contains pulmonary neuroendocrine cells, some of which are associated with the processes of nerve cells and are called NEBs (neuroepithelial bodies), which are capable of releasing immunotropic amines and peptides. The role of NEBs in the development of the lungs during inflammatory diseases, as well as the ability of NEBs to act as stem cells have been proven (Yeger et al., 2019).
It was shown that the innervation of NEBs, by analogy with the vascular endothelium, is carried out by sensory fibers of the nodose and jugular ganglia of n. vagus, which express membrane receptors for neurotropic factors and purines, respectively. Activation of fibers by endogenous and exogenous mediators causes depolarization of neurons, followed by the release of neuropeptides both in the lungs and near the secondary neurons of the nucleus of the solitary pathway, triggering a reflex cough. Low conduction (~1 m/s) unmyelinated C fibers that express TRPV1 receptors are also involved in the development of cough (Nassenstein et al., 2018).
Parasympathetic efferent regulation of neuroimmune processes in the lungs is anatomically and histologically provided by preganglionic neurons of the dorsal motor nucleus of n. vagus reaching the intramural ganglia of the trachea containing acetylcholine-producing neurons and non-cholinergic neurons of Auerbach’s plexus near the outer longitudinal muscular layer of the esophagus. The latter release vasoactive intestinal peptide (VIP) and NO as neurotransmitters (Yildiz-Pekoz and Ozsoy, 2017).
Unlike neuronal acetylcholine, non-neuronal acetylcholine release is non-quantum. Thus, in an experimental model of bronchial asthma in animals, the use of anticholinergic drugs provided a bronchodilatation effect, a decrease in mucus and inflammation, and remodeling through the process of blocking the M3R. The pro-inflammatory effect of acetylcholine, including an increase in neutrophil chemotaxis and stimulation of the production of reactive oxygen species, is probably also mediated by M3R activation, as confirmed using pharmacological probes (Shen et al., 2020).
VIP-ergic parasympathetic nerve fibers are extensively co-localized with a peptide that activates pituitary adenylate cyclase. Both mediators increase vasodilation and bronchodilatation. In addition, mast cells and Th2 lymphocytes synthesize VIP which has an anti-inflammatory effect that has been confirmed in various models of pneumonia. Thus, prophylactic intratracheal injection of a VIP analogue reduced the recruitment of inflammatory cells by 70% in the bronchoalveolar lavage fluid (Prescott et al., 2020). Late interaction between sensory neurons and eosinophils, as well as antigen-presenting cells, was confirmed histologically. Eosinophils influence the branching of sensory neurons, presumably through excessive secretion of nerve growth factor. In turn, it has been proven that nerve fibers stimulate the migration of eosinophils due to the secretion of eotaxin-1. A significant role in the organization of neuroimmune interactions belongs to mast cells, which can activate the TRPV1 receptor through the neuronal H1 receptor.
The clinical significance of neuroimmune interaction in providing inflammatory processes is confirmed by the clinical success of refractory therapy of bronchial asthma by bronchial thermoplasty and vagus nerve stimulation (Caravaca et al., 2019). Neuroimmune interactions at the anatomical level, acting as targets for the pharmacological correction of inflammation are schematically shown in Fig. 3.

Schematic representation of neuroimmune interactions that act as targets for the pharmacological correction of inflammation. P2XR—ATP-gated P2X receptor cation channel family; P2YR—ATP-gated P2Y receptor cation channel family; AR—adrenoreceptors; M2R—muscarinic acetylcholine receptor M2; M3R—cholinergic/acetylcholine receptor M3; CGRP—calcitonin gene-related peptide; β2AR—β2 adrenoreceptor; VIP—vasoactive intestinal peptide; NO—nitric oxide; TRPV1R—transient receptor potential vanilloid subtype 1; TRPA1R—transient receptor potential cation channel subfamily V member 1; TRPM8R—transient receptor potential cation channel subfamily M (melastatin) member 8; NKA—neurokinin A; SP—substance P; 5-HT—serotonin.
Stimulation of parasympathetic fibers induces bronchiospasm, mucus hypersecretion, and vasodilation of pulmonary vessels through activation of acetylcholine muscarinic M3 receptors (CHRM3). Muscarinic acetylcholine receptors M2 (CHRM2) are present on presynaptic terminals, and their activation reduces the release of acetylcholine and prevents bronchiospasm. Many biological factors, such as parainfluenza viruses, disrupt the function of CHRM2, which leads to an increase in the release of acetylcholine and an increase in smooth muscle tone. Taking into account the ability of pulmonary macrophages to express CHRM2 and CHRM3, it can be concluded that there is a correlation between the activity of the acetylcholine system and the severity of inflammation (Koarai et al., 2012). The positive feedback is confirmed by the ability of IFNγ, TNF-α, and IL-1β to increase CHRM2 expression, which has been shown in models of viral infections, including SARS-CoV-2 (Rynko et al., 2014). Activation of C-fibers containing calcitonin gene-related peptide CGRP, tachykinins (substance P and neurokinin A) causes spasm of airway smooth muscles, vasodilation, excessive mucus formation, which together form the clinical manifestations of nonspecific inflammation syndrome. It has been shown that P2RY1+ neurons of the vagus nerve trigger the inflammatory reaction cascade in response to inhalation intake of various irritants, while Piezo2 airway stretch receptors integrate and coordinate protective reflexes, in particular, laryngospasm (Nonomura et al., 2017). Sensory cells rich in NEBs with bioactive mediators—bombesin, serotonin, and CGRP participate in the implementation of non-specific inflammation under the influence of damaging factors (Noguchi et al., 2020).
Promising targets for pharmacological correction of reflex-dependent inflammation may be cationic channels acting through a transient receptor potential channel (TRP channel): vanilloid (TRPV1), ankyrin (TRPA1), and melastatin (TRPM8), as well as receptors for mediators of efferent fibers of the lung tissue (substance P and CGRP). TRPA1, TRPV1, and TRPM8, localized on the nociceptive peripheral neurons are responsible for the perception of acute and chronic pain, the initiation of the reflex cough, and the development of asthma attacks, lung damage, and aseptic inflammation (Grace et al., 2013). Their peculiarity is the possibility of activating inflammatory processes without connection with a specific pulmotoxicant, but only through ingress of particles with a diameter of less than 2.5 mm, the so-called fine particles (FP 2.5). It has been experimentally shown that prophylactic treatment with antagonists of TRPV1- and TRPA1 receptors effectively prevents the pneumonia and bronchial hypersensitivity caused by intranasal instillation of mice with FP 2.5 a dose of 7.8 mg/kg (Xu et al., 2019).
Another member of the TRP receptor family, TRPC6, is expressed on eosinophils, neutrophils, mast cells, and CD4+ lymphocytes, and its activation is involved in the pathogenesis of asthma and allergic inflammation. In turn, activation of TRPV1 on the C fibers of sensory neurons leads to the release of tachykinins and increased recruitment of immune cells (Jia and Lee, 2007).
TRPA1 receptors are activated by lachrymators (CS, CN, CR), drugs (paracetamol, diphenhydramine), combustion products, chemotherapeutic agents, cigarette smoke, and other pollutants (Belvisi and Birrell, 2017). Currently, the following are considered as the most likely candidates capable of blocking TRP channels: AG489 isolated from the venom of the Agelenopsis aperta spider, HCRG21 isolated from the sea anemone Heteractis crispa, JYL-1421, and AMG8562 (Kvetkina et al., 2019).
ALTERNATIVE APPROACHES TO THE CORRECTION OF NONSPECIFIC INFLAMMATION
Blood Coagulation Factors
Blood coagulation factors are classified as pro-inflammatory factors, while substances that have the opposite effect have an anti-inflammatory action, as shown in various models of lung injury (Choi et al., 2008). Thus, in a model of LPS-induced lung damage in rats, the administration of heparin by a nebulizer at a dose of 1000 UI/kg provided a decrease in the expression of the plasminogen gene, as well as the effectors TGFβ—Smad 2, Smad 3—NF-κB—P-selectin, and CCL2. Preventive inhalation of heparin had a pronounced protective effect (Chimenti et al., 2017). In addition, heparin had an anti-asthma effect in models of allergen-, adenosine-, and exercise-induced asthma by preventing mast cell degranulation (Mousavi et al., 2015). Inhalation of heparin in conditions accompanied by inflammation of the lung tissue and systemic hypercoagulability is becoming a mandatory indication for patients with severe forms of COVID-19 (van Haren et al., 2020). It has been proven that inhalations of unfractionated heparin provide an anti-inflammatory effect due to the following:
— block of heparan sulfate-containing receptors of the cytolemma, considered as a translocation cofactor of the peplomer spike protein to the receptor-binding domain of the angiotensin-converting enzyme 2 receptor (ACE2) (Clausen et al., 2020);
— decreased expression of pro-inflammatory mediators and inhibition of the complement system due to interaction with the C1 component (Shi et al., 2021);
— disaggregation of DNA and actin followed by activation of endogenous DNase and a decrease in electrostatic interactions between mucin molecules (Broughton-Head et al., 2007);
— inactivation of thrombin, factor Xa, kallikrein, serine proteases and reduction of excessive fibrin deposition (Camprubí-Rimblas et al., 2018).
In addition to heparin, antithrombin is of practical interest, which, with a similar anti-inflammatory activity to heparin, is devoid of the side effects of the latter (osteoporosis and thrombocytopenia). In a model of pneumonia in rats caused by Streptococcus pneumoniae, intravenous administration of antithrombin led to a decrease in the number of neutrophils in bronchoalveolar lavage, and after 48 hours it led to a significant drop in the levels of TNF-α, IL-6, and cytokine-induced neutrophil chemoattractant 3, CINC-3, also in the model of acute lung injury by endotoxin it led to a decrease in the expression of ERK1/2 and p38-MAPK (Sun et al., 2009).
Modulators of the Integrated Stress Response
Each eukaryotic cell, when exposed to damaging factors, reacts in an integrated way by reducing protein synthesis with two goals: both preventing the accumulation of defective forms and preventing the replication of the bioagent, as well as increasing gene expression, which makes it possible to specifically adapt either to damaging factors or to factors that trigger apoptosis (Emanuelli et al., 2020). For various stress factors, there are intracellular sensor kinases that trigger the integrated stress response (ISR). In the presence of double-stranded DNA (dsDNA) in the cytosol, cytosolic protein kinase R (PKR) and PKR-like endoplasmic reticulum kinase activate kinases of the eukaryotic initiation factor 2α (eIF-2α), in the case of oxidative stress and Fe2+ deficiency, the heme-regulated inhibitor (HRI) is activated, in the event of a deficiency of amino acids, the kinase that controls the general control nonderepressible 2 (GCN2) is activated. After phosphorylation of the α subunit of translation initiation factor 2 (eIF-2), the latter interacts with methionine transfer RNA (tRNA) on a ribosome and starts translation.
Normally, eIF-2α hydrolyzes bound GTP, which is replenished by the guanine nucleotide exchange factor eIF-2β. However, under the influence of damaging factors, eIF-2α is mainly represented by a phosphorylated form which easily binds to eIF-2β and inhibits further GTP metabolism.
Therefore protein synthesis can slow down significantly, which contributes to the activation of the transcription factors ATF4 and CHOP and an increase in the expression of the damage-induced protein (DIP) by the GADD34 gene. DIP forms a complex with protein phosphatase PP1 and G-actin, ensuring the functioning of eIF-2α-specific phosphatases and maintenance of translation. This mechanism allows cells to temporarily activate the ISR (whereas in the absence of such interaction, cell death can be observed) (Pakos-Zebrucka et al., 2016).
The function of ISR is viewed from opposite positions relating to viral invasion. This is due, as a rule, to the very nature of pathogens. PKR activation of viral DNA is considered part of antiviral immunity, but many viruses have improved systems to counteract this mechanism. For viruses of the families Togaviridae, Reoviridae, and hepatitis C virus, eIF-2α phosphorylation can promote translation of their mRNA (Fusade-Boyer et al., 2019). For other rapidly replicating viruses, on the contrary, the implementation of such a mechanism significantly slows down the intracellular assembly of the protein capsid of viral particles.
ISR is a universal nonspecific defense mechanism in response to the action of damaging factors of various nature. However, excessive ISR may be unfavorable for the cell and may not be adaptive. In this regard, pharmacological tools are being developed to reduce the intensity of ISR in the form of so-called inhibitors of the integrated stress response, ISRIB. Their mechanism of action is to bind by the allosteric site to eIF-2β, the main target of phosphorylated eIF-2α, and to enhance the recruitment of guanidine residues in the presence of phosphorylated eIF-2α (Zyryanova et al., 2021). Therefore, ISRIB converts an inactive complex involved in protein synthesis into an active one, restoring the cell’s potential to synthesize protective proteins. ISR dysfunction can underlie conditions associated with insufficient function of the endogenous defense system against damaging factors of various nature, which requires the study of the issue of targeted pharmacological correction of dysfunctional ISR. To date, several ISR-modulating compounds have been synthesized that are structurally similar to ISRIB.
Although ISR represents a novel target in pneumonia protection (Van’t Wout et al., 2014), there is already evidence for the effectiveness of ISR modulators in various respiratory pathologies. Thus, ISRIB at a dose of 2.5 mg/kg, when administered intraperitoneally, reduced the severity of bleomycin- and asbestosis-induced pulmonary fibrosis, including that caused by a decrease in excess collagen formation. In addition, ISRIB promotes the transformation of type 2 alveolar macrophages into type 1 alveolar macrophages, which is necessary to maintain gas exchange in the lungs (Watanabe et al., 2021). ISRIB, like salubrinal and Sal003, inhibits eIF-2α phosphatase. Sephin1 (a derivative of guanabenz) selectively blocks GADD34-phosphatase and stress-induced protein phosphatase 1 (PPP1R15A), as a result, the released serine/threonine phosphatase PP1 dephosphorylates eIF-2α. Apparently, due to the additional effect on protein phosphatase, the latter, after a course of administration for 11 days at a dose of 5 mg/kg, showed antiviral activity in the model of infection of rabbits with the myxoma virus. Thus, the use of ISR modulators to prevent fibrotic processes in the lungs after inhalation exposure to pathological factors may be promising . The anti-inflammatory potential of this class of compounds is well realized, especially in diseases associated with protein misassembly (folding), in storage diseases, however, there is still no unequivocal opinion regarding bacterial and viral lesions (Pierre, 2019).
Multimodal Agents
Since the inflammatory process is characterized by the involvement of a huge number of effector cells and their signaling pathways, it may be justified to use anti-inflammatory drugs of a multimodal type of action, for example, plant-derived substances related to flavonoids: quercetin and its analogue, dihydroquercetin (Adhikari et al., 2021).
Using a model of LPS-induced damage to the TC-1 cell line, it was shown that taxifolin reduces the cytotoxic effect and the level of lethality through modulation of NF-κB signaling (Liu et al., 2020). Using computer simulation, the potential of taxifolin and rhamnetin to act as inhibitors of SARS-CoV-2 main protease (Mpro) with satisfactory tolerability characteristics has been proven (Fischer et al., 2020). Taxifolin showed the ability to suppress oxidative stress and pneumonia caused by benzo[a]pyrene (125 mg/kg) when administered to mice at a dose of 20–40 mg/kg for 14 days. The corrective effect was due to the high level of expression of NF-E2-related factor 2 (Nrf2), NAD(P)H quinone dehydrogenase 1 (NQO1), heme oxygenase 1 (HO-1), and superoxide dismutase (SOD), where Nrf2 plays the leading role in the suppression of inflammation through inhibition of the NF-κB signaling pathway (Islam et al., 2021).
Quercetin exhibits anti-inflammatory, antioxidant properties, and also blocks lipid peroxidation, platelet aggregation, and vascular permeability. Its efficacy has been demonstrated in the model of LPS-induced TNF-α production in macrophages, LPS-induced IL-9 production in A54 lung cells, LPS-induced increase in mRNA and TNF-α in glial cells, FcεRI-mediated release of pro-inflammatory cytokines, tryptases, and histamine from mast cell culture (Jafarinia et al., 2020). Quercetin has a bronchodilatory effect, presumably by enhancing downstream signals from β-adrenergic receptors of bronchial smooth muscle and by inhibiting PDE4. Exposure was increased by introduction via inhalation with PBS buffer solution using a nebulizer and prevented methacholine induced increase in airway resistance. The noted ability of the substance to increase isoprenaline-induced relaxation of the bronchi may be required in the development of tachyphylaxis to β-adrenergic agonists, which complicates the treatment of not only an attack of bronchial asthma, but also treatment of damage by air pollutants.
Antidepressants, Sigma-1 Receptor Modulators
Fluvoxamine, a selective serotonin reuptake inhibitor (SSRI), has a high affinity for sigma-1 receptors at therapeutically relevant doses. The mechanisms of its anti-inflammatory and immunomodulatory properties are not fully understood, however, the ability to block the transfer of SARS-CoV-2 through endolysosomes and hypercoagulability in COVID-19 indicates a potential impact upon lysosome membrane formation (Marčec and Likić, 2021). In a double-blind, randomized study of adult patients with COVID-19, individuals treated with fluvoxamine had a lower likelihood of clinical deterioration within 15 days. Later, it was suggested that the positive therapeutic effect may also be associated with the inhibition of acid sphingomyelinase and the ability to influence membrane protonation and thereby promote the retention of the viral particle in the lysosome (Sukhatme et al., 2021). It is known that SSRIs affect platelet aggregation by reducing their serotonin content. After 12 weeks of therapy with fluvoxamine at a dose of 100–150 mg/day, the concentration of serotonin in platelets decreases by 86%, and in blood plasma, by 60% (Celada et al., 1992). By reducing the concentration of serotonin in platelets, SSRIs can reduce their potential for aggregation and prevent the state of hypercoagulability against the background of inflammation caused by damaging factors. The additional anti-inflammatory effect of fluoxamine is associated with the ability to inhibit the breakdown of melatonin in the liver, which leads to an increase in its concentration in blood plasma. By blocking mast cell degranulation, it reduces histamine-mediated tissue edema and hypercoagulability. On the other hand, fluvoxamine reduces the manifestations of hypercytokinemia in a model of LPS-induced sepsis. This effect may be mediated by the impact of fluvoxamine upon the endoplasmic reticulum resident protein, the sigma-1 receptor, which is considered a natural factor in inhibition of cytokine production (Rosen et al., 2019). The ability of fluvoxamine to lower levels of pro-inflammatory cytokines explains the reduction in excess IL-6 in patients who have had COVID-19.
Thus, the order of application of therapeutic agents aimed at reducing the activity of pro-inflammatory cytokines should be adjusted based on specific periods of potential growth in their level after exposure to damaging factors.
CONCLUSION
Acute and chronic effects of damaging factors of chemical and biological origin on the respiratory system and the body as a whole, are ubiquitous and represent a significant proportion in the structure of overall morbidity and mortality (Fig. 4). The use of standardized treatment regimens does not always lead to complete recovery of affected individuals. For example, those who have recovered from COVID-19 may display long-term consequences that reduce their quality of life. In this regard, there is a need to study and develop universal (non-specific) means of treating pneumonia, providing for the blockade of pathogenetic cascades, and targeted therapeutic and prophylactic effects on the key stages of inflammation in order to accelerate the solution of inflammation and interrupt maladaptive reflex reactions.

Chronology of molecular and cellular changes during pneumonia. ABB—air-blood barrier; damaging factors displayed from top to bottom: bacteria, cigarette smoke, industrial air pollutants, pulmonotoxicants like paraquat, viruses.
General directions for the treatment of nonspecific inflammation should include: reflexogenic therapy immediately after exposure to damaging factors; then, anti-inflammatory therapy in combination with multimodal drugs; then, pro-resolving therapy in combination with drugs that prevent fibrosis.
REFERENCES
Adhikari, B., Marasini, B.P., Rayamajhee, B., et al., Potential roles of medicinal plants for the treatment of viral diseases focusing on COVID–19: a review, Phytother. Res., 2021, vol. 35, no. 3, pp. 1298–1312.
Anfinogenova, N.D., Quinn, M.T., Schepetkin, I.A., and Atochin, D.N., Alarmins and c-Jun N-terminal kinase (JNK) signaling in neuroinflammation, Cells, 2020, vol. 9, no. 11, p. 2350.
Bakulina, L.S., Litvinenko, I.V., Nakatis, Ya.A., et al., Sepsis: Pozhar i bunt na tonushchem v shtorm korable. Chast’ 1. Triggery vospaleniya. Retseptsiya triggerov vospaleniya i signal’naya transduktsiya (Sepsis: Fire and Riot on a Ship Sinking in a Storm. Part 1. Triggers of Inflammation. Reception of Inflammatory Triggers and Signal Transduction), St. Petersburg: Severo-Zapad. Gos. Med. Univ. im. I.I. Mechnikova, 2017.
Basharin, V.A., Chepur, S.V., Tolkach, P.G., et al., Toksikologiya produktov goreniya polimernykh materialov (Toxicology of Combustion Products of Polymeric Materials), St. Petersbufg: Levsha, 2022.
Belvisi, M.G. and Birrell, M.A., The emerging role of transient receptor potential channels in chronic lung disease, Eur. Respir. J., 2017, vol. 50, no. 2, p. 1601357.
Broughton-Head, V.J., Shur, J., Carroll, M.P., et al., Unfractionated heparin reduces the elasticity of sputum from patients with cystic fibrosis, Am. J. Physiol.: Lung Cell. Mol. Physiol., 2007, vol. 293, no. 5, pp. L1240–L1249.
Camprubí-Rimblas, M., Tantinyà, N., Bringué, J., et al., Anticoagulant therapy in acute respiratory distress syndrome, Ann. Transl. Med., 2018, vol. 6, no. 2, p. 36.
Caravaca, A.S., Gallina, A.L., Tarnawski, L., et al., An effective method for acute vagus nerve stimulation in experimental inflammation, Front. Neurosci., 2019, vol. 13, p. 877.
Cargnello, M. and Roux, P.P., Activation and function of the mapks and their substrates, the MAPK-activated protein kinases, Microbiol. Mol. Biol. Rev., 2011, vol. 75, no. 1, pp. 50–83.
Celada, P., Dolera, M., Alvarez, E., and Artigas, F., Effects of acute and chronic treatment with fluvoxamine on extracellular and platelet serotonin in the blood of major depressive patients. Relationship to clinical improvement, J. Affective Disord., 1992, vol. 25, no. 4, pp. 243–249.
Chepur, S.V., Pluzhnikov, N.N., Saiganov, S.A., et al., The hypothesis of the aperiodic polysaccharides matrix synthesis, Usp. Sovrem. Biol., 2019, vol. 139, no. 6, pp. 583–593.
Chiang, N. and Serhan, C.N., Specialized pro-resolving mediator network: an update on production and actions, Essays Biochem., 2020, vol. 64, no. 3, pp. 443–462.
Chimenti, L., Camprubí-Rimblas, M., Guillamat-Prats, R., et al., Nebulized heparin attenuates pulmonary coagulopathy and inflammation through alveolar macrophages in a rat model of acute lung injury, Thromb. Haemostasis, 2017, vol. 117, no. 11, pp. 2125–2134.
Choi, G., Hofstra, J.J., Roelofs, J.J., et al., Antithrombin inhibits bronchoalveolar activation of coagulation and limits lung injury during Streptococcus pneumoniae pneumonia in rats, Crit. Care Med., 2008, vol. 36, no. 1, pp. 204–210.
Clausen, T.M., Sandoval, D.R., Spliid, C.B., et al., SARS-CoV-2 infection depends on cellular heparan sulfate and ACE2, Cell, 2020, vol. 183, no. 4, pp. 1043–1057.
Codagnone, M., Cianci, E., Lamolinara, A., et al., Resolvin D1 enhances the resolution of lung inflammation caused by long-term Pseudomonas aeruginosa infection, Mucosal Immunol., 2018, vol. 11, no. 1, pp. 35–49.
De Virgiliis, F. and Di Giovanni, S., Lung innervation in the eye of a cytokine storm: neuroimmune interactions and COVID-19, Nat. Rev. Neurol., 2020, vol. 16, no. 11, pp. 645–652.
Di Salvo, E., Di Gioacchino, M., Tonacci, A., et al., Alarmins, COVID-19 and comorbidities, Ann. Med., 2021, vol. 53, no. 1, pp. 777–785.
Emanuelli, G., Nassehzadeh-Tabriz, N., Morrell, N.W., and Marciniak, S.J., The integrated stress response in pulmonary disease, European Respiratory Review, 2020, vol. 29, no. 157, p. 200184.
Fischer, A., Sellner, M., Neranjan, S., et al., Potential inhibitors for novel coronavirus protease identified by virtual screening of 606 million compounds, Int. J. Mol. Sci., 2020, vol. 21, no. 10, p. 3626.
Fusade-Boyer, M., Dupré, G., Bessière, P., et al., Evaluation of the antiviral activity of Sephin1 treatment and its consequences on eIF2α phosphorylation in response to viral infections, Front. Immunol., 2019, vol. 10, p. 134.
Grace, M.S., Dubuis, E., Birrell, M.A., and Belvisi, M.G., Pre-clinical studies in cough research: role of transient receptor potential (TRP) channels, Pulm. Pharmacol. Ther., 2013, vol. 26, no. 5, pp. 498–507.
Hsiao, H.M., Sapinoro, R.E., Thatcher, T.H., et al., A novel anti-inflammatory and pro-resolving role for resolvin D1 in acute cigarette smoke-induced lung inflammation, PLoS One, 2013, vol. 8, no. 3, p. e58258.
Hsiao, H.M., Thatcher, T.H., Colas, R.A., et al., Resolvin D1 reduces emphysema and chronic inflammation, Am. J. Pathol., 2015, vol. 185, no. 12, pp. 3189–3201.
Hu, X., Shen, H., Wang, Y., et al., Aspirin-triggered resolvin D1 alleviates paraquat-induced acute lung injury in mice, Life Sci., 2019, vol. 218, pp. 38–46.
Imai, Y., Kuba, K., Neely, G.G., et al., Identification of oxidative stress and Toll-like receptor 4 signaling as a key pathway of acute lung injury, Cell, 2008, vol. 133, no. 2, pp. 235–249.
Islam, J., Shree, A., Vafa, A., et al., Taxifolin ameliorates benzo[a]pyrene-induced lung injury possibly via stimulating the Nrf2 signalling pathway, Int. Immunopharmacol., 2021, vol. 96, p. 107566.
Jafarinia, M., Hosseini, M.S., Kasiri, N., et al., Quercetin with the potential effect on allergic diseases, Allergy, Asthma, Clin. Immunol., 2020, vol. 16, p. 36.
Jia, Y. and Lee, L.-Y., Role of TRPV receptors in respiratory diseases, Biochim. Biophys. Acta, Mol. Basis Dis., 2007, vol. 1772, no. 8, pp. 915–927.
Kandadi, M.R., Frankel, A.E., and Ren, J., Toll-like receptor 4 knockout protects against anthrax lethal toxin-induced cardiac contractile dysfunction: role of autophagy, Br. J. Pharmacol., 2012, vol. 167, no. 3, pp. 612–626.
Koarai, A., Traves, S.L., Fenwick, P.S., et al., Expression of muscarinic receptors by human macrophages, Eur. Respir. J., 2012, vol. 39, no. 3, pp. 698–704.
Krishnamoorthy, N., Abdulnour, R.E., Walker, K.H., et al., Specialized proresolving mediators in innate and adaptive immune responses in airway diseases, Physiol. Rev., 2018, vol. 98, no. 3, pp. 1335–1370.
Kvetkina, A.N., Kaluzhskiy, L.A., Leychenko, E.V., et al., New targets of Kunitz-type peptide from sea anemone Heteractis magnifica, Dokl. Biochem. Biophys., 2019, vol. 487, pp. 260–263.
Liu, J.H., Cao, L., Zhang, C.H., et al., Dihydroquercetin attenuates lipopolysaccharide-induced acute lung injury through modulating FOXO3-mediated NF-κB signaling via miR-132-3p, Pulm. Pharmacol. Ther., 2020, vol. 64, p. 101934.
Marčec, R. and Likić, R., Could fluvoxamine keep COVID-19 patients out of hospitals and intensive care units?, Croat. Med. J., 2021, vol. 62, no. 1, pp. 95–100.
Mousavi, S., Moradi, M., Khorshidahmad, T., and Motamedi, M., Anti-inflammatory effects of heparin and its derivatives: a systematic review, Adv. Pharmacol. Sci., 2015, vol. 2015, p. 507151.
Nassenstein, C., Krasteva-Christ, G., and Renz, H., New aspects of neuroinflammation and neuroimmune crosstalk in the airways, J. Allergy Clin. Immunol., 2018, vol. 142, no. 5, pp. 1415–1422.
Noguchi, M., Furukawa, K.T., and Morimoto, M., Pulmonary neuroendocrine cells: physiology, tissue homeostasis and disease, Dis. Models Mech., 2020, vol. 13, no. 12, p. dmm046920.
Nonomura, K., Woo, S.H., Chang, R.B., et al., Piezo2 senses airway stretch and mediates lung inflation-induced apnoea, Nature, 2017, vol. 541, no. 7636, pp. 176–181.
Pakos-Zebrucka, K., Koryga, I., Mnich, K., et al., The integrated stress response, EMBO Rep., 2016, vol. 17, no. 10, pp. 1374–1395.
Panigrahy, D., Gilligan, M.M., Huang, S., et al., Inflammation resolution: a dual-pronged approach to averting cytokine storms in COVID-19?, Cancer Metastasis Rev., 2020, vol. 39, no. 2, pp. 337–340.
Perrin-Cocon, L., Aublin-Gex, A., Sestito, S.E., et al., TLR4 antagonist FP7 inhibits LPS-induced cytokine production and glycolytic reprogramming in dendritic cells, and protects mice from lethal influenza infection, Sci. Rep., 2017, vol. 7, p. 40791.
Pierre, P., Integrating stress responses and immunity, Science, 2019, vol. 365, no. 6448, pp. 28–29.
Prescott, S.L., Umans, B.D., Williams, E.K., et al., An airway protection program revealed by sweeping genetic control of vagal afferents, Cell, 2020, vol. 181, no. 3, pp. 574–589.
Rosen, D.A., Seki, S.M., Fernández-Castañeda, A., et al., Modulation of the sigma-1 receptor-IRE1 pathway is beneficial in preclinical models of inflammation and sepsis, Sci. Transl. Med., 2019, vol. 11, no. 478, p. eaau5266.
Rynko, A.E., Fryer, A.D., and Jacoby, D.B., Interleukin-1β mediates virus-induced M2 muscarinic receptor dysfunction and airway hyperreactivity, Am. J. Respir. Cell Mol. Biol., 2014, vol. 51, no. 4, pp. 494–501.
Schloer, S., Hübel, N., Masemann, D., et al., The annexin A1/FPR2 signaling axis expands alveolar macrophages, limits viral replication, and attenuates pathogenesis in the murine influenza A virus infection model, FASEB J., 2019, vol. 33, no. 11, pp. 12188–12199.
Selfridge, B.R., Wang, X., Zhang, Y., et al., Structure-activity relationships of (+)-naltrexone-inspired Toll-like receptor 4 (TLR4) antagonists, J. Med. Chem., 2015, vol. 58, no. 12, pp. 5038–5052.
Shen, X., Wang, Q., Lin, Y., et al., Voltage-dependent modulation of TRPA1 currents by diphenhydramine, Cell Calcium, 2020, vol. 90, p. 102245.
Shi, C., Tingting, W., Li, J.P., et al., Comprehensive landscape of heparin therapy for COVID-19, Carbohydr. Res., 2021, vol. 254, p. 117232.
Sukhatme, V.P., Reiersen, A.M., Vayttaden, S.J., and Sukhatme, V.V., Fluvoxamine: a review of its mechanism of action and its role in COVID-19, Front. Pharmacol., 2021, vol. 12, p. 652688.
Sun, H.M., Hong, L.Z., Shen, X.K., et al., Antithrombin-III without concomitant heparin improves endotoxin-induced acute lung injury rats by inhibiting the activation of mitogen-activated protein kinase, Chin. Med. J. (Engl.), 2009, vol. 122, no. 20, pp. 2466–2471.
Van Haren, F.M.P., Page, C., Laffey, J.G., et al., Nebulised heparin as a treatment for COVID-19: scientific rationale and a call for randomized evidence, Crit. Care, 2020, vol. 24, no. 1, p. 454.
Wang, L., Yuan, R., Yao, C., et al., Effects of resolvin D1 on inflammatory responses and oxidative stress of lipopolysaccharide-induced acute lung injury in mice, Chin. Med. J. (Engl.), 2014, vol. 127, no. 5, pp. 803–809.
Wang, X., Zhang, Y., Peng, Y., et al., Pharmacological characterization of the opioid inactive isomers (+)-naltrexone and (+)-naloxone as antagonists of toll-like receptor 4, Br. J. Pharmacol., 2016, vol. 173, no. 5, pp. 856–869.
Wang, H., Anthony, D., Yatmaz, S., et al., Aspirin-triggered resolvin D1 reduces pneumococcal lung infection and inflammation in a viral and bacterial coinfection pneumonia model, Clin. Sci. (Lond.), 2017, vol. 131, no. 18, pp. 2347–2362.
Watanabe, S., Markov, N.S., Lu, Z., et al., Resetting proteostasis with ISRIB promotes epithelial differentiation to attenuate pulmonary fibrosis, Proc. Natl. Acad. Sci. USA, 2021, vol. 118, no. 20, p. e2101100118.
Watkins, L.R., Wang, X., Mustafa, S., and Hutchinson, M.R., In vivo veritas: (+)-naltrexone’s actions define translational importance: a letter in response to Skolnick et al., “Translational” potential of naloxone and naltrexone as tlr4 “antagonists,” Trends Pharmacol. Sci., 2014, vol. 35, no. 9, pp. 432–433.
Van’t Wout, E.F., Hiemstra, P.S., and Marciniak, S.J., The integrated stress response in lung disease, Am. J. Respir. Cell Mol. Biol., 2014, vol. 50, no. 6, pp. 1005–1009.
Xia, H., Wang, J., Sun, S., et al., Resolvin D1 alleviates ventilator-induced lung injury in mice by activating PPARγ/NF-κB signaling pathway, BioMed. Res. Int., 2019, vol. 2019, p. 6254587.
Xu, M., Zhang, Y., Wang, M., et al., TRPV1 and TRPA1 in lung inflammation and airway hyperresponsiveness induced by fine particulate matter (PM2.5), Oxid. Med. Cell. Longevity, 2019, vol. 2019, p. 7450151.
Yao, H., De Boer, W.I., and Rahman, I., Targeting lung inflammation: novel therapies for the treatment of copd, Curr. Respir. Med. Rev., 2008, vol. 4, no. 1, pp. 57–68.
Yeger, H., Pan, J., and Cutz, E., Precursors and stem cells of the pulmonary neuroendocrine cell system in the developing mammalian lung, in Airway Chemoreceptors in the Vertebrates. Structure, Evolution and Function, Zaccone, G., Cutz, E., Adriaensen, D., Nurse, C.A., and Mauceri, A., Eds., Boca Raton, FL: CRC Press, 2019, pp. 291–310.
Yildiz-Pekoz, A. and Ozsoy, Y., Inhaled heparin: therapeutic efficacy and recent formulations, J. Aerosol Med. Pulm. Drug Delivery, 2017, vol. 30, no. 3, pp. 143–156.
Zhuo, Y., Zhang, S., Li, C., et al., Resolvin D1 promotes SIRT1 expression to counteract the activation of STAT3 and NF-κB in mice with septic-associated lung injury, Inflammation, 2018, vol. 41, no. 5, pp. 1762–1771.
Zingoni, A., Molfetta, R., Fionda, C., et al., NKG2D and its ligands: “One for all, all for one,” Front. Immunol., 2018, vol. 9, p. 476.
Zyryanova, A.F., Kashiwagi, K., Rato, C., et al., ISRIB blunts the integrated stress response by allosterically antagonising the inhibitory effect of phosphorylated eIF2 on eIF2B, Mol. Cell, 2021, vol. 81, no. 1, pp. 88–103.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest. The authors declare that they have no conflicts of interest.
In carrying out this work, all ethical standards were observed.
This article does not contain any studies involving humans and animals as objects.
Additional information
Translated by A. Ostyak
Rights and permissions
About this article

Cite this article
Yakovlev, O.A., Yudin, M.A., Chepur, S.V. et al. Non-Specific Targets for Correction of Pneumonia Caused by Aerosols Containing Damaging Factors of Various Nature. Biol Bull Rev 12, 649–660 (2022). https://doi.org/10.1134/S207908642206010X
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S207908642206010X